
Abstract
Breast cancer is a major cause of cancer-related mortality in women. There are major discrepancies concerning the usefulness of various antibodies in detecting breast cancer susceptibility gene 1 (BRCA1) protein and its subcellular localization. The aim of the present study was to determine the specificity and sensitivity of immunohistochemistry (IHC) as a screening method for demonstrating BRCA1 expression. BRCA1 gene expression in archival paraffin-embedded breast cancer tissues was studied simultaneously at the protein and mRNA levels, and the two findings were compared. Forty-eight archival paraffin-embedded breast cancer tissues were studied for BRCA1 gene expression at protein level by IHC using four different antibodies against different BRCA1 epitopes and at mRNA level using real-time RT-PCR. BRCA1 mRNA expression was reduced or absent in 79% of the samples, and this finding correlated significantly with loss of BRCA1 protein expression in 83% of breast cancer tissues using one BRCA1 antibody studied (AB-1, against N-terminus epitope). The specificity of this antibody was 91.3%, and its sensitivity was 66.6%. There was no significant correlation between BRCA1 mRNA and protein expression as demonstrated by the remaining three antibodies. Antibody 8F7 had the highest sensitivity of 100%, but its specificity was 30.4% if mRNA levels were considered as the reference standard.
B
There are major discrepancies concerning the usefulness of various antibodies in detecting BRCA1 protein expression and localization. Different studies have generated conflicting results related to BRCA1 expression and the subcellular localization of this protein. This controversy is even more pronounced when paraffin-embedded tissues are used (Wilson et al. 1999). Thus, although some researchers conclude that the specificity of some of the BRCA1 antibodies is adequate to consider immunohistochemistry (IHC) as a valuable screening method (Yoshikawa et al. 1999), others believe that commercially available BRCA1 antibodies lack the specificity required to identify the BRCA1 protein (Perez-Valles et al. 2001).
BRCA1 has been claimed to be exclusively a nuclear protein in both normal and cancer cells (Scully et al. 1996; Thomas et al. 1996; Thakur et al. 1997; Wilson et al. 1999), a nuclear protein in normal cells but an aberrantly localized cytoplasmic protein in breast and ovarian tumor cells (Chen et al. 1995; Lee et al. 1999), or a cytoplasmic protein found in tube-like structures that invaginate the nucleus (Coene et al. 1997). The role of BRCA1 in DNA repair is becoming clearer: BRCA1 protein is involved in several DNA repair pathways and also has an effect on global genomic repair, involving transcriptional regulation of nucleotide excision repair genes (Hartman and Ford 2002,2003; Zhang et al. 2004). Also, it has recently been reported that BRCA1 expression influences the choice of chemotherapeutic agents used in the treatment of breast cancer (Egawa et al. 2003; Zhou et al. 2003). Therefore, it is now becoming clear that the knowledge of BRCA1 expression in breast cancer has important clinical ramifications.
This study focused on BRCA1 expression status in paraffin-embedded breast cancer tissues by using RT-PCR and IHC and correlated the results of the two. Such a comparison could be useful in assessing the specificity of commercially available BRCA1 antibodies.
Materials and Methods
Forty-eight archival formalin-fixed paraffin-embedded breast cancer tissues of female patients (aged 29–65 years) were randomly chosen and used for the study. The genetic background of the population studied was unknown. Sections of 5-μm thickness were cut from each tissue. One section was stained with hematoxylin and eosin to confirm the diagnosis, type, and grading of the tumor. The remaining sections were stored for use in IHC and RNA extraction. Histological grading of the carcinomas was done according to the system of Scarff-Bloom-Richardson (Dalton et al. 1994). The MCF-7 cell line, grown in RPMI medium (Gibco BRL; Carlsbad, CA) and known to express BRCA1 (Scully et al. 1996), was used as positive control for IHC. In addition, RNA extracted from this cell line was used as positive control RNA in the real-time RT-PCR method.
Immunohistochemistry
To study BRCA1 protein expression, four antibodies were applied to formalin-fixed paraffin-embedded breast cancer sections (Table 1). Breast cancer sections were deparaffinized, then rehydrated through three different concentrations of alcohol and 0.3% H2O2 for 30 min to block endogenous peroxidase. Epitope retrieval was carried out in 0.1 M citrate buffer at 95C in water bath for 20 min. Nonspecific binding was blocked using rabbit anti-mouse serum before monoclonal antibodies (AB-1, AB-8F7), and swine anti-rabbit serum before polyclonal antibodies (AB-D20, AB-C-terminus) for 30 min. After overnight incubation, biotinylated antibody (link antibody) was added, followed by streptavidin (dilution 1:500). Diaminobenzene was used as chromogen. Sections from MCF-7 breast cancer cell line paraffin blocks were used as positive control. For negative controls, the antibodies were substituted with the corresponding serum. It is worthy of note that several buffers and antigen retrieval methods were tested before this optimal protocol was achieved. These methods include microwave treatment at 90C for 10 min and at 95C for 20 min.
Subcellular localization of staining was categorized as cytoplasmic, nuclear, or both cytoplasmic and nuclear.
RNA Extraction and Real-time RT-PCR
Of the 48 breast cancer tissues studied using IHC, 29 samples had enough tissue for successful RNA extraction and RT-PCR amplification of GAPDH. To control for RNA degradation, GAPDH RNA had to be amplified in RNA samples extracted from breast tissue sections before BRCA1 expression in that tissue was deemed negative. Macrodissection of tumor tissue from several paraffin sections was used to minimize the influence of surrounding normal tissues. In these samples, 70% or more of the tissue sections contained tumor. The sections were rehydrated and scraped off the glass slide, using a needle, into an Eppendorf tube containing RNase inhibitor, proteinase K buffer (pH 8.3), 1% Tween surfactant, and proteinase K (10 mg/ml). The tubes were kept at 60C in water bath overnight, then mixed with 400 μl Trizol and chloroform to precipitate the RNA. There was no need to treat the samples with Dnase, because the BRCA1 probe and GAPDH primers spanned an exon–exon junction. RT-PCR master mixes were prepared according to the Invitrogen ThermoScript Platinum Quantitative One-Step RT-PCR System (Invitrogen; Carlsbad, CA) BRCA1 primers (Egawa et al. 2001) (forward, 5'-ACAGCTGTGTGGTGCTTCTGTG-3', reverse 5'-CATTGTCCTCTGTCCAGGCATC-3') and BRCA1 probe labeled with FAM (Egawa et al. 2001) (5'-CATCATTCACCCTTGGCACAGGTGT-3') were used to amplify and detect BRCA1 mRNA. GAPDH primers (forward 5'-TCATTGACCTCAACTACATGGTTT-3', reverse 5'-GAAGATGGTGATGGGATTTC-3') and GAPDH probes labeled with JOE [TaqMan] (JOE-CAAGCTTCCCGTTCTCAGCC-TAMRA) were used as internal control for RT-PCR. Samples for no-RT and no-template were also included in each test to detect any DNA or RNA contamination. The Applied BioSystems ABI7000 real-time sequence detection system (Applied Biosystems, Foster City, CA) was used to detect amplifications. Amplification conditions were 60C for 60 min for RT, followed by 95C for 5 min, 45 PCR cycles at 95C for 15 sec, and 60C for 1 min. The cut-off value for BRCA1 mRNA negativity was when there was no specific or significant reduction in BRCA1 signals detected even after 45 cycles of amplification. Each sample was assayed in triplicate in independent reactions.
Details of four different anti-BRCA1 antibodies

Histological grade, BRCA1 mRNA and protein expression, and BRCA1 protein subcellular localization of breast cancer tissues studied
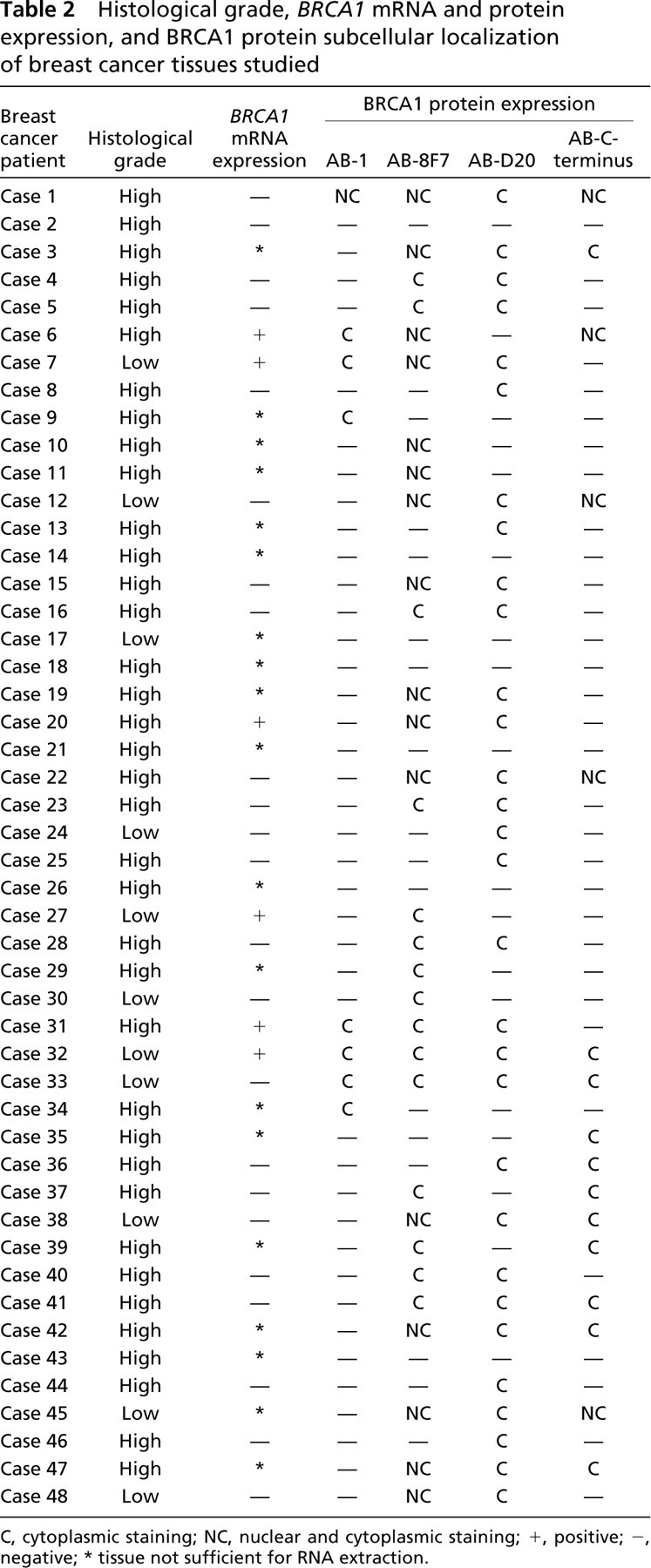
C, cytoplasmic staining; NC, nuclear and cytoplasmic staining; +, positive; -, negative, ∗ tissue not sufficient for RNA extraction.
Statistical Analysis
The data were analyzed using the Statistical Package for Social Sciences (SPSS version 11.01) software. The association between BRCA1 mRNA and protein expression was analyzed using the χ2 test. A p-value less than 0.05 was considered significant.
Results
Breast cancers were categorized according to their histological type. Five main histological types were present in this study: infiltrative ductal carcinoma (35 cases), intraductal carcinoma (1 case), medullary carcinoma (7 cases), infiltrative lobular carcinoma (3 cases), and tubulo-lobular carcinoma (2 cases). The most common histological type was infiltrative ductal carcinoma, which accounted for 72.8% of cases. In this study, cancers with total score of 3 to 5 were considered low grade; and those with scores of 6 to 9 were considered high grade. Of the 48 breast cancer cases, 37 (77%) were of high histological grade (Table 2).
Immunohistochemistry
Table 3 summarizes the BRCA1 protein expression and its subcellular localization in 48 breast cancer tissues using the four different antibodies.
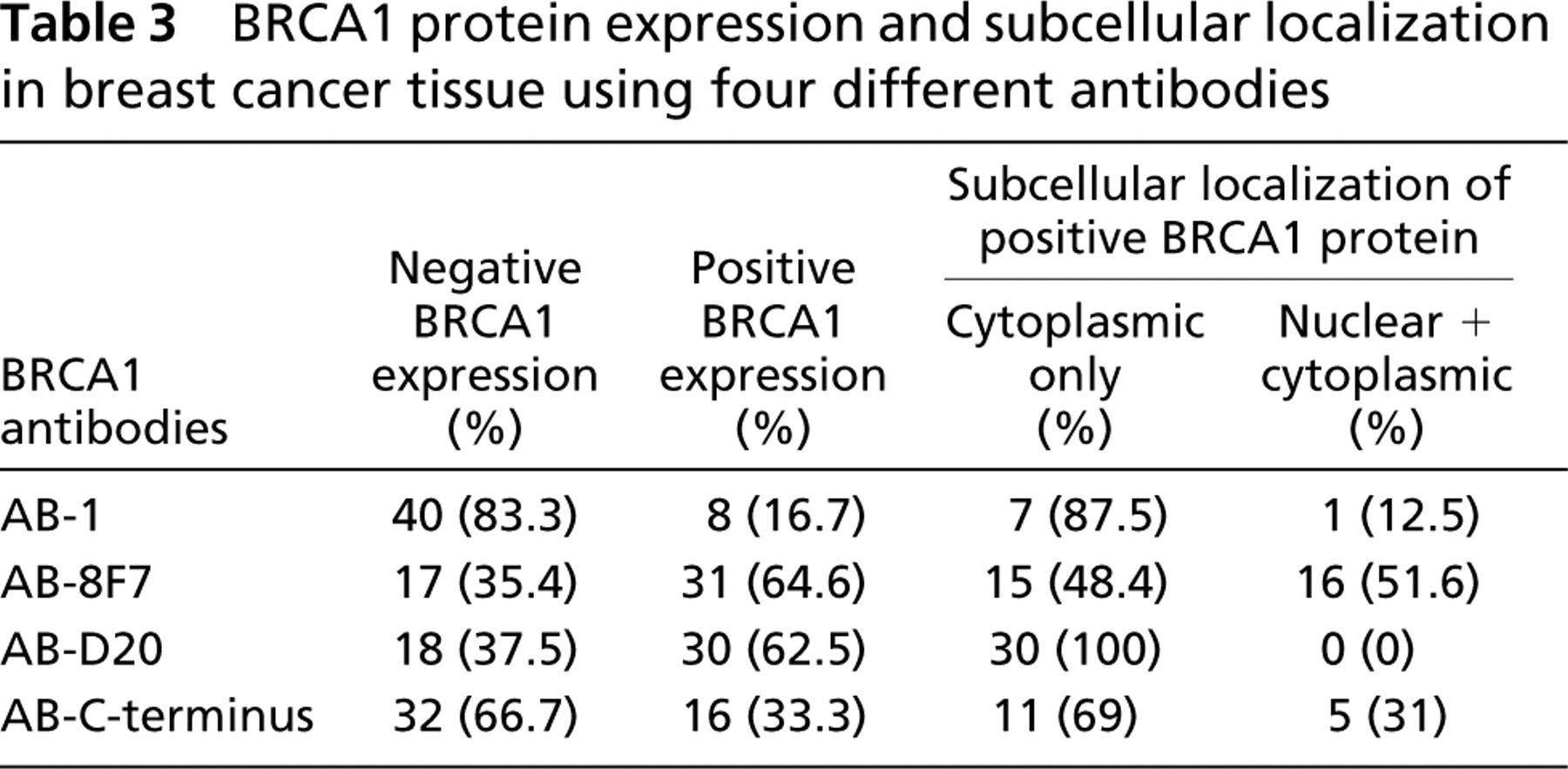
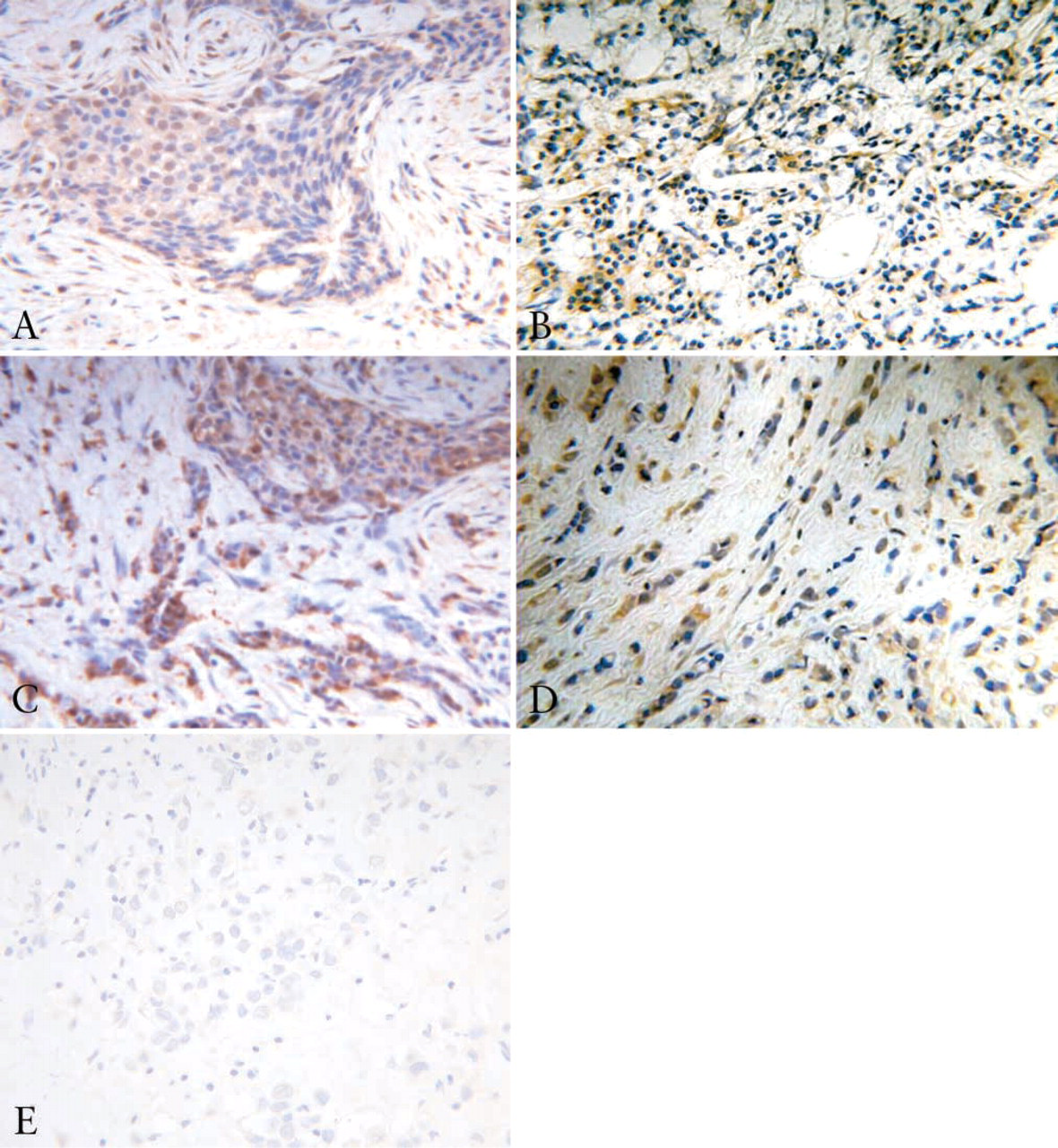
Monoclonal Antibody against N-terminus, AB-1. IHC using monoclonal antibody against N-terminus, AB-1 showed positive staining for BRCA1 protein in only 16.7% of cases. In 87.5% of these cases, staining was localized in the cytoplasm. The remaining 12.5% showed staining in both the cytoplasm and nuclei (Figure 1A).
BRCA1 protein expression and subcellular localization in breast cancer tissue using four different antibodies
BRCA1 IHC using four different antibodies. (
Monoclonal Antibody against Exon-11, AB-8F7. Monoclonal antibody against exon-11, AB-8F7 showed positive BRCA1 protein staining in 64.6% of cases. In half of these cases, staining was nuclear and cytoplasmic. In the remaining half, staining was cytoplasmic only (Figure 1B).
Polyclonal Antibody against N-terminus, AB-D20. BRCA1 protein staining with polyclonal antibody against N-terminus, AB-D20 was positive in 62.5% of cases, and staining was localized in the cytoplasm in all these cases (Figure 1C).
Polyclonal Antibody against C-terminus, AB-C-terminus. Polyclonal antibody against AB-C-terminus showed positive staining for BRCA1 protein in 33.3% of cases. In 69% of these cases, staining was localized only in the cytoplasm. Staining in the remaining 31% was localized in both the nucleus and cytoplasm (Figure 1D).
Cytoplasmic staining appeared to be the most common pattern, with or without nuclear staining, and none of the antibodies demonstrated nuclear staining only (Tables 2 and 3).
Real-time RT-PCR
Given the generally poor quality of RNA extracted from formalin-fixed paraffin-embedded tissues, several steps were implemented to ensure the accuracy of the results. First, only samples that had successful amplification of GAPDH, a housekeeping gene, were considered for subsequent BRCA1 amplifications. This step ensured that samples with only high-quality RNA were considered and represented a control for RNA degradation. Second, to ensure amplification of the less common transcript, namely, BRCA1, the number of amplification cycles was increased to 45. Third, the MCF7 cell line and a paraffin-embedded tissue known to express BRCA1 were used in all reactions as positive controls. Finally, to ensure reproducibility, three independent RT-PCR reactions were performed for each sample. Such stringent criteria reduced our analyzable tissues to 29 of the 48 for which IHC was successful. Of 29 breast cancer tissues analyzed for BRCA1 mRNA expression using real-time RT-PCR, only 6 (21%), showed detectable BRCA1 mRNA. Twenty-three breast cancer tissues (79.3%) showed no BRCA1 mRNA amplification, even after 45 cycles. The amplification curves of RNA extracted from the breast cancer MCF-7 cell line and paraffin-embedded breast cancer tissues using BRCA1 primers are demonstrated in Figure 2.
Correlation between BRCA1 mRNA and Protein Expression
Table 4 compares BRCA1 mRNA expression with corresponding protein expression. BRCA1 mRNA expression in breast cancer tissues showed a significant relationship with protein expression by only one of the antibodies, AB-1 (p = 0.002). Thus, 72% of breast cancers, which also did not demonstrate BRCA1 at mRNA level, did not show BRCA1 protein expression by IHC using AB-1. No significant relationship could be detected between BRCA1 mRNA and BRCA1 protein expression using the remaining three antibodies (AB-8F7, AB-D20, and AB-C-terminus).
Specificity and Sensitivity of the Four Different BRCA1 Antibodies
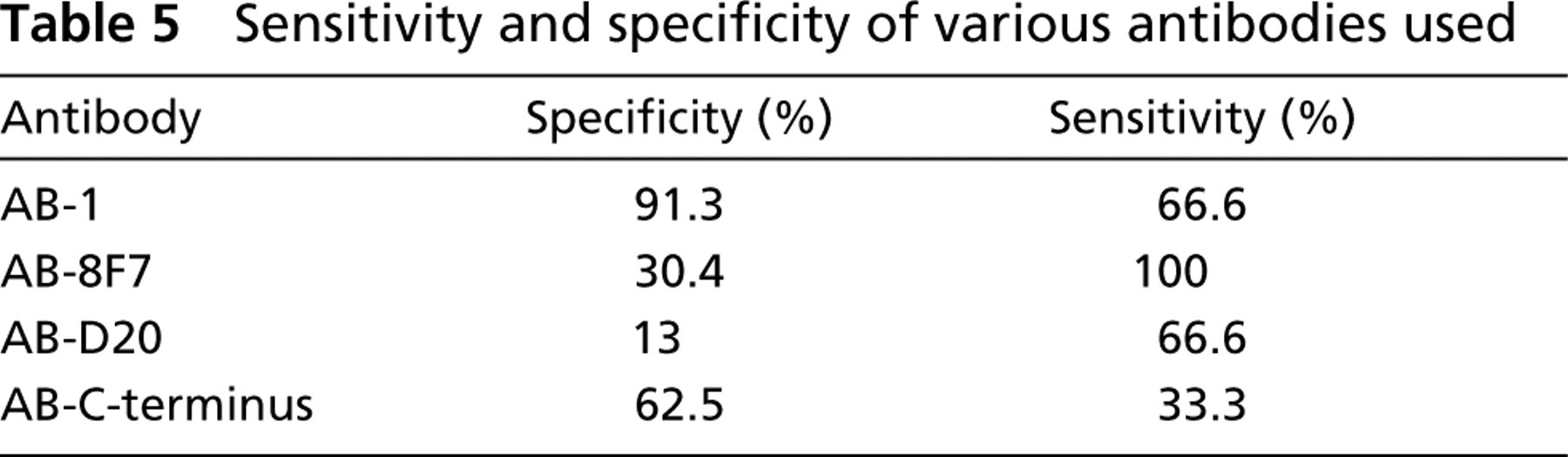
To evaluate the specificity and sensitivity of the four anti-BRCA1 antibodies, the RT-PCR results were considered as standard. AB-1 has the best combined specificity (91.3%) and sensitivity (66.6%) in detecting BRCA1 protein (Table 5). Although AB-8F7 had 100% sensitivity, its specificity was low (30.4%).
Discussion
The present study shows lack of both BRCA1 mRNA and protein expression in the majority of breast cancers studied. This finding is consistent with other reports, which have shown that BRCA1 mRNA and protein are decreased in mammary tumors compared with matched normal breast tissue (Thompson et al. 1995; Rio et al. 1999). The negative BRCA1 mRNA and protein expression in this study was associated with infiltrative ductal carcinoma, although this relationship was not statistically significant. On the other hand, there was a significant, albeit marginal, relationship between BRCA1 mRNA expression and high histological grade of breast cancer (p = 0.05). This finding is consistent with the observation that the majority of high-grade breast cancers show weak or no BRCA1 expression (Jacquemier et al. 1995; Wilson et al. 1999; Lakhani et al. 2000), suggesting that absence of BRCA1 may contribute to the pathogenesis of a significant percentage of breast cancers. Seventy-seven percent of our patients had a high histological grade of breast cancer; although this could indicate a selection bias, this represents a consistent observation in Kuwaiti patients (unpublished data). It follows that our low BRCA1 detection rate, although consistent with the study of Wilson et al. (1999), could be attributed to this selection bias.
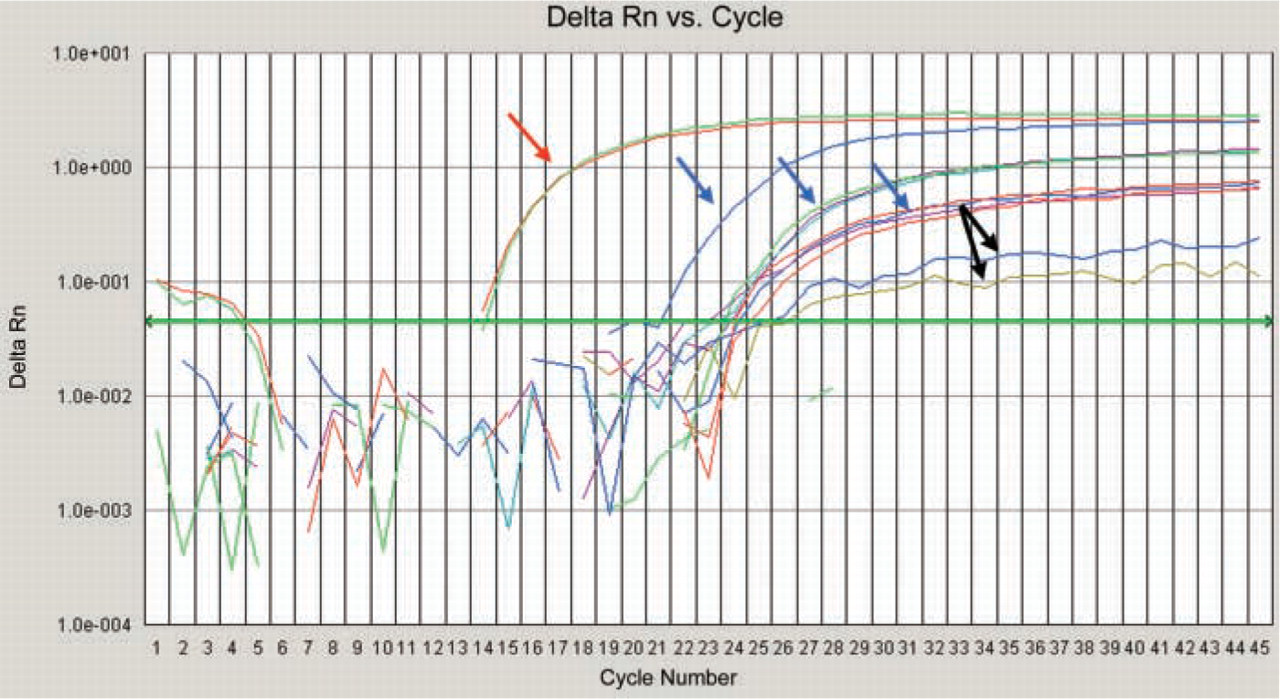
Amplification curves of RNA extracted from breast cancer MCF7 cell line (red arrow) and paraffin-embedded breast cancer tissues from several patients (blue arrows) using BRCA1 primers. The x-axis represents the PCR cycle number at which the cDNA started the amplification; the y-axis represents Delta-Rn, which is the normalized reporter signal minus the baseline signal. Blue arrow corresponds to the BRCA1 mRNA amplification curve of the breast cancer tissues shown in Figure 1. The last two curves are samples considered negative for BRCA1 expression (black arrows).
Comparison between BRCA1 mRNA and protein expression in breast cancer tissues

One of the major tasks of this study was to choose the appropriate and reliable antibody for BRCA1 protein IHC. There is a major controversy about the usefulness and specificity of different BRCA1 antibodies. This controversy is even more pronounced when paraffin-embedded tissues are used. In the present study, four anti-BRCA1 antibodies against different BRCA1 epitopes were used. The results obtained were compared to determine which antibody is most reliable for detecting BRCA1 protein on paraffin-embedded breast cancers. To evaluate the specificity and sensitivity of the four anti-BRCA1 antibodies, the RT-PCR results were considered as a standard. Our results demonstrate a significant relationship between BRCA1 mRNA and BRCA1 protein expression with only one of the antibodies, monoclonal AB-1. Almost all breast cancers that had no or significantly reduced BRCA1 mRNA expression in this study were also negative for BRCA1 protein expression with AB-1 (21/23, 91%; p = 0.002). Accordingly, AB-1 has the best combined specificity (91.3%) and sensitivity (66.6%) in detecting BRCA1 protein (Table 5). In a study using four commercially available anti-BRCA1 antibodies on paraffin-embedded breast cancers (including AB-1 and AB-D20), Perez-Valles et al. (2001) reported that available BRCA1 antibodies lack the specificity required to identify the BRCA1 protein. Others, however, have used AB-1 for immunolocalization of BRCA1 on breast cancer and have reported that this antibody is the most reliable antibody for detecting BRCA1 protein (Lee et al. 1999; Wilson et al. 1999; Niwa et al. 2000). Our data are consistent with this finding. IHC results using the other three antibodies have shown no statistically significant relationship with BRCA1 mRNA. Of all the antibodies, AB-8F7 had the best sensitivity, but its specificity was low (30.4%). Results reported by Yoshikawa et al. (1999) suggest that N-terminus AB-1 could be useful in prescreening tumors for BRCA1 mutations because of a high detection rate (7 of 19, 37%) of alterations in the BRCA1 gene product in breast cancer. Our data are also consistent with the findings that BRCA1 loss is at mRNA level (Russell et al. 2000; Baldassarre et al. 2003). Interestingly, we have found that some of the breast cancers with positive BRCA1 mRNA expression had no BRCA1 protein expression with AB-1. This finding supports the hypothesis that there might be other mechanisms by which BRCA1 expression is controlled at translational or posttranslational levels (Miyamoto et al. 2002; Sobczak and Krzyzosiak 2002). Nevertheless, loss of both mRNA and BRCA1 protein could indicate that the genetic control of BRCA1 expression in the breast cancer analyzed is at transcriptional level.
Sensitivity and specificity of various antibodies used
The subcellular localization of BRCA1 has been also been controversial. BRCA1 has been claimed to be an exclusively nuclear protein in both normal and cancer cells (Scully et al. 1996; Thomas et al. 1996; Thakur et al. 1997; Wilson et al. 1999), a nuclear protein in normal cells but an aberrantly localized cytoplasmic protein in breast and ovarian tumor cells (Chen et al. 1995; Lee et al. 1999), and a cytoplasmic protein found in tubelike structures that invaginate the nucleus (Coene et al. 1997). The variation in the subcellular localization of BRCA1 might be attributable to several causes, including the specificity of the antibodies used to localize the protein, antibody cross-reactivity (Smith et al. 1996; Bernard-Gallon et al. 1997; Wilson et al. 1999), and the presence of splice variant isoforms (Thakur et al. 1997; Wilson et al. 1997). The present study confirms that BRCA1 is expressed as both nuclear and cytoplasmic antigen in breast cancer tissues. Cytoplasmic staining was a consistent feature in the breast cancers positive for BRCA1, but the nuclear expression of BRCA1 protein ranged from 37% to 83% of breast cancers (depending on the type of antibody used) and was absent in a subset of them. AB-1 monoclonal N-terminus antibody, which showed a nuclear dot pattern both in normal and cancer cell lines in other studies (Scully et al. 1996,1997; Wilson et al. 1999; Hsu et al. 2001), resulted in only 16.7% positive staining for BRCA1. In 87.5% of these cases, staining was located in the cytoplasm only; in the remaining 12.5%, staining was in both the cytoplasm and nuclei. Differences in subcellular localization of the BRCA1 protein can affect the interpretation of the IHC results (Chen et al. 1995). Wilson et al. (1997) compared 19 BRCA1 antibodies and detected nuclear foci in both a normal breast epithelial cell line and a breast malignancy–derived cell line. In accord with Wilson's study are the studies of Thomas et al. (1996), Ruffner and Verma (1997), and Zhang et al. (1997), which have also reported the nuclear subcellular localization of BRCA1 protein on cell lines. On the other hand, Chen et al. (1995) have reported that BRCA1 is a nuclear protein in normal cells and becomes aberrantly localized in cytoplasm of breast and ovarian tumor cells. In the present study, none of the breast cancer tissues showed BRCA1 nuclear localization alone. It can be argued that breast cancer cell lines, not archival paraffin-embedded breast cancer tissues, were used in the majority of these studies. Subcellular localization of proteins in cell lines cannot be compared with formalin-fixed, paraffin-embedded tissue, because several factors affect the protein localization in the tissue. The staining pattern in the paraffin-embedded tissues could be altered by changes in tissue fixation conditions and antigen retrieval methods; differences in immunostaining methodology, antibody concentrations, specificity of the antibodies used to localize the protein, and antibody cross-reactions (Scully et al. 1996; Smith et al. 1996; Bernard-Gallon et al. 1997; Wilson et al. 1997,1999); the presence of splice variant isoforms (Lu et al. 1996; Wang et al. 1997; Orban and Olah 2001; Fabbro et al. 2002); and the problems of working with preserved archival tissue (Scully et al. 1996; Thomas et al. 1996; Bernard-Gallon et al. 1997; Wilson et al. 1999). According to Yoshikawa et al. (1999), results obtained by using C-terminus antibody should be evaluated carefully because of false-positive immunostaining demonstrated. BRCA1 protein localization is a complex issue, but the antibodies used in this study were chosen against different BRCA1 protein epitopes (N-terminus, exon 11, and C-terminus), which enabled us to recognize different protein isoforms. In addition, the realtime probe used for BRCA1 detects all known mRNA variants (because it hybridizes to the junction of exon 22–exon 23).
In conclusion, the clinical benefits of establishing BRCA1 expression status and its effects on breast cancer treatment, prophylaxis, and prognosis are obvious (Lafarge et al. 2001; Egawa et al. 2003; Zhou et al. 2003). Thus, IHC can be a valuable preliminary test for detecting the reduction in BRCA1 protein expression. Among the different available BRCA1 antibodies, we consider AB-1 to be the best anti-BRCA1 antibody that can be applied on formalin-fixed paraffin-embedded tissues, with 91.3% specificity and 66.6% sensitivity. Nevertheless, AB-8F7 detected six of six tumors positive for BRCA1 mRNA, making it highly sensitive. The antibody, however, also produced signals in another 16 tumors that were negative for BRCA1 mRNA, which indicates low specificity if one considers, as we did, RT-PCR to be the reference standard. We acknowledge that the small number of positive RT-PCR values constitutes a limitation of the study. A further, larger study focusing on identifying various BRCA1 splice variants and their correlation with a range of antibodies raised against a larger number of epitopes is now warranted.
Footnotes
Acknowledgements
This study was funded by the College of Graduate Studies of Kuwait University and the Kuwait Institute for the Advancement of Sciences, project number 990707 (FA-M), and Shared Facility Grant, number GM/0101.
We wish to thank Dr. Josley George, Dr. Shirley George, Mrs. Bency John, and Mrs. Tessy Saji for their technical support.