
Abstract
Reduced leptin (Ob protein) signaling is proposed to be a stimulus for the activation of neuropeptide Y (NPY) gene activity and increased expression of mRNA for the long form of the leptin receptor (Ob-Rb) in the hypothalamic arcuate nucleus. To determine if Ob-Rb protein is expressed in arcuate nucleus NPY neurons, we developed an affinity-purified polyclonal antibody against amino acids 956-1102 of human Ob-Rb. This antibody specifically recognizes the cytoplasmic tail of Ob-Rb and does not react with shorter leptin-receptor variants. Western immunoblots of Ob-Rb-transfected COS cells showed a single 150-kD band, and immunofluorescence revealed intense perinuclear staining in the cytoplasm. A 150-kD band was also present in Western immunoblots of hypothalamus. Immunocytochemical staining of brain slices revealed immunoreactive Ob-Rb protein concentrated in many neuronal cell bodies in the same regions of the forebrain that also express Ob-Rb mRNA. In the hypothalamus, Ob-Rb-positive cell bodies were abundant in the arcuate nucleus and ventromedial nucleus, with lesser numbers in the dorsomedial nucleus and paraventricular nucleus. Immunostaining was also detected in cell bodies of pyramidal cell neurons of the pyriform cortex and cerebral cortex, in neurons of the thalamus, and on the surface of ependymal cells lining the third ventricle. The choroid plexus, which expresses the short Ob-Ra form, was negative. Combined immunocytochemistry for Ob-Rb protein and fluorescence in situ hybridization for NPY mRNA identified arcuate nucleus neurons containing both NPY mRNA and Ob-Rb protein. The present finding of Ob-Rb protein in neurons that express NPY mRNA supports the hypothesis that arcuate nucleus NPY neurons are direct targets of leptin and play an important role in regulation of food intake and body weight.
A
Much attention is currently focused on the effects of leptin on the expression of arcuate nucleus neuropeptide Y (NPY), a potent hypothalamic stimulator of food intake. We have hypothesized that reduced leptin signaling in the hypothalamus is a stimulus for the activation of NPY gene activity in arcuate nucleus neurons (Schwartz et al. 1996b; Baskin et al. 1998). This is consistent with evidence that NPY mRNA levels in the arcuate nucleus are elevated in response to low leptin levels, as occurs in the leptin-deficient ob/ob mouse (Stephens et al. 1995; Rohner-Jeanrenaud et al. 1996; Schwartz et al. 1996b) and fasted rodents (Schwartz et al. 1996b,1998), whereas elevated leptin levels during ad libitum feeding inhibit arcuate nucleus NPY gene activity and food intake, as shown by administration of leptin to leptin-deficient or fasted rodents (Campfield et al. 1995; Halaas et al. 1995; Pelleymounter et al. 1995; Weigle et al. 1995; Cusin et al. 1996; Schwartz et al. 1996a; Seeley et al. 1996) and by direct injections of leptin into the arcuate nucleus and adjacent hypothalamic regions (Jacob et al. 1997; Satoh et al. 1997).
A role for leptin in regulating the activity of arcuate nucleus NPY gene expression is supported by evidence that mRNA encoding the long form of the leptin receptor (Ob-Rb) (Tartaglia 1997) is expressed in the arcuate nucleus. Ob-Rb is considered to be the major signaling form of the leptin receptor in the brain in contrast to Ob-Ra, a shorter splice variant with a truncated cytoplasmic tail and diminished signaling capabilities (Bjorbaek et al. 1997; Tartaglia 1997; White et al. 1997). Ob-Ra, but not Ob-Rb, is expressed at high levels in the choroid plexus and is believed to be responsible for the high levels of choroid plexus leptin binding (Banks et al. 1996; Devos et al. 1996; Corp et al. 1996). Whereas both Ob-Ra and Ob-Rb splice-variant transcripts are present in the hypothalamus, Ob-Rb mRNA is more abundant than Ob-Ra mRNA in the arcuate nucleus (Huang et al. 1996; Mercer et al. 1996a,b; Fei et al. 1997; Guan et al. 1997; Baskin et al. 1998).
Finding Ob-Rb receptor protein in arcuate nucleus NPY neurons would strengthen the hypothesis that the mechanism of leptin's effects on energy balance involves NPY neurons in the hypothalamus. Ob-Rb mRNA has been detected in arcuate nucleus NPY neurons by in situ hybridization (Mercer et al. 1996a,b). Leptin receptor-like immunoreactive proteins have been associated with arcuate nucleus NPY cell bodies by immunocytochemistry, but the antibodies that were used in previous studies do not distinguish between Ob-Ra and Ob-Rb forms of the receptor protein (Håkansson et al. 1996; Shioda et al. 1998; Yarnell et al. 1998). Therefore, whether the leptin receptor protein in the arcuate nucleus includes Ob-Rb, the major signaling form of the receptor in brain neural tissue, and whether the Ob-Rb receptor protein is expressed in arcuate nucleus NPY neurons have remained important unsolved problems.
To determine if the Ob-Rb receptor protein is expressed in arcuate nucleus neurons (NPY neurons in particular) we developed an affinity-purified antibody that specifically recognizes the cytoplasmic tail of Ob-Rb and does not recognize amino acid sequences found in shorter leptin receptor splice variants. The specificity of this Ob-Rb IgG was validated by analysis of transfected COS cells that express Ob-Rb and ObRa. We used this Ob-Rb-specific IgG for immunocytochemical staining of rat brain, and combined Ob-Rb immunostaining with in situ hybridization for NPY mRNA.
Materials and Methods
Reagents
Chemicals were obtained from Sigma (St Louis, MO), unless otherwise noted. Labeled antisera, second antibodies, and normal sera were obtained from Jackson ImmunoResearch (West Grove, PA), except where indicated.
Animals
Male Wistar rats (280-300 g) (Simonsen Labs; Gilroy, CA) were housed individually and maintained on a 12:12-hr day: night cycle (0700-0900 hr) with ad lib access to standard rodent chow and water before the study. For collection of brains, animals were sacrificed in the morning by decapitation under CO2 inhalation. The Animal Research Committees of the Veterans Affairs Puget Sound Health Care System and the University of Washington approved all procedures.
Antiserum
To prepare an antibody that is specific for Ob-Rb, a cDNA encoding amino acids 956-1102 of human Ob-Rb was fused to glutathione S-transferase (GST) (pGEX4T-1; Pharmacia, Uppsala, Sweden) and expressed in E. coli MC1061. The GST/Ob-Rb fusion protein was injected into rabbits (Pocono Animal Farms; Pocono, PA) to make Ob-Rb antiserum. The IgG was purified from the antiserum (Osborne et al. 1995), after which affinity-purified anti-Ob-Rb was prepared by passage over immobilized GST/Ob-Rb.
COS Cell Studies
To verify the specificity of the anti-Ob-Rb IgG for Ob-Rb protein, extracts of COS cells transfected with cDNAs encoding Ob-Rb or Ob-Ra (gift of L. Tartaglia) and grown for 48 hr (Osborne et al. 1995) were incubated for 1 hr with anti-Ob-Rb IgG (1 μg/ml) or leptin-Nugel beads (gift of P. Bailon), followed by protein G-Sepharose for 1 hr and SDS-PAGE (4-20% gel). After transfer to nitrocellulose membranes, the blots were incubated in 1 μg/ml anti-Ob-Rb IgG and visualized by chemiluminescence (Trophix; South Plain-field, NJ). Western immunoblotting controls for specificity of the anti-Ob-Rb IgG in detecting Ob-Rb in transfected COS cells included (a) the use of mock-transfected COS cells and (b) substitution of affinity-purified anti-GST IgG (0.5 μg/ml) directed against the GST portion of the GST/Ob-Rb fusion protein in place of the affinity-purified anti-Ob-Rb IgG. Specificity of the affinity-purified Ob-Rb antiserum for immunostaining was demonstrated with COS cells transfected with Ob-Rb or Ob-Ra. The COS cells were fixed in 4% paraformaldehyde, permeabilized with 1% Triton X-100 (Hakimi et al. 1990), and immunostained with affinity-purified anti-Ob-Rb IgG (1 μg/ml), followed by donkey anti-rabbit IgG-Cy3. Immunostaining controls included (a) substitution of anti-GST IgG for the affinity-purified Ob-Rb antiserum, (b) immunostaining of COS cells transfected with Ob-Ra or mock-transfected COS cells, and (c) immunostaining of unpermeabilized cells.
Western Immunoblotting of Tissue
To characterize the reactivity of the anti-Ob-Rb IgG with rat brain proteins, hypothalamus and liver were homogenized at 4C in a lysis buffer (50 mM Hepes, pH 7.5, with 137 mM NaCl, 10 mM Na3VO4, 100 mM NaF, 100 mM tetrasodium pyrophosphate, 10 mM EDTA, 0.1% Triton X-100, 2 mM PMSF, and 100 μg/ml leupeptin and aprotinin, and centrifuged at 15,000 X g. Supernatants (with equivalent protein concentrations) were immunoprecipitated overnight at 4C with anti-Ob-Rb IgG (0.5 μg/ml) and mixed with Gammabind G-Sepharose (Pharmacia) for 90 min at 4C, followed by SDS-PAGE in a 7.5% gel. Proteins were transferred to PVDF membranes and blocked overnight in 5% dried milk, then immersed for 3 hr in anti-Ob-Rb IgG (0.5 μg/ml) followed by goat anti-rabbit IgG-alkaline phosphatase and detected by chemiluminescence (Trophix). Controls for the specificity of anti-Ob-Rb in Western immunoblots of brain and liver included absorption of the anti-Ob-Rb IgG with the GST/Ob-Rb fusion protein and substitution of anti-GST IgG for the affinity-purified Ob-Rb IgG.
Immunostaining of Ob-Rb in Brain
Brains (n = 12) were fixed by vascular perfusion in 4% paraformaldehyde, sectioned coronally by a cryostat at 10 μm, thaw-mounted on slides, and stored at −80C until use. Sections for immunocytochemical staining were selected from the region between stereotaxic levels −2.5 and −3.5 mm posterior to the bregma (Paxinos and Watson 1986). Slides were thawed to room temperature (RT) and placed in 0.05 M phosphate buffer containing 0.9% NaCl (PBS) at 4C. They were then immersed in a blocking buffer containing PBS with 5% normal donkey serum for 40 min and rinsed in PBS. Sections were incubated in anti-Ob-Rb IgG (0.2 μg/ml) overnight at 4C. For detection of Ob-Rb-like protein by brightfield microscopy, the ABC-peroxidase method was used (Baskin et al. 1993,1994). For fluorescence detection of Ob-Rb immunoreactive sites, incubation in anti-Ob-Rb IgG was followed by a rinse in cold PBS and then immersion in donkey anti-rabbit IgG-Cy3 (1:200) at RT for 1 hr. Sections observed by fluorescence were covered with a glycerol-based mounting medium in glycine buffer (pH 9.0) containing poly-vinyl alcohol to retard fading. Immunocytochemical staining controls included (a) substitution of affinity-purified IgG to the GST portion of the fusion protein (0.2 μg/ml) in place of anti-Ob-Rb IgG, (b) absorption of the anti-Ob-Rb IgG with the full GST/Ob-Rb fusion protein (10 μg/ml), and (c) omission of anti-Ob-Rb IgG from the immunocytochemical staining protocol.
Combined Immunocytochemistry and In Situ Hybridization
Sections of paraformaldehyde-perfused rat brains were obtained from the specimens prepared for immunocytochemical staining of Ob-Rb protein, as described above, and placed in freshly prepared 0.1 M TEA, pH 8.0. The sections were acetylated for 10 min in 350 ml TEA containing 875 μl acetic anhydride and were rinsed in 2 × SSC. Sections were dehydrated in ethanol and delipidated with chloroform, followed by 100% and 95% ethanol (2 min each) and PBS at 4C. Sections were immunostained with affinity-purified anti-Ob-Rb IgG and donkey anti-rabbit IgG-Cy3 (with the addition of 0.25% Tween in the buffer), as described above, followed by three 5-min rinses in PBS without Tween. The sections were then subjected to the fluorescence in situ hybridization (FISH) protocol to detect NPY mRNA (Baskin et al. 1998; Hahn et al. 1998), including the appropriate controls for specificity of the immunostaining and in situ hybridization (see above). Cell nuclei were visualized with Hoechst 33258 dye (Molecular Probes; Eugene, OR).
Results
Characterization of the Anti-Ob-Rb IgG in COS Cells
Immunoblots of Ob-Rb-transfected COS cells showed a single broad band of approximately 150 kD when incubated in anti-Ob-Rb IgG. This band was also present when Ob-Rb protein was concentrated from Ob-Rb-transfected COS cell extracts by immunoprecipitation with anti-Ob-Rb IgG or with leptin immobilized on Nugel beads. The 150-kD band was absent in mock-transfected COS cells and in COS cells transfected with ObRa (Figure 1A). Immunofluorescence with anti-Ob-Rb IgG revealed intense perinuclear staining in Ob-Rb-transfected COS cells (Figure 1B). In controls, immunofluorescent staining was absent when anti-GST IgG was substituted for anti-Ob-Rb IgG and when Ob-Ra-transfected COS cells or mock-transfected cells were used (Figure 1C). Ob-Rb immunostaining was also observed in COS cells transfected with mouse Ob-Rb cDNA (not shown).
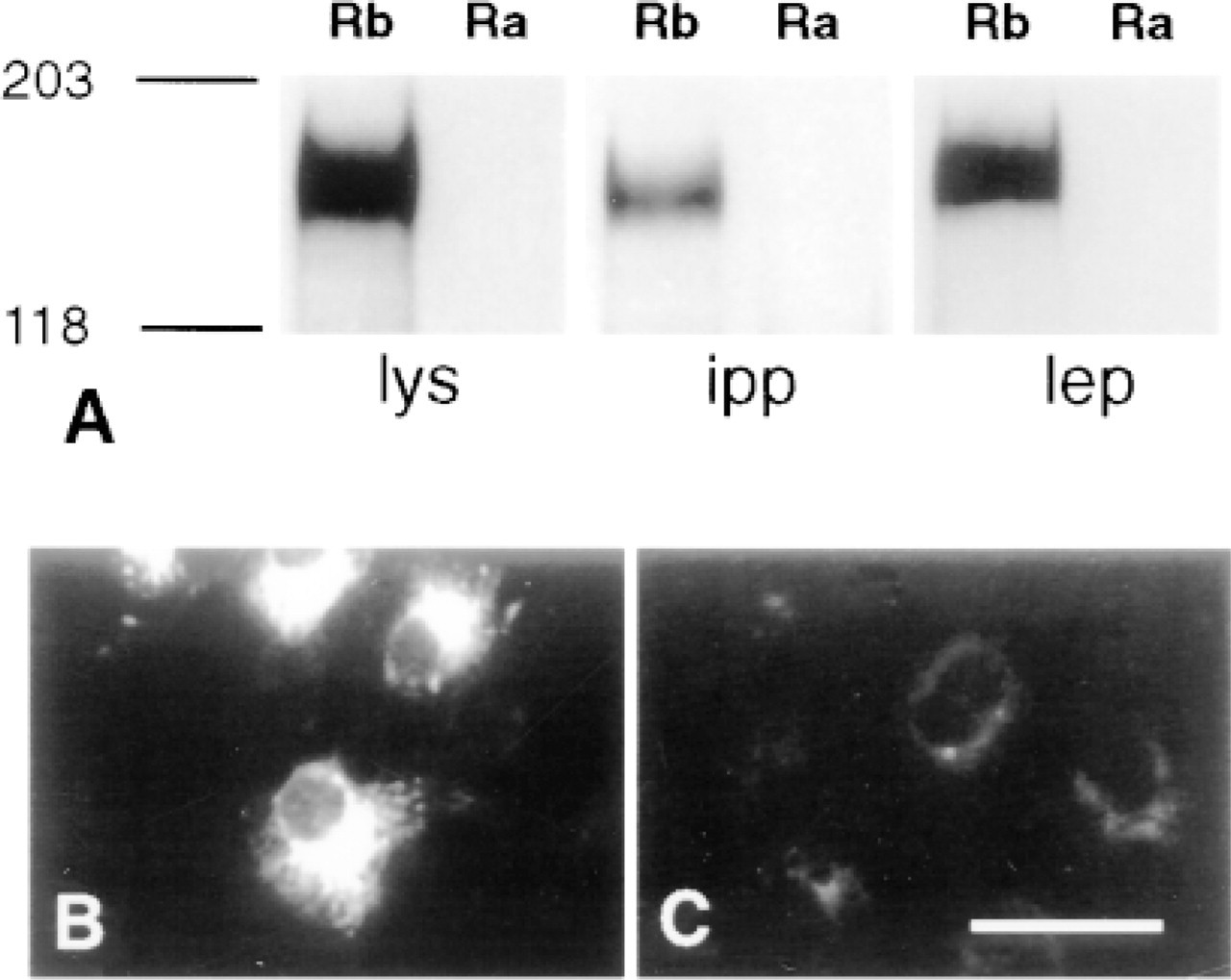
Detection of Ob-Rb with anti-Ob-Rb IgG in COS cells transfected with Ob-Rb (Rb) but not in COS cells transfected with Ob-Ra (Ra). (A) Immunoblots show a single 150-kD band in Ob-Rb transfected cell lysate (lys), in extracts immunoprecipitated with anti-Ob-Rb IgG (ipp), and in extracts mixed with leptin-Nugel beads (lep). (B) Anti-Ob-Rb IgG detects cytoplasmic Ob-Rb-like protein in Ob-Rb COS cells but not in Ob-Ra COS cells (C). All controls resembled C. Bar = 25 μm.
Immunocytochemical Staining of Ob-Rb in Brain
Brain sections that were incubated in with anti-Ob-Rb IgG showed many positively stained neuronal cell bodies throughout the arcuate nucleus (Figures 2A and 2D). A high percentage of arcuate nucleus neuronal cell bodies contained Ob-Rb-like immunoreactive protein, although the precise numbers were not determined quantitatively. Staining was absent when anti-GST IgG was substituted for the anti-Ob-Rb IgG (Figure 2B) and when the anti-Ob-Rb IgG was absorbed with the GST/Ob-Rb fusion protein (Figure 2E). In individual neuronal cell bodies the immunoreactive Ob-Rb-like protein was concentrated intracellularly in a perinuclear distribution (Figures 2C and 2F). No neuronal cell bodies showed a staining pattern suggestive of surface labeling, although a surface location of the immunoreactivity could not be precluded. This staining pattern was consistently found in all rats and was detected by both peroxidase and fluorescence immunocytochemical staining methods in all regions of the forebrain that were examined.
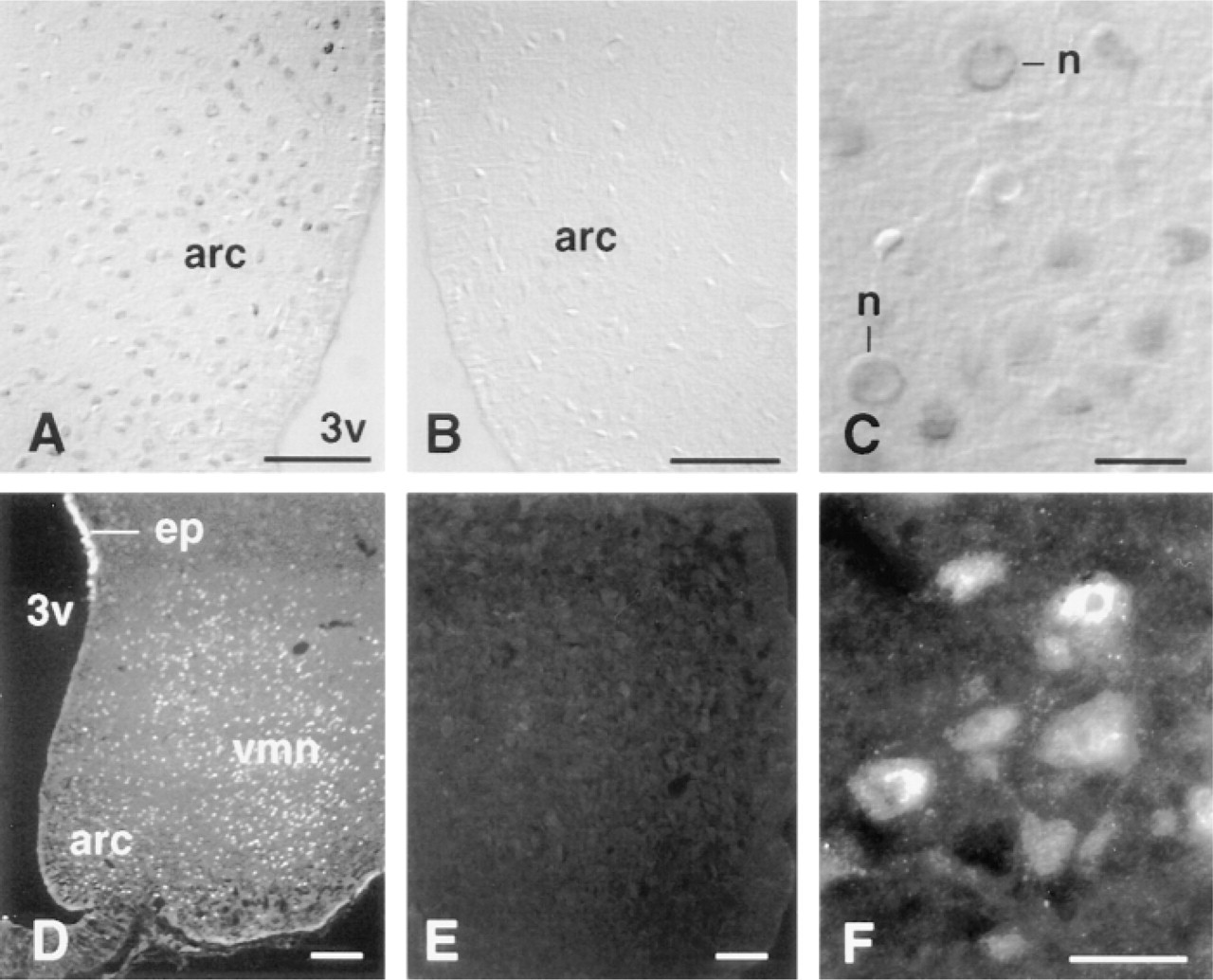
Immunocytochemical staining with anti-Ob-Rb IgG in the arcuate nucleus, using immunoperoxidase (A-C) and immunofluorescence (D-F) methods. (A,D) Detection of Ob-Rb-positive neuron cell bodies in the arcuate nucleus (arc) and ventromedial nucleus (ventromedial nucleus) adjacent to third ventricle (3v). The surface of the ependymal lining of the ventricle also has Ob-Rb-like staining (ep). Control sections immunostained with anti-GST control serum (B) and anti-Ob-Rb IgG absorbed with the Ob-Rb/GST fusion protein (E) are negative. (C,F) Higher magnification of arcuate nucleus, showing Ob-Rb-like protein concentrated in neuronal cell bodies (n). Bars: A,B,D,E = 100 μm; C,F = 50 μm.
Positive staining of Ob-Rb-like immunoreactive protein was also observed in other hypothalamic regions. In the paraventricular nucleus, Ob-Rb-like immunoreactive protein was detected as small foci of bright fluorescence that appeared to be located in parvocellular neuronal cell bodies (Figure 3A). The intensity and abundance of Ob-Rb-like immunoreactive protein were relatively weak in the paraventricular nucleus compared to the arcuate nucleus. In contrast, the ventromedial nucleus showed many positively stained neuronal cell bodies with intense intracellular immunoreactivity to the anti-Ob-Rb IgG (Figure 3C). Positive immunostaining was seen in neuronal cell bodies in the supraoptic nucleus and dorsomedial nucleus (not shown). The surface of the ependymal cells lining the third ventricle in the region adjacent to the ventromedial nucleus was intensely stained with the anti-Ob-Rb IgG (Figure 2D). The ependymal immunoreactivity was absent in the region of the median eminence and arcuate nucleus. Throughout the hypothalamus, staining was at background levels in cells and neuropil when anti-GST IgG was substituted for the anti-Ob-Rb IgG and when the anti-Ob-Rb IgG was absorbed with the GST/Ob-Rb fusion protein (Figures 3B and 3D).
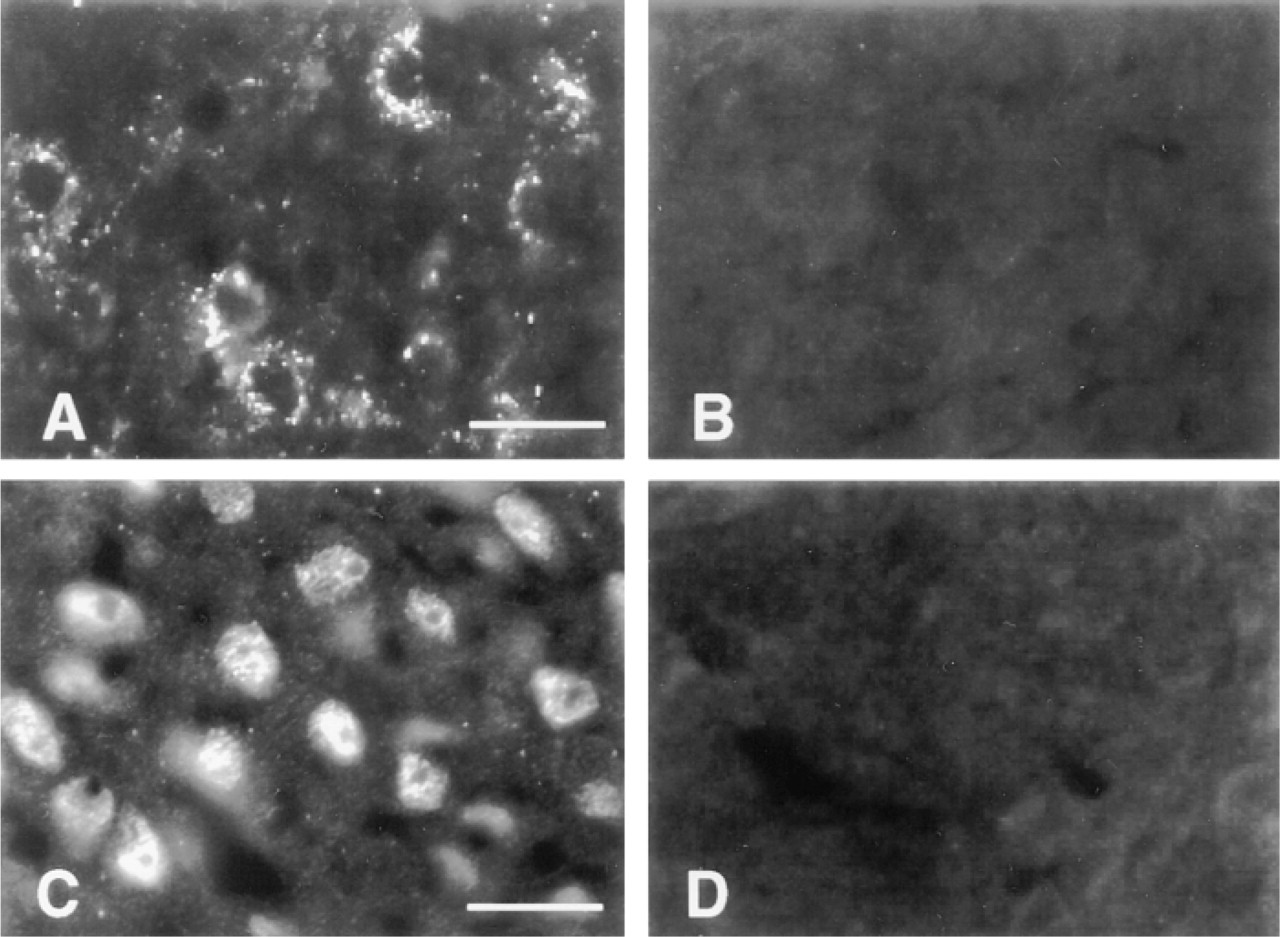
Immunocytochemical staining of paraventricular nucleus parvocellular neuronal cell bodies (A) and cell bodies in the ventromedial nucleus (C) is positive with anti-Ob-Rb IgG but negative with anti-GST antiserum (B) or anti-Ob-Rb absorbed with the Ob-Rb/GST fusion protein (D). Bars = 50 μm.
Neurons in other forebrain regions also showed positive immunostaining for the Ob-Rb splice-variant protein. Immunostaining was markedly strong in cell bodies of pyramidal cell neurons in Layer III of the pyriform cortex (Figure 4A), in pyramidal cell bodies scattered throughout the cerebral cortex (Figure 4B), and throughout the thalamus, particularly in the anteromedial thalamic nucleus (Figure 4C). As in the hypothalamus, Ob-Rb-like immunoreactive protein was concentrated in the cytoplasm of the cell bodies. Cells showing surface labeling only were not detected. Staining of dendrites and terminals was not detected. The choroid plexus was negative for Ob-Rb-like immunoreactivity, whereas the ependymal lining of the ventricles showed bright positive immunocytochemical staining (Figure 4D). Immunostaining in nonhypothalamic regions was absent in all controls.
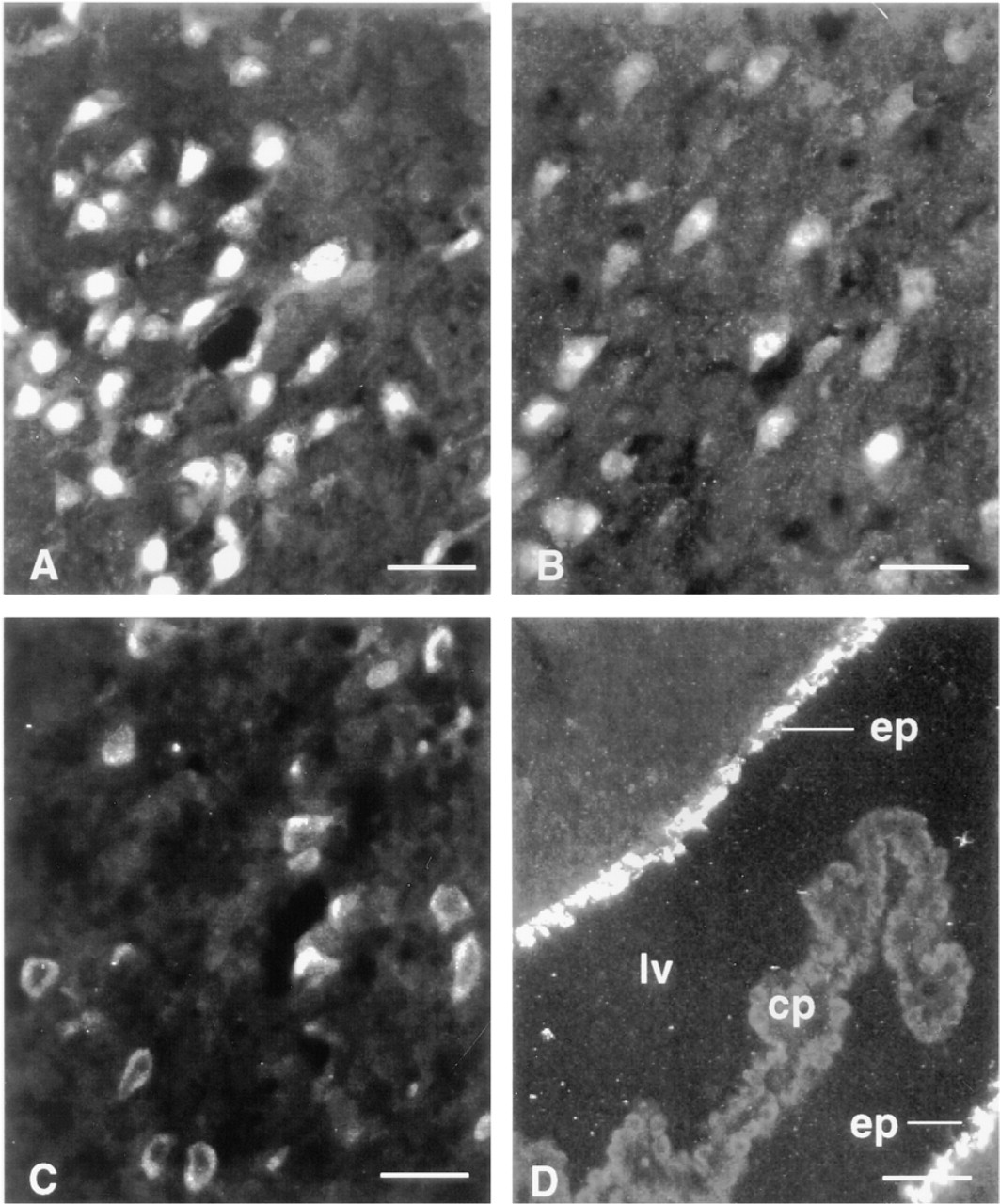
Immunocytochemical staining of pyramidal cell bodies with anti-Ob-Rb IgG in the pyriform cortex (A) and cerebral cortex (B), and cell bodies in the thalamus (C). The surfaces of ependymal cells (ep) lining the lateral ventricle (lv) are also positive but the choroid plexus (cp) is negative. Bars = 50 μm.
Co-localization of Ob-Rb Protein and NPY mRNA
To determine if Ob-Rb splice-variant protein is expressed by NPY neurons in the arcuate nucleus, we performed immunocytochemistry for Ob-Rb and FISH for NPY mRNAs in the same brain sections (Figure 5). Although the results were not quantified, visual examination suggested that about 75% of arcuate nucleus neuronal cell bodies that showed FISH for NPY mRNA also showed positive immunocytochemical staining for Ob-Rb-like immunoreactive proteins. Ob-Rb immunostaining was present in many arcuate nucleus neurons that did not express NPY mRNAs. Likewise, some neurons with NPY mRNA did not show immunocytochemical staining for Ob-Rb. Neurons with Ob-Rb-like immunoreactive protein were found in all regions of the arcuate nucleus.
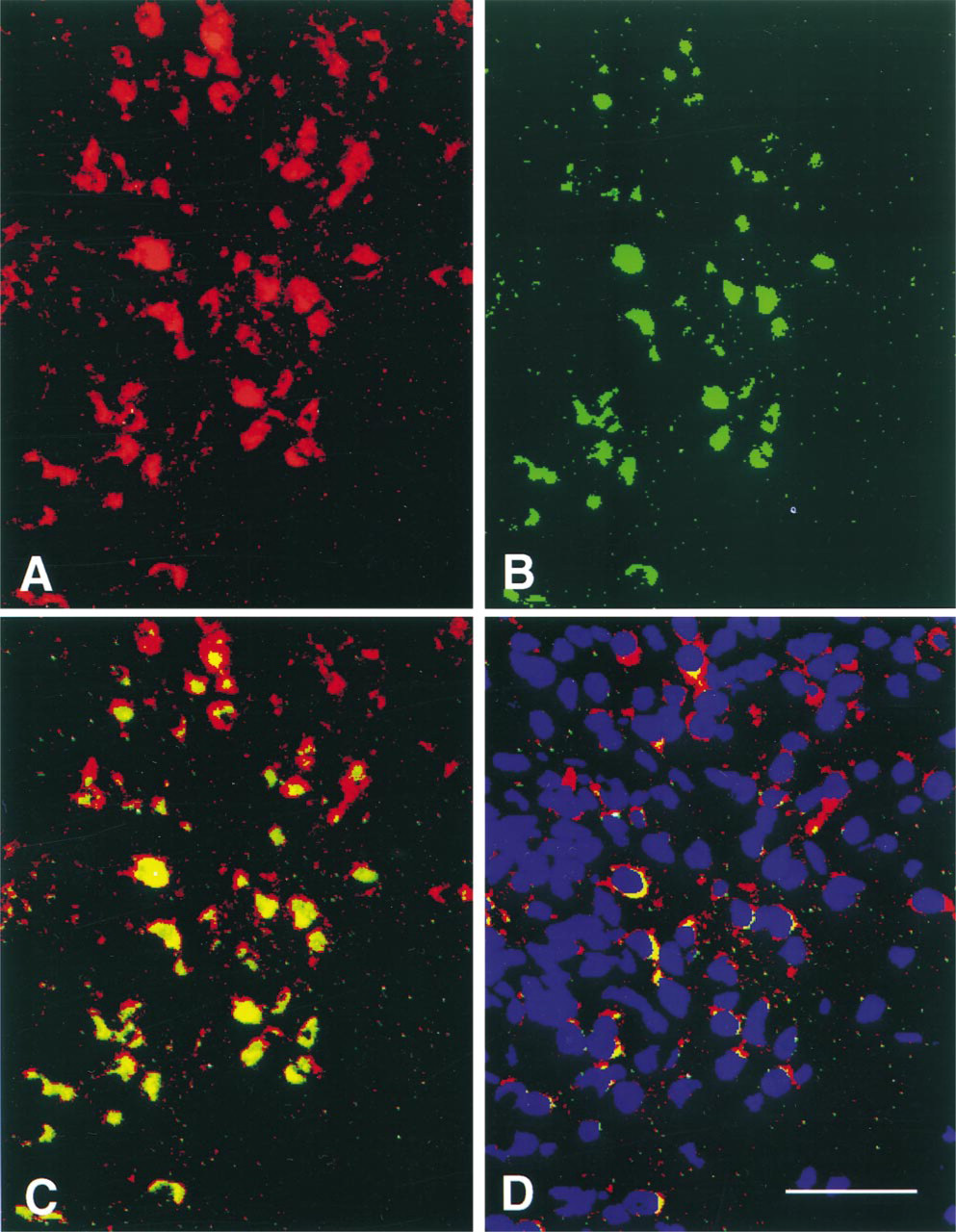
Pseudocolor image showing co-localization of NPY mRNA by FISH (red) (A) and Ob-Rb by immunostaining (green) (B) in cell bodies of the arcuate nucleus. Cells that contain both NPY mRNA and Ob-Rb protein are yellow when A and B are fused by the MCID image analysis system (C). Blue fluorescent staining with the Hoechst dye shows the location of cell nuclei (D). Bar = 100 μm.
Western Immunoblotting of Hypothalamus and Liver
Western immunoblots of hypothalamic extracts that were immunoprecipitated with anti-Ob-Rb IgG showed a prominent band representing a protein of about 150 kD, similar to the single band observed in COS cells transfected with Ob-Rb (Figure 6A). Additional bands representing smaller proteins (approximately 78, 100, and 120 kD) were also present in hypothalamic extracts. The 100-kD protein was sometimes present as a doublet. All of these bands were present in immunoprecipitates of boiled hypothalamus and in extracts prepared from both fresh and frozen hypothalamus. A variety of protease inhibitors and various buffer conditions were used in an attempt to eliminate the smaller molecular weight bands, but they were persistently present. However, all of these bands were absent when the blots were incubated in anti-Ob-Rb IgG that was absorbed with the Ob-Rb/GST fusion protein and when affinity-purified anti-GST IgG was substituted for the anti-Ob-Rb IgG. (Figure 6B). A smaller nonspecific band of about 66 kD was also present in controls. However, it probably did not contribute to the immunostaining because all immunostaining was abolished by these controls. As an additional control, Western immunoblots were performed on rat liver, which shows low expression of Ob-Rb. In immunoblots prepared from rat liver immunoprecipitated with anti-Ob-Rb IgG, the 150-kD protein band was very weak compared to hypothalamus (Figures 6C vs 6D).
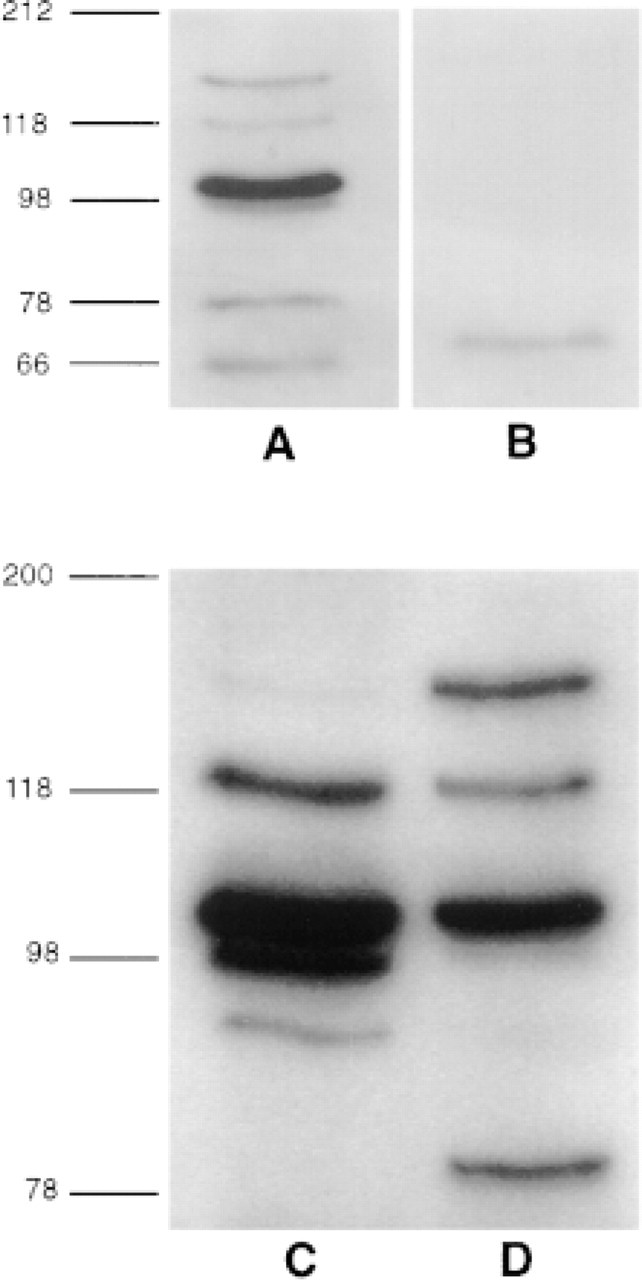
Western immunoblots of hypothalamus (A,B) and liver (C,D). Hypothalamic extracts that were immunoprecipitated with anti-Ob-Rb IgG and immunoblotted with anti-Ob-Rb IgG (A) or anti-GST IgG (B). Extracts that were immunoprecipitated with anti-GST IgG or in which anti-Ob-Rb IgG was absorbed with the Ob-Rb/ GST fusion protein resembled B. Liver (C) and hypothalamic (D) extracts immunoprecipitated and blotted with anti-Ob-Rb IgG. The liver has a weak 150-kD band, which is strong in the hypothalamus, representing Ob-Rb.
Discussion
The absence of direct evidence that Ob-Rb protein is present in the hypothalamus and expressed specifically by NPY neurons has been a significant gap in the data needed to validate the hypothesis that leptin acts in the arcuate nucleus to regulate the activity of NPY neurons. There is no doubt, on the basis of in situ hybridization analyses, that mRNA encoding the Ob-Rb form of the leptin receptor is abundant in the arcuate nucleus (Huang et al. 1996; Mercer et al. 1996a,b; Fei et al. 1997; Baskin et al. 1998). Until now, however, it has not been known whether these transcripts are translated into Ob-Rb receptor protein and whether the Ob-Rb protein is associated with NPY neurons in the arcuate nucleus. The results of the present study provide strong evidence that arcuate nucleus neurons, the NPY neurons in particular, are direct targets of leptin.
Previous immunocytochemical studies of Ob-Rb receptor protein in the arcuate nucleus used antisera that recognized amino acid sequences common to both Ob-Ra and Ob-Rb (Håkansson et al. 1996; Couce et al. 1997; Shioda et al. 1998). Therefore, immunostaining with these antisera potentially identified all known leptin-receptor splice-variant proteins in the brain. In contrast, the present investigation used an antibody directed against an amino acid sequence that is unique to the Ob-Rb splice-variant protein and does not recognize the Ob-Ra form. The results therefore constitute strong evidence that the Ob-Rb splice-variant protein is present in arcuate nucleus neurons, consistent with the expression of Ob-Rb mRNA in the arcuate nucleus (Huang et al. 1996; Mercer et al. 1996a,b; Fei et al. 1997; Guan et al. 1997; Baskin et al. 1998). Furthermore, our results indicate that the Ob-Rb splice-variant protein is expressed in many arcuate nucleus NPY neurons, which are proposed to be direct targets of leptin (Schwartz et al. 1996a; Baskin et al. 1998). This observation is consistent with earlier evidence for Ob-Rb mRNA in arcuate nucleus NPY neurons (Mercer et al. 1996b).
The present findings suggest that the relative levels of Ob-Rb receptor protein are roughly proportional to the levels of mRNA as shown by in situ hybridization in the hypothalamus and elsewhere in the forebrain. Although precise quantitation was not performed in the present analysis, Ob-Rb immunocytochemical staining was observed to be most abundant, both in terms of relative numbers of stained cells and in the intensity of the staining within cells, in the arcuate nucleus, where Ob-Rb mRNA levels are also highest (Baskin et al. 1998). Strong Ob-Rb immunocytochemical staining of neurons was also observed in the ventromedial nucleus, dorsomedial nucleus, thalamus, and pyriform cortex, which also have dense concentrations of leptin receptor mRNA in the rat brain (Schwartz et al. 1996a; Fei et al. 1997). Furthermore, the finding of Ob-Rb-like protein associated with the ependymal lining of the ventricles raises the possibility that the functions of these cells may be influenced by leptin.
Our finding of Ob-Rb protein by immunocytochemical staining in the parvocellular neuron cell bodies of the paraventricular nucleus is significant because leptin potently activates c-fos gene expression in this nucleus (Van Dijk et al. 1996; Elmquist et al. 1997). Because parvocellular paraventricular nucleus neurons project to hindbrain sympathetic and para-sympathetic preganglionic neurons, the finding of Ob-Rb protein in these paraventricular nucleus neurons is consistent with a direct action of leptin on paraventricular nucleus neurons involved in food intake and body weight regulation. Therefore, although Ob-Rb mRNA and protein expression is low in the paraventricular nucleus, neurons in this nucleus (particularly the parvocellular subdivision) appear to be direct targets of leptin action. The identity and connections of the paraventricular nucleus neurons that express Ob-Rb remain to be determined.
In neuronal cell bodies of the hypothalamus and elsewhere in the forebrain, most of the Ob-Rb-like immunoreactive protein was found in the cytoplasm. The distribution of immunoreactive protein was largely peri-nuclear, in a pattern consistent with localization in an intracellular vesicular compartment and in the Golgi apparatus. Although these neurons have an abundance of Ob-Rb in the cytoplasm, the degree to which Ob-Rb is expressed at the surface, where the receptor can interact with leptin, is unknown and cannot be determined from these results. The functional meaning of this distribution requires further investigation, but a similar intracellular distribution of immunoreactive Ob-Rb protein has been reported for endothelial cells (Sierra-Honigmann et al. 1998). Furthermore, the finding of most of the immunoreactive Ob-Rb receptor protein in a perinuclear region that corresponds to the Golgi apparatus has precedent in the intracellular location of δ-opioid receptors in dorsal root ganglion neurons (Zhang et al. 1998). Clearly, defining the dynamics of Ob-Rb trafficking in brain neurons is a critical step towards understanding the regulation of leptin sensitivity in the hypothalamus and the role of leptin in food intake and body weight.
The observations from the immunocytochemical staining and Western immunoblotting analyses of transfected COS cells support the conclusion that the anti-Ob-Rb IgG recognizes Ob-Rb. The antibody detected a single band in the Western immunoblots from COS cells that express Ob-Rb but not from COS cells that express Ob-Ra. Furthermore, only the Ob-Rb-transfected COS cells showed positive immunocytochemical staining with the anti-Ob-Rb IgG. The mobility of the Ob-Rb band in the immunoblots of Ob-Rb-COS cells suggested a molecular size of about 150 kD, which is larger than the predicted size (approximately 130 kD) based on the amino acid composition of Ob-Rb but is consistent with previous reports (Bjorbaek et al. 1997; Ghilardi and Skoda 1997). This suggests that the Ob-Rb protein may be glycosylated, as predicted by the primary sequence (Tartaglia 1997). A 150-kD protein was also recognized by the anti-Ob-Rb IgG in Western immunoblots of hypothalamus, which is consistent with the immunocytochemical staining of Ob-Rb protein in the hypothalamus. However, the 150-kD band was only weakly represented in liver, which agrees with the relatively low expression of Ob-Rb in liver (Hoggard et al. 1997). The immunoblots of brain and liver show additional bands that may represent degradation products or multiple forms of Ob-Rb, perhaps as post-translational modifications or undiscovered alternative splice products. Characterization of these variants requires further analysis. However, the specificity of the anti-Ob-Rb IgG for Ob-Rb-like proteins in the brain is supported by the negative staining that resulted when the anti-GST IgG was substituted for anti-Ob-Rb IgG and when the anti-Ob-Rb IgG was absorbed with the GST/Ob-Rb fusion protein. The high concordance of Ob-Rb immunocytochemical staining with the distribution of Ob-Rb mRNA, as shown by in situ hybridization studies (Huang et al. 1996; Mercer et al. 1996a,b; Fei et al. 1997; Guan et al. 1997; Baskin et al. 1998), as well as the absence of Ob-Rb immunocytochemical staining in the choroid plexus, which expresses mainly Ob-Ra (Tartaglia 1997), are further evidence that the anti-Ob-Rb IgG detects Ob-Rb protein in the brain.
An increasing body of evidence indicates that the mechanism of leptin's effects in the hypothalamus involves the altered expression of hypothalamic neuropeptides (Ahima et al. 1996; Légrádi et al. 1997; Quintela et al. 1997; Thornton et al. 1997; Mizuno et al. 1998; Sahu 1998), including NPY mRNA (Schwartz et al. 1996a–c,1998) and proopiomelanocortin mRNA (Schwartz et al. 1997) in the arcuate nucleus. An effect of leptin on hypothalamic neurons has also been suggested by evidence that leptin activates c-fos expression in the hypothalamus (Van Dijk et al. 1996; Elmquist et al. 1997). Finding evidence for the Ob-Rb protein in hypothalamic neurons is an important link between these observations on mRNA expression and biological activity, because the presence of the translated receptor protein is presumably a prerequisite for leptin signaling to these neurons. Furthermore, the present finding of Ob-Rb protein in neurons that express NPY mRNA, together with evidence that these cells also express Ob-Rb mRNA, supports the hypothesis that arcuate nucleus NPY neurons are direct targets of leptin.
Footnotes
Acknowledgements
Supported by the Merit Review and Career Scientist Programs of the Department of Veterans Affairs and by NIH grants DK-17047, DK-12829, DK-17844, DK-17847, HD-09171, and NS-32273.
We thank P. Bailon, W.J. Fung, and L. Tartaglia for valuable reagents.