
Abstract
Unlike in crustacean freshwater species, the structure and ultrastructure of the excretory antennal gland is poorly documented in marine species. The general organization and ultrastructure of the cells and the localization of Na+,K+-ATPase were examined in the antennal gland of the adult lobster
From an osmoregulatory point of view, the juveniles and adults of homarid lobsters are osmoconformers in seawater and are slight hyperregulators in dilute media (Dall 1970; Charmantier et al. 1984, 2001). Osmoregulation in these animals is apparently achieved through hyperregulation of Na+ and, to a lesser extent, of Cl-, K+ is hyporegulated at high salinities, and Mg2+ is strongly hyporegulated at all salinities (Charmantier et al. 1984).
Because of the active implication of the antennal glands in osmoregulation of freshwater decapods through the production of dilute urine, their structure has been more intensively studied in these species than in closely related marine species. This is particularly true in the Astacidea, in which the structure of the antennal glands has been described in several species of crayfish (review in Khodabandeh et al. 2005a). These studies have shown that each antennal gland is composed of a single nephron-like unit including the coelomosac, the labyrinth, the nephridial tubule (nephridial canal), and the urinary bladder. Ultrafiltration occurs across the wall of the coelomosac, which presents typical podocytes. The filtrate then flows through the different parts of the efferent duct, where it is modified through absorption and secretion processes. The labyrinth, nephridial tubule, and even the bladder are involved in salt reabsorption, which results in the production of urine that is hyposmotic to the hemolymph of crayfish (Riegel 1966; Riegel and Cook 1975; Susanto and Charmantier 2000, 2001; Mc Mahon 2002; Vogt 2002; Khodabandeh et al. 2005b).
Although several physiological studies have been conducted on the antennal glands of homarid lobsters, no structural information is available on them except for the pioneering observations of Waite (1899) and Peters (1935). The nephridial canal, with its salt-reabsorbing function in crayfish, does not appear as compulsory in marine species such as homarid lobsters (Burger 1957; Dall 1970).
The aim of the present study was to: (a) describe the structure and ultrastructure of the different parts of the antennal glands in the adult European lobster
Materials and Methods
Animals
Adult
Histology
The antennal glands were surgically removed from cold-anesthetized lobsters. For light microscopy observations, sample fixations and other histological processes were performed as previously described (Khodabandeh et al. 2005a).
Electron Microscopy
For transmission electron microscopy, samples were fixed in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer for 24 hr at room temperature, pH 7.4, adjusted to 1030 mos-mol/kg by NaCl to avoid osmotic shock. They were then rinsed in sodium cacodylate buffer, and postfixed for 1 hr in a mixture (v/v) of 2% osmium tetroxide and 0.45 M sodium cacodylate buffer at room temperature. Other processes of the ultrastructural observations techniques have been described in Khodabandeh et al. (2005a).
For scanning electron microscopy, samples were placed in cold 4% glutaraldehyde in 0.1 M phosphate buffer, pH 7.4, containing 5% sucrose. After an initial 1 hr fixation, followed by rinsing in 0.1 M phosphate buffer containing 5% sucrose, the samples were postfixed for 1 hr in 2% osmium tetroxide in 0.1 M phosphate buffer, pH 7.4, containing 5% sucrose.
Immunocytochemistry
Immunolocalization of the Na+,K+-ATPase was performed through immunofluorescence light microscopy using a mouse monoclonal antibody IgGα5 raised against the α-subunit of the chicken Na+,K+-ATPase (Takeyasu et al. 1988) obtained from the Development Studies Hybridoma Bank, developed under the auspices of the NICHD, and maintained by the University of Iowa. In crustaceans, this antiserum has previously been used to quantify the expression of the α subunit after a dilute transfer in
After 24 hr in Bouin's fixator and embedment in paraplast, sections of 3–5 μm were cut on a Leitz Wetzlar microtome and collected on poly-L-lysine-coated slides. The immunocytochemistry procedure has been previously described (Lignot et al. 2001, 2005; Khodabandeh et al. 2005b). Posterior gills of the crab
Results
General Organization
In adult
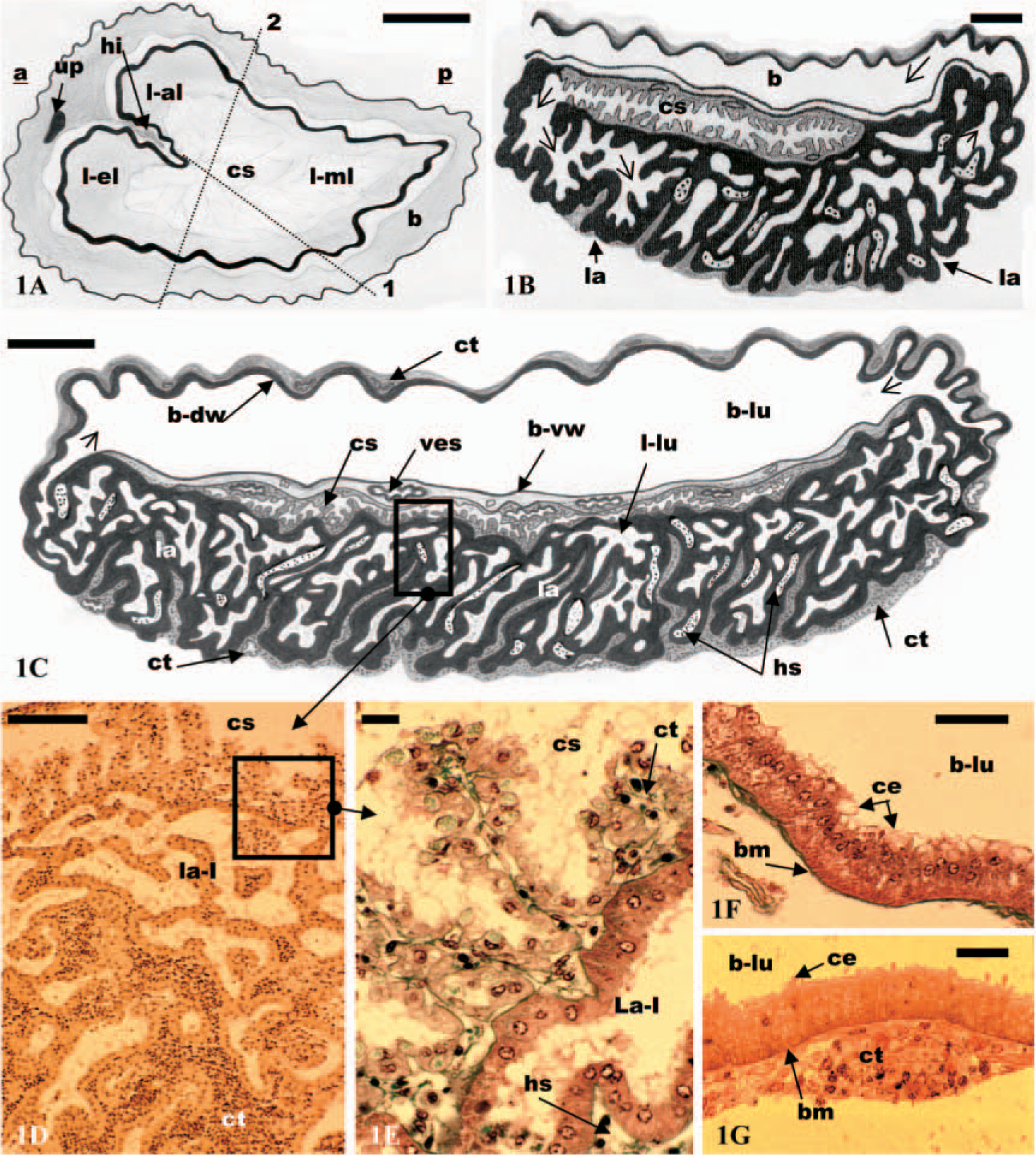
Antennal gland of
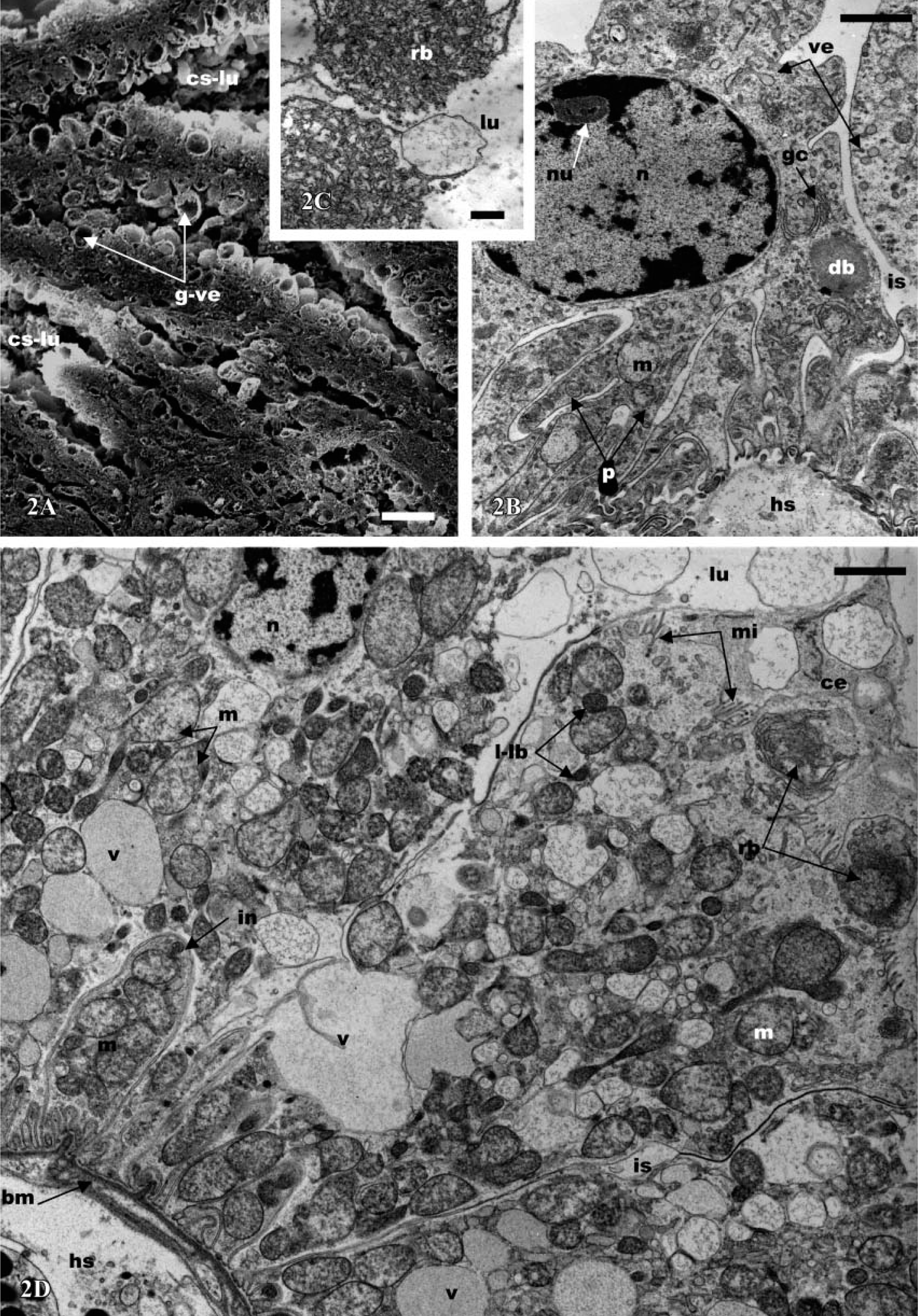
Antennal gland of
The labyrinth constitutes the greater bulk of the gland (Figures 1B and 1C). Its ventral part is covered by a connective tissue (Figures 1C and 1D). This tissue is highly vascular, mainly formed by the walls of hemolymph vessels and hemolymph spaces (not shown). The labyrinth (Figures 1B–1D) consists of a very complicated spongy-like tissue. The space between the epithelial lining is occupied by a connective tissue (Figures 1C and 1D). The dorsal face of the labyrinth is covered by the coelomosac, except in the border areas (Figure 1C). In this region, two subregions have been observed, labyrinth I and II. Labyrinth I immediately follows the coelomosac and forms the main part of the labyrinth. Labyrinth II is present only in the lateral border of the labyrinth where it connects with the bladder (Figure 1C). The lumen of this subregion is considerably reduced compared with the lumen of labyrinth I (not shown).
The bladder (Figure 1C) is a large reservoir, firmly attached to the dorsal face of the gland. Between the basal membrane of the epithelium of the bladder (ventral wall) and the coelomosac lies a rich plexus of vessels. The bladder (Figures 1F and 1G) is lined by a single layer of epithelial cells covered by conjunctive tissue, and a system of hemolymph vessels is present in between. The thickness of the conjunctive layer is more variable than that of the epithelial layer (Figure 1G). It is composed of flattened cells with a centrally located nucleus, organized into one to several cellular layers.
Ultrastructure
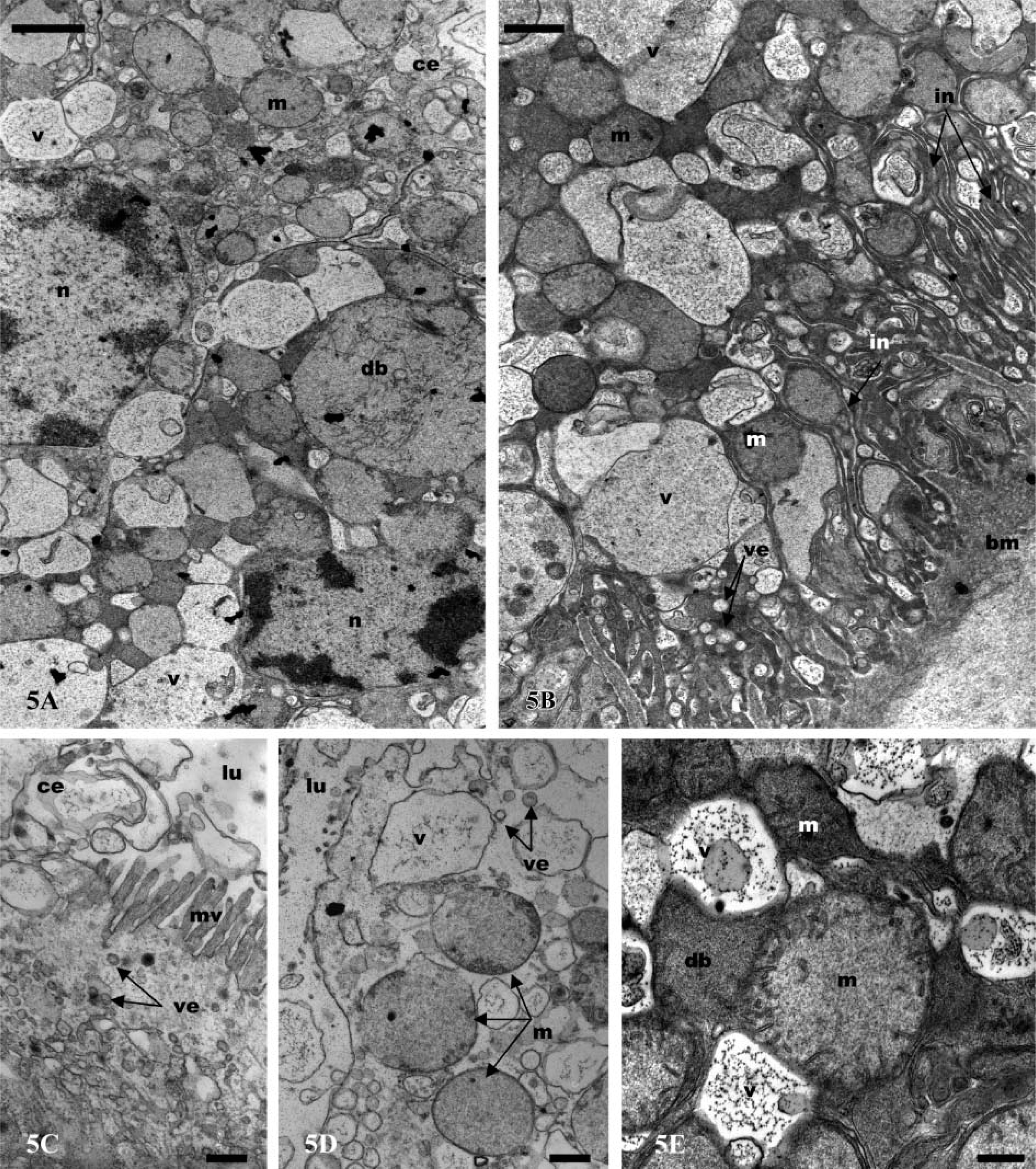
Ultrastructurally, the coelomosac cells (podocytes) present a variable shape. They possess distinctive pedicels or foot processes (Figure 2B) that extend to the basal plasma membrane separating the coelomosac cell from the underlying hemolymph space. The apical portions of all cells are devoid of microvilli and expand into voluminous globules, which are often two times larger in diameter than the basal portion (Figures 1A and 1B). These globular vesicles contain spherical structures that become detached into the lumen (Figure 2C). Consequently, the lumen of the coelomosac is more or less filled with different and various-sized globules that contain residual bodies (Figures 2A and 2C). The cells are separated by large intercellular space, but they are bound together by desmosome-like intercellular attachments. The main parts of the cells contain large intracellular dense bodies, mitochondria, vesicles, and vacuoles (Figure 2B). Typical Golgi complex systems are generally observed, consisting of the usual stacks of concentric or flattened cisternae (Figure 2B). The nucleus, round or oval with a voluminous nucleolus, is usually situated in the apical half part of the cells (Figure 2B).
In all parts of the labyrinth and of the bladder, the cells present several common features, including apical microvilli, apical cytoplasmic extrusions (Figure 2D; Figure 3A; Figures 4A and 4B; Figures 5A and 5B), and basal membrane infoldings associated with mitochondria (Figure 2A; Figure 3C; Figure 4C; Figure 5B). The arrangement of apical microvilli is usually disturbed by the formation of cytoplasmic extrusions that become detached and pass into the lumen (Figure 2D; Figure 3A; Figure 4A; Figure 5A). The cytoplasm contains lysosome-like bodies, abundant clear vacuoles of different sizes, dense bodies, mitochondria, and glycogen granules. Beside the typical small-sized mitochondria, other clear and voluminous mitochondria with small tubular cristae are observable (Figure 4D and Figure 5E). In the labyrinth I, two cell types are observable, columnar cells and cuboidal cells. (a) Columnar cells (Figure 2D) (average height 22 μm) possess an apical nucleus, dilated intercellular spaces, irregular apical microvilli, and irregular basal membrane infoldings associated with mitochondria. The large globular vesicles and cytoplasmic extrusions in these cells contain numerous small vesicles, vacuoles, and, frequently, dense spherical residual bodies. As in the coelomosac region, spherical residual bodies are often observed in the globular vesicles and in the lumen (not shown). (b) In cuboidal cells (Figures 3A-5E) (height 20 μm), the nucleus is centrally located; the main features of these cells include the presence of regular basal infoldings associated with round mitochondria, globules of glycogen, and lysosome-like bodies (Figure 3C). They display many apical swollen mitochondria, tight junctions, and zonula adherens (Figure 3E). Their apical large globular vesicles contain many vacuoles and swollen mitochondria, and no spherical structure is observable (Figure 3B). Vacuoles, lysosome-like bodies, glycogen globules, and mitochondria are consistently observable in close contact (Figure 3D). The labyrinth II cells (Figures 4A-4E) are columnar-shaped (average height 27 μm) and their cytoplasm is usually clear. On their basal side, short basal infoldings are associated with mitochondria, vesicles, and numerous and large vacuoles containing fine particles (Figure 4C). Their apical portion and cytoplasmic extrusions contain clear vacuoles, small vesicles, mitochondria, and lysosome-like bodies (Figures 4A and 4D). Besides these cytoplasmic extrusions, well-developed vacuoles and large, globular vesicles containing fine particles of labyrinth II cells bulge into the lumen and are free from the cell body (Figures 4B and 4E).
The bladder cells (Figures 5A-5E) present different sizes according to their location. The ventral wall of the bladder is lined by a thin epithelium (average height 15 μm), whereas the lateral and dorsal walls are limited by a thick epithelium (average height 45 μm). The bladder cells are columnar with an apical nucleus and irregular apical microvilli. They are filled with numerous and clear vacuoles of different sizes containing fine particles, frequently surrounded by an electron-dense cisterna, or associated with dense bodies and lysosome-like bodies (Figures 5B and 5E). They possess apical cytoplasmic extrusions into the lumen that contain clear vacuoles; there is no apical globular vesicle (Figures 5C and 5D). Small endocytic vesicles are frequently present in the apical region (Figures 5C and 5D). As in the labyrinth cells, clear and voluminous mitochondria with small tubular cristae are observable in the apical extrusions (Figures 5A and 5D). The basal side of the cells is dense and presents deep membrane infoldings associated with mitochondria, sparse vacuoles, and small, clear vesicles (Figure 5B). The bladder cells possess a thick basal lamina (Figure 5B).
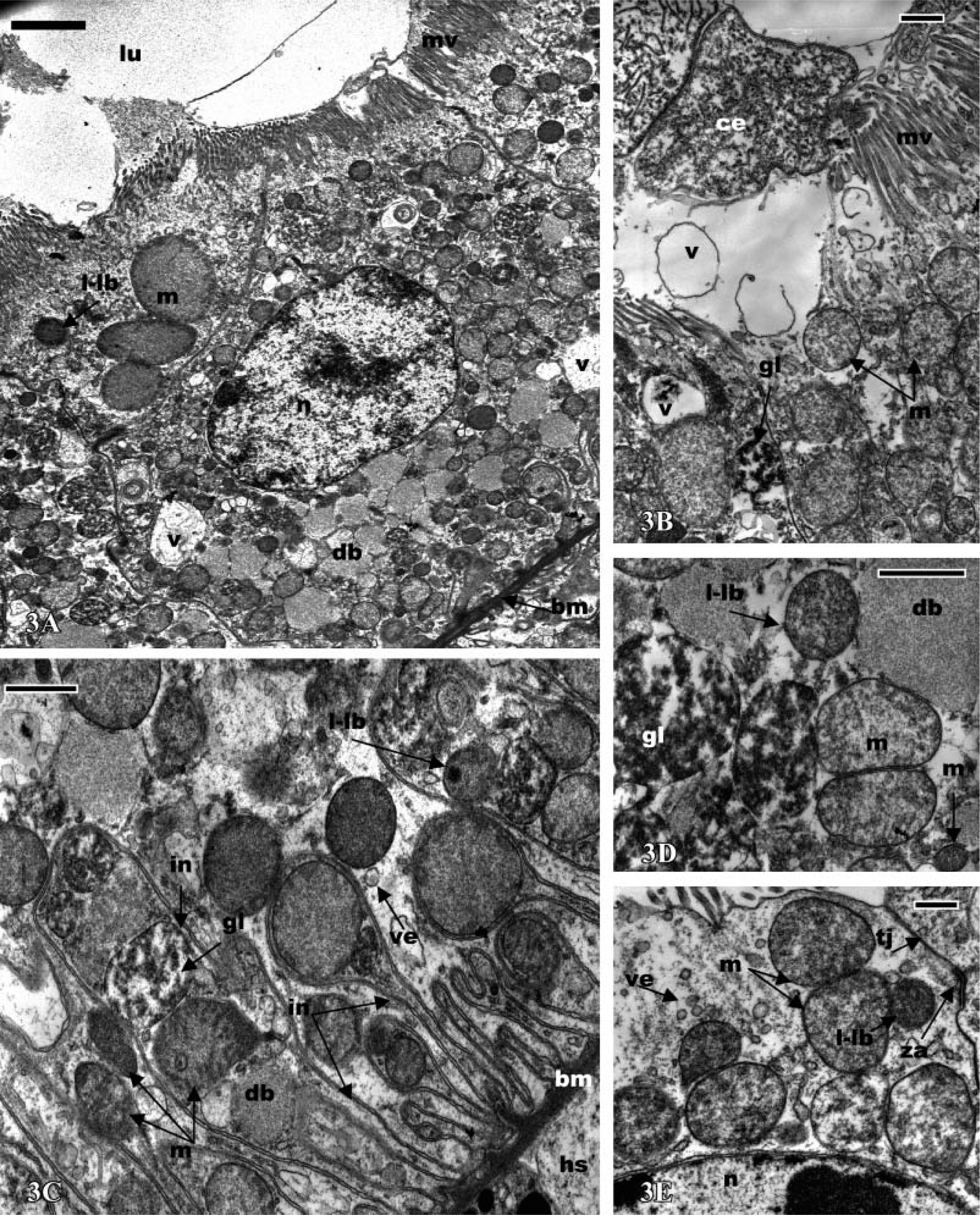
Antennal gland of
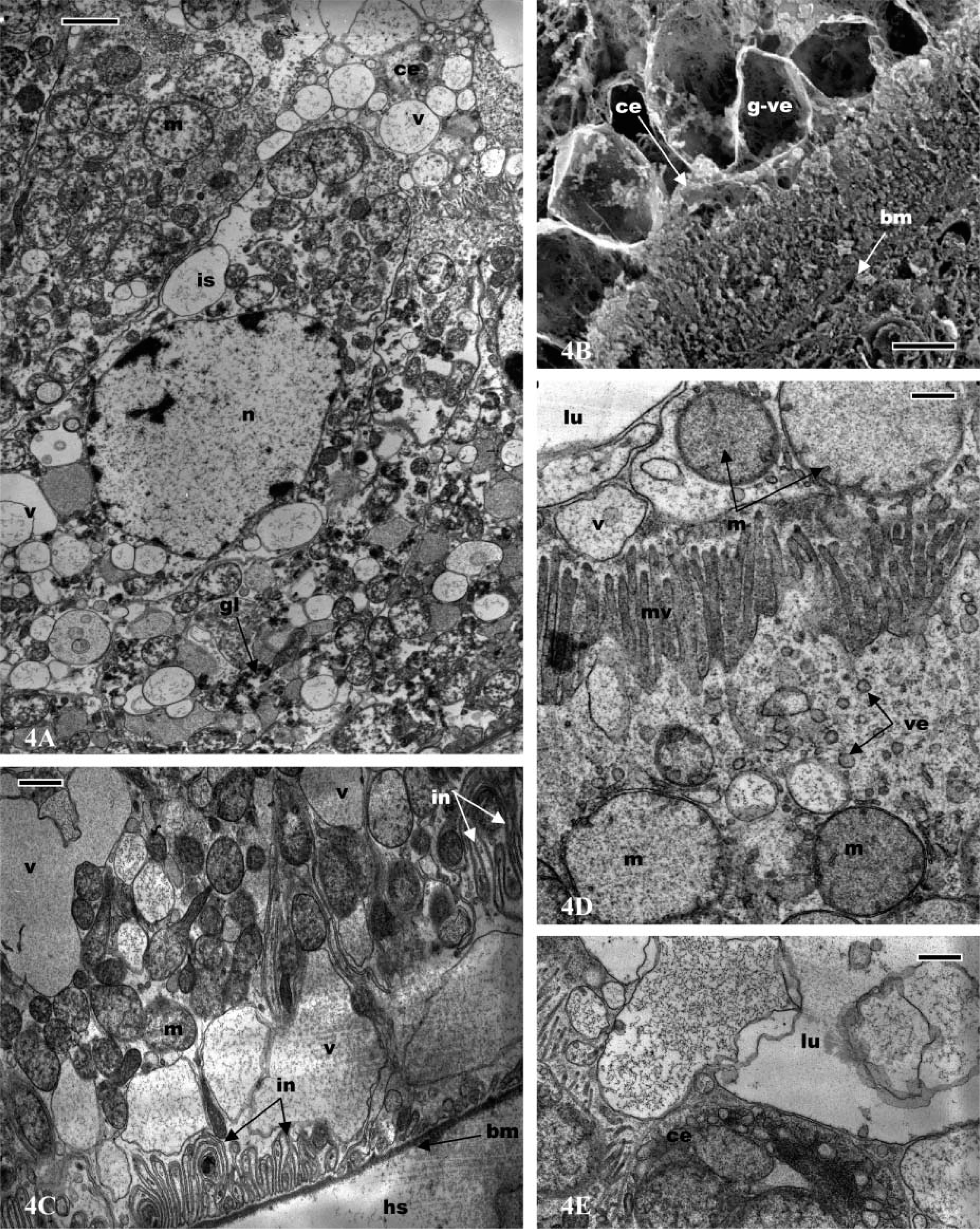
Antennal gland of
Antennal gland of
Immunocytochemistry
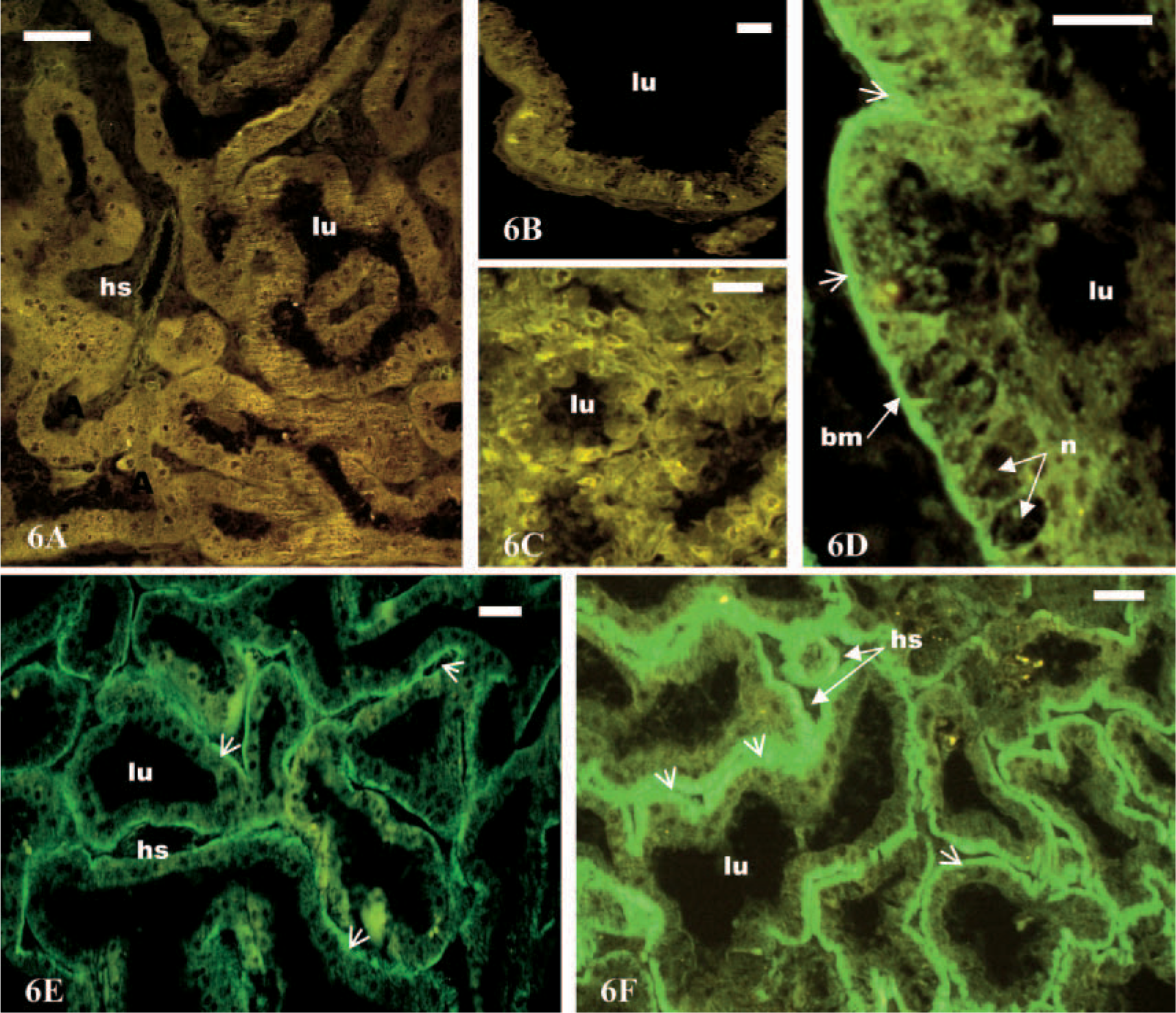
Immunofluorescence was used for the localization of the Na+,K+-ATPase, and the fixation and staining process leaves a good antigenicity. Positive control crab sections were constantly brightly immunostained (not shown). Negative control sections showed no specific binding within the different parts of the antennal gland (Figures 6A and 6B). No immunoreactivity was detected in the coelomosac cells (Figure 6C). Na+,K+-ATPase was localized in the labyrinth and the bladder regions of the antennal glands (Figures 6D-6F). The cells of labyrinth I showed weak immunostaining (Figure 6E). A stronger immunoreaction was observed in the cells of labyrinth II (Figure 6F) and in those of the thick wall of the bladder (Figure 6D). In all immunostained cells, the basal region showed a much higher fluorescence than the apical side, and the nucleus was free of fluorescent marker (Figures 6D–6F).
Discussion and Conclusion
To our knowledge, this study is the first ultrastructural investigation of the antennal glands of homarid lobsters. In contrast to the situation in freshwater species, no nephridial tubule was observed between the labyrinth and the bladder. In crayfish, the main function of the nephridial tubule is salt reabsorption, which results in the production of hyposmotic urine, a key adaptation to the low ion concentration of freshwater (Peterson and Loizzi 1974; Sesma et al. 1983; Fuller et al. 1989; Khodabandeh et al. 2005a). This compartment is not functionally necessary in
The most striking features of the coelomosac cells (podocytes) are the presence of basal pedicles, large lateral channels, intracellular large dense bodies, Golgi complex, vesicles, and vacuoles. The cells possess also endocytic vesicles and residual bodies that become detached and pass (as apical globular vesicles) into the lumen of the coelomosac. They are similar to podocytes described in other species of crustaceans particularly in crayfish (Khodabandeh et al. 2005a). Their cytological features and previous investigations in crabs and crayfish have shown that the coelomosac cells perform an ultrafiltration and secretory activity similar to that of the vertebrate podocytes (Johnson 1980; Fuller et al. 1989; Khodabandeh et al. 2005a). The apical globular globules (“formed bodies” of Riegel 1966) are analogous to the dense bodies extruded by vertebrate podocytes. No Na+,K+-ATPase immunoreactivity was detected in the coelomosac cells, a result in agreement with the ultrastructure of these cells. Previous investigations regarding the activity of Na+,K+-ATPase in the antennal glands of
The general common features of the epithelial cells of the labyrinth and bladder as observed in the present study are apical microvilli, apical cytoplasmic extrusions, endocytosis vesicles, cytoplasmic vacuoles, and basal plasma membrane infoldings associated with mitochondria. In general, these characters suggest the existence of active ion transport, potentially associated with reabsorption, secretion, or uptake of materials such as glucose, amino acids, and small proteins, and reabsorption of solute and water, respectively. Morphological differences were observed between the different regions of labyrinth I and II. In labyrinth I, we observed two different cell types: columnar and cuboidal. They might be similar cells, but at different physiological states, as previously described in the labyrinth epithelium of the blue crab
The bladder cells present different sizes (15-50 μm) according to their location. Morphological similarities between the cells of labyrinth II and the bladder suggest functional analogies between these two sites. Because Na+,K+-ATPase was detected in the bladder cells, a last modification of the filtrate would occur in this region of the antennal gland before the release of urine. Investigations of the bladder cells in crayfish and crabs have shown that they are involved in active ion transport (Johnson 1980; Miller 1989; Khodabandeh et al. 2005a,b) or in the transport of solutes such as carbohydrates (Gross 1967; Holliday and Miller 1984; Sarver et al. 1994) and organic acids (Pritchard and Miller 1991). Compared with the labyrinth cells, the bladder cells present more developed basal infoldings and a denser network of vacuoles. Using the same antibody concentration, the immunoactivity of Na+,K+-ATPase was always more intense in the bladder than in the labyrinth, as in
In conclusion, labyrinth and bladder cells are involved in different excretory and reabsorption functions. The presence of Na+,K+-ATPase in both structures could be explained in two ways: (a) the enzyme provides only the driving force for the transport of solutes such as sugar or amino acids or 2) the labyrinth and bladder cells are involved in active ion exchanges
Lobsters can be found in coastal waters and estuarine habitats (Jury et al. 1994; Lawton and Lavalli 1995), where salinity fluctuates. At low salinity, they are able to slightly hyperosmoregulate (Charmantier et al. 1984) and to maintain their urine concentration slightly below the hemolymph osmolality by 20–50 mosm/kg (Dall 1970). This difference might originate from limited selective ion reabsorption through the labyrinth and bladder cells that present features of ionocytes including the presence of Na+,K+-ATPase. In conclusion, the antennal glands of adult
Footnotes
Acknowledgements
Thanks are due to the University of Tarbiat Modarres and Ministry of Science, Research and Technology, Islamic Republic of Iran, for financial aid and support. Special thanks also to Dr. Khosrov Piri, responsible for Iranian student service in Paris, and F. Aujoulat, C. Blasco, E. Grousset, J.P. Selzner of the University of Montpellier II for their technical help.