
Abstract
Among the monoclonal antibodies (MAbs) prepared against human sperm extracts, MAb 4F7 was found to be specific to the human and Macaca fascicularis sperm cytoskeletal fibrous sheath (FS). In Western blotting, MAb 4F7 stains a doublet of polypeptides of about Mr 95 × 103 in extracts of human sperm cells. These polypeptides are not recognized by the KL1 anti-cytokeratin MAb, nor by the MAbs known to bind to the carboxy terminal (IFA) and to the amino terminal (ME101) rod domain of intermediate filaments. Sequential extraction procedures shows that the FS polypeptides recognized by MAb 4F7 are exposed after treatment with 8 M urea. 4F7 immunoreactivity is lost after treatment with high ionic solutions (NaCl, KCl, KI). Immunogold electron microscopy reveals that this protein is present throughout the FS. This FS antigenic determinant first accumulates in an FS proximal body in late spermatids, then in granules extending distally along the flagellum. Staining of spermatozoa with flagellar dysgenesis reveals that this FS protein colocalizes with actin no matter what the location of their abnormal assembly. These data suggest that the transient microtubule-like spindle-shaped body of as yet unknown function could be involved in FS protein deposition and that the assembly of the FS and actin could be under the control of some common morphogenetical factor(s). MAb 4F7 should allow further investigations of this periaxonemal structure in both normal and pathological conditions.
IN MOST VERTEBRATES, the flagellar axoneme of spermatozoa is surrounded by unique cytoskeletal structures. Two such structures are the fibrous sheath (FS) and the outer dense fibers of the spermatozoa of reptiles and mammals (Furieri 1970). In mammals, the FS comprises a succession of ribs and two longitudinal columns (Fawcett 1975) which have a filamentous substructure (Olson et al. 1976; Sapsford et al. 1970). In humans, the FS is present on about 88% of the flagellum length and comprises three distinct regions depending on the presence or absence of the columns (Serres et al. 1983). The FS could play a role in the modulation of the flagellar curvature in species with internal fertilization (Phillips 1972).
FS biogenesis occurs in late spermiogenesis (Oko and Clermont 1989; Fenderson et al. 1988; Sakai et al. 1986; Irons and Clermont 1982). In rodents, FS assembly begins with the formation of a framework, the definitive FS being formed by deposition of material onto the framework (Irons and Clermont 1982). In humans, a flagellar tubulofilamentous complex of as yet undefined function is transiently observed during FS assembly (Wartenberg and Holstein 1975). Simultaneous anomalies of the axoneme and periaxoneme suggest that the FS spatial organization is controlled by its association with the axoneme (Escalier and Serres 1985) and in relation to distinct tubulin domains (Gallo et al. 1986). In mice, flagellar anomalies suggest that the axoneme is required for FS organization (Phillips et al. 1993).
Both human (Kim et al. 1995; Jassim 1991, 1994; Jassim and Chen 1994; Beecher et al. 1993) and rodent (Brito et al. 1989; Fenderson et al. 1988; Oko 1988; O'Brien and Bellvé 1980; Olson et al. 1976) FS contain many polypeptides ranging from Mr 11 × 103 to Mr 120 × 103. Crossreactions of anti-rat FS antibody with the outer dense fibers reveals common antigenic determinants between these flagellar cytoskeletal structures (Tres and Kierszenbaum 1996; Oko 1988). Amino acid analysis has shown that the rat FS is characterized by a high content of aspartic acid and glutamic acid (Kim et al. 1995) and of phosphoserine, the latter suggesting that the FS contains phosphoprotein (Brito et al. 1989). Human FS phosphorylation (Pariset and Weinman 1994; Jassim et al. 1991) and glycosylation (Jassim and Bottazzo 1994) are also found. In the rat, the Mr 80 × 103 (major) and 120 × 103 (minor) FS proteins bind the regulatory subunit (RII) of a Type II cAMP-dependent protein kinase (A-kinase) (MacLeod et al. 1994) and the prominent mouse Mr 82 × 103 FS protein shows homology to A-kinase anchoring protein (Carrera et al. 1994). FS association with a distinct isoform of Type II cAMP-dependent protein kinase has been found in human (Pariset and Weinman 1994).
Disulfide content and resistance to solubilization have long suggested that the FS may be related to proteins of the intermediate filament (IF) family (Calvin 1975), and an antibody directed against a keratin-like protein of the dense fibers has recently been found to label the FS longitudinal columns (Tres and Kierszenbaum 1996). However, few antibodies directed against proteins of several classes of somatic IFs recognize the FS in sperm cells (Eddy et al. 1991; Longo et al. 1987; Franke et al. 1979), including an anti-neurofilament antibody (Jassim 1991; Jassim et al. 1991). Moreover, characterization of the Fsc1 cDNA, possibly coding for the mouse Mr 78–80 × 103 major FS polypeptide, reveals no significant homology with other known cytoskeletal proteins, except for a region with >44% identity with a stretch of a centromeric protein-E (CENP-E) (Fulcher et al. 1995). FSC1 protein exhibits potential phosphorylation sites and is rich in cysteinyl residues, suggesting extensive posttranslational modifications of the protein (Fulcher et al. 1995).
Data on the mouse FS therefore reveal that spermatogenic cells contain unique cytoskeletal protein(s) (Fulcher et al. 1995). In humans, cDNA for FS proteins has not yet been found and the FS assembly processes are still unknown, although several types of FS anomalies lead to male infertility (for review see Escalier 1985). In this study, a monoclonal antibody, MAb 4F7, elicited against human sperm cell extracts, was shown to specifically recognize the FS of monkey and human spermatozoa. MAb 4F7 recognizes an FS polypeptide doublet of about Mr 95 × 103, which is resistant to urea extraction and which is not recognized by an antibody directed against pan-cytokeratins and two anti-IF antibodies directed against the consensus sequence on the IF coil. The biochemical characterization, ultrastructural location, and biogenesis of this FS component have been studied. Analysis of pathological models of FS dysgenesis reveals actin and FS co-localization, even if they fail to surround the axoneme. MAb 4F7 should be useful to further investigate the events of human spermiogenesis and their failures in clinical conditions.
Materials and Methods
Production of Monoclonal Antibodies
Balb/c mice were immunized with the insoluble fraction from homogenates of human spermatozoa performed in 0.1 M HEPES, 1 mM MgSO4, 2 mM EGTA, 2 M glycerol, pH 6.9, and 1% Nodinet P-40 (Gallo and Schrével 1985). Hybridomas were produced by fusion of spleen cells with the myeloma cell line × 63 Ag 8–653 and culture supernatants were screened by indirect immunofluorescence as previously described (Gallo et al. 1991). Antibody isotyping was performed by means of an ImmunoType kit (Sigma; St. Louis, MO).
Sperm Samples and Testicular Biopsies
Human semen samples were collected after 3 days of sexual abstinence. They were from eight donors whose sperm parameters were considered normal according to standard criteria of semen analysis and from four infertile patients with spermatozoa exhibiting FS dysgenesis whose characteristics were defined by electron microscopy as previously described (Escalier and David 1984). Motile human spermatozoa from donors were selected by swim-up migration into B2 medium (Menezo B2 medium; BioMerieux, Lyon, France) for 1 hr at 37C. Spermatozoa were harvested by centrifugation (800 × g, 10 min) and washed in PBS.
Testis biopsies were obtained from two men with obstructive azoospermia and a normal germinal epithelium as revealed by light microscopy. Spermatozoa from rodent species (rat, mouse, hamster, and guinea pig) and from the monkey Macaca fascicularis were obtained by perfusion of caudae epididymes with PBS at pH 7.2. Spermatozoa from horse, bull, and boar were obtained by sexual stimulation, then washed in PBS.
Sperm Treatments for Indirect Immunofluorescence
Washed motile sperm cells were air-dried onto microscope slides. Some samples were directly immunolabeled, and others were treated with the following procedures before immunostaining: (a) fixation with acetone or methanol or 5% acetic acid in ethanol at −20C for 7 min; (b) permeabilization with 0.1% (w/v) Triton X-100 in PBS at 4C for 10 min; (c) incubation in PBS containing 2 M NaCl or 2 M KCl or 1 M potassium iodide or 3 M guanidine hydrochloride (grade I; Sigma) or 6 M urea (Bio-Rad Laboratories; Richmond, CA) at 4C for 1 hr; and (d) three-step FS extractions at 4C (see below). FS extraction on slides was performed according to Fenderson et al. (1988) from the method of FS isolation in suspension of Olson et al. (1976). After each treatment, cells were washed with PBS and processed for immunofluorescence microscopy.
Sperm cells from infertile patients were air-dried onto slides and treated with 0.1% (w/v) Triton X-100 in PBS for 10 min, then washed with PBS. A sample of each biopsy was Bouin-fixed and paraffin-embedded and another was frozen as previously described (Escalier et al. 1991). Testis sections were treated, or not, with 0.1% Triton X-100 or 3% paraformaldehyde and washed with PBS before immunolabeling.
Indirect Immunostaining
Sperm samples processed as above were preincubated in PBS containing 1% (w/v) BSA (grade V; Sigma) for 15 min and then with MAb 4F7 (undiluted culture supernatant) for 45 min at room temperature (RT). After extensive washing in PBS, fluorescein isothiocyanate-conjugated goat anti-mouse immunoglobulins (GAM/FITC; Nordic, Tilburg, The Netherlands) diluted 1:35 were added for another 45 min. After extensive washing in PBS, cells were fixed with 3% formaldehyde in PBS. Slides were counterstained with 0.02% Evans blue and mounted in Citifluor–PBS–glycerol solution (Citifluor; London, UK). Samples from infertile men were also labeled with an antiactin MAb (N 350; Amersham, Poole, UK) diluted 1:100 in PBS/BSA. The secondary antibody was an anti-mouse IgM (Biosys SA; Compiègne, France) diluted 1:40 in PBS. Testis sections were labeled by the three-step immunoperoxidase technique using the biotin–avidin system (Vectastain ABC kit PK-4002; Vector Laboratories, Burlingame, CA) as previously described (Escalier et al. 1991, 1992). Sections were incubated for 1 hr with MAb 4F7 (undiluted culture supernatant). Nonspecific antibody binding was blocked by 5% BSA in PBS. Controls were performed using unrelated MAb 4D4, an anti-proacrosin MAb produced in the same fusion as MAb 4F7 (Gallo et al. 1991) and Mab 1B41 directed against β-tubulin of Trypanosoma b. brucei (Gallo et al. 1988), which does not recognize human sperm proteins. These antibodies were used as undiluted culture supernatants and ascites fluid diluted 1:200 (4D4).
Immunoelectron microscopy
Before pre-embedding immunolabeling, pellets of sperm cells from donors were treated by one of the three following methods: (a) one cycle of freezing-thawing in liquid nitrogen; (b) 0.05% (w/v) Triton X-100 in PBS for 10 min; and (c) 0.1 % (w/v) Triton X-100 in PBS for 15 min, then 6 M urea and 100 μg/ml DNAse I (grade II; Boehringer, Mannheim, Germany) in PBS for 2 hr. In this latter method, a cocktail of protease inhibitors was added to the incubating solutions (see below). After each treatment, sperm cells were washed in PBS. Pre-embedding immunolabelings were performed as previously reported (Gallo et al. 1991). Briefly, cells were incubated for 3 hr at RT with MAb 4F7 (undiluted culture supernatant), then for 1 hr with a colloidal gold-labeled antibody to mouse IgGs (Auroprobe EM GAM IgG G10; Amersham) diluted 1:20 in PBS/BSA. Then cells were processed for electron microscopy. Controls were performed either by using unrelated MAbs (4D4, 1B41) or by omission of MAb 4F7 incubation.
Sperm Extraction Procedures for Immunoblotting
Extraction of sperm proteins from motile spermatozoa was performed at 4C after several methods of sequential extraction. Protease inhibitors were added: 2 mM phenyl-methyl-sulfonyl-fluoride (PMSF), 10 mM aprotinin, 5 mM soybean trypsin inhibitor (Type IS), 1 mM benzamidine hydrochloride, 7 mM α2 macroglobulin (Sigma). In addition, 100 μg/ml DNAse I (grade II, Boehringer) was also present in solutions. The first method for sperm cell extraction was the three-step method of FS isolation according to Olson et al. (1976): (a) 50 mM sodium borate and 1% (w/v) Triton X-100 for 1 hr; (b) 0.6 M potassium thiocyanate (Sigma) for 2 hr; and (c) 4 M urea for 16 hr. These three solutions contained 2 mM dithiothreitol (Bio-Rad) and the incubations were performed at 4C. The second method was a two-step ionic strength extraction method. In the first step, incubation was performed in solutions containing 10 mM NaCl, 5 mM EDTA, 10 mM Tris-HCl, pH 7.4, and 1% (w/v) Triton X-100 for 30 min and in the second step in 1.5 M KCl, 0.5% (w/v) Triton X-100 for 1 hr. The third method was used by Aynardi et al. (1984) to extract IFs: (a) the lysis buffer (pH 8.0) contained 0.6 M KCl, 10 mM MgCl2, 1% Triton X-100 (w/v), and 10 mM Tris-HCl (pH 7.4) and (b) the solubilization buffer (pH 8.0) contained 8 M urea and 10 mM Tris-HCl (pH 7.4). Single-step methods were also tried using the following solutions: (a) 6 M urea, 2 mM DTT, 50 mM Tris-HCl (pH 8.0) for 1 hr and (b) 8 M urea, 1% (w/v) Triton X-100, Tris-HCl 10 mM for 3 hr. After each incubation, extracts were centrifuged at 12, 000 × g for 2 min and the pellets were washed in the buffer of the preceding extraction solution and centrifuged again. Supernatants resulting from all treatments and pellets were then boiled in SDS sample buffer (Laemmli 1970). Pellets of untreated spermatozoa were directly boiled in SDS sample buffer.
Electrophoresis and Immunoblotting
Samples were electrophoresed in 8.5% (w/v) polyacrylamide gels according to Laemmli (1970) and electroblotted onto nitrocellulose according to Towbin et al. (1979). Nitrocellulose sheets were incubated with a 1:5 dilution of 4F7 culture supernatant as primary antibody. The following monoclonal antibodies were also used: (a) anti-cytokeratin (Pan, KL1; diluted 1:200) (Immunotech; Marseille, France); (b) anti-intermediate filament IFA (IgG1; culture supernatant diluted 1:15) (Pruss et al. 1981) and ME101 (IgM; ascites fluid diluted 1:100) (Escurat et al. 1989). Secondary antibody was a goat anti-mouse IgG (or IgM for ME101) conjugated to peroxidase (Biosys SA). Antibody binding was detected by the immunoperoxidase method using diaminobenzidine as a chromogen, as previously described (Escalier et al. 1991). Controls were performed using mouse IgG (IgM for ME101).
Results
MAb 4F7 Recognizes a Flagellar Component of Primate Spermatozoa
The hybridoma MAb 4F7 secreted an Ig (belonging to the IgG1 class) that stained the flagellar principal piece of permeabilized human spermatozoa (Figure 1). Unrelated 4D4 and 1B41 MAbs did not reveal any staining of the flagellum. Unfixed and unpermeabilized spermatozoa remained unstained, indicating that MAb 4F7 did not react with the cell surface but with an intracellular antigen. MAb 4F7 did not stain the blood cells and desquamated epithelial cells present in some ejaculates. Permeabilized spermatozoa from M. fascicularis were stained with MAb 4F7 in a pattern similar to that observed for human spermatozoa. By contrast, spermatozoa from horse, boar, bull, rat, mouse, hamster, and guinea pig were not stained with MAb 4F7, regardless of the chemical treatment used before labeling (see Materials and Methods) (data not shown).
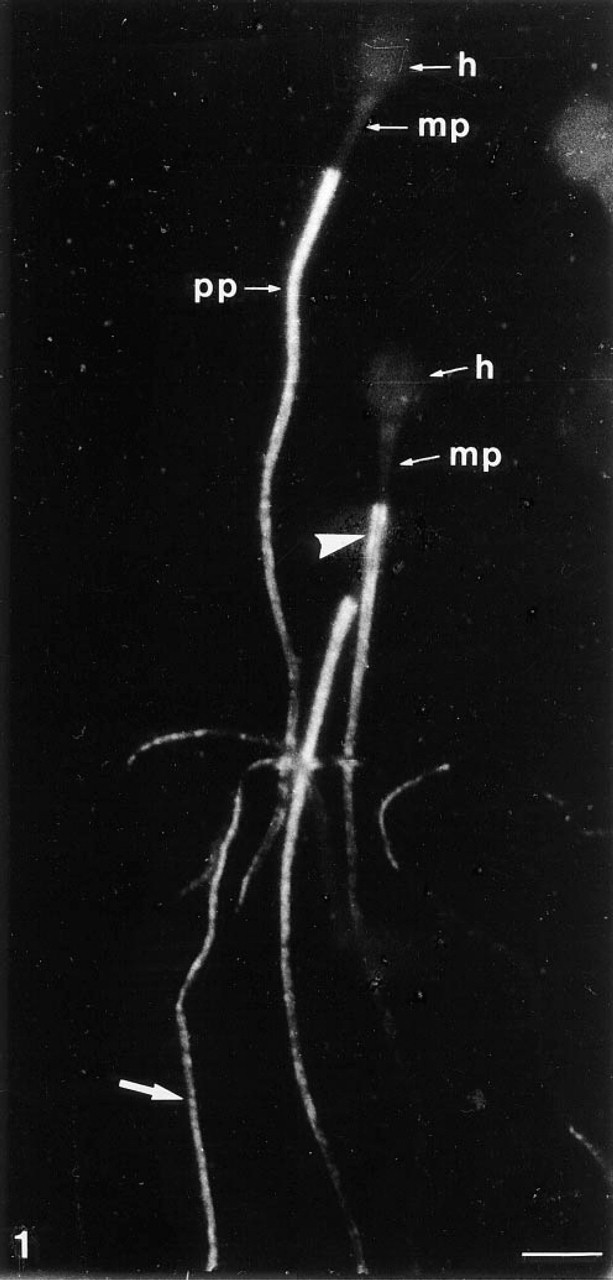
MAb 4F7 labeling pattern of human spermatozoa. Indirect immunofluorescence after cell permeabilization (5% acid acetic in ethanol). The staining is located only on the flagellar principal piece (pp) and surrounds an unstained inner flagellar compartment (arrowhead). The labeling exhibits a transverse striated pattern (arrow). h, sperm head; mp, flagellar middle piece. Bar = 5 μm.
MAb 4F7 Recognizes an M r 95 × 103 Polypeptide Doublet
From sperm polypeptides separated by electrophoresis, a band at about Mr 95 × 103 could be observed in the FS-containing fraction isolated after treatment with 8 M urea and 1% Triton X-100 (Figure 2A, Lane 1). This band was absent in the soluble sperm fraction (Figure 2A, Lane 2) and was poorly distinguishable in the whole sperm extract (Figure 2A, Lane 3). In Western blotting, this Mr 95 × 103 doublet was also present in the pellet obtained after treatment with 8 M urea and 1% Triton X-100 (Figure 2B, Lane 1). Total sperm cell extracts directly boiled in SDS sample buffer also showed that MAb 4F7 reacted with a polypeptide doublet of about Mr 95 × 103 (Figure 2B, Lane 3). Pellets obtained after treatment with 6 M urea and 2 mM DTT, or after the IF extraction method, exhibited the labeled Mr 95 × 103 doublet and an additional fainter doublet of about Mr 82 × 103 (Figure 2B, Lane 4). A similar result was obtained after the three-step extraction method (data not shown). Soluble fractions obtained after each of these various treatments did not exhibit MAb 4F7 labeling except for small amounts after potassium thiocyanate treatment for the three-step extraction method (data not shown). By contrast, the insoluble fraction obtained by the two-step method of extraction with 1.5 M KCl (Step 2) showed predominantly the lower band of the Mr 95 × 103 doublet and also several labeled bands at about Mr 82 × 103, 74 × 103, and 45 × 103, and minor bands at Mr 68 × 103, 59 × 103 and 49 × 103 (Figure 2B, Lane 5). Increasing KCl concentration to 2 M led to very faint or no labeling (data not shown). Together, these data suggest that the additional bands corresponded to degraded products of the FS polypeptide studied.
4F7-Related Polypeptides Are Not Recognized by Anti-intermediate Filament Antibodies
Known crossreaction of FS proteins with IF antibodies (Eddy et al. 1991; Jassim 1991) and resistance to prolonged urea treatment of the 4F7-related polypeptides led us to investigate whether these latter could be related to IF proteins. To investigate the presence of IF epitopes on FS proteins, several anti-IF antibodies were used in Western blotting on sperm extracts (Figure 3). The antibody KL1 recognizes Types I and II cytokeratins (pan-cytokeratin) in somatic tissues (Viac et al. 1983). The IFA recognizes the consensus sequence common to every IF protein and located at the car-boxy terminal end of the rod domain (Pruss et al. 1981). The antibody ME101 recognizes the amino terminal half containing the consensus sequence located on IF coil 1a of all the major classes of vertebrate IF proteins (Escurat et al. 1989). FS polypeptides recognized by MAb 4F7 were not recognized by these three anti-IF antibodies. However, other unidentified sperm proteins reacted with these antibodies (Figures 3B–3B). The pattern of the proteins recognized by the anti-IFs differed according to the sperm protein extraction procedures used. Controls were negative (data not shown).
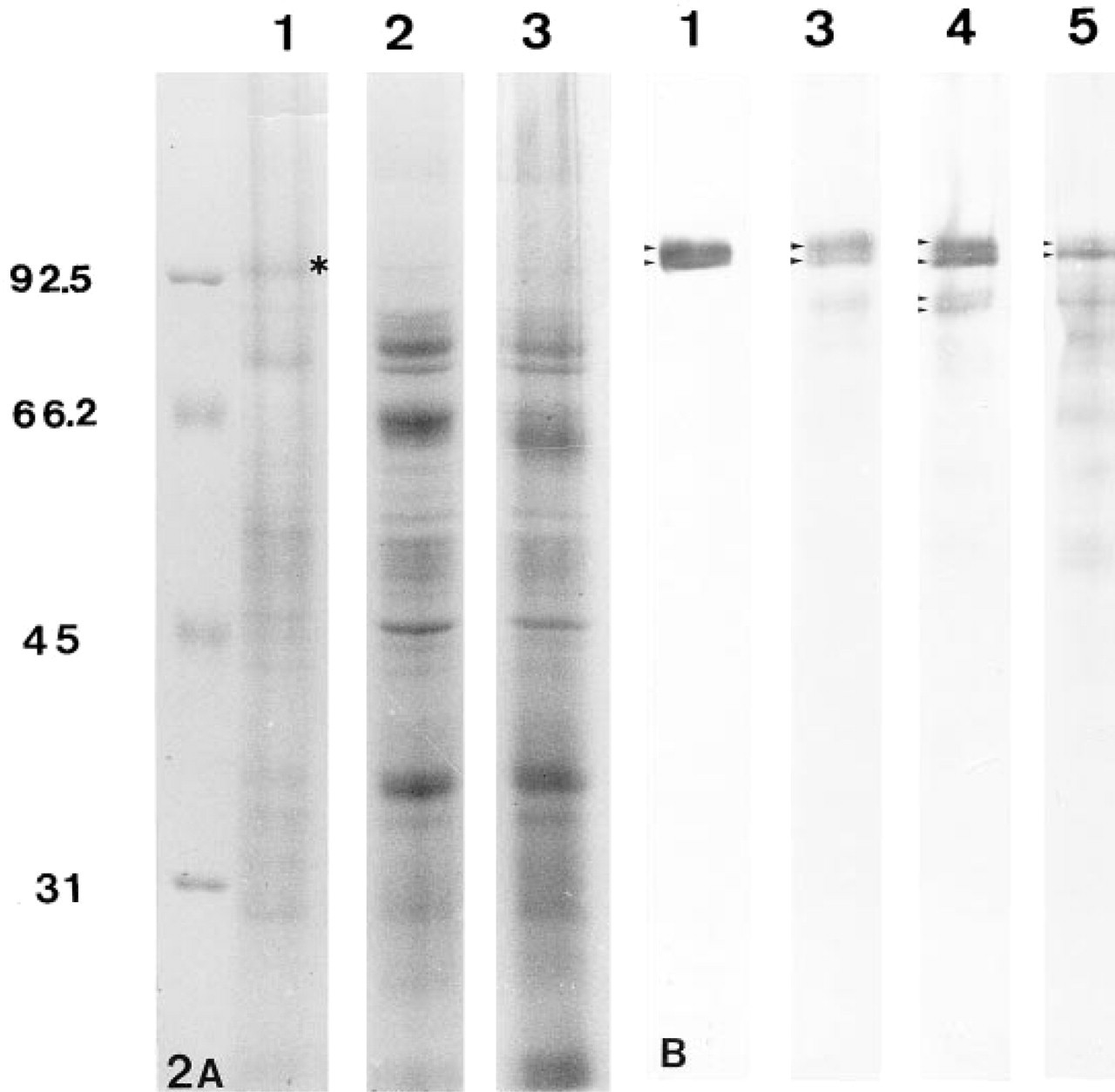
Identification of the 4F7-immunoreactive polypeptide in human spermatozoa.
The 4F7-immunoreactive Protein Is Resistant to Denaturing Solutions but Not to High Ionic Strength Treatments
Characterization of the somatic IF after biochemical treatments has established that IFs are resistant to high-salt buffers (see review in Steinert et al. 1982). The failure of the IF antibodies used to recognize the 4F7-immunoreactive polypeptides in Western blotting (Figure 3) led us to investigate some other biochemical characteristics of these FS polypeptides by 4F7 labeling of sperm cells treated on slides.
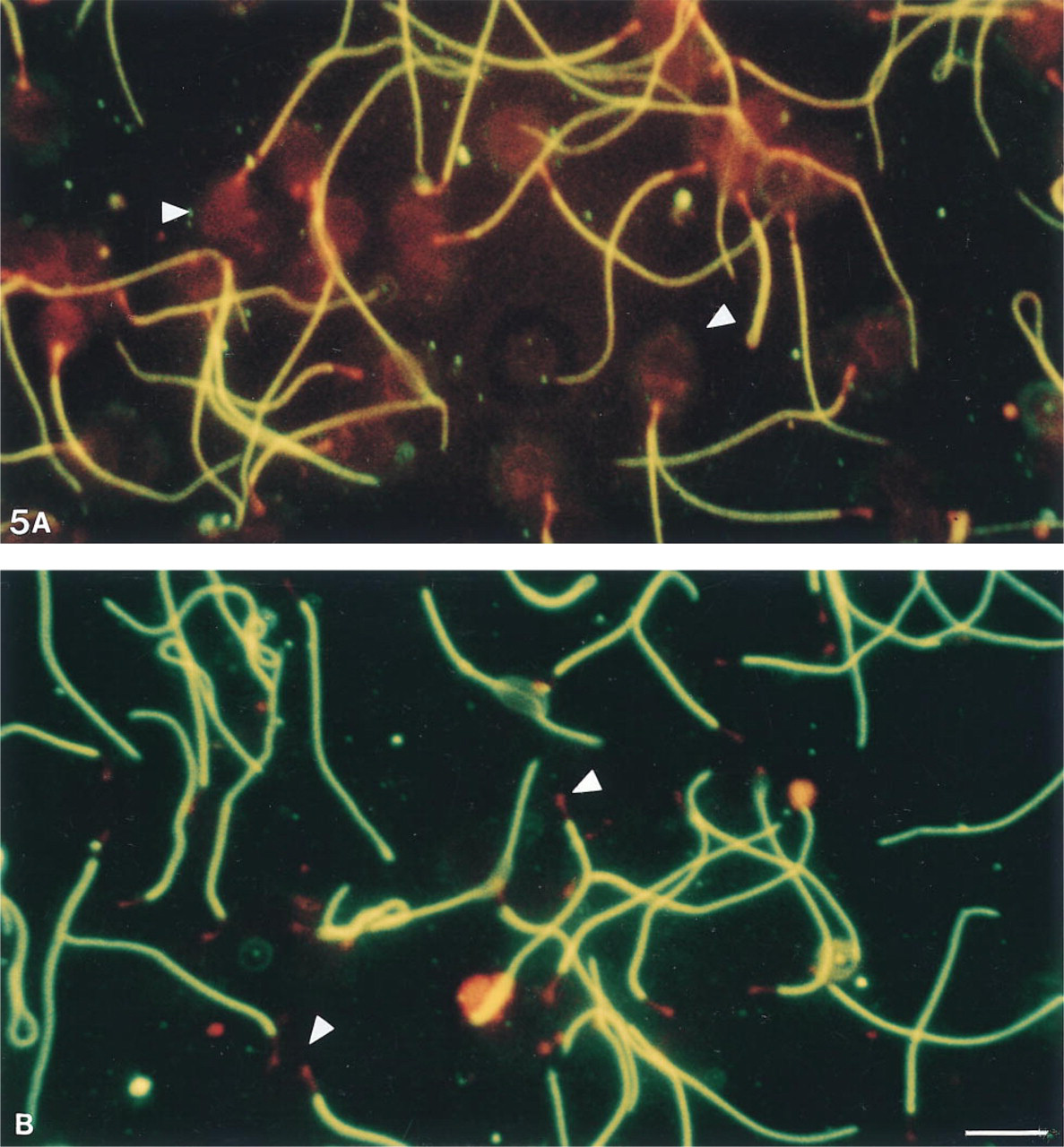
Moderate and variable 4F7 labeling was obtained with air-dried spermatozoa permeabilized by acetone or methanol. The 4F7 labeling was increased after treatment with 5% acetic acid in ethanol (Figure 1). Very intense labeling was found after treatment with 6 M urea (Figure 4A) or 3 M guanidine (Figure 4B) or with 0.1% Triton X-100 alone. Spermatozoa were poorly stained after treatment with 2 M KCl (Figure 4C) or 1 M potassium iodide, and they lost their staining after 2 M NaCl treatment (Figure 4D). Immunolabeling with MAb 4F7 after the three-step FS extraction method (Figure 5) showed that sperm cells were (a) weakly stained after 50 mM sodium borate incubation (Step 1), (b) intensely stained after 0.6 M potassium thiocyanate incubation (Step 2) (Figure 5A), and (c) strongly stained after 4 M urea treatment (Step 3) (Figure 5B). For all the above treatments, no staining was observed in regions of the sperm cells other than the principal piece of the flagellum.
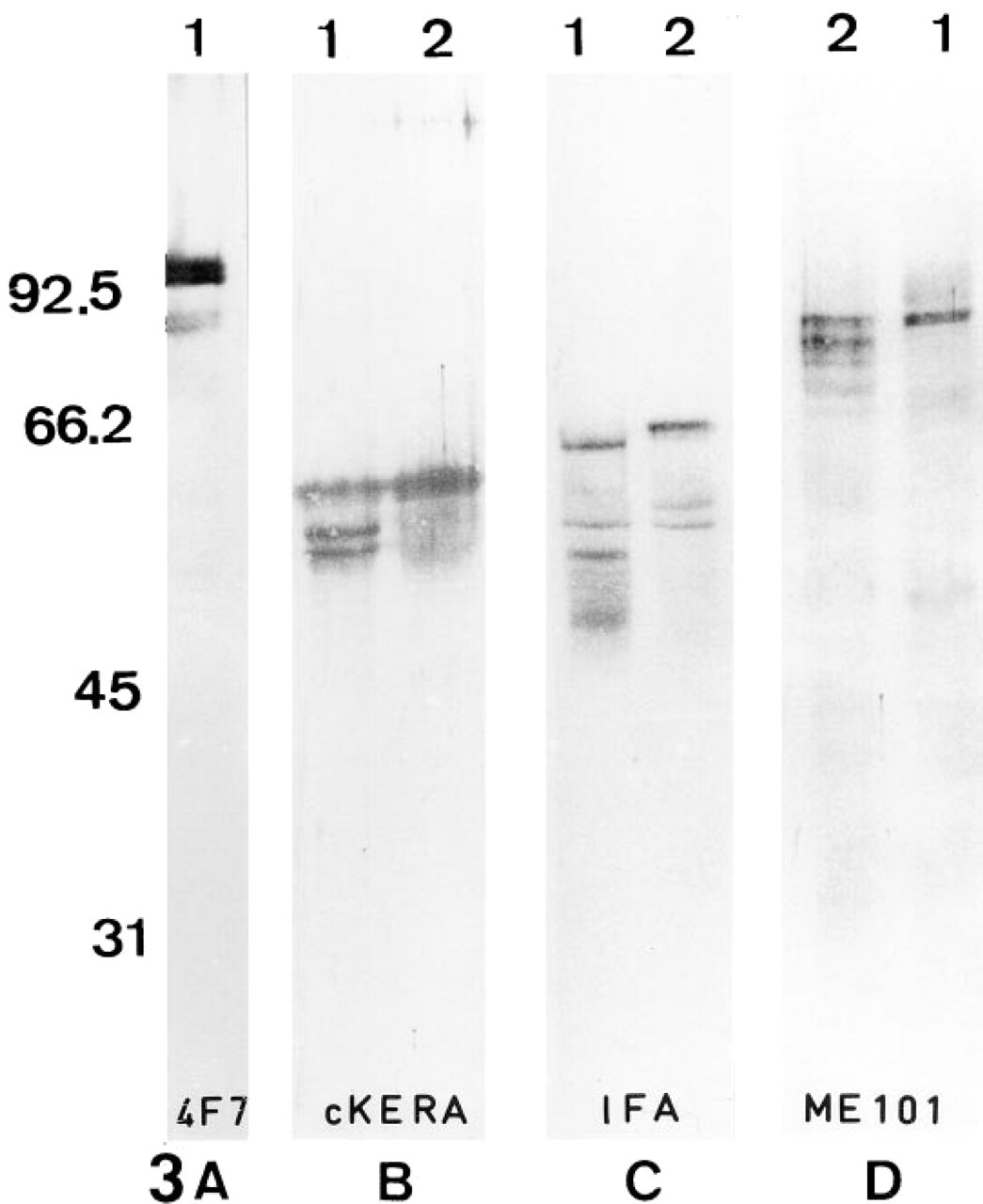
The FS polypeptides recognized by MAb 4F7 do not react with anti-intermediate filament antibodies.
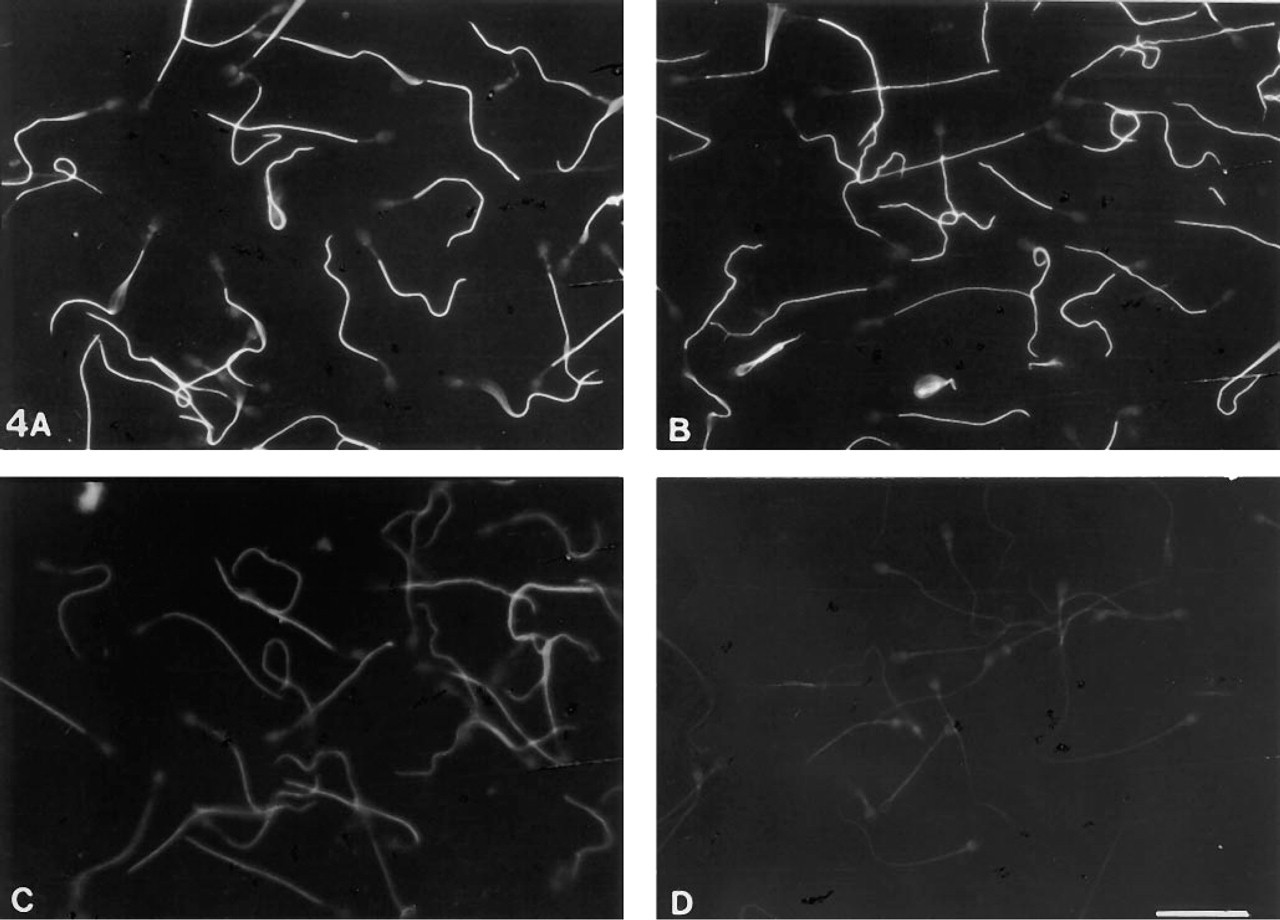
Effects of dissociating agents and ionic solutions on the epitope recognized by MAb 4F7 in human sperm fibrous sheath. Detection was performed by indirect immunofluorescence. MAb 4F7 binding to the fibrous sheath is preserved and intense after sperm treatment with urea 6 M
MAb 4F7 Binds Specifically to the Flagellar Fibrous Sheath
Immunogold labeling performed on pre-embedded human sperm samples showed that the epitope recognized by MAb 4F7 was located on the FS (Figure 6A). The other flagellar components such as mitochondria, the connecting piece, the outer dense fibers, and the axoneme were unstained by MAb 4F7. FS isolated by 6 M urea showed many gold particles on both the ribs and columns of the FS (Figure 6B) and on outer and inner FS surfaces (Figure 6C). This staining restricted to the FS surfaces could be due to the tight packing of FS filaments, forming a barrier to the gold particles. Indeed, sections exhibiting FS packaging alteration due to prolonged urea treatment showed that the 4F7-related polypeptides are structural components of the FS (Figure 6D). Controls omitting MAb 4F7 did not show any staining. Samples permeabilized by either Triton X-100 or freeze-thawing showed very sparse labeling (data not shown).
Sequential Expression of 4F7-Immunoreactive Protein During Fibrous Sheath Biogenesis
4F7 labeling was obtained in frozen sections from testis biopsies. The avidin–biotin–peroxidase labeling procedure revealed different labeling patterns and allowed visualization of the cell maturation steps (Figure 7).
Only spermatids with a condensed nucleus and an acrosomal cap were labeled with MAb 4F7 at the flagellar level, whereas the flagellar spermatid cytoplasm was free of labeling. The labeling was restricted to the principal piece (Figure 7A). Several labeling patterns were observed in distinct epithelial seminiferous regions: (a) an intense staining of the proximal part of the principal piece and, in addition, enlarged staining beneath the middle piece whose location and size corresponded to the spindle-shaped body (0.8 μm wide vs 0.4 μm for the FS) (Figure 7B); (b) staining extending over the principal piece but with either a granular (Figures 7C and 7D) or a striated pattern (Figures 7E and 7F); and (c) more homogeneous staining extending to the whole flagellar principal piece (Figure 7A). Staining of only the distal principal region of the flagella (i.e., of the flagellar part emerging into the tubule lumen) was never seen, indicating that human FS assembly occurs first in the proximal flagellar region.
Unmasking of the MAb 4F7-related epitope after human sperm fibrous sheath isolation by the three-step method on slides (according to Fenderson et al. 1988; Olson et al. 1976). Indirect fluorescence using MAb 4F7 (green) and counterstaining of the sperm structures with Evans blue (red).
Comparative Distribution of the 4F7-Immunoreactive Protein and Actin
The N 350 anti-actin MAb has revealed the presence of actin in FS in normal human spermatozoa (Flaherty et al. 1988), but the role of actin in FS is unknown. In somatic cells, actin associates with other cytoskeletal proteins (see review in Carmo-Fonseca and David-Ferreira 1990). The strategy using pathological models was chosen to investigate whether the flagellar cytoskeletal component recognized by MAb 4F7 could have some relationship with actin, particularly in human spermatozoa with morphogenetic anomalies of the FS. Three cases with defective FS were selected by electron microscopy: one case with a reduced FS and two cases in which the FS failed to assemble around the axoneme and was occasionally found in residual bodies (Escalier 1985; Pedersen et al. 1971) (data not shown). In the first case, the labeling with both MAb 4F7 (Figure 8A) and anti-actin MAb (Figure 8B) was either absent or was present in only 4–10 μm of the principal piece (vs 50 μm in normal flagella; see Figure 1). In the two other cases, intense 4F7 and actin staining was found in isolated cytoplasmic bodies, whereas the principal pieces were unstained with MAb 4F7 and poorly stained with the anti-actin MAb (Figures 8C–8F). This residual actin may be related to the known presence of actin in the axoneme (Sugase et al. 1996). Therefore, these pathological models of FS dysgenesis showed that the FS component studied and the FS-associated actin were affected in their biogenesis in a similar pattern.
Discussion
The MAb 4F7 selected among MAbs directed against human sperm extracts recognizes an Mr 95 × 103 polypeptide doublet in the human FS. This FS component is insoluble in urea but is degraded by high ionic strength treatments. MAb 4F7 crossreacts with the sperm flagella of M. fascicularis but not in many other mammal species. A failure of cross-species reactivity of FS antibodies has been frequently observed, even among rodent species (Fenderson et al. 1988).
Until now, immuno-analysis of the FS has been studied mainly in rodents (Eddy et al. 1991; Clermont et al. 1990; Oko and Clermont 1989; Fenderson et al. 1988; Oko 1988). The GDA-J/F3 MAb (Jassim et al. 1990) and the FS K 32 MAb (Sakai et al. 1986; Koyama et al. 1984; Jones et al. 1983) recognize proteins of the human FS but are not reactive in Western blotting. MAbs S69 and S70 recognize human 68-kD and 54–51-kD major FS polypeptides (Beecher et al. 1993) and AJ-FS1 multiple bands ranging between 97 and 28 kD (Jassim and Chen 1994). Unlike MAb 4F7, AJ-FS1 (Jassim and Chen 1994) and S69 (Beecher et al. 1993; Homyk and Herr 1992) bind only to the outer side of the FS. The RT97 anti-neurofilament antibody recognizes a single 97-kD peptide of the human FS, showing that the human FS shares an epitope common to anti-200-kD neurofilament (Jassim et al. 1991, 1992; Jassim 1991). MAb 4F7, which was produced from human spermatozoa, does not react with various seminiferous components and germ cell nuclei as was observed with the RT97 MAb (Jassim 1991).
Western blotting reveals that MAb 4F7 labels two bands around Mr 95 × 103 in human FS extracts. These two polypeptides are found after various conditions of sperm extraction. Because the two bands are of equal intensity, the 4F7-labeled polypeptides should correspond to a dimer whose peptide chain monomers were separated. The presence of a dimeric protein in the FS has already been suggested for the mouse (Fenderson et al. 1988). Another possibility would be the presence of isoforms. It has already been discussed whether the six or more peptides found to be stained by anti-FS antibodies in rodents are degraded FS protein products or represent a family of related proteins (Fenderson et al. 1988; Oko 1988). Data from this study show that FS polypeptides have a propensity to be reduced in a cascade of several degradation products.
The MAb 4F7 epitope is accessible only after treatment with detergents or dissociating solutions, such as urea or guanidine. The recognized FS protein may be associated with other flagellar proteins, such as those of the cross-filaments of the cytoplasmic matrix which link the FS to the other flagellar components (Escalier 1984). The presence of masked antigens in the human FS has already been observed (Jassim 1994). Another possibility would be that the inacessibility of the antigenic determinant is the result of the three-dimensional configuration of the protein in situ.
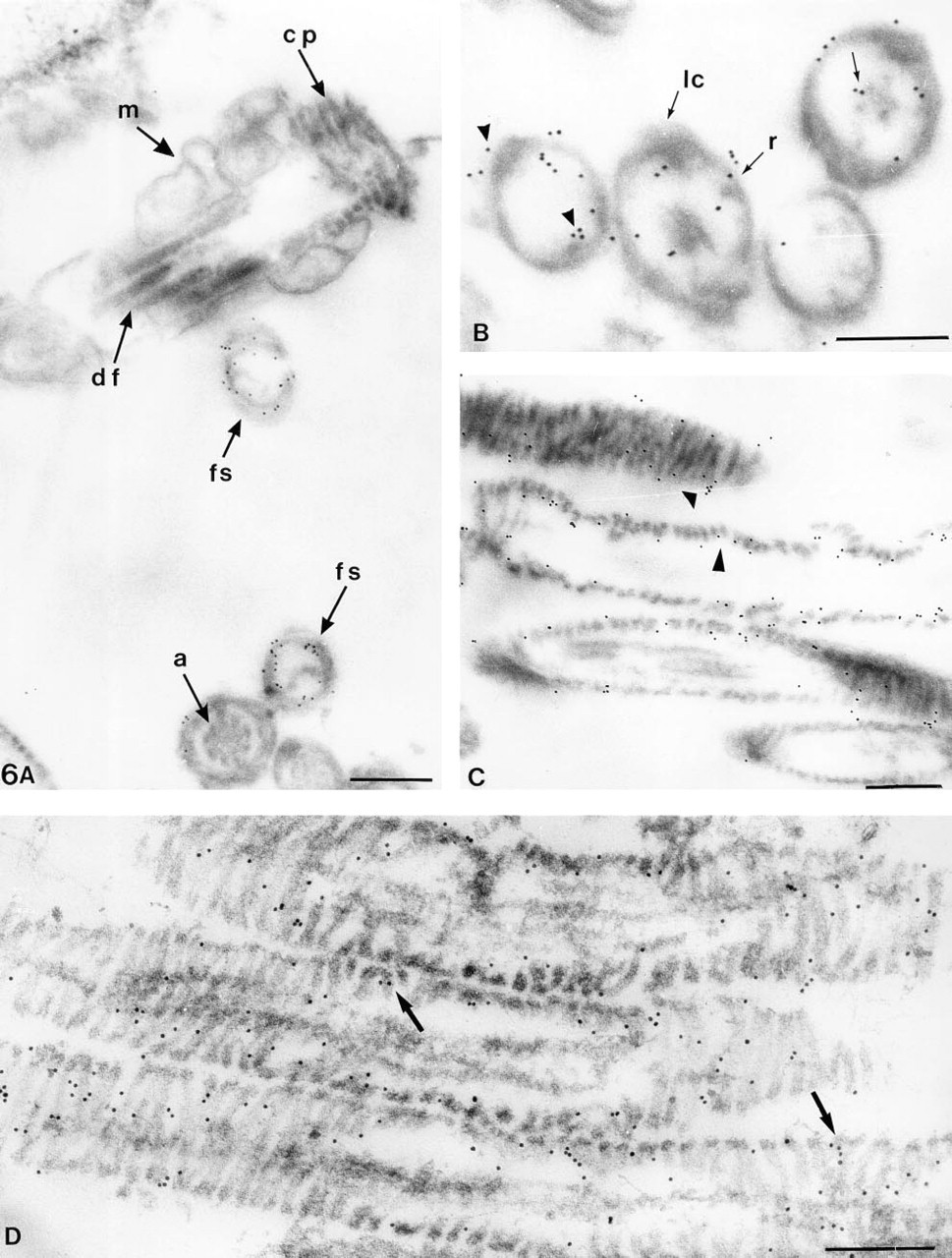
Ultrastructural immunogold localization of the MAb 4F7 epitope (pre-embedding immunogold labeling of human spermatozoa after treatment with 0.1% Triton X-100 and 6 M urea).
The FS polypeptides recognized by MAb 4F7 do not crossreact with an antibody recognizing a large panel of cytokeratins in somatic cells nor with two antibodies recognizing IF common domains: the IFA, at the carboxy terminal end of the rod domain (Pruss et al. 1981) and the ME101 antibody at the amino terminal half (Escurat et al. 1989). It has already been observed that sperm antigens crossreacting with IF antibodies have a different molecular weight than that found in somatic cells, suggesting that germ cells may express cell-specific IF genes (Eddy et al. 1991). The somatic IFs are resistant to high-salt buffers (see review in Steinert et al. 1982). On the contrary, the MAb 4F7-immunoreactive protein is either modified or extracted by high ionic strength, as seen with 2 M NaCl, 2 M KCl, and 1 M KI. In addition, the fact that the FS is composed of 5–6-nm-wide filaments (Olson et al. 1976) argues against their somatic IF relationship. Moreover, the available FS cDNA sequence data obtained from the mouse predicted an FS protein that does not exhibit homology with known cytoskeletal proteins (Fulcher et al. 1995).
The N 350 anti-actin antibody labels the connecting piece and the fibrous sheath of the human spermatozoon. As for the MAb 4F7, the N 350 MAb stains both the inner and the outer FS sides (Flaherty et al. 1988). In cases of FS deficiency, actin and the 4F7-immunoreactive FS protein have the same behavior: according to the type of flagellar dysgenesis, both are present either in a common reduced part of the principal piece or in ejaculated cytoplasmic bodies. These data suggest that their assembly may be under control of some common morphogenetical factor(s). A close association of actin filaments and other cytoskeletal components is well known in somatic cells (see review in Carmo–Fonseca and David–Ferreira 1990). Their association also suggests a role in their spatial organization, as in the case of the interaction of the vimentin tail with actin-containing structures (Cary et al. 1994).
The FS appears after initiation of condensation of the spermatid nucleus. This is also the case for the FS component detected by MAb 4F7. It has been suggested that mRNAs coding for FS proteins are first stored, to be translated later in spermiogenesis (Oko and Clermont 1990). The absence of ribosomes in the extruded flagellar part, in which the FS appears, had suggested that FS precursors are produced in the spermatid cytoplasm (Clermont et al. 1990; Oko and Clermont 1989; Fenderson et al. 1988; Sakai et al. 1986). That an immature form of the 4F7-related protein may be present in the spermatid cytoplasm could not be excluded. Nevertheless, in the human, FS antigens recognized by the 4F7 (this study) and RT97 MAbs (Jassim et al. 1991) appear directly in the flagellar compartment. Differences among mammalian species in the biogenesis of FS are also suggested by the fact that the FS appears first in the proximal flagellar part in the mouse (Sakai et al. 1986) and the human (as seen with MAb 4F7) but not in the rat (Oko and Clermont 1989; Irons and Clermont 1982).
Therefore, the location of FS protein synthesis and the mechanism of their eventual transport from a cytoplasmic compartment and/or along the flagellum remain to be elucidated. However, an as yet unknown step in FS development has been revealed by MAb 4F7, which shows that the FS arises from granules regularly distributed along the flagellum. In addition, MAb 4F7 reveals that in the course of FS biogenesis the region proximal to the FS exhibits a transient accumulation of the 4F7-related FS component, in a size, form, and location that could correspond to the spindle-shaped body (Wartenberg and Holstein 1975). This body is filled with microtubule-like elements and is well developed in primates, unlike the case in rodents (Irons and Clermont 1982). Its possible involvement in FS protein deposition is strongly suggested from the present data, and MAb 4F7 should be a unique tool to investigate this spermatid-specific transient cytoskeletal structure.
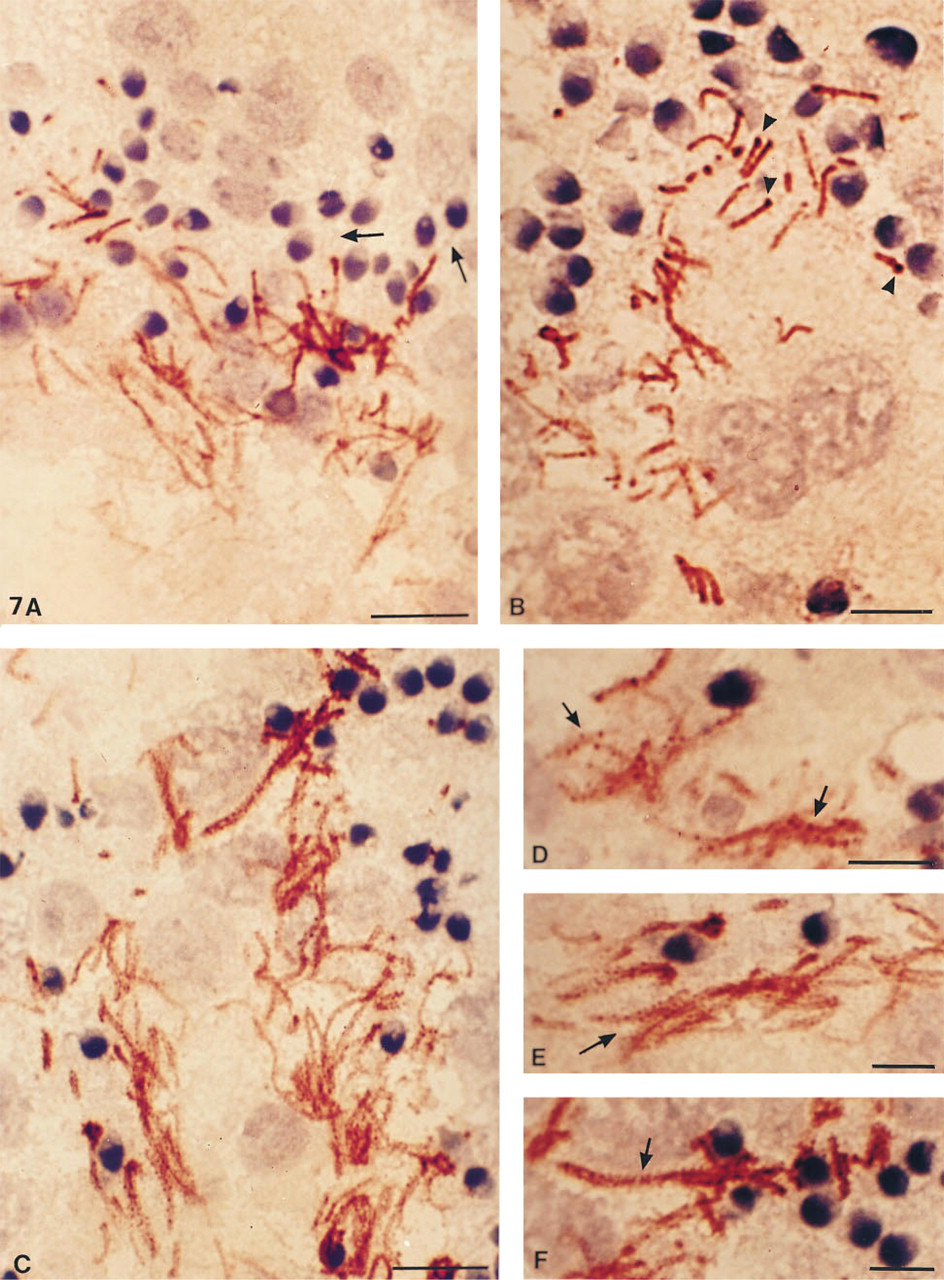
MAb 4F7 reveals developmental steps of fibrous sheath formation. Frozen testis sections fixed with paraformaldehyde and labeled by MAb 4F7 according to the three step method using biotin–avidin–peroxidase.
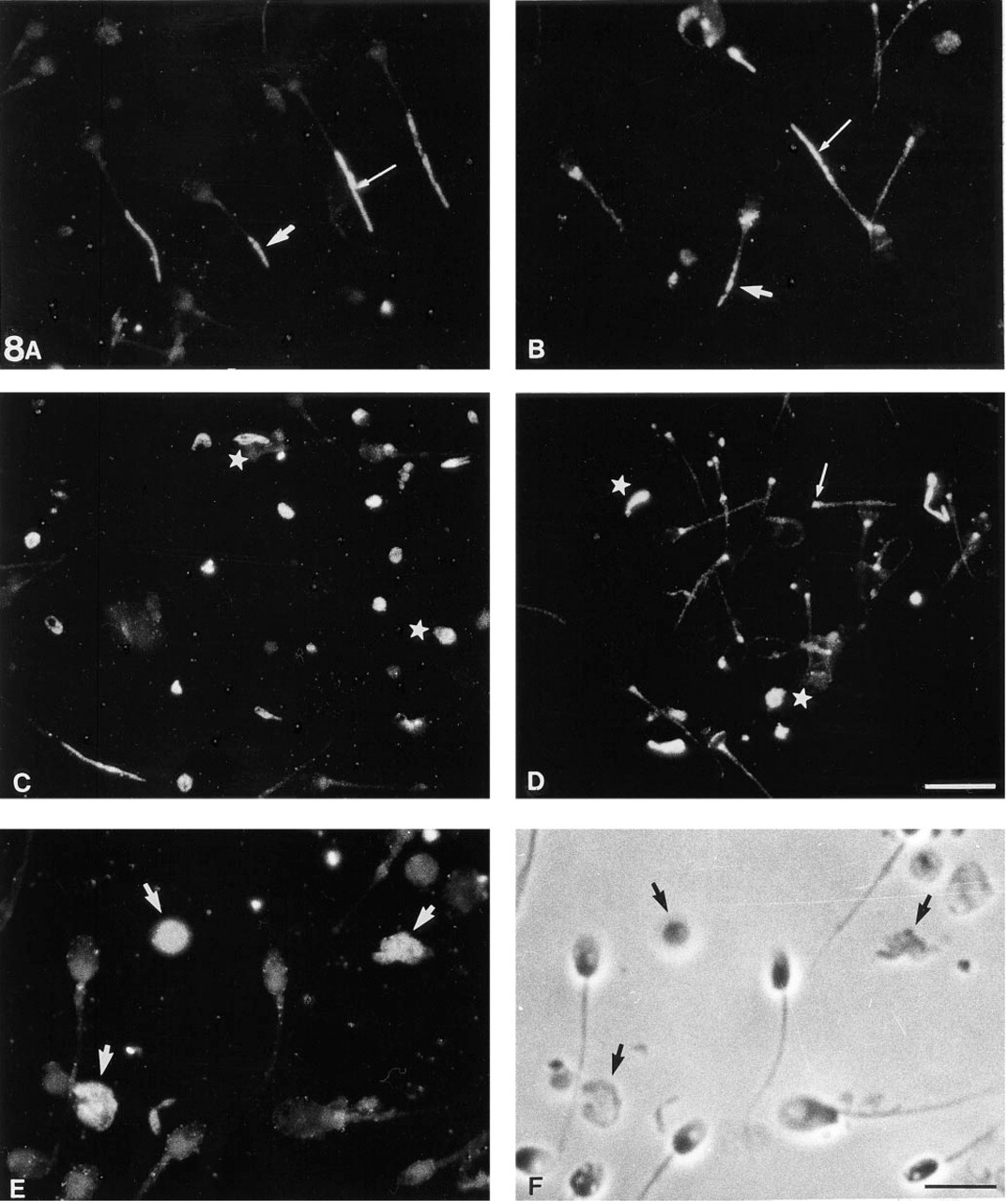
Pathological models reveal assembly relationships between actin and FS Protein(s). Indirect immunofluorescence using MAb 4F7 (A, C, E) or an MAb anti-actin
In conclusion, the protein of the sperm fibrous sheath recognized by MAb 4F7 could represent a spermatogenic-specific type of cytoskeletal proteins. A cDNA clone for a gene encoding the FS has been obtained only in the mouse (Fulcher et al. 1995). The availability of MAb 4F7 should offer the possibility of cloning the gene encoding the 4F7-immunoreactive protein and thus could be useful to analyze this particular cytoskeletal structure at the level of both molecular biology and germ cell differentiation.
Footnotes
Acknowledgements
We thank Dr M.M. Portier (Collège de France) for the gift of anti-IF antibodies. We also thank Dr J. Selva (CHU Bicětre, France), Dr J.L. Dacheux (INRA, Nouzilly, France), Dr D. Goffaux (Ecole Vétérinaire, Maison Alfort, France), and Dr M. Albert (UER Biomédicale des Saints Pères, Paris, France) for providing non-human semen samples.