
Abstract
LMO4 belongs to the LIM-only family of zinc finger proteins that have been implicated in oncogenesis. The LMO4 gene is overexpressed in breast cancer and oral cavity carcinomas, and high levels of this protein inhibit mammary epithelial differentiation. Targeted deletion of Lmo4 in mice leads to complex phenotypic abnormalities and perinatal lethality. To further understand the role of LMO4, we have characterized Lmo4 expression in adult mouse tissues by immunohistochemical staining using monoclonal anti-Lmo4 antibodies. Lmo4 was highly expressed within specific cell types in diverse tissues. Expression was prevalent in epithelial-derived tissues, including the mammary gland, tongue, skin, small intestine, lung, and brain. High levels of Lmo4 were frequently observed in proliferating cells, such as the crypt cells of the small intestine and the basal cells of the skin and tongue. Lmo4 was highly expressed in the proliferative cap cell layer of the terminal end buds in the peripubertal mammary gland and in the lobuloalveolar units during pregnancy. The expression profile of Lmo4 suggests that this cofactor is an important regulator of epithelial proliferation and has implications for its role in the pathogenicity of cancer.
T
Targeted deletion of Lmo4 in mice results in perinatal lethality and complex phenotypic abnormalities. Approximately 50% of Lmo4 nullizygous mice exhibit exencephaly due to failure of the neural tube to close (Hahm et al. 2004; Tse et al. 2004), whereas all mice show sphenoid bone defects (Hahm et al. 2004). Homeotic transformations were evident in Lmo4-null embryos and newborn mice, including cranial nerve abnormalities and skeletal defects in the rib cage and cervical vertebrae (Hahm et al. 2004). These transformations are reminiscent of defects that occur in Hox-deficient mice (Trainor and Krumlauf 2000), suggesting that Lmo4 regulates the expression or activity of Hox proteins in specific cell types.
The importance of LMO proteins in regulating normal development is underscored by the findings that deregulated expression of these proteins can be oncogenic. The LMO1 and LMO2 genes were discovered by their translocation in acute T-cell leukemia and overexpression of either gene in transgenic mice results in T-cell tumors (Rabbitts 1998). Furthermore, two SCID (Severe Combined ImmunoDeficiency) patients undergoing gene therapy developed T-cell leukemia due to the ectopic activation of the LMO2 gene by retroviral integration (Hacein-Bey-Abina et al. 2003). LMO4 was originally identified in an expression screen using autologous serum from a breast cancer patient (Racevskis et al. 1999), and we have demonstrated that elevated levels of LMO4 occur in greater than 50% of primary breast tumors (Visvader et al. 2001). The mechanism underlying LMO4 overexpression in breast cancer reflects increased promoter activity in addition to aberrant activation of a second promoter (Wittlin et al. 2003). Somatic mutations in the LMO4 gene occur infrequently in breast cancers (Sutherland et al. 2003). Increased levels of LMO4 have also been reported in cancers of the oral cavity, in which high levels were apparent in less-differentiated tumors (Mizunuma et al. 2003).
In contrast to the more restricted expression profiles of the other LMO genes, Lmo4 mRNA is widely expressed in mouse embryonic and adult tissues (Grutz et al. 1998; Kenny et al. 1998; Sugihara et al. 1998; Racevskis et al. 1999; Bulchand et al. 2003). Interestingly, the pattern of Lmo4 expression during embryogenesis defined uncommitted cells, indicating that one function of Lmo4 may be to inhibit differentiation (Kenny et al. 1998). To perform a comprehensive analysis of Lmo4 expression in adult mouse organs at the protein level, we generated monoclonal antibodies (MAbs) specific for Lmo4. Immunohistochemical analysis revealed that although Lmo4 is widely distributed, its expression is restricted to specific cell subtypes. Lmo4 protein was found to be particularly prevalent in epithelial tissues, with high levels occurring in proliferating cells. Our results imply a role for Lmo4 in regulating proliferation in different epithelia.
Materials and Methods
Experimental Animals
All experiments with animals were conducted according to the guidelines of our Institutional Animal Ethics Committee. Wistar rats, C57BL/6, and BALB/c mice were obtained from our Institute's breeding facility at Kew (Victoria, Australia). Adult female mice were subjected to timed pregnancies that were scored by the observation of vaginal plugs and confirmed by examination of embryos when mammary glands were collected. The null Lmo4 and Lmo4:LacZ KI mice were a generous gift from K. Hahm and S. Orkin (Hahm et al. 2004).
Expression Constructs and Protein Purification
Full-length mouse Lmo4 cDNA was amplified by PCR using forward 5′-CGC GGA TCC AGC TCG CAG CCG CCC CCG GTG-3′ and reverse 5′-CGC GGA TCC TCA GCA GAC CTT CTG GTC TGC CAG-3′ primers, then subcloned into the BamHI site of pGEX-2T (Amersham Biosciences, Inc.; Uppsala, Sweden). The production of GST-Lmo4 protein and subsequent purification on a glutathione-Sepharose column (Amersham Biosciences, Inc.) was performed according to standard protocols. The expression constructs encoding either FLAG-tagged full-length LIM domain 1 (LIM1) or LIM domain 2 (LIM2) of mouse Lmo4 have been described (Sum et al. 2002).
Immunization and Hybridoma Fusion
Wistar rats were initially immunized by subcutaneous (SC) injection of 100 μg of purified GST-Lmo4 protein dissolved in complete Freund's adjuvant (Difco; Detroit, MI). Two subsequent boosts of the immunogen were injected SC 3 and 6 weeks later. A final boost of protein dissolved in phosphate-buffered saline (PBS) was given by intravenous and intraperitoneal injection 4 weeks later. Hybridomas were generated 3 days after the final boost by fusing spleen cells from immunized rats with the SP2/0 myeloma cell line (Galfre et al. 1977; O'Reilly et al. 1998). Hybridomas producing MAbs to Lmo4 were identified and their isotype determined by a FACScan screening strategy that we have previously described (O'Reilly et al. 1998; O'Reilly et al. 2002). Briefly, 293T cells were transiently transfected with an expression construct containing mouse Lmo4 cDNA with an N-terminal FLAG tag. Transfected cells were fixed in 1% paraformaldehyde, permeabilized with 0.3% saponin (Sigma), and stained with hybridoma supernatants. Bound antibodies were detected by fluorescein-isothiocyanate (FITC)-conjugated goat anti-rat Ig antibodies (Southern Biotechnology Associates, Inc.; Birmingham, AL) and analyzed in a FAC-Scan analyzer (Becton Dickinson: Bedford, MA). Hybridomas producing Lmo4-specific antibodies were cloned twice and subsequently adapted for growth in low-serum medium. For production of large amounts of antibodies, hybridomas were cultured for several weeks in the miniPERM classic 12.5 kD production and nutrient module (Heraeus; Asheville, NC). Antibodies were purified on a protein G-Sepharose column (Amersham Biosciences, Inc.) according to the manufacturer's protocols.
Flow Cytometry
Balb/c mouse-derived thymocytes were incubated with a mixture of PE-conjugated CD4+ (BD Pharmingen; San Diego, CA) and FITC-conjugated CD8+-specific MAbs (kindly provided by A. Strasser) for 30 min at 4C. The immunostained thymocytes were subjected to a four-way-sort on a FACS sorter (DIVA; Becton Dickinson) after gating for PI-positive cells to isolate each of the four thymic subsets: CD4−CD8− double negative cells, CD4+CD8+ double-positive cells, and single-positive CD4+ and CD8+ cells. Protein lysates (from 105 cells) were subjected to Western blot analysis.
Western Blotting
Protein lysates from wild-type and Lmo4-deficient mouse E16.5 embryos were prepared by crushing frozen embryos in liquid nitrogen with a mortar and pestle before transfer into 1 ml ice-cold lysis buffer (150 mM NaCl, 5 mM EDTA, 50 mM Tris/HCl (pH 7.5), 1% NP40, 1 mM DTT, supplemented with Complete protease inhibitor tablet (Roche Diagnostics; Mannheim, Germany), 10 mM NaF, and 1 mM Na3VO4). For the preparation of tissue lysates, frozen tissues derived from adult mouse organs were pulverized in liquid nitrogen before transfer into ice-cold ONYX lysis buffer (20 mM Tris-HCl (pH 7.4), 135 mM NaCl, 1.5 mM MgCl2, 1 mM EDTA, 10% glycerol, and 1% Triton X-100; supplemented with Complete inhibitor tablet, 50 mM NaF, and 1 mM Na3VO4) or directly homogenized in ONYX (ONYX Pharmaceuticals; Richmond, CA) lysis buffer at 4C as detailed elsewhere (Print et al. 1998).
Tissue lysates (30-50 μg total protein) for Western blot analysis were denatured by boiling in sodium dodecyl sulfate (SDS) loading buffer and separated on polyacrylamide gels (Novex; Carlsbad, CA) before transfer to polyvinylidene difluoride membranes (Millipore; Bedford, MA). Nonspecific binding of proteins to membranes was blocked by incubation in PBS containing 5% skim milk and 0.1% Tween-20. The membranes were then probed with anti-Lmo4 MAbs (1-2 μg/ml), followed by horseradish peroxidase-coupled secondary antibodies (Amersham Biosciences, Inc.), and developed by ECL (Amersham Biosciences, Inc.). To control for the integrity of proteins in cell and tissue lysates, the blots were probed with mouse anti-Hsp70 N6 (kindly provided by Dr R. Anderson, Peter MacCallum Cancer Centre, Melbourne, Australia and Dr. W. Welch, UCSF, San Francisco, CA), anti-tubulin (Sigma; St. Louis, MO), or anti-actin (Sigma) MAbs.
Immunohistochemistry
For immunohistochemistry, mouse tissues were fixed in 4% paraformaldehyde. Sections were deparaffinized, rehydrated, and subjected to antigen retrieval by boiling in 10 mM citrate buffer (pH 6.0) for 20 min. After cooling to room temperature (RT), sections were treated with 3% hydrogen peroxide to block endogenous peroxide activity, followed by a blocking step in PBS containing 0.2% Triton X-100 (T-PBS) and 10% normal rabbit serum (NRS). Sections were then incubated overnight at 4C with rat anti-Lmo4 20F8 or 16H2 (5-10 μg/ml), rat anti-mouse Ki-67 (1:50 dilution; DAKO) or an isotype-matched control rat antibody (IgG2a/κ, Pharmingen) diluted in T-PBS containing 5% NRS. Staining was detected by incubating with biotinylated rabbit anti-rat IgG antibody (DAKO), followed by HRP-conjugated strepavidin (LSAB2, DAKO). Finally, sections were stained with diaminobenzidene (DAKO), counterstained with hematoxylin, and mounted in DPX (BDH; Poole, UK).
Wholemount Staining for β-Galactosidase Activity
Tissues from Lmo4:LacZ KI mice were fixed in PBS containing 2% paraformaldehyde and 0.25% glutaraldehyde for 2 hr at 4C prior to staining for β-galactosidase activity by incubation with X-gal solution (5 mM K3Fe, 5 mM K4Fe, 2 mM MgCl2, and 0.02% NP40 in PBS, supplemented with 0.5 mg/ml X-gal) at 37C overnight. These tissues were then postfixed in 4% paraformaldehyde overnight at 4C and then embedded in paraffin. Sections (7 μm) were prepared and counterstained with nuclear fast red.
Results
Characterization of Monoclonal Antibodies to Lmo4
We have identified several monoclonal antibodies (MAbs) specific for native Lmo4 using an immunofluorescence staining and flow cytometric screening protocol, as previously described (O'Reilly et al. 2002). These antibodies recognize both the mouse and human LMO4 proteins because the amino acid sequences of these proteins are identical. Based on initial studies, we selected four MAbs (11E2, 12B7, 16H2, and 20F8) for further characterization. All four MAbs recognized Lmo4 in Western blot analysis of protein lysates from 293T cells transfected with an expression construct encoding FLAG-tagged mouse Lmo4 (data not shown). Similarly, Western blot experiments using 293T cells transfected with constructs encoding either the first or second LIM domains of Lmo4 revealed that 11E2, 16H2, and 20F8 were specific to LIM1, whereas 12B7 recognized LIM2 (data not shown). Immunoblotting of protein lysates from E16.5 mouse embryos demonstrated that the MAbs 16H2 and 20F8 detected a single protein of ~17 kD, corresponding to endogenous Lmo4. These MAbs were highly specific for Lmo4 because no cross-reactivity to other proteins was observed and Lmo4 protein was not detected in lysates from Lmo4-deficient embryos at E16.5 (Figure 1A) (Hahm et al. 2004). Each of the four selected MAbs, (11E2, 12B7, 16H2, and 20F8) was capable of immunoprecipitating both Flag-tagged Lmo4 protein from transfected 293T cells (data not shown) and endogenous LMO4 protein from these cells. A summary of the different applications of Lmo4 MAbs 11E2, 12B7, 16H2, and 20F8 is given in Table 1.
Western Analysis of Lmo4 Expression in Mouse Tissues
To examine the expression of Lmo4 in mouse tissues, we performed Western blot analysis with 20F8 MAb (Figure 1B). Lmo4 was expressed widely, corresponding to its mRNA expression profile in mouse and human tissues (Grutz et al. 1998; Kenny et al. 1998; Sugihara et al. 1998; Racevskis et al. 1999; Bulchand et al. 2003). High levels of Lmo4 were evident in brain, salivary gland, lung, liver, pancreas, small intestine, colon, and lymphoid organs (thymus, spleen, and lymph node) (Figure 1B). Lower levels of Lmo4 protein were found in kidney and testes, whereas no LMO4 expression was detected in the heart (Figure 1B). Because expression was high in lymphoid organs, we determined the phenotype of Lmo4-expressing cells in the thymus. Analysis of FACS-sorted thymocytes revealed highest levels in immature CD4− and CD8− negative cells and CD4+CD8+ double-positive cells compared with that in the mature single-positive CD4+ and CD8+ populations (Figure 1C). These data are compatible with previous findings that Lmo4 mRNA is expressed in specific thymic sublineages using reverse transcription-PCR (Kenny et al. 1998).
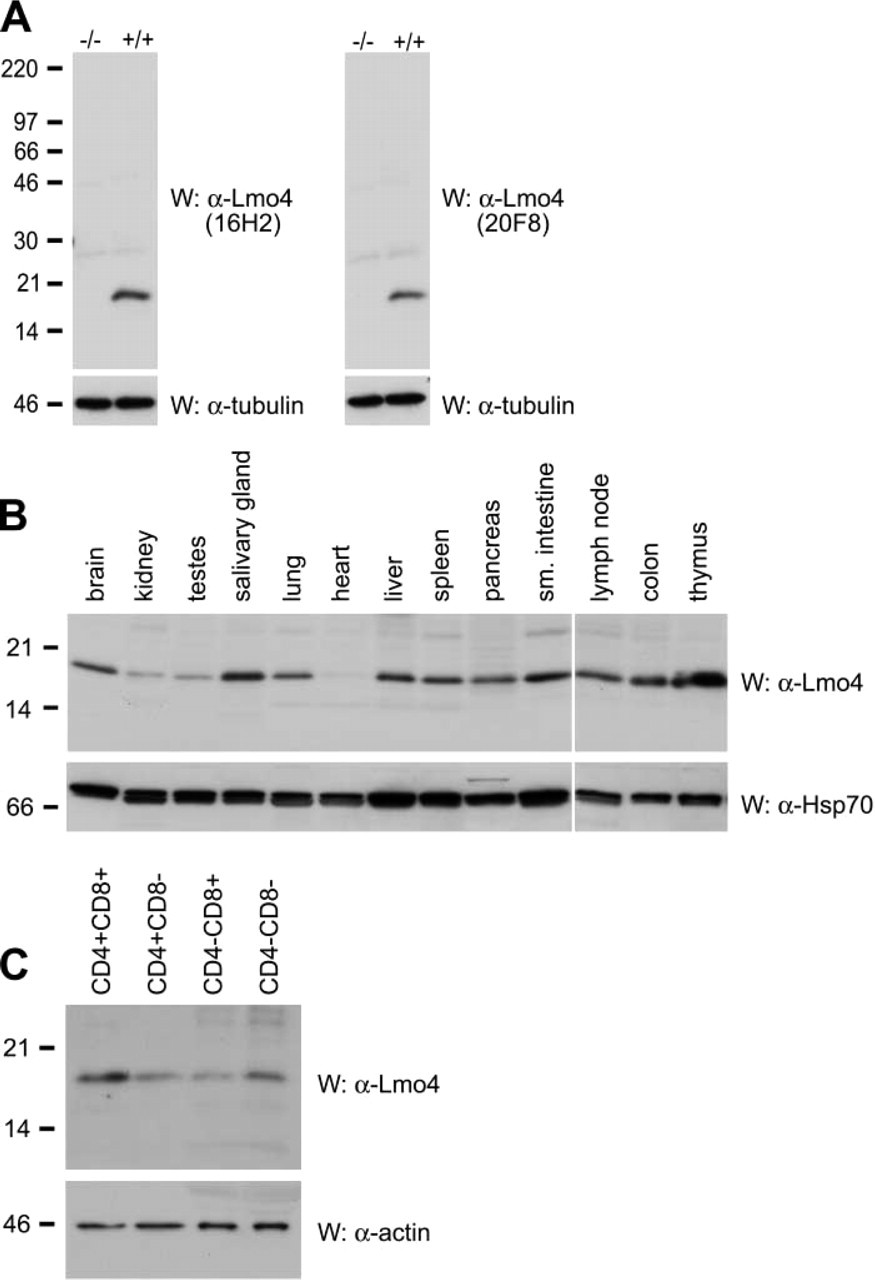
Expression of Lmo4 in mouse organs.
Immunohistochemical Analysis of Lmo4 Expression in Mouse Tissues
The Developing and Adult Central Nervous System. The specificity of Lmo4 immunostaining by 20F8 MAb was validated using sections from wild-type and Lmo4-deficient E16.5 mouse embryos (Figures 2A-2D). As expected, no immunostaining was detectable in Lmo4-deficient embryos (Figures 2B and 2D), whereas strong staining was evident in wild-type tissue (Figures 2A and 2C). Approximately 50% of these embryos exhibited exencephaly (Hahm et al. 2004), as evident in Figure 2D. The immunohistochemical profile of Lmo4 was confirmed using a second anti-Lmo4 MAb, 16H2 (data not shown). In addition, an isotype-matched rat non-immune IgG provided a negative control for each experiment (Figure 2, insets).
Applications of LMO4 monoclonal antibodies

IP, immunoprecipitation; IF, immunofluorescence; IHC, immunohistochemistry; W, Western blot analysis; hu, human; mo, mouse; LIM1 and LIM2, there are two LIM domains in LMO4 (see text).
In the developing mouse brain, Lmo4 immunostaining was predominantly observed in the postmitotic neurons of the neopallial cortex (future cerebral cortex) of wild-type E16.5 embryos, with some staining of migrating neurons in the subventricular zone of the differentiating field (Figure 2C). In the olfactory bulb and spinal cord, abundant Lmo4 protein was evident in the postmitotic and migrating neurons in the differentiating field (data not shown). This pattern of Lmo4 protein expression is consistent with in situ RNA hybridization studies of the developing mouse brain (Kenny et al. 1998; Sugihara et al. 1998; Hermanson et al. 1999; Bulchand et al. 2003).
In the adult central nervous system (CNS), abundant Lmo4 expression was observed in neurons and astrocytes in the hippocampus (Figures 2E and 2G) and cerebral cortex (data not shown). Lmo4 primarily localized to the nuclei in these cells, with little or no staining of the cytoplasm. Conversely, the larger motor neurons and Purkinje cells (Figure 2H) exhibited high levels of cytoplasmic Lmo4 protein. Within chick embryos, Lmo4 has been localized to both muscle sensory and motor neurons (Chen et al. 2002). Lmo4 was also detected in the choroid plexus and ependymal cell layers, which are derived from neuroepithelium (Figure 2F), but no Lmo4 protein was detected in oligodendrocytes (Figures 2E-2G). This expression profile is consistent with that described for LMO4 in both mouse and human brain (Hermanson et al. 1999; Leuba et al. 2004).
Lmo4 Expression in Various Mouse Organs
The widespread expression of Lmo4 determined by Western blot analysis (Figure 1B) was further characterized by immunohistochemistry using the MAb 20F8 (Figures 3–5 and Table 2). The expression profile of Lmo4 was also confirmed by staining tissue sections from Lmo4:LacZ KI mice, in which the lacZ gene is driven by the endogenous Lmo4 promoter (Hahm et al. 2004) for β-galactosidase activity (Figures 3–5). An identical pattern of expression was observed in all cases. In addition, staining with an isotype-matched rat non-immune IgG provided a negative control for each tissue (Figures 3–5).
Hematopoietic Tissues. Heterogeneous expression levels of Lmo4 in the thymus were observed by LacZ staining of Lmo4:LacZ KI mice (Figure 3A) and immunohistochemical analysis using 20F8 MAb (Figure 3B). Consistent with Western blot analysis of sorted thymocytes (Figure 1C), moderate to strong Lmo4 immunostaining was detected in cortical thymocytes, which contain the more immature pre-T-cell population (Figure 3B). In contrast, Lmo4 expression was lower in the thymic medulla where the more mature single positive T cells are located (Figure 3B). There was no evidence of Lmo4 expression in the epithelial cells of Hassall's corpuscle in the thymic medulla. In the lymph node, strong Lmo4 expression was confined to the T-cell zone of the paracortex (Table 2). No Lmo4 protein was detected in lymphoid follicles, which predominantly consist of B lymphocytes. Lmo4 expression in the spleen was evident in periarteriolar lymphoid sheaths of the white pulp, comprised mainly of T lymphocytes (Table 2). Consistent with expression in the lymph node, Lmo4 immunostaining was absent in splenic lymphoid follicles. Megakaryocytes scattered in the red pulp did not reveal any Lmo4 expression.
Respiratory Tract. Lmo4 expression was confined to the low columnar epithelial cells lining the trachea and lumen of the respiratory bronchioles and alveoli in the lung (Figures 3D and 3E). Lmo4 immunostaining was also present in the non-ciliated Clara cells, identified by a characteristic bulging into the lumen, as well as in the terminally differentiated ciliated cells (Figure 3E).
Pancreas. Intense Lmo4 immunostaining was detected in the endocrine islet of Langerhans cells but was negligible in the exocrine acinar cells (Figure 3H), consistent with the expression of LacZ in pancreatic tissue from LMO4:LacZ KI mice (Figure 3G). A lower level of Lmo4 protein expression was also detected in epithelial cells lining the pancreatic ducts.
Kidney. Lmo4 expression was readily detectable in all tubular structures of the kidney, including the proximal and distal convoluted tubules, by both LacZ staining and immunohistochemistry using 20F8 MAb (Figures 3J and 3K). Stronger staining of juxtaglomerular tubules was observed (Figure 3K), whereas the glomeruli were devoid of Lmo4 expression. The majority of cells in the kidney expressed relatively low levels of Lmo4, consistent with the level observed by Western analysis (Figure 1B).
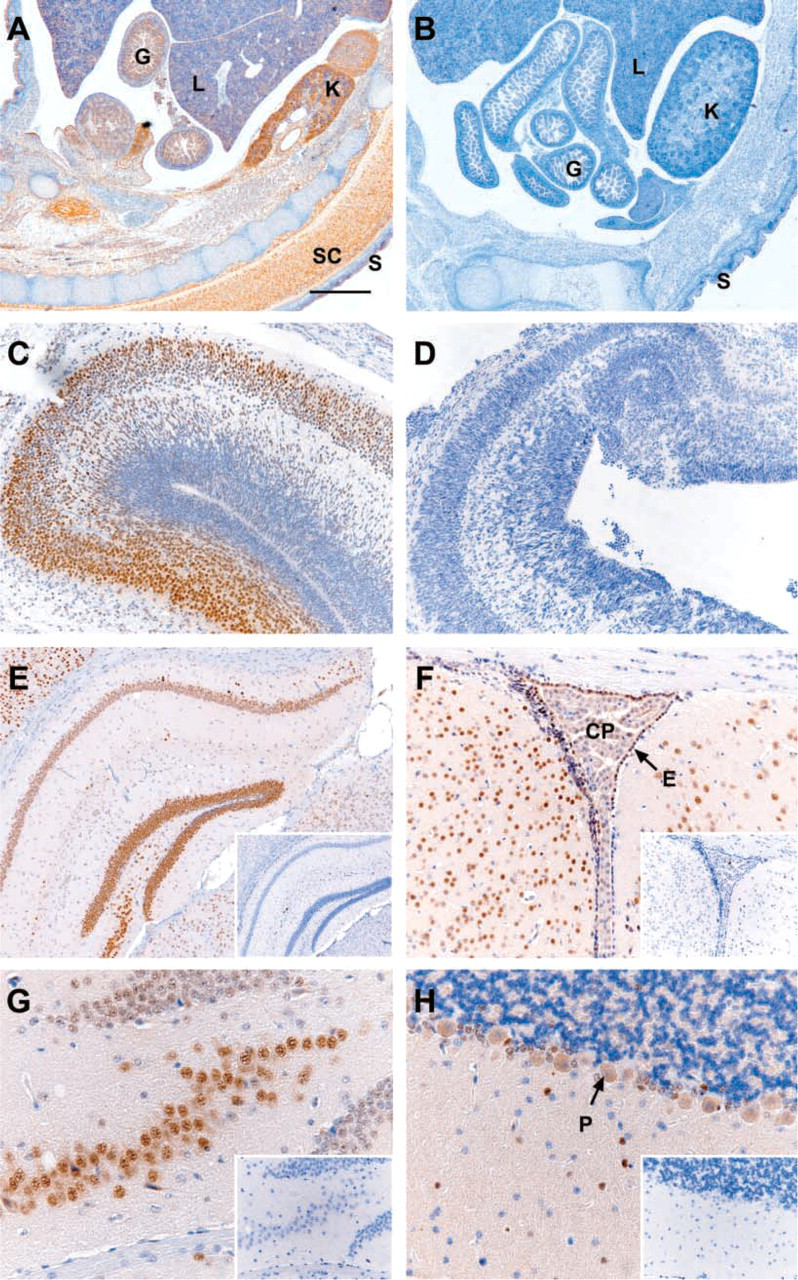
Lmo4 immunostaining of the central nervous system.
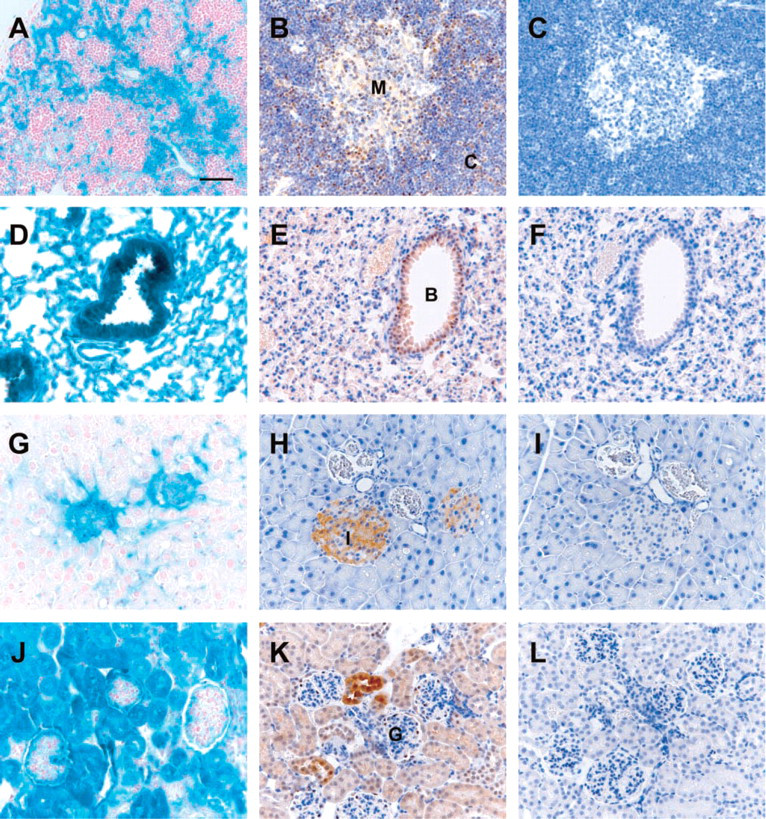
Expression of Lmo4 in adult mouse tissues.
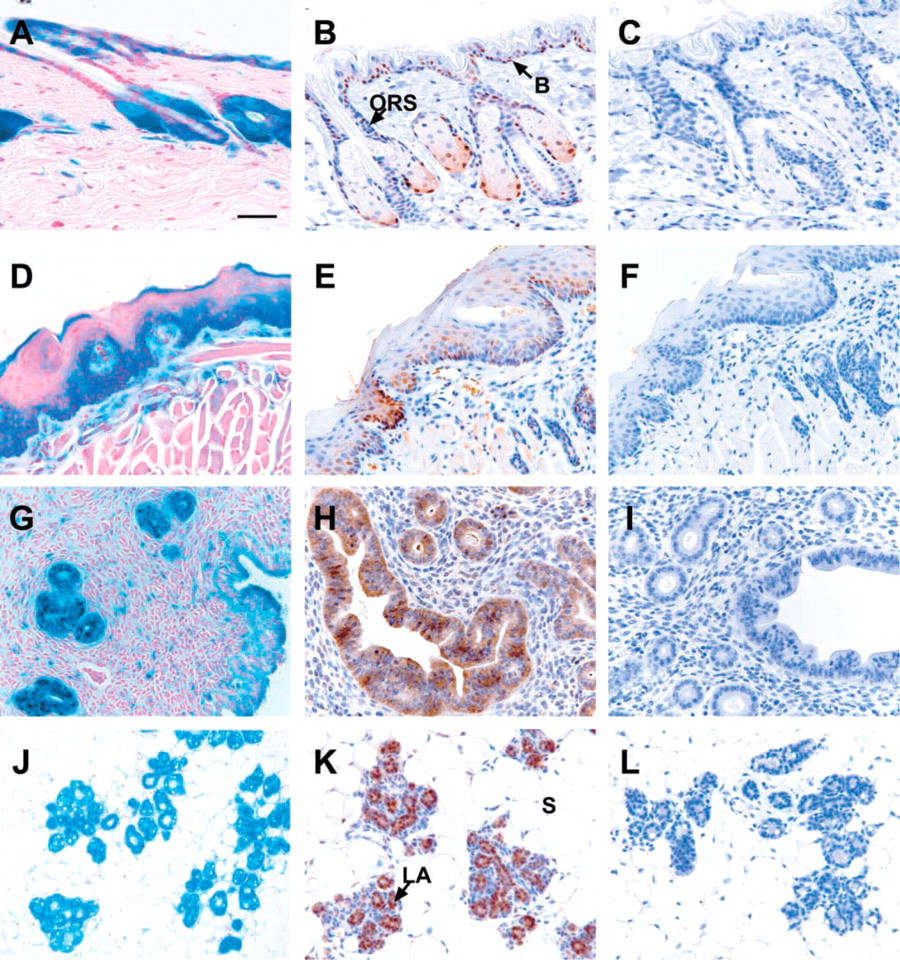
Lmo4 expression in epithelial tissues from adult (10-week-old) mice.
Lmo4 Is Highly Expressed in Proliferating Cells of Epithelial Tissues
Skin and Hair Follicles. Lmo4 was predominantly expressed in hair follicles (Figures 4A and 4B) and squamous epithelial cells in the basal layer of the epidermis (Figures 4A, 4B, and 5B). Within the hair follicle, Lmo4 immunostaining was evident in the sebaceous glands, undifferentiated bulb cells, and the outer epithelial root sheath (Figure 4B), the latter of which may contain a keratinocyte stem cell population (Rochat et al. 1994). Epidermal basal cells have abundant Lmo4 protein and are highly proliferative as indicated by the expression of the proliferation marker Ki-67 (Figure 5A).
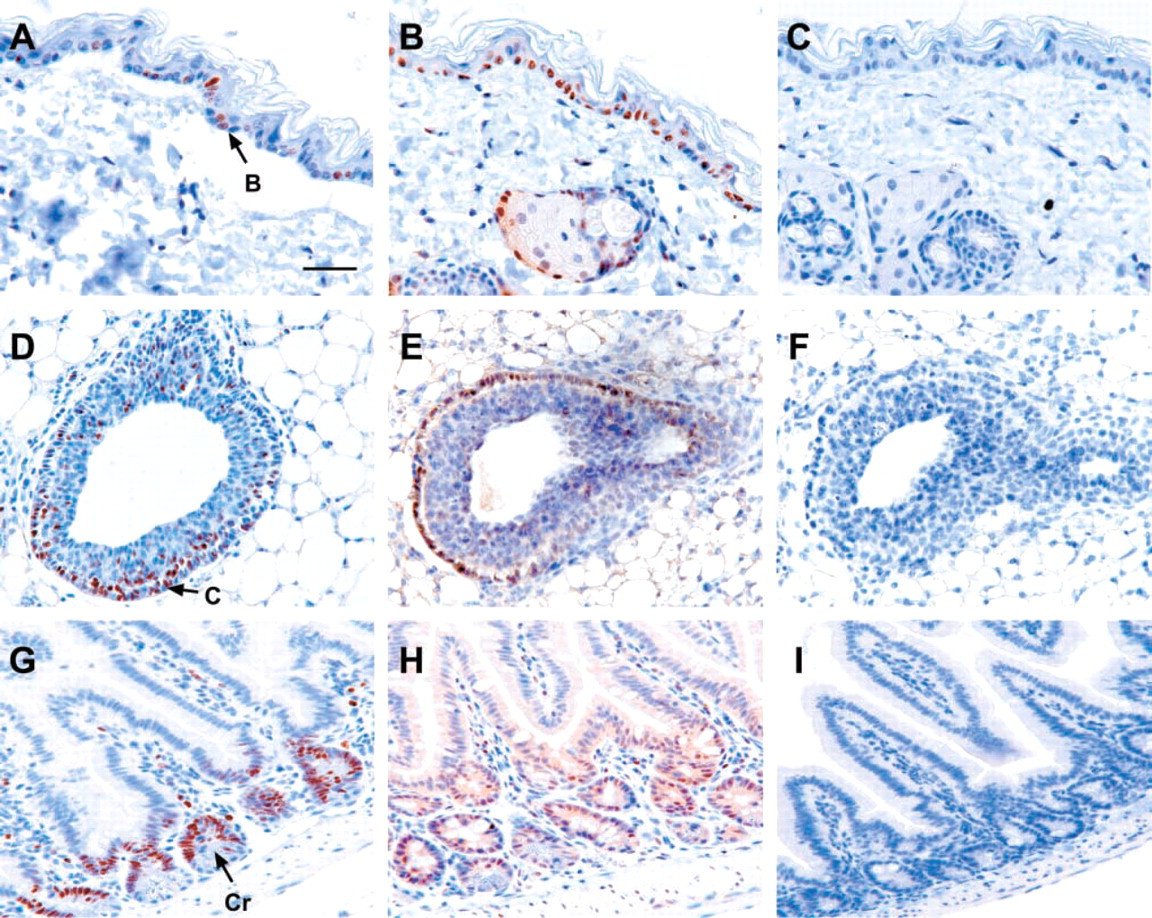
Lmo4 is expressed in proliferating epithelial cells.
Tongue. Lmo4 expression was evident in the stratified squamous epithelial cells of the tongue, with the majority of expression confined to the basal layer of the germinativum (Figures 4D and 4E). Nuclear Lmo4 expression was also detected in taste buds and serous glands within the tongue (Figure 4E), but no expression was evident in the filiform or dermal papilla.
Reproductive Organs. Lmo4 was expressed in both the uterine myometrium and endometrium in the female reproductive system (Figures 4G and 4H). Strong cytoplasmic immunostaining was observed in the endometrial cells lining the uterine glands and the lumen of the uterus (Figure 4H), both of which are highly proliferative and turn over regularly during the estrus cycle. Expression of Lmo4 in the ovary was less striking, with low to moderate staining of the corpus luteal and follicular cells and negligible staining of the single epithelial layer (Table 2). In male reproductive organs, Lmo4 expression was confined to spermatogonium and spermatocytes in the testes, whereas low levels were evident in the epididymis and epithelial cells lining the dorsal and ventral prostate (Table 2).
Mammary Gland. The mid-pregnant mammary gland exhibited prominent Lmo4 expression in both the ductal and lobuloalveolar epithelial cells, with substantially lower levels in the myoepithelium and surrounding stroma (Figures 4J and 4K). Lmo4 expression was also detected within the terminal end bud (TEB) of the peripubertal mammary gland (Figure 5E). Intense Lmo4 staining was observed in the cap cells, which form the outer monolayer at the tip of the bulbous structure (Figure 5E). These are highly proliferating cells, as shown by their expression of the Ki-67 antigen (Figure 5D).
Localization and intensity of Lmo4 expression in adult mouse tissues by immunohistochemistry

-, no detectable expression; +, low; ++, moderate; + + +, high expression.
Small Intestine. Strong Lmo4 expression was detected in the crypts of Lieberkühn located at the base of villi (Figure 5H). These epithelial crypt cells also express abundant Ki-67 (Figure 5G), indicating a high proliferation rate. In addition to crypt cells, nuclear Lmo4 immunostaining was detected in the smooth muscle surrounding the small intestine (Figure 5H). Brunner's glands within the duodenum, representing modified extensions of crypt cells, also expressed moderate levels of Lmo4 (Table 2).
Discussion
Lmo4, in parallel with Lmo2 in hematopoietic cells, has been shown to act as a negative regulator of differentiation, suggesting a role for this gene in maintaining the proliferative state. We have demonstrated that overexpression of Lmo4 in SCp2 mammary epithelial cells blocks differentiation to milk-producing cells (Visvader et al. 2001). Similarly, neuroblastoma cells expressing high levels of LMO4 had diminished capacity to differentiate into neurites (Vu et al. 2003). Deregulation of LMO4 protein expression has been implicated in oncogenesis, whereby elevated levels of LMO4 have been reported in breast cancer (Visvader et al. 2001) and carcinomas of the oral cavity (Mizunuma et al. 2003). Furthermore, we have evidence from RNA interference studies that downregulation of LMO4 expression in breast cancer cells inhibits proliferation (unpublished data). Here we report that Lmo4 is highly expressed in proliferating epithelial tissues as well as in specific cell types of diverse mouse organs.
High levels of Lmo4 protein occur in epithelial tissues and in cells that appear to have a high proliferative capacity. The epithelial tissues include lung, kidney, skin, tongue, small intestine, uterus, and mammary gland. In these tissues, Lmo4 was localized primarily to the nucleus or distributed throughout both the nucleus and cytoplasm. Within several of the epithelial tissues, high levels of Lmo4 were observed in proliferating cells, particularly in their nuclei. Stratified squamous epithelial cells of the tongue and uterine endometrium exhibit rapid turnover and express abundant Lmo4. Mitotically active cells in the basal layer of the epidermis and undifferentiated cells in the outer root sheath of the hair follicle, thought to contain a stem cell population (Rochat et al. 1994), exhibited strong nuclear staining of Lmo4. Similarly, crypt cells at the base of the villi in the small intestine and cap cells of TEBs in the peri-pubertal mammary gland showed prominent Lmo4 expression. These cells have a high proliferative index as indicated by staining for the Ki-67 antigen. In the mammary gland, both ductal and lobuloalveolar epithelial cells expressed abundant Lmo4. The formation and expansion of the lobuloalveolar units during pregnancy is accompanied by a high rate of proliferation.
Lmo4 expression is not restricted to proliferating cells. High Lmo4 protein levels were detected in postmitotic and migrating neurons of the developing brain and in several different neuronal cell types within the adult CNS, including astrocytes, motor neurons, and Purkinje cells. Interestingly, a high level of Lmo4 expression has been reported in the entorhinal cortex and CA1 hippocampal region of the human brain, which are altered in Alzheimer's disease (Leuba et al. 2004). Comparison of Lmo4 expression in these regions revealed reduced levels in brains from Alzheimer's disease patients relative to control brains (Leuba et al. 2004). Lmo4 has been found to be differentially expressed at an early stage in the development of muscle sensory neurons and may actively participate in their differentiation by regulating the transcriptional activity of LIM homeodomain proteins (Chen et al. 2002). In several other tissues, such as smooth muscle, kidney and pancreas, mature cells were also found to express abundant Lmo4. Thus, Lmo4 may have a dual physiological role in which it can either promote proliferation or differentiation.
Lmo4 is widely expressed in both embryonic and adult mouse tissues but in restricted cell types and at levels that vary markedly. The expression profile of Lmo4 suggests that this protein is multifunctional and that it influences both proliferation and differentiation, dependent on cellular context. The underlying cause of death in Lmo4 null mice, all of which succumb within a few minutes of birth, is yet to be established. Histological examination of major organs, including the hematopoietic compartment, did not reveal any gross morphological defects in these mice (Hahm et al. 2004; Tse et al. 2004). On the basis of high Lmo4 expression in the lungs and brain, it is tempting to speculate that proper lung expansion or brain stem function may be affected in these mutants, leading to perinatal lethality. Specific deletion of the Lmo4 locus in lung or specific regions of the brain should provide insight into the physiological role of Lmo4 in these organs.
Footnotes
Acknowledgments
This work was supported by the Victorian Breast Cancer Research Consortium Inc. and the National Health and Medical Research Council (Australia).
We wish to thank F. Feleppa and U. Devisekera for expert technical assistance and S. Mihajlovic for histology.