
Abstract
Ezrin is a membrane-cytoskeletal linker belonging to the ezrin-radixin-moesin (ERM) family and has been suggested to be involved in tumorigenesis. In this study we investigated ezrin expression pattern in normal esophageal mucosa and esophageal squamous cell carcinoma (ESCC) and the correlation with clinical characteristics. Immunohistochemical staining showed a tendency for ezrin to translocate from membrane to cytoplasm in the progression from normal epithelium to invasive carcinoma of the esophagus. By Western blot, we found that ezrin expression was downregulated in 13 ESCC specimens and upregulated in 36 others. Moreover, quantitative real-time RT-PCR demonstrated that ezrin mRNA level in normal esophageal mucosa was 3.60 ± 3.60 times that in ESCC (p<0.001). Proliferating cell nuclear antigen (PCNA) expression level was higher in ezrin downregulated group compared with that in ezrin upregulated group (p<0.05). However, there was no significant association between ezrin expression and clinical characteristics. The results suggested that the localization of ezrin by immunohistochemistry may be useful in the diagnosis of ESCC, and ezrin may play a suppressive role in the tumorgenesis of ESCC.
Keywords
T
Ezrin usually concentrates at actin-rich surface structures such as the apical domain of polarized cells (Bretscher et al. 1997). Certain circumstances can stimulate the translocation of ezrin, and there is evidence suggesting that the subcellular redistribution of ezrin significantly correlates with tumorigenesis (Tokunou et al. 2000; Moilanen et al. 2003).
Except for the redistribution, the expression level of ezrin was also suggested to be associated with tumorigenesis as well as motility and invasive behavior of malignant cells. There are two different opinions about the role of ezrin in tumor progression. One opinion supported that ezrin was a suppressor with ezrin down-regulated in some tumor cells (Tokunou et al. 2000; Guan et al. 2002; Moilanen et al. 2003) and inhibition of ezrin expression leading to a reduced cell-cell adhesiveness together with a gain in their motile and invasive behavior (Hiscox and Jiang 1999). The other opinion regarded ezrin as an enhancing factor with ezrin exhibiting strong expression in a variety of invasive human cancers and being actively involved in regulating the growth and metastatic capacity of cancer cells (Geiger et al. 2000; Tynninen et al. 2004; Elliott et al. 2005; Ilmonen et al. 2005; Weng et al. 2005).
Our group previously proved ezrin was overexpressed in malignantly transformed esophageal epithelial cell line compared with immortalized phase cell line (Shen et al. 2003). Until now, however, there have been few reports about how ezrin was expressed in esophageal squamous cell carcinoma (ESCC) tissues and what its relation was with the clinical and pathological parameters such as lymph node status, histopathological grades, and clinical grades. Thus, based on our previous study of ezrin on esophageal epithelial cell lines, we continued our work in an attempt to identify ezrin expression pattern and its clinical significance in esophageal epithelial tissues.
Materials and Methods
Patients and Tissue Specimens
Eighty nine paraffin sections including 40 ESCC, 42 normal esophageal mucosa, and 7 special paraffin sections of full-length mucosa layer from the distant margin to the cancer focus of the excised esophagus were obtained for immunohistochemical staining from the Clinical Pathology Department of the First Affiliated Hospital of Shantou University from 2001 to 2003. The sections of normal mucosa, which had been histologically evaluated by hematoxylin and eosin staining, were taken from the matched distal resected margin of ESCC samples. All specimens were fixed in 10% formaldehyde solution, embedded in paraffin blocks, and then cut into 4-μm-thick sections.
Frozen ESCC tissues and adjacent normal esophageal mucosa tissues (taken as far as possible from neoplasia) were collected for Western blot and real-time RT-PCR from 49 patients with primary ESCC who underwent surgery in the Department of Cardiothoracic Surgery of the First Affiliated Hospital of Shantou University from 2003-2004. All samples were immediately frozen in liquid nitrogen following surgery and stored at −70C until RNA isolation.
All tumors were confirmed as ESCC by the Clinical Pathology Department of the Hospital. All cases were classified according to the International Union Against Cancer Tumor-Node-Metastasis Classification, 5th ed. Evaluation of tumor differentiation was based on histological criteria of the guidelines of WHO Pathological Classification of Tumors. The study was approved by the Ethics Committee of the First Affiliated Hospital of Shantou University, and only patients who gave written informed consent were included. None of the patients had radiotherapy or chemotherapy prior to surgery.
Immunohistochemical Staining
Immunohistochemical staining was done using a modified streptavidin-peroxidase conjugated method based on the poly-horseradish peroxidase (HRP) anti-mouse/rabbit IgG detection system of the Polymer Detection System (Golden Bridge International, Inc.; Mukilteo, WA) Briefly, each tissue section was deparaffinized in dimethylbenzene, rehydrated through a graded ethanol series, and incubated with fresh 3% hydrogen peroxide for 10 min. After rinsing in PBS, antigen retrieval from the tissue was carried out by autoclaving in 0.01 M sodium citrate buffer (pH 6.0) at 120C for 3 min. Next, sections were blocked in 10% normal goat serum for 15 min at room temperature (RT) and then incubated overnight at 4C with mouse anti-human ezrin monoclonal antibody (1:800 dilution; Lab Vision Corporation, Fremont, CA). After PBS rinse, slides were incubated for 15 min at RT with polymer helper solution (Polymer Detection System Kit; Golden Bridge International) and then rinsed in PBS. This was followed by 20-min incubation at RT with streptavidin-peroxidase-conjugated anti-mouse IgG (Polymer Detection System Kit, Golden Bridge International). Subsequently, they were stained with 0.003% 3,3-diaminobenzide tetrahydrochloride and 0.005% hydrogen peroxide in 0.05 M Tris-HCl (pH 7.2), counterstained with Mayer's hematoxylin, dehydrated, and then mounted.
As negative controls, one was prepared by substituting PBS for the primary antibody, and the other was applied using preimmune serum (1:1000). A metastatic breast carcinoma shown previously to have immunoreactivity was used as a positive control to confirm immunoreactivity in each series of experiments.
Ezrin immunoreactivity was evaluated. Either membrane or cytoplasmic staining was considered positive. In each case, the intensity of membrane or cytoplasmic staining was scored as follows: no visible staining or appreciable staining in <5% of cells, 0; faint staining detected in >5% of cells, 1+; weak to moderate staining in >5% of cells, 2+; strong staining in >5% of cells, 3+. Cases classified as 0 were considered “negative,” and those scored as 1+, 2+, or 3+ were considered “positive.” All sections were evaluated independently by two investigators who had no prior knowledge of the patients' clinical information. When the opinions of the two evaluators were different, agreement was reached by careful discussion.
Western Blot Analysis
Tissues were lysed in a sample buffer (50 mM Tris-HCl, pH 8.0,150 mM NaCl, 1% Triton X-100, 100 μg/ml PMSF) on ice for 30 min. Lysates were then centrifuged for 5 min (12,000 × g, 4C). Protein concentration was estimated by the Bradford method. An equal amount of tissues lysates (50 μg) was electrophoresed on 10% polyacrylamide gel and transferred to polyvinylidene difluoride membranes (Millipore; Bedford, MA). The membranes were then blocked with 5% skim milk-PBST (0.01 M PBS, pH 7.4, 0.05% Tween 20) for 1 hr and incubated at RT for 1 hr with mouse anti-human ezrin monoclonal antibody (Lab Vision Corporation). The membrane was subsequently incubated at RT for 1 hr with HRP-linked goat anti-mouse IgG and analyzed using Western blotting luminol reagent (Santa Cruz Biotechnology; Santa Cruz, CA). Image acquisition and quantitative analysis were carried out with the FluorChem 8900 image analysis system (Alpha Innotech; Miami, FL). To evaluate the approximate proliferation level of cells in every sample, PCNA level was measured with anti-PCNA monoclonal antibody (DAKO; Glostrup, Denmark). To verify the relative amounts of protein in each lane, β-actin level as an internal control was measured with anti-β-actin monoclonal antibody (Sigma; St Louis, MO). Protein expression level in tumors and in the adjacent normal tissues were calculated from the signal intensity. Expression levels of ezrin and PCNA were finally evaluated by the signal intensity ratio of their presence in the tumor to the adjacent normal epithelial tissue (T/N ratio).
Preparation of Total RNA and Real-time RT-PCR
Total RNA was extracted from frozen stored tissues with TRIzol reagent (Invitrogen; Carlsbad, CA) in accordance with the manufacturer's instructions. Reverse transcription was performed in a total volume of 20 μl using 1 μg of total RNA, oligo-dT primer, random primers, and AMV Reverse Transcriptase (Promega; Madison, WI). All PCR reactions were performed using an ABI Prism 7000 Sequence Detection System (Perkin-Elmer Applied Biosystems; Foster City, CA) based on the TaqMan technology. For normalization of our results, β-actin was used as the internal control. Primers and probes for ezrin and β-actin were chosen with the assistance of Primer Express 2.0 software (Perkin-Elmer Applied Biosystems). To avoid amplification of contaminating genomic DNA, one of the two primers of each gene was intron spanning. PCR primers used for ezrin were as follows: forward primer 5î-GCTTTTTGATCAGGTGGTAAAGACT-3î and reverse primer 5î-TCCACATAGTGGAGGCCAAAGT-3î. According to the gene sequence of ezrin, a PCR product of 69 bp was obtained. For β-actin, the forward primer was 5î-GCGCGGCTACAGCTTCA-3î and the reverse primer was 5î-TCTCCTTAATGTCACGCACGAT-3î. The sequence of the labeled probe was as follows: 5î- TCGGCCTCCGGGAAGTGTGG-3î for ezrin and 5î- CACCACGGCCGAGCGGGA-3î for β-actin. The 50-μl volume PCR reaction mixture was preheated at 93C for 3 min, followed by 40 cycles at 93C for 45 sec and 55C for 1 min. A non-template control and the standard curve group with 10-fold serial dilutions of pGEM-T Easy Vector (Promega) containing truncated ezrin cDNA as templates were included in each PCR run.
Statistical Analysis
In immunohistochemistry study, relationship between ezrin-positive rate and cell differentiation was analyzed using X 2 test. In Western blot analysis, the relationships among ezrin and PCNA expression levels and clinical characteristics were analyzed by using independent sample t-test. In the investigation of real-time RT-PCR, the extent of ezrin mRNA expression was analyzed by using paired sample t-test. Significance level was set at p<0.05.
Results
Translocalization of Ezrin in the Progression of ESCC
A significant difference of ezrin distribution was observed in normal esophageal mucosa and ESCC by antibody staining. Ezrin immunoreactivity was apparent in the cell membranes of 41/42 (97.6%) normal esophageal mucosa cases (Figures 1A and 1B). In contrast, ESCC samples showed mainly cytoplasmic staining in 31/40 (77.5%) or both membranous and cytoplasmic staining in 9/40 (22.5%). Of the 40 cases of ESCC, 9 were well differentiated and 31 were moderately differentiated, and ezrin was found to be mainly diffused in the cytoplasm in 29/31 (93.5%) moderately differentiated samples (Figure 1D), whereas it was preferentially localized both in the membranous region and in the cytoplasm in 7/9 (77.8%) well-differentiated ESCC (Figure 1C). A significant difference of ezrin distribution was found between well- and moderately differentiated groups (p< 0.05). Results of these immunohistochemical examinations are summarized in Table 1.
We also investigated ezrin distribution in the progression from normal epithelium to invasive carcinoma of the esophagus using seven special paraffin sections of full-length mucosa layer from the distant margin to the cancer focus of the excised esophagus. In addition to the normal epithelium (7/7) and invasive cancer of esophagus (7/7), these seven special paraffin sections contained esophageal lesions of simple hyperplasia (3/7), low-grade dysplasia (1/7), and carcinoma in situ (3/7). In normal epithelium, positive immunostaining was mainly apparent on the membrane in all cases (Figures 2A and 2B). For simple hyperplasia (2/3), positive signal was predominantly observed in both membrane and the cytoplasm of the basal layer and the spinous layer cells (Figure 2C). In low-grade dysplastic lesions and all carcinoma in situ and invasive cancers, most cells demonstrated only diffusely cytoplasmic staining (Figures 2D-2F).
Ezrin Protein Level in ESCC and the Adjacent Normal Mucosa
We performed Western blot to identify the expression level of ezrin in esophageal tissues by using the frozen stored ESCC and the adjacent normal epithelial tissues. The intensity ratio of ezrin to β-actin was scored. Each specimen was evaluated by the ratio of the ezrin protein level in the tumor to that in the adjacent normal epithelial tissues (T/N ratio). Ezrin expression was upregulated (T/N ratio >1.0) in 13 (26.5%) cases and downregulated (T/N ratio <1.0) in 36 (73.5%) cases (Figure 3 and Figure 4).
Ezrin mRNA Level in ESCC and the Adjacent Normal Mucosa
With real-time RT-PCR, the relative copy number of ezrin mRNA was evaluated by the ratio of the absolute copy number of ezrin mRNA to the copy number of β-actin mRNA. The result demonstrated that ezrin mRNA level in normal esophageal mucosa was 3.60 ± 3.60 times that in ESCC (p<0.001).
Correlation between Level of Ezrin Protein and Cell Proliferation
Overexpression of PCNA, a biomarker for cell proliferation (Kimos et al. 2004), was observed in 87.76% (43/49) of the tumor tissues compared with the adjacent normal epithelium (Figure 3). PCNA expression level was higher in ezrin downregulated group (10.06 ± 9.74, mean ± SD) than in ezrin upregulated group (5.01 ± 4.47, mean ± SD) (p<0.05) (Table 2). These results demonstrated that the expression level of ezrin correlated inversely with cell proliferative potency (Figure 4).
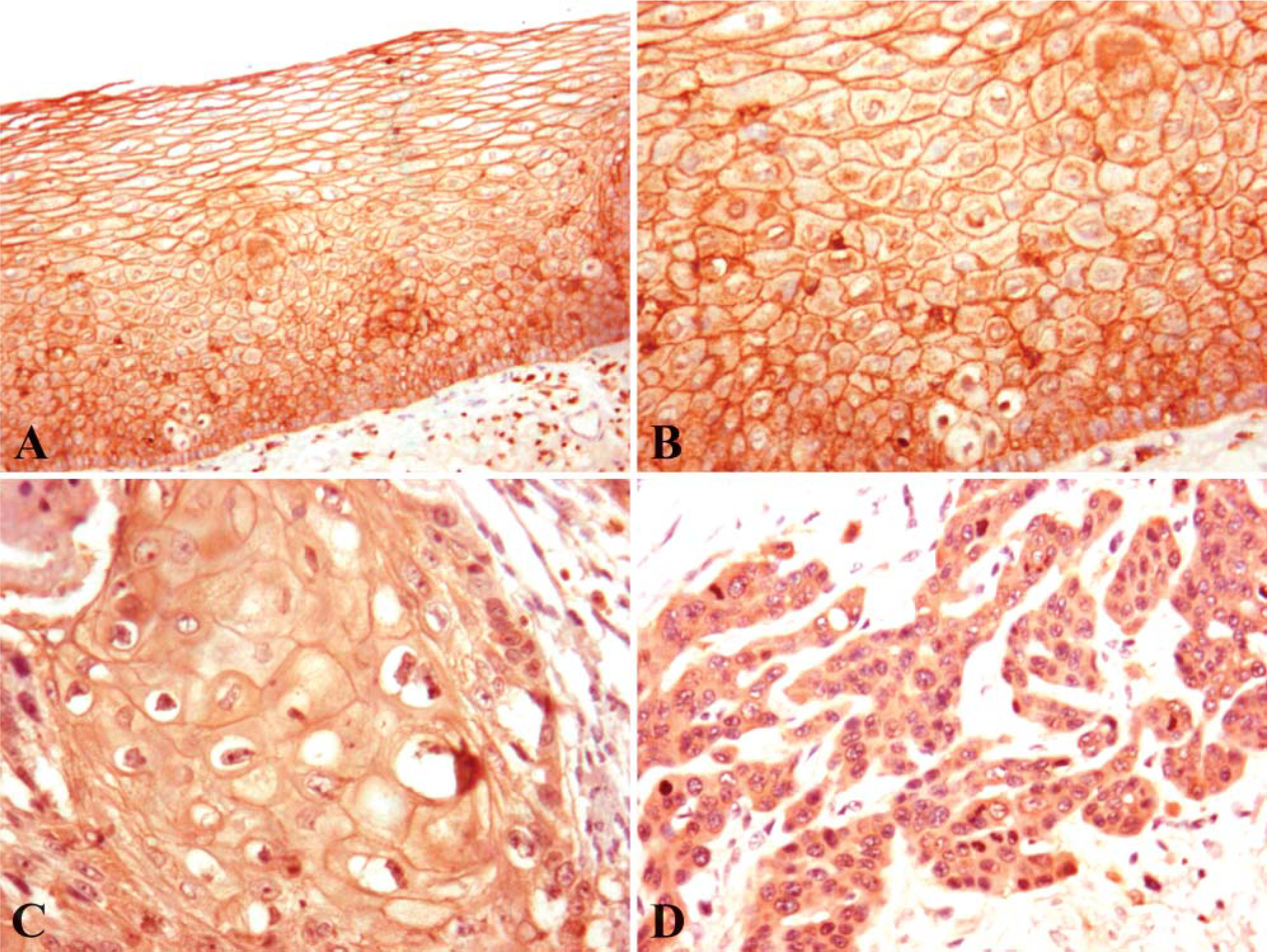
Immunohistochemical staining of ezrin expression in human normal esophageal mucosa and esophageal squamous cell carcinoma (ESCC) tissues. Paraffin-embedded tissue sections were stained by a modified streptavidin-peroxidase conjugated method, as described in Materials and Methods. In normal esophageal epithelium, ezrin immunoreactivity was apparent in the membrane (
Correlation between Clinical Characteristics and Level of Ezrin Protein in ESCC
Correlation between ezrin protein expression and clinical characteristics such as pathologic grades, tumornode-metastasis, and stages was simultaneously investigated (Table 3). However, there was no significant association between ezrin expression and any of the clinical characteristics.
Distribution of ezrin in esophageal squamous cell carcinoma (ESCC) and the adjacent normal esophageal mucosa

Membrane staining.
Cytoplasmic staining.
Positive staining both in membrane and the cytoplasm.
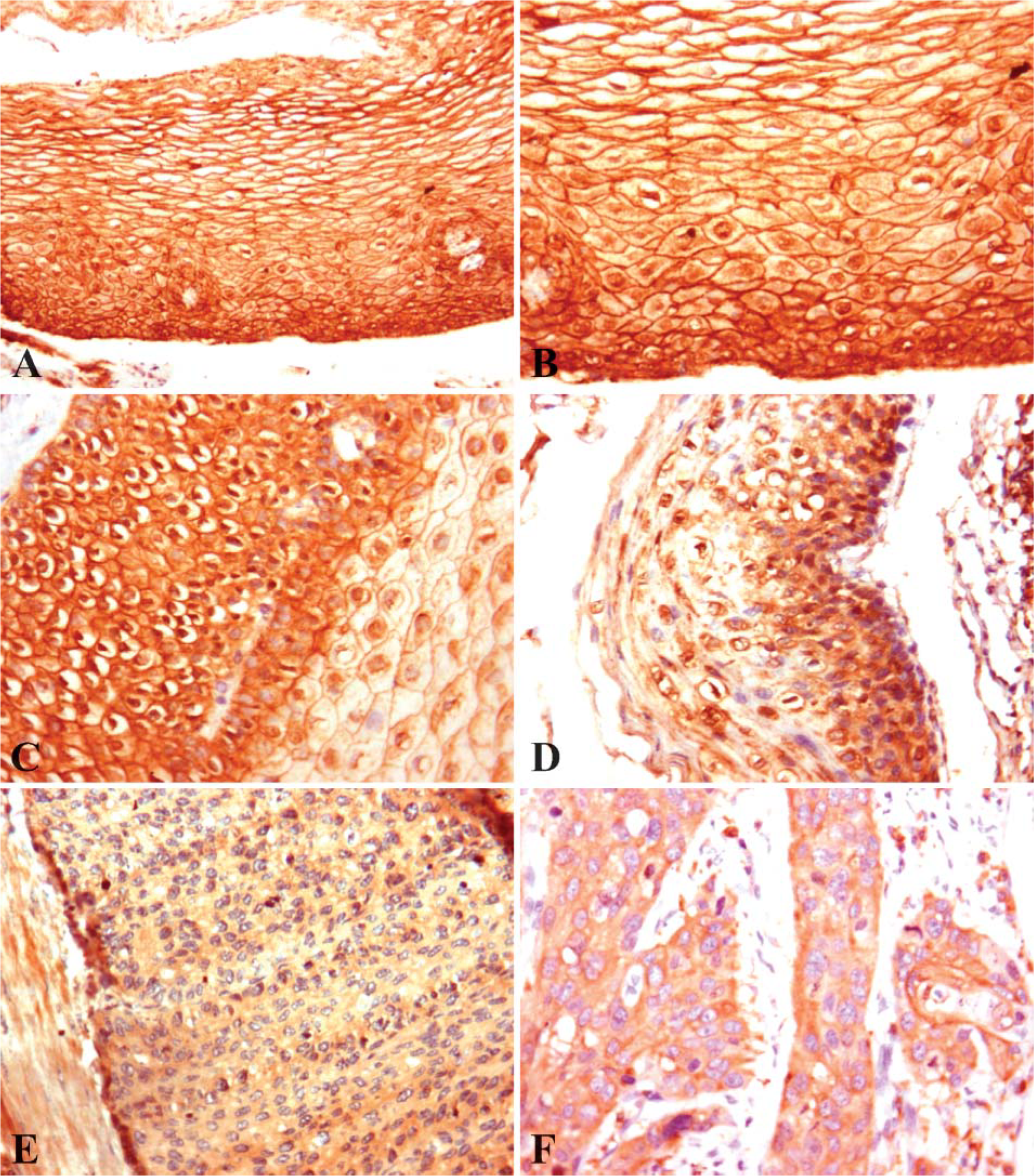
Ezrin distribution in the progression from normal epithelium to invasive carcinoma of esophagus. Seven special paraffin sections of full-length mucosa layer from the distant margin to the cancer focus of the excised esophagus were immunohistochemically investigated as described in Materials and Methods. The normal epithelium mainly showed a membranous staining pattern (
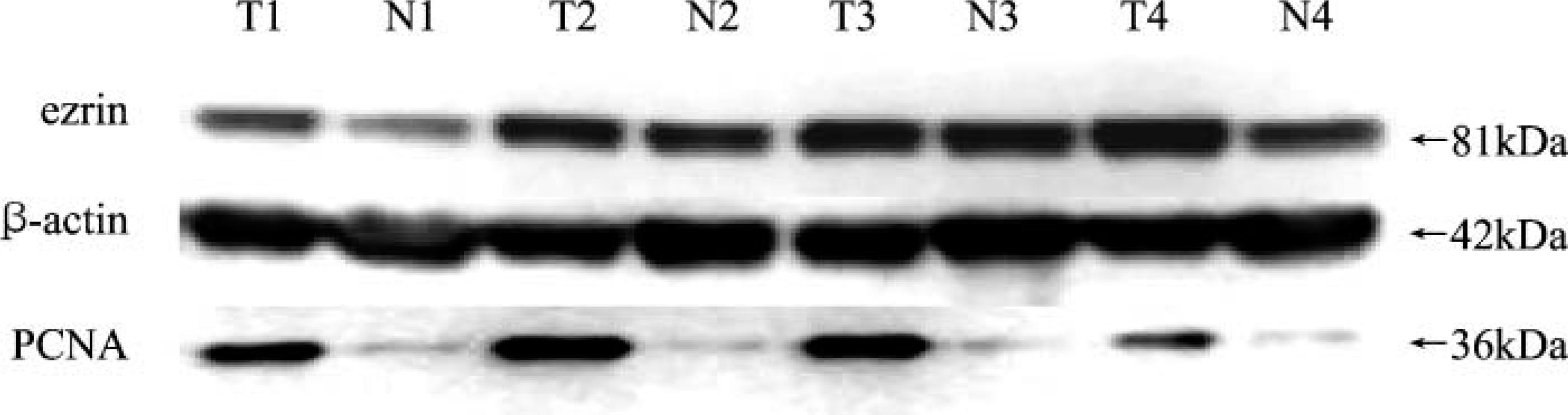
Western blot analysis of ezrin and proliferating cell nuclear antigen (PCNA) levels in four representative cases of ESCC tissues (T) and the adjacent normal esophageal mucosa tissues (N). The molecular weights(kDa) of the immunoreacitivity are shown on the right. Expression of β-actin served as an internal control for equal loading.
Discussion
In this report we showed the differences of ezrin expression at transcriptional and translational levels and ezrin subcellular localization in the progression from normal epithelium to invasive carcinoma of the esophagus. We also demonstrated that ezrin expression level inversely correlated with cell proliferation. From immunohistochemistry data, we found that ezrin had a tendency to translocate from membrane to cytoplasm in the malignant transformation progress of esophageal epithelium. In comparison to normal esophageal epithelium, which demonstrated a mainly membranous staining pattern, ezrin distribution in simple hyperplasia lesions extended to the cytoplasm. In case of low-grade dysplasia, carcinoma in situ, and invasive cancer, ezrin was prominently cytoplasmic diffuse. Furthermore, in most tumor cells, ezrin protein and mRNA levels were reduced.
According to previous studies of our group, ezrin was upregulated in the malignantly transformed esophageal epithelial cell line (Shen et al. 2003), which was contrary to our present finding. This paradox may be due to the different survivable microenvironments between the cell lines and the cells in tissue specimens. The cell lines employed in previous studies of our group originated from the progressive induction of human embryonic esophageal epithelium by the E6E7 genes of human papillomavirus (HPV) type 18 (Shen et al. 2002), and the inductor might be the main regulatory factor modulating the expression of genes including ezrin in the malignantly transforming cell lines. Nevertheless, as for the tumorigenesis of ESCC in the human body, there may be many factors, perhaps including HPV, in the intricate in vivo environment affecting ezrin expression. It would be interesting to see what factors account for the aberrant expression pattern of ezrin in human ESCC.
Ezrin has been reported to be concentrated in actin-rich surface structures such as microvilli, membrane ruffles, and protrusions and can regulate the structure of the cortical cytoskeleton to control cell surface topography with subcellular redistribution (Bretscher et al. 1997). Tokunou et al. (2000) showed the altered expression of ezrin protein in lung adenocarcinoma by immunohistochemical analysis: ezrin was localized in the apical surface of non-neoplastic bronchiolar and alveolar epithelial cells, whereas tumor cells with disorganized structures were either negative or diffusely positive for ezrin in the cytoplasm. Moilanen et al. (2003) also described the subcellular redistribution of ezrin in serous ovarian carcinoma. Similarly, we found an obvious translocation of ezrin from membrane to the cytoplasm during the development of ESCC. The above observations imply that in the transformation and development of tumors, ezrin not only acts as a membrane-cytoskeletal linker to strengthen the cells' ability to retain their normal growth pattern but also is involved in cytoskeleton reorganization (Tokunou et al. 2000). Huang et al. (2003) reported that ezrin localization, which was regulated by Foxj1, was related to cell differentiation in airway epithelial cells. Likewise, a significant difference of ezrin distribution between well- and moderately differentiated ESCC was observed in our present study. The evidence suggests that subcellular translocation of ezrin plays an important role during cell differentiation. However, the assumptions require further investigation. Mechanism of ezrin redistribution in ESCC may be associated with the conformational changes of ezrin protein. Ezrin can exist in an active or dormant state in cells. The active form is involved in interactions with the membrane molecules, whereas the dormant form is present in the cytoplasm (Louvet-Vallée 2000). According to that, we can deduce from our observation that ezrin may predominantly exist in an active state in normal esophageal mucosa epithelium, whereas concomitant actin cytoskeleton breaks down in ESCC cells, mainly in a dormant state.
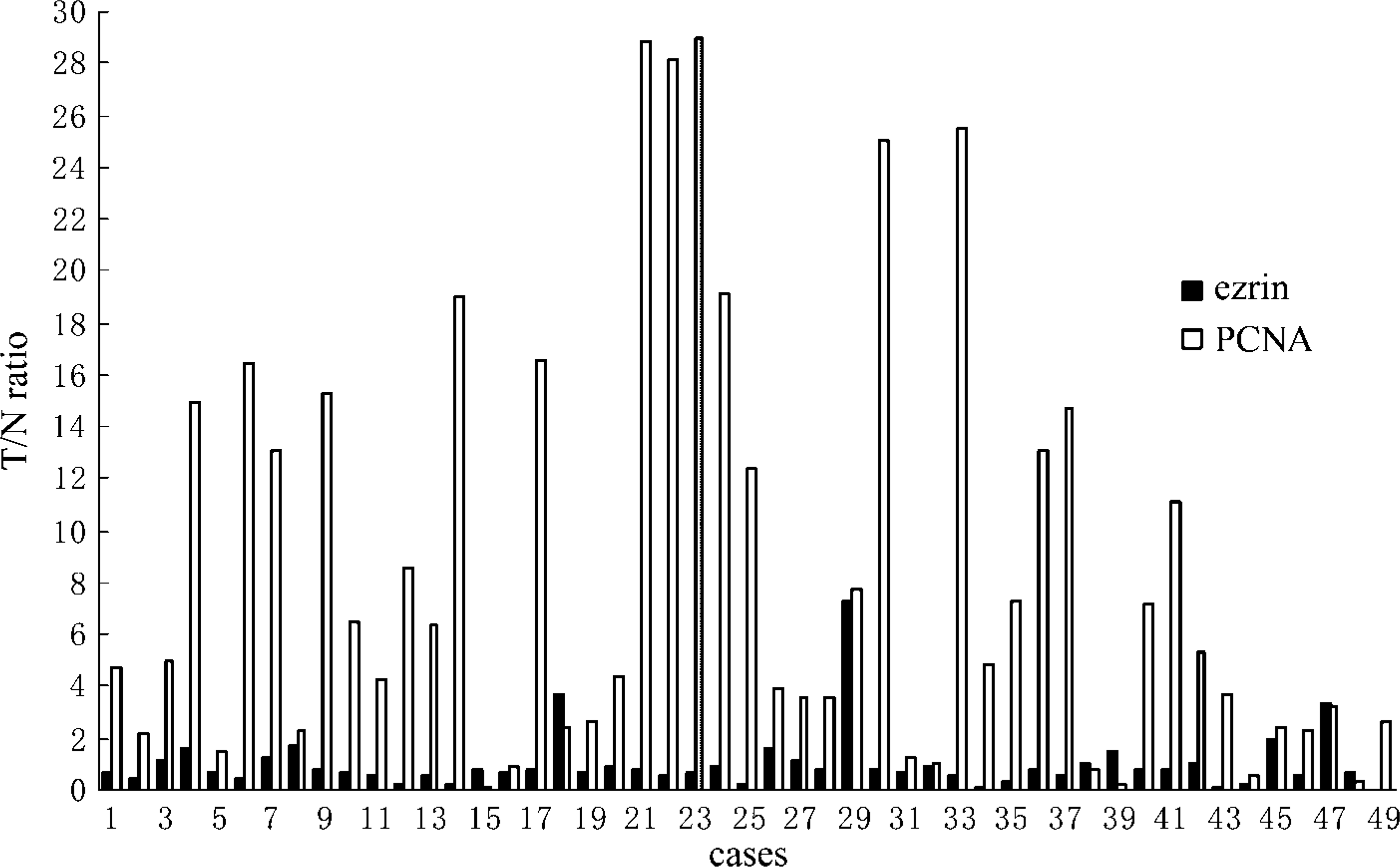
Relative expression levels of ezrin and PCNA in the 49 paired cases analyzed by Western blot. β-actin served as an internal control, and levels of ezrin and PCNA expression were evaluated by T/N ratio as described in Materials and Methods. Distribution of ezrin protein expression among the 49 patients was as follows: 13 cases had T/N ratio >1.0, 36 cases had <1.0. PCNA expression level was higher in the group of ezrin T/N ratio <1.0 than in the group of ezrin T/N ratio >1.0.
Relationship between the expression levels of ezrin and PCNA

PCNA, proliferating cell nuclear antigen.
Relationship between clinical characteristics and ezrin expression level in Western blot
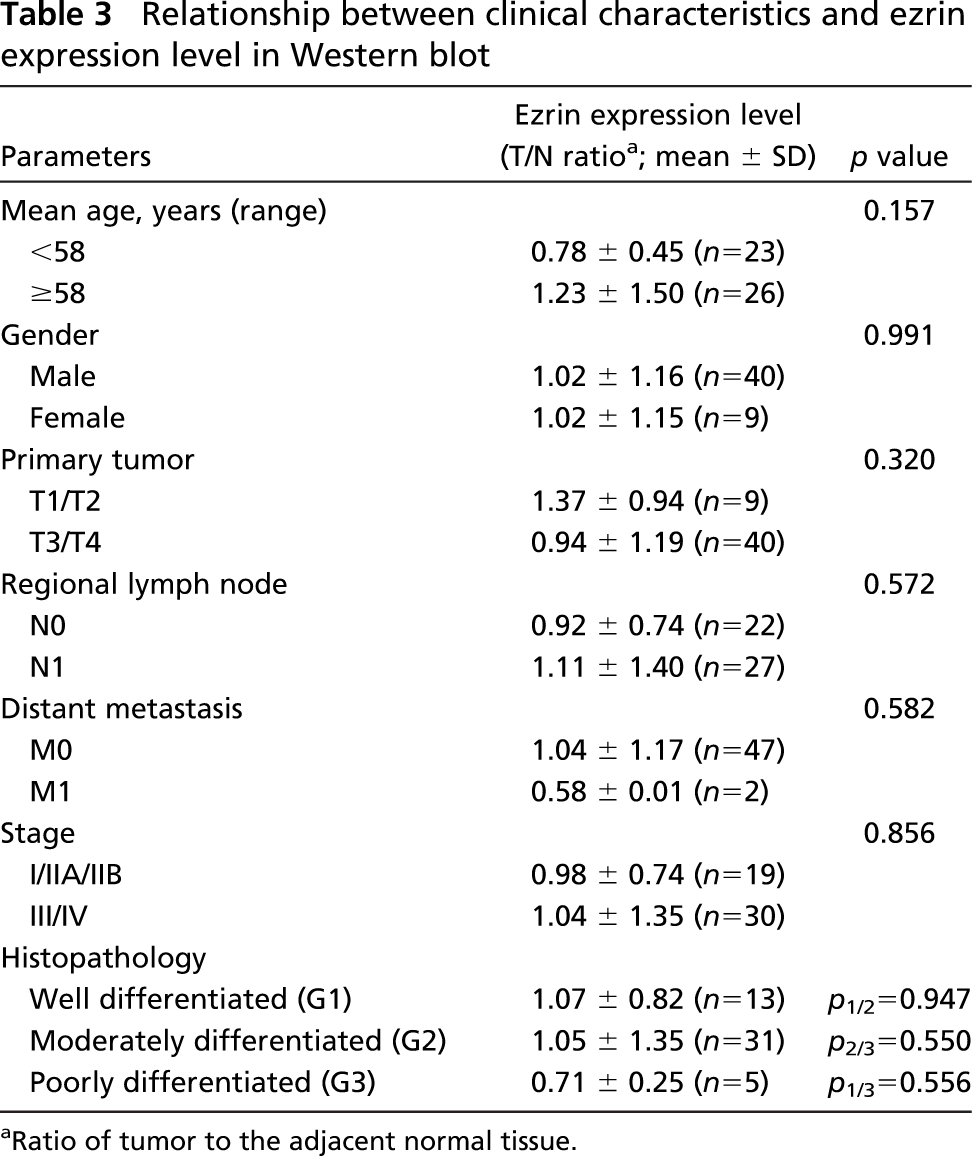
Ratio of tumor to the adjacent normal tissue.
Ezrin was demonstrated overexpressing in some tumor tissues or cell lines and was required for metastasis (Khanna et al. 2004; Yu et al. 2004; Elliott et al. 2005). Evidence suggested that ezrin was downregulated and played a suppressive role in tumor progression (Tokunou et al. 2000; Guan et al. 2002; Moilanen et al. 2003). Our present study showed that ezrin protein expression level in normal esophageal epithelium was higher than in ESCC in 73.5% of the 49 samples. In addition, ezrin mRNA level in normal esophageal mucosa was 3.60 ± 3.60 times that in ESCC. The inverse prognosis of ezrin expression in tumors indicates that there are cell-specific functions for ezrin in tumorigenesis (Hunter 2004).
Even though the mechanism of ezrin downregulation in tumor cells is not clear, it is almost certain that this aberrant expression of ezrin can be explained partly by a corresponding alteration on the mRNA level, as shown by real-time RT-PCR. Whether the reduced mRNA level in ESCC is owing to a decrease in the copy number of the gene or the upstream promoter region modulating ezrin transcription merits further studies.
Ezrin downregulation in tumor cells may have a significant role in tumorigenesis. Ezrin has been found to participate in the CD95-mediated apoptosis (Parlato et al. 2000; Lozupone et al. 2004) and T-cell activation (Autero et al. 2003). We also observed a higher proliferative potency in the ezrin downregulation group in our present study. The reduced expression of ezrin in tumor cells could result in decreased CD95-mediated apoptosis along with vigorous proliferation, as well as the lower level activation of T cells; thus, the tumor cells might obtain the ability to avoid immunosurveillance and immortalization. Recently, Yang and Hinds (2003) found that ezrin expression increased upon pRb-induced senescence and, more significantly, translocation of ezrin from the cytoplasm to membrane was concomitant with acquisition of the senescent phenotype. This translocation appeared to be the consequence of direct phosphorylation of T235 of ezrin by CDK5, which was activated in response to pRb expression. Furthermore, the study by Hiscox and Jiang (1999) showed that following inhibition of ezrin expression in colorectal cancer cells using antisense oligonucleotides, these cells displayed a reduced cell-cell adhesiveness together with a gain in their motile and invasive behavior and also displayed increased spreading over matrixcoated surfaces. The studies suggest that ezrin may perform a suppressive role in tumor progression. In fact, diminished ezrin expression in serous ovarian carcinoma was associated with shorter survival and a higher histological grade of the disease (Moilanen et al. 2003). However, we could not find a significant correlation between ezrin expression level and the clinical parameters. The results may be due to the limited amount of samples.
In conclusion, the redistribution of ezrin and reduced expression of ezrin both at mRNA and protein levels in ESCC suggest that the different ezrin staining patterns may be useful for the diagnosis of ESCC, and ezrin may play a suppressive role in the transformation and development of ESCC. Additional studies on a larger series of ESCC samples are warranted to confirm these findings, and further investigations are required to elucidate the mechanisms of translocation and downregulation of ezrin in ESCC.
Footnotes
Acknowledgements
This work was supported by the National Natural Science Foundation of China (No. 39900069, No. 30170428, No. 30370641, No. 30570849) and Guangdong Scientific Fund Key Items (No. 37788, No. 05104541).
We thank DaAn Gene Co., Ltd. of Sun Yat-sen University for their kind assistance in real-time RT-PCR and Lijun Tan and Dawn Harp, University of Michigan, for their help in reviewing the paper. We thank Professor MingYao Wu, Department of Pathology, Medical College of Shantou University for his kind help in pathological image analysis.