
Abstract
In rat lung, the definitive alveoli are established during development by the outgrowth of secondary septa from the primary septa present in newborn; however, the mechanism of alveolar formation has not yet been fully clarified. In this study, we characterize the septal interstitial cells in developing alveoli. During the perinatal period, alpha-SMA–containing slender cells were found in the primitive alveolar septa. Alpha-SMA–containing cells were detected at the tips of the septa until postnatal day 21, when the alveolar formation was almost completed, but disappeared in adult. Immunoelectron microscopy demonstrated that alpha-SMA is localized mainly in the cellular protrusions, which are connected with the elastic fibers around the interstitial cells. Developmentally regulated brain protein (drebrin) is also located in the cell extensions containing alpha-SMA in immature alveolar interstitial cells. In adult lung, alpha-SMA–positive cells are located only at the alveolar ducts but are not found in the secondary septa. Desmin is expressed only in alpha-SMA–containing cells at the alveolar ducts but not in those at the tip of alveolar septa. These results suggest that a part of the septal interstitial cells are temporarily alpha-SMA– and drebrin-positive during maturation. Alpha-SMA– and drebrin-containing septal interstitial cells (termed septal myofibroblast-like cells) may play an important role in alveolar formation.
D
In the stages of lung maturation, specific temporal–spatial interactions between mesenchymal and epithelial cells are required to establish an effective air–blood barrier (Chuang and McMahon 2003). In lung, alveolar epithelial cells secrete platelet-derived growth factor (PDGF), the receptor of which is expressed in interstitial fibroblasts (Bostrom et al. 1996; Lindahl et al. 1997). A mouse PDGF-A null allele has been shown to be homozygous lethal. Postnatally surviving PDGF-A–deficient mice develop lung emphysema because of the failure of alveolar septation. This is apparently caused by the loss of specialized interstitial fibroblasts in alveoli and associated elastin fiber deposits (Bostrom et al. 1996). Although the mechanism that governs the outgrowth of secondary septa is not known, interstitial matrix proteins, especially elastin, seem to be required for the initiation and progression of alveolization (James et al. 1998; Noguchi et al. 1989). It is suggested that the precursor of elastin, tropoelastin, is synthesized and secreted by a specialized interstitial cells (fibroblasts) in the alveoli (Myers et al. 1983); however, the interstitial fibroblasts that play a role in alveolar maturation are not fully characterized.
Generally, fibroblasts are responsible for the production and secretion of extracellular matrix components, proteolytic enzymes, and various cytokines (Sappino et al. 1990). A line of evidence suggests that fibroblasts are likely to have specialized functions in each of these tissues. It has been accepted that the fibroblasts are heterogeneous (Komuro 1990). Several kinds of fibroblasts exist in the alveolar interstitial space, but it is difficult to distinguish the fibroblasts in the tissue by their morphological characteristics.
Alpha–smooth muscle actin (alpha-SMA) is a differentiation marker of smooth muscle cells and is also present in a special type of fibroblast called myofibroblast. Previous reports demonstrated that cells containing alpha-SMA are found in maturing alveolar interstitium (Vaccaro and Brody 1978; Mitchell et al. 1990; Wagner et al. 2003). Myofibroblasts expressing alpha-SMA are instrumental in wound contraction during normal wound healing (Powell et al. 1999; Tomasek et al. 2002). Alpha-SMA within myofibroblasts becomes organized in filamentous bundles, called stress fibers, that allow the retractile movement producing wound contraction (Grinnell 1994).
Recently, an actin-binding protein, developmentally regulated brain protein (drebrin), has been localized in the specialized interstitial cells of renal glomerulus (i.e., mesangial cells) (Peitsch et al. 2003). Drebrins are a family of actin-binding proteins originally identified in neuronal cells (Peitsch et al. 2001; Shirao et al. 1992). Drebrin has been involved in the regulation of actin filament organization, especially in the formation of neuronal cell processes and cell protrusions of motile cells, especially lamellipodia and filopodia (Peitsch et al. 2001). Electron microscopy (EM) observation indicates that both mesangial cells and alveolar interstitial cells are tightly associated with the neighboring vasculature by their elongated cell protrusions. This finding led us to investigate the expression of drebrin in alveolar interstitial cells.
Skalli et al. (1989) have distinguished four different types of myofibroblasts according to their expression of alpha-SMA and the intermediate filaments, desmin and/or vimentin. Thus, we tried to examine which kinds of intermediate filament proteins are expressed in alveolar interstitial cells during maturation. The expression of intermediate filaments depends on cell type at the phase of cellular differentiation. In general, epithelial cells express cytokeratin-type intermediate filaments, whereas mesenchymal cells express vimentin-type ones. Desmin-type intermediate filaments, which are expressed predominantly in skeletal muscle cells, cardiac myocytes, and certain smooth muscle cells, are also found in some non-muscle cells. A major function shared by several types of cytoplasmic intermediate filaments is to stabilize cellular architecture against the mechanical forces to which it is subjected (Fuchs and Weber, 1994). The alveoli need steady and tidy skeletal elements to maintain their unique structure in such a severe environment; however, the function of intermediate filaments in lung has not been clarified.
In the present study, we characterized the alveolar interstitial cells expressing unique cytoskeletal proteins during postnatal alveolar maturation using immunocytochemistry and EM.
Materials and Methods
Reagents and Antibodies
Bovine serum albumin (fraction V) was obtained from Sigma-Aldrich (St. Louis, MO). Mouse monoclonal anti-alpha-SMA antibody (IgG) and rabbit anti-rat elastin antibody were from Chemicon (Temecula, CA). Rabbit anti-laminin antibody and mouse monoclonal anti-desmin antibody (IgG) were from Dako (Carpinteria, CA). Guinea pig anti-drebrin antibody was from Progen (Heidelberg, Germany). Rhodamine (TRITC)-conjugated donkey anti-mouse IgG F(ab')2 fragment, fluorescein (FITC)-conjugated donkey anti-rabbit IgG F(ab')2 fragment, rhodamine-conjugated donkey anti-guinea pig IgG F(ab')2 fragment, and peroxidase-conjugated goat anti-mouse IgG F(ab')2 fragment were from Jackson Immunoresearch Laboratories (West Grove, PA).
Animals
All the procedures performed on laboratory animals were approved by the Institutional Animal Care and Use Committee of Juntendo University School of Medicine, and all the animal experiments were carried out in compliance with the guidelines for animal experimentation of Juntendo University School of Medicine. Wistar rats (embryonic day 20 to 6 weeks old) were obtained from Charles River Japan (Kanagawa, Japan). They were kept in an air-conditioned room and maintained on a commercial stock diet and tap water ad libitum. Gestational age was calculated from the first day when a vaginal plug was seen.
Confocal Laser-scanning Microscopy
Rat lungs were perfused with 4% paraformaldehyde fixative buffered with 0.1 M phosphate buffer (PB, pH 7.4) under anesthesia with Nembutal (Dainippon Pharmaceutical; Osaka, Japan) and then were cut into small pieces. These samples were immersed in the same fixative for approximately 30 min. After washing with PBS, the tissue was immersed successively in PBS solutions containing 10%, 15%, and 20% sucrose for 4 hr, 12 hr, and 4 hr, respectively. Cryosections (thickness, 5–10 μm) were cut using a Jung Frigocut 2800E (Leica; Wetzlar, Germany) and then mounted on silane-coated glass slides (Dako; Carpinteria, CA). The cryosections were rinsed with PBS and blocked in blocking solution (0.1% BSA in PBS). The sections were incubated for 2 hr at RT with mouse monoclonal anti-alpha-SMA antibody (1:100), mouse monoclonal anti-desmin antibody (1:100), rabbit anti-laminin antibody (1:100), rabbit anti-rat elastin antibody (1:50), or goat polyclonal anti-drebrin antibody (1:100). Subsequently, the sections were incubated for 1 hr at RT with TRITC-conjugated anti-mouse IgG (1:200), TRITC-conjugated anti-rabbit IgG (1:200), or FITC-conjugated anti-rabbit IgG (1:100). Fluorescence specimens were viewed with confocal laser scanning microscope LSM510 (Carl Zeiss; Oberkochen, Germany).
Transmission Electron Microscopy
Rat lungs were perfused with 2.5% glutaraldehyde fixative buffered with 0.1 M PB (pH 7.4) under anesthesia with Nembutal and immersed in the same fixative for approximately 12 hr. The specimens were sectioned at 200-μm thickness with a Microslicer (Dosaka; Kyoto, Japan) and processed further by modified cold dehydration technique. The samples were successively immersed in 0.1% OsO4 in 0.1 M PB for 30 min, in 5% extracts of oolong tea in 0.05 M maleate buffer for 3 hr, and in 1% uranyl acetate in the same maleate buffer solution. Subsequently, the specimens were dehydrated with a graded series of acetone at 0C to −30C before being embedded in Epon 812 (Oken; Tokyo, Japan). These procedures enabled detailed morphological observations of the extracellular matrices and intracellular fibrils. Ultrathin sections (80 nm) were processed with a diamond knife, transferred to copper grids (50 mesh) that had been coated with Formvar membrane (Oken), stained with uranyl acetate and lead citrate, and observed in a Hitachi 7100 transmission electron microscope (Hitachi; Tokyo, Japan).
Immunoperoxidase Labeling on Cryostat Sections
Paraformaldehyde-fixed lung tissues from embryonic day 20 and postnatal day 2, 5, 12, and 21 animals were cut on a cryostat into 50-μm thick sections. Cryostat sections were incubated overnight at RT with mouse monoclonal anti-alpha-SMA antibody (1:50) followed by incubation for 2 hr in F(ab')2 fragments of goat anti-mouse IgG conjugated to horseradish peroxidase (1:100). The sections were fixed in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.4), reacted with diaminobenzidine and were processed for EM.
Results
Immunohistochemical Characterization of Septal Interstitial Cells
The developmental changes in lung structure were examined histochemically in prenatal and postnatal rats (from embryonic day 20 to postnatal week 6) using antibodies against alpha-SMA and laminin (Figure 1) or against alpha-SMA and elastin (Figure 2). Anti-laminin antibody was used to stain the basement membrane. At day 20 of gestation, the pulmonary tissue was composed of primitive alveolar cavities separated from each other by thick primitive septa. The primitive cavities corresponded to both the definitive alveolar ducts and alveolar sacs (Figures 1A and 2A). At birth, the primitive cavities were enlarged significantly, and the primitive septa contained numerous empty blood capillaries that frequently bulged into the primitive alveolar sacs as well as small ridges that protruded into the primitive cavities (Figures 1B and 2B). Until postnatal day 21, the small ridges elongated continuously to become the secondary septa that subdivided the primitive alveolar cavities into the primary and secondary alveolar saccules (Figures 1C, 1D, 2C, and 2D). In adult rats, the thickness of definitive alveolar septa decreased considerably (Figures 1E and 2E).
The signal for laminin was observed along the airway and capillary endothelial cells at all examined stages of the lung (Figures 1A–1E). At day 20 of gestation as well as until postnatal day 12, the primitive septa contained slender, elongated alpha-SMA–positive cells in the interstitial space (Figures 1A–1C). In addition to the slender alpha-SMA–positive cells, a population of round alpha-SMA–positive cells appeared at the tips of the developing secondary septa on postnatal day 12 and 21, whereas the slender alpha-SMA–positive interstitial cells disappeared on postnatal day 21 (Figures 1C and 1D). In adult rat lung, alpha-SMA–positive cells had disappeared from the tips of secondary septa, and a third population of alpha-SMA–positive cells emerged at the surface of the alveolar duct (Figure 1E).
Elastic fibers are abundant in septal interstitium, and the relationship between elastin and alpha-SMA–positive cells was examined by double immunostaining with anti-elastin antibody and anti-alpha-SMA antibody (Figure 2). Until postnatal day 12 (Figures 2A, 2B, and 2C), a large amount of elastic fibers surrounded the primitive alveolar cavities, and alpha-SMA–positive slender cells were always associated with the elastic fibers. At the period of alveolar maturation (Figures 2C and 2D), the elastic fibers were concentrated in the tips of the elongated secondary septa with alpha-SMA–positive round cells. In adult rat lung, the tips of the secondary septa contained a concentrated population of elastic fibers and exhibited no signals for alpha-SMA. In the region of alveolar ducts, the third type of alpha-SMA–positive cells were found together with considerable amounts of elastic fibers (Figure 2E).
Electron Microscopy of Septal Interstitial Cells
Alpha-SMA–positive round cells at the tips of secondary alveolar septa were found on postnatal days 12 and 21. Electron microscopy showed that the alpha-SMA–positive cells had well-developed rough endoplasmic reticulum and Golgi apparatus in the cytoplasm and that their elongated processes were surrounded by elastic fibers and collagen fibers on postnatal days 12 and 21 (Figures 3A and 3B). In the adult rat lung, alpha-SMA–positive cells were not seen in the alveolar septa, but interstitial cells surrounded by elastin and collagen fibers were still located in the interstitium (Figures 3C and 3D).
EM observation showed that there were two distinct types of interstitial cells at the tips of the developing alveolar septa. One type possessed well-developed rough endoplasmic reticulum and Golgi apparatus in the cytoplasm, and the other had lipid droplets in the cytoplasm. When we introduced anti-alpha-SMA antibodies in the developing lung, only the former type of cells expressed alpha-SMA (Figure 4B). Alpha-SMA was distributed in the cell periphery facing the blood capillary and in the cell protrusions (Figures 4A, 4C, and 4D).
Two Types of Interstitial Cells Are Found in the Septal Interstitium
To characterize the alpha-SMA–positive interstitial cells at the tips of secondary septa on postnatal days 12 and 21 and the alpha-SMA–positive interstitial cells in the alveolar duct in the adult rat, we examined the developing and mature pulmonary tissue histochemically using antibodies against desmin intermediate filament and alpha-SMA (Figure 5). On postnatal days 2 and 12, desmin-positive cells were occasionally found that were completely distinct from alpha-SMA–positive ones (Figures 5A–5C). Interestingly, in the adult rat, the alpha-SMA–positive interstitial cells beneath the alveolar duct expressed desmin (Figure 5D).
Alpha-SMA–positive cells in septal interstitium are characterized by elongated cell processes. Actin-binding proteins are involved in cell process formation. One of the actin-binding proteins, drebrin, is a family of actin-binding proteins originally identified in neurons. In the nervous system, drebrins play a role in the formation of neurites. We examined the expression of drebrin in rat lung and found that drebrin was located in the cell processes of alpha-SMA–positive cells during the maturation of alveolar septa (Figures 6A–6F). Drebrin was not expressed in the interstitial cells in alveoli and alveolar ducts of adult rat lung (Figures 6G–6I).
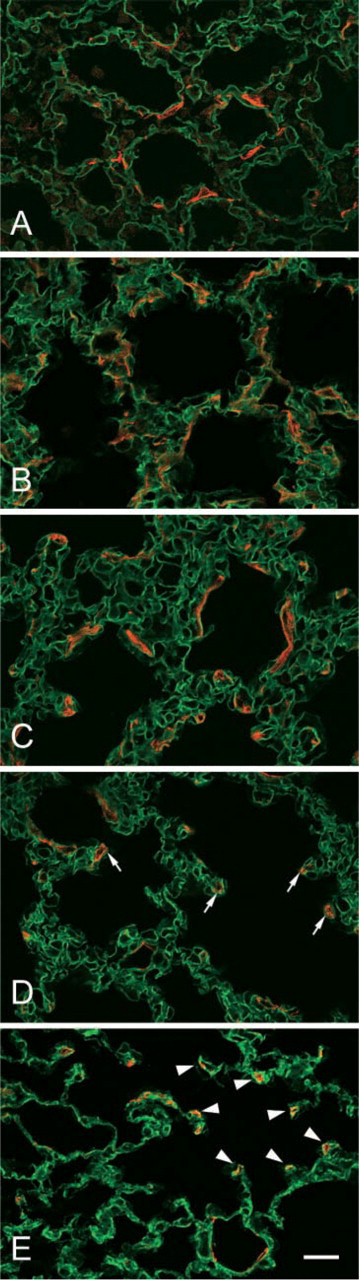
Localization of alpha-SMA and laminin in developing rat lung. The developing and mature rat lungs are labeled with antibodies against alpha-SMA (TRITC) and laminin (FITC). In all of the stages observed in this study, laminin is localized along the alveolar capillaries. In the prenatal and postnatal rats [(
Discussion
In mammals, including humans, the formation of pulmonary alveoli occurs substantially after birth (Thurlbeck 1975). In the present study, we identified the interstitial cells expressing alpha-SMA and drebrin at the tips of the saccular interstitium during alveolar maturation. The expression of alpha-SMA and drebrin in cells located in the septal interstitium was transient. We could detect the cells only in the immature lung, not in the adult one. On the other hand, alpha-SMA–containing cells were seen only at the alveolar duct in mature lung. Desmin, a kind of intermediate filaments, is often used for the classification of interstitial cells (Kapanci et al. 1992; Mitchell et al. 1990). Alpha-SMA–positive cells at the alveolar duct expressed desmin, whereas the septal alpha-SMA–positive cells did not. The dominant intermediate-type filament of visceral smooth muscle cells is desmin (Gabbiani et al. 1981). The similar localization of desmin in alpha-SMA–positive cells has been reported (Mitchell et al. 1990). Therefore, an alpha-SMA–containing cell at the alveolar duct is thought to be a kind of smooth muscle cell, as previously reported by Leslie et al. (1990). Furthermore, we could detect the desmin-positive but alpha-SMA–negative cells alongside the alpha-SMA–positive cells at the septal interstitium. Desmin-positive cells at the septal interstitium were identified as lipid-laden interstitial cells by immunoelectron microscopy. Based on the ultrastructural observation, Vidic et al. (1989) also reported two types of the cells in the septal interstitium, myofibroblasts and lipofibroblasts. Based on the morphology of the subcortical cytoskeletons, it has been proposed that the lipofibroblasts are the precursors of the myofibroblasts (Kaplan et al. 1985); however, we could distinguish two types of the septal interstitial cells by using desmin staining. Thus, it is thought that alpha-SMA–positive but desmin-negative cells are precursors of myofibroblasts and that alpha-SMA–negative but desmin-positive cells are precursors of lipofibroblasts. These results suggest that desmin is a good marker for distinguishing the septal alpha-SMA–containing cells (septal myofibroblast-like cells) from other types of cells (e.g., lipofibroblasts and the smooth muscle cells at the alveolar duct) at the level of light microscopy.
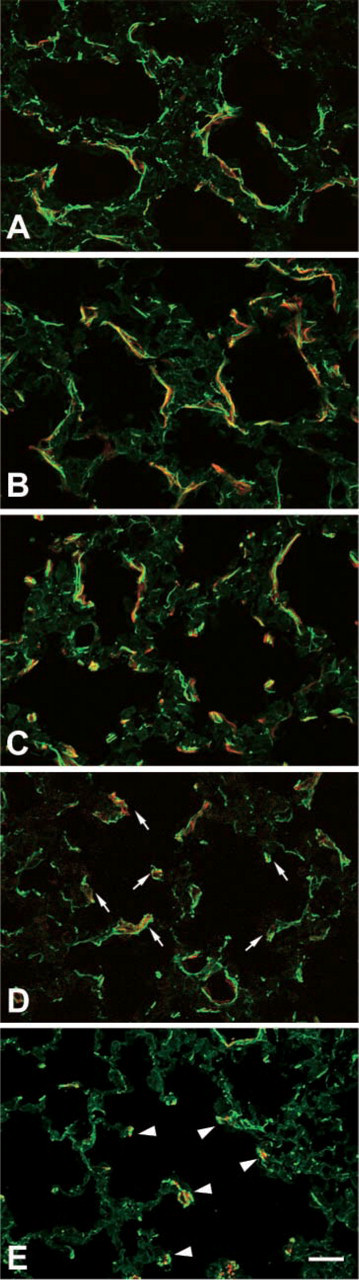
Localization of elastin and alpha-SMA in septal interstitium of developing rat lung. The developing and mature rat lungs are labeled with antibodies against alpha-SMA (TRITC) and elastin (FITC). During the septal formation, the alpha-SMA–positive elongated cells (TRITC) in the primitive alveolar septa (
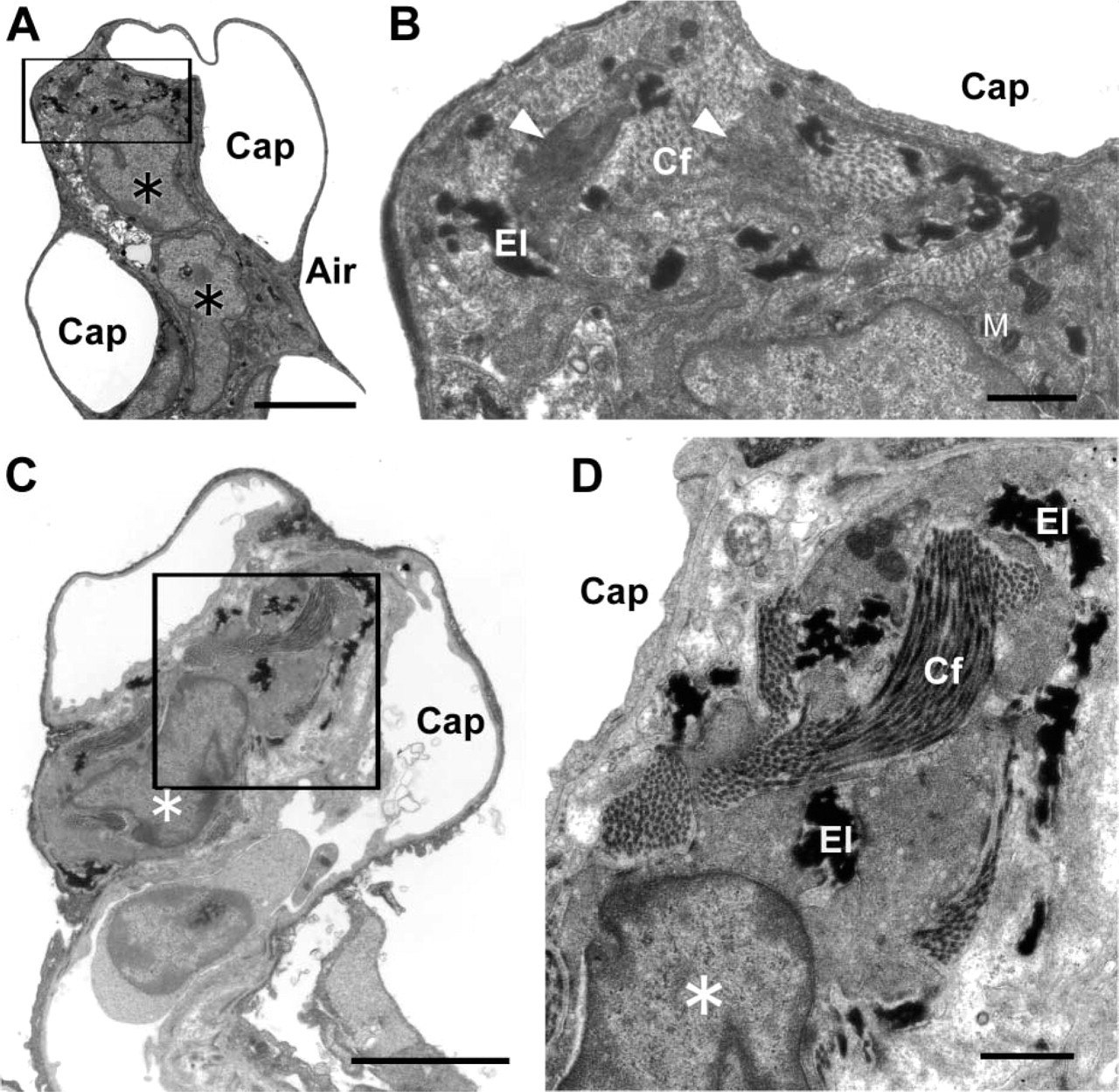
Electron microscopy (EM) of septal interstitial cells. As postnatal day 12, the alpha-SMA-containing cells (asterisk) are located at the tip of the secondary septum. EM observation shows that the septal interstitial cells have well-developed rough endoplasmic reticulum and Golgi apparatus in the cytoplasm, and their elongated processes are surrounded by elastic fibers (El) and collagen fibers (Cf) through the alveolar maturating period (
Our data on the cellular localization of alpha-SMA during lung maturation indicate that alpha-SMA is located exclusively at the protrusions of the septal myofibroblasts. Myofibroblasts are recognized to play a central role in closing wound tissue through their capacity to produce a strong contractile force (Serini and Gabbiani 1999). Exogenous expression of alpha-SMA was induced to enhance fibroblast contractile activity (Hinz et al. 2001). Alpha-SMA expressed in the interstitial cells located within the alveolar walls might play a role in making the tensile forces of the alveolar parenchyma (Adler et al. 1989). EM observations also suggest that the filopodia-like protrusions of the cells contain numerous actin bundles. Cell process formation is closely related to cellular adhesion to other cells or to the extracellular matrix and to cell movements. These events are achieved by the assembly of actin and actin-binding proteins (Tomasek et al. 2002). We could detect that drebrin, a family of actin-binding proteins, is expressed in septal alpha-SMA–positive cells. We found that drebrin is located in the cell processes of septal myofibroblast-like cells during alveolar maturation. The transient expression of drebrin in the myofibroblasts suggests that the formation of cellular projection is related to the elongation of the secondary septum. Drebrin is also found in the mesangial cells, which provide the mechanical support for glomerular capillaries by generating an inwardly directed counter-force and regulating the glomerular capillary wall tension by contraction and relaxation (Kriz et al. 1990). Like mesangial cells, septal myofibroblasts might play a role in regulating the capillary wall tension via the interaction between the cell processes and the elastic fibers located beneath the capillary. Interestingly, drebrin is not expressed in the interstitial cells in the alveolar ducts expressing alpha-SMA during maturation, suggesting that the alpha-SMA–positive interstitial cells in the alveolar ducts are distinct from the septal myofibroblast-like cells.
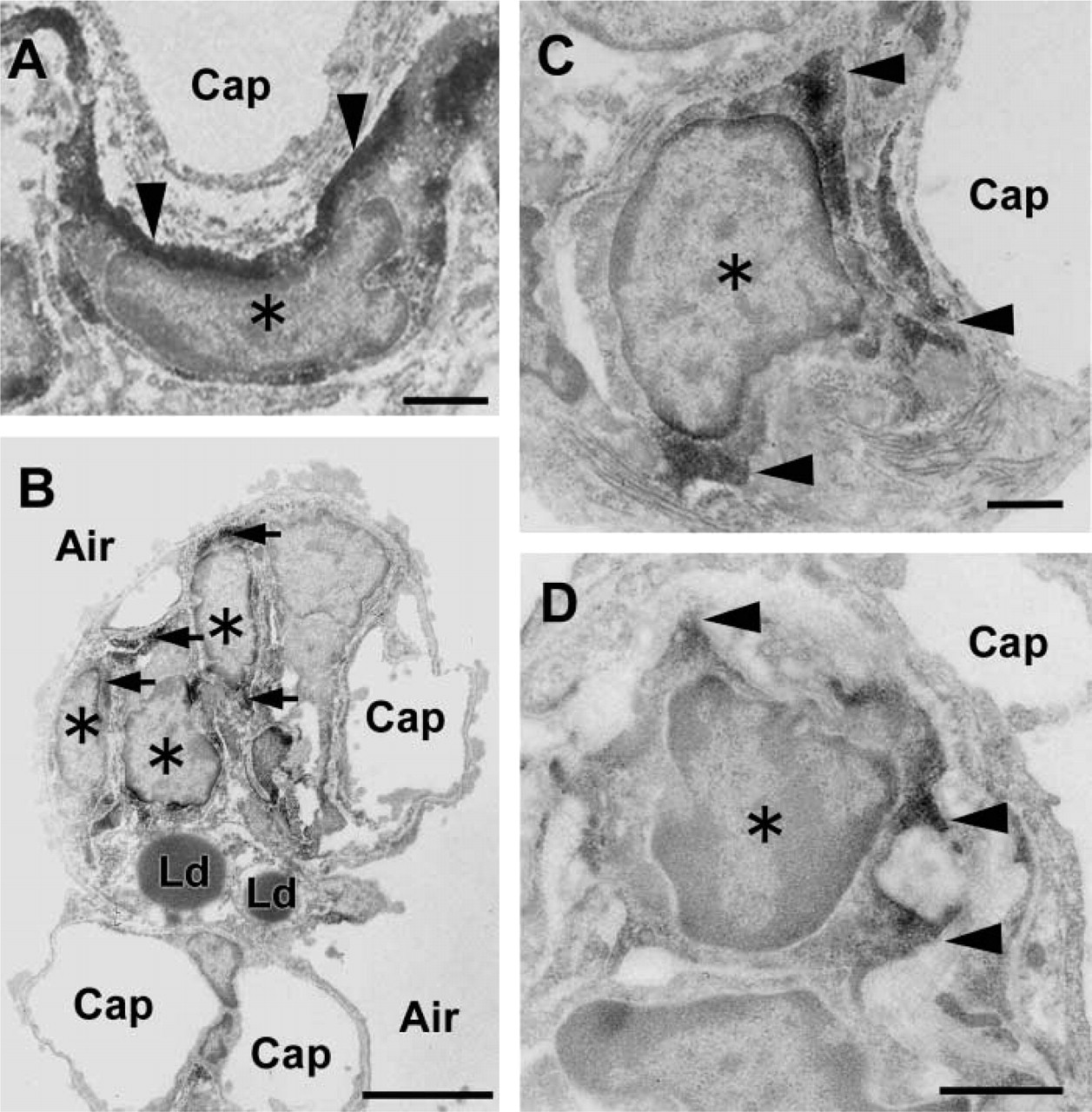
Immunoelectron microscopy of alpha-SMA in the interstitial cell at the tip of the secondary septum. (
The septal myofibroblast-like cells are always surrounded by elastic fibers. The cells elongate their cell processes containing alpha-SMA to the elastic fibers during the septal formation. Because it appears that alpha-SMA–positive fibroblasts have much larger focal adhesions with the substratum than those of alpha-SMA–negative fibroblasts in vitro (Dugina et al. 1998), it is thought that the interaction between the cellular protrusions and elastic fibers is important to make a contractile force. Elastic properties of the lung result from the presence of elastic fibers in the extracellular space. The 72-kDa biosynthetic precursor, tropoelastin, is secreted into the extracellular space where it becomes highly cross-linked into a rubberlike network through the activity of the copper-requiring enzyme lysyl oxidase (Rosenbloom et al. 1993). Myers et al. (1983) have demonstrated that the maximal rates of elastin synthesis are observed in lung explants from 7- to 12-day-old rats, and the rate of elastin synthesis during this period was five to eight times greater than the rate observed in adult rat lung. Furthermore, Noguchi and Samaha (1991) reported that with alveolar septal formation the message for elastin in the interstitium increased progressively from day 17 of gestation, reaching a peak at postnatal days 7 to 11. In general, the signal declined significantly by postnatal day 21, and elastogenesis was virtually absent in the adult. These findings are consistent with our immunocytochemical data for alpha-SMA, suggesting that the expression of alpha-SMA in interstitial cells is closely associated with elastogenesis in the interstitium during septal formation.
It would be interesting to know the fate of alpha-SMA–containing cells (e.g., septal myofibroblast-like cells) at the septal interstitium after lung maturation. We checked the cellular loss of alpha-SMA–containing cells during maturation using the in situ terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling assay, which is useful for detecting apoptotic cells. As a result, we were unable to obtain any evidence on the apoptosis of alpha-SMA–containing cells throughout the whole period. On the contrary, we found cells showing similar ultrastructural characteristics surrounded by elastic fibers located in the septal interstitium in adult rat lung. These results indicate that phenotypical changes in alpha-SMA–positive cells occur after alveolar maturation.
PDGF-A–deficient mice develop lung emphysema secondary to the failure of alveolar septation. Bostrom et al. (1996) reported that these mice lack lung alveolar alpha-SMA-positive cells and exhibit reduced deposition of elastin fibers in the lung parenchyma. The PDGFs appear to regulate the generation of specific populations of myofibroblasts during mammalian development. PDGF-A is crucial for alveolar myofibroblast ontogeny. They also proposed that lung PDGF-R alpha + cells are progenitors of the tropoelastin-positive alveolar SMC, and that postnatal alveogenesis failure in PDGF-A−/- mice is due to a prenatal block in the distal spreading of PDGF-R alpha + cells along the tubular lung epithelium during the canalicular stage of lung development. Our findings also indicate that the alpha-SMA–containing cells are positive against anti-PDGF-A receptor antibody, and that PDGF signaling might be necessary to induce the expression of alpha-SMA and specialized actin filament organization for cell protrusion in which alpha-SMA and drebrin are involved.
Taken together, the unique localization and transient expression of alpha-SMA suggests that the septal myofibroblasts play an important role in septal formation.
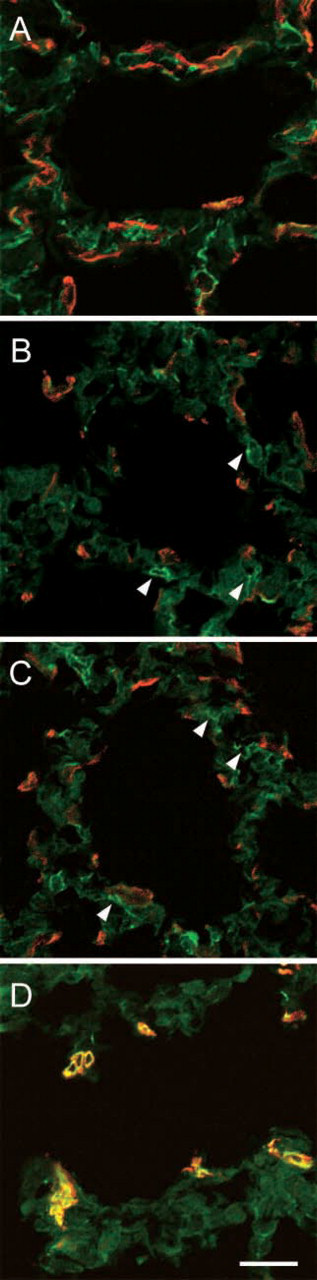
Localization of desmin (FITC) and alpha-SMA (TRITC) in developing rat lung. During the maturation of alveolar septa (
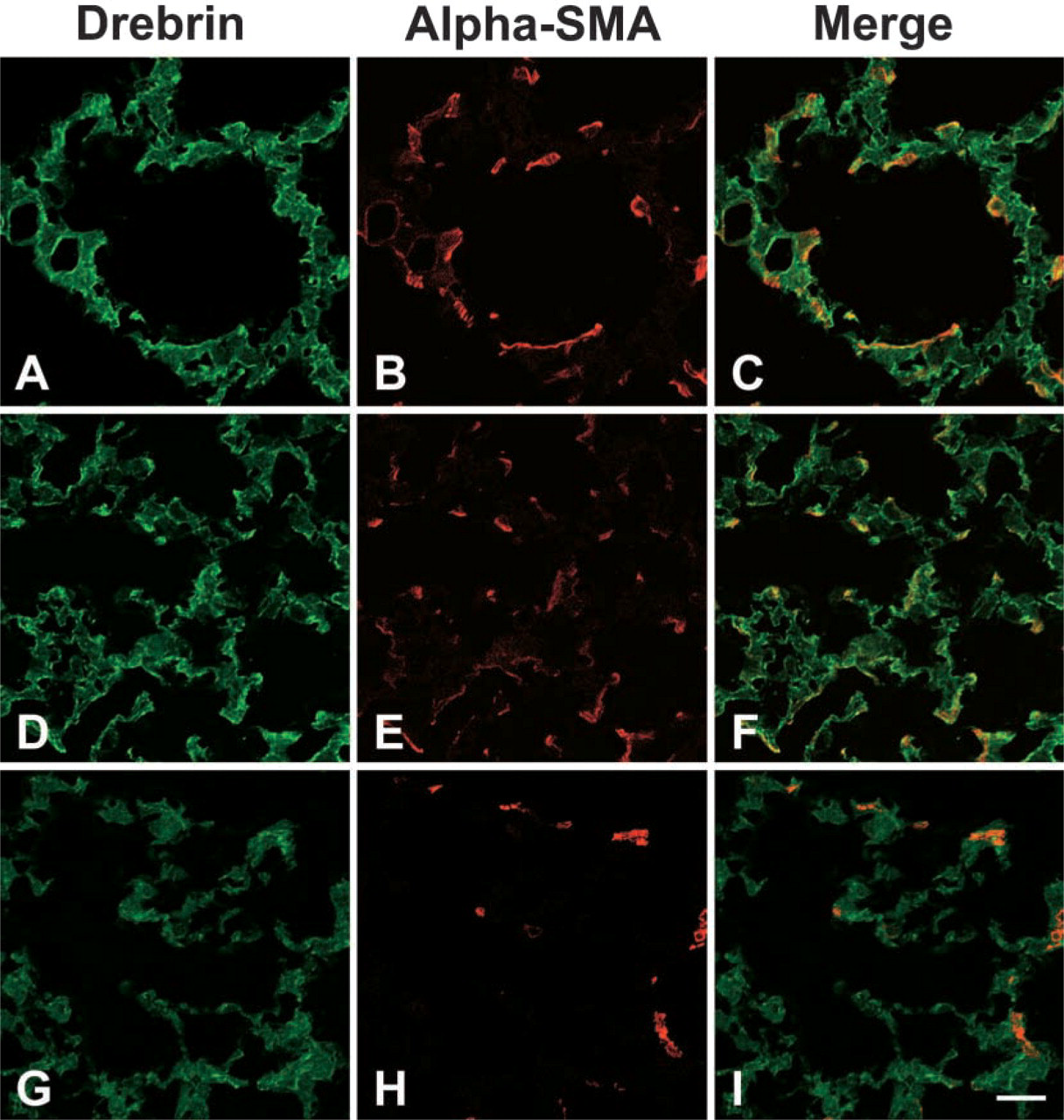
Localization of drebrin and alpha-SMA in developing rat lung. Drebrin is located in cell processes of alpha-SMA-positive cells during the maturation of alveolar septa (