
Abstract
Osteopontin (OPN), a major non-collagenous matrix protein of bone, is also found in tissue fluids and in the circulation. It is still not clear whether circulating OPN contributes to bone formation. To elucidate this question, rat OPN was tagged with dinitrophenol groups and administered to rats either intravenously or by infusion with an osmotic minipump through a “surgical window” in the bone of the hemimandible. Dinitrophenylated rat albumin (ALB) was used as a control. The presence and distribution of tagged proteins were revealed by immunogold labeling on sections of tibia and alveolar bone. Tagged molecules of OPN were found in mineralization foci, surfaces and interfaces, and matrix accumulations among calcified collagen fibrils. Even though dinitrophenylated ALB was administered at several-fold higher concentrations, it did not accumulate in these sites. These results show that circulating OPN can be incorporated into specific compartments of forming bone and suggest that such molecules may play a more important role than previously suspected.
O
Osteopontin is not only present in tissues but is also dissolved in serum and tissue fluids (Sodek et al. 2000; Rittling and Chambers 2004). The potential contribution of these circulating forms of OPN to calcified tissue biology has received little attention. Indeed, a number of bone matrix components are normally found in the circulation, but these are generally regarded as metabolic byproducts of bone formation and resorption that have no function at distant sites. To our knowledge, there is only one study that has explored the fate of circulating OPN (VandenBos et al. 1999). This light microscope study showed that intravenously administered [125I]OPN can be transported via the circulation and deposited in a number of calcified tissues. The amount of tracer administered was “three orders of magnitude” greater than the quantity of free OPN reported in human serum (concentrations in rat serum are not known). Under these conditions, some [125I]OPN was also found in enamel, a compartment in which the presence of OPN has not been revealed by biochemical assays and immunohistochemical techniques. Therefore, the authors concluded that they “could not exclude the possibility that the relatively high dose of injected OPN could have led to a somewhat artificial distribution pattern.” These results nonetheless clearly highlighted the possibility that OPN in calcified tissues is not only derived from local cellular sources but may also be recruited from outside the local environment via the circulation.
Proteins have been tagged with chemical groups other than 125I to visualize them. One such alternative method is dinitrophenylation, involving the covalent addition of dinitrophenol (DNP) groups to the ε-lysine residues of proteins (Little and Eisen 1967). This reaction, like iodination, generally does not alter the physiochemical properties of the tagged molecules (Kessler et al. 1982). Thereafter, detection of tagged proteins is highly sensitive, because several DNP groups can be attached to a protein and the antigenicity of those groups is resistant to tissue processing conditions (Kessler et al. 1982; Ghitescu and Bendayan 1992). Dinitrophenylated albumin (ALB) has been administered to study vascular permeability (Ghitescu and Bendayan 1992; Arshi et al. 2000). In calcified tissues, DNP-tagged ALB was used to investigate the uptake of proteins by ameloblasts and odontoblasts (Nanci et al. 1996). These cells, as well as osteoblasts, were shown to possess high levels of endocytotic activity and to take up protein non-selectively from the interstitial fluids.
The objective of the present study was to test the hypothesis that circulating forms of OPN may participate in bone formation. Tracer protocols such as those described above generally involve intravenous injections of relatively large amounts of proteins to saturate tissues throughout the body in quantities large enough to be detected. Such large dosages are rarely physiological. Our laboratory has developed an experimental system that allows the controlled administration of biological and chemical agents through a “surgical window” in the rat hemimandible (Vu et al. 1999; Orsini et al. 2001). This system was used to infuse near-physiological amounts of DNP-tagged OPN and to demonstrate that the tracer molecules reach and are incorporated into the same sites at which endogenous OPN is believed to accumulate and act (Nanci 1999).
Materials and Methods
Dinitrophenylation
Albumin (Sigma-Aldrich Canada; Oakville, ON, Canada) and bone-extracted rat OPN (Goldberg and Sodek 1994) were tagged with DNP according to the method described by Little and Eisen (1967). Briefly, distilled water, potassium carbonate (BDH; Toronto, ON, Canada), 2,4-dinitrobenzene sulfonic acid (Sigma-Aldrich), and proteins were mixed in a 0.2:1:2:4 ratio by weight for 48 hr at room temperature, shielded from light. At the end of the reaction, the unreacted DNP was removed by dialysis for 72 hr against 0.01 M phosphate-buffered saline (PBS) containing 0.85% NaCl, pH 7.2, in 12 kD dialysis tubing (Sigma-Aldrich). The complexes were concentrated by centrifugation at 300-350 × g, 4C for 20 min, in Centricon YM-10 ultrafiltration tubes (Millipore; Bedford, MA).
Surgical Procedures
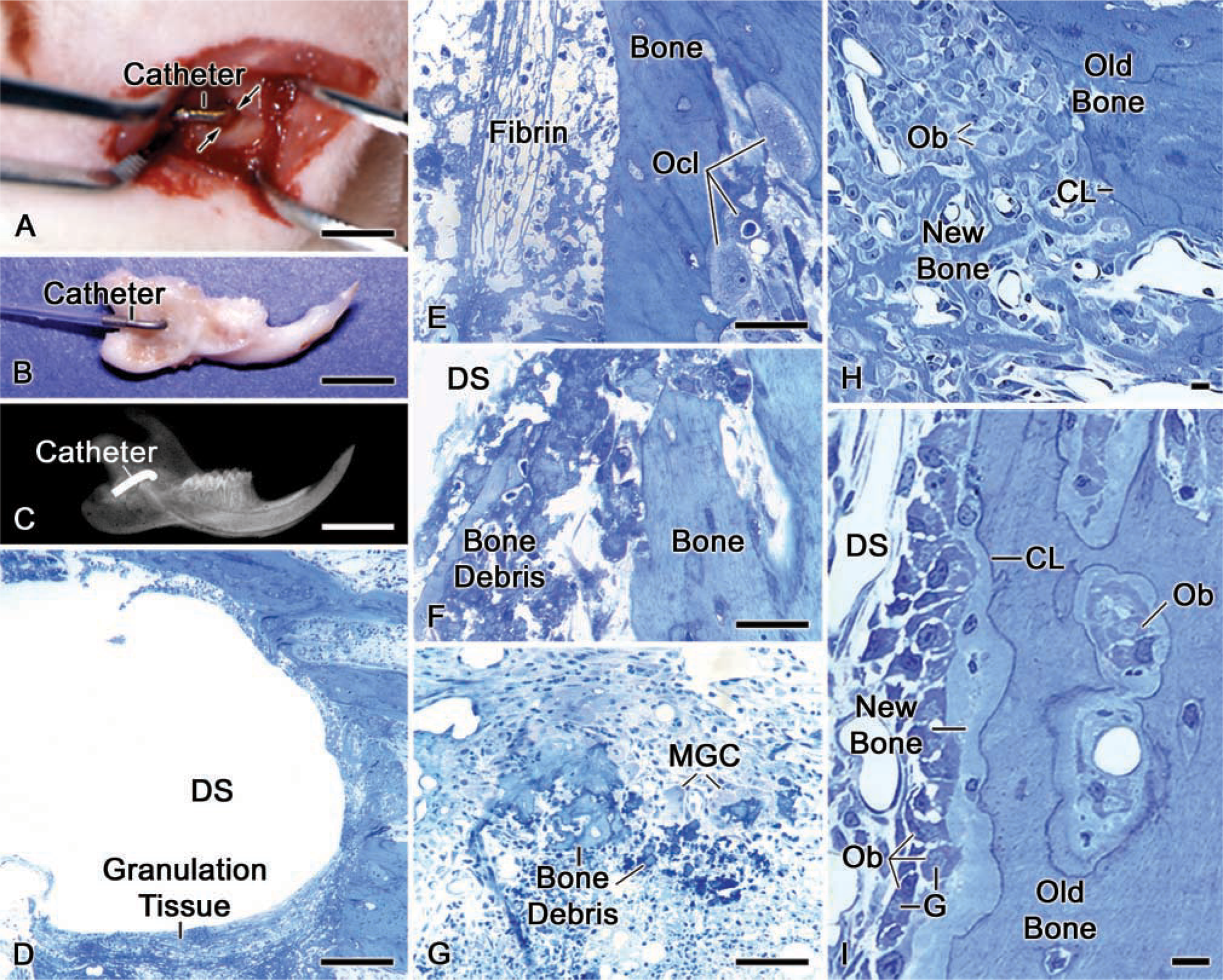
Juvenile (5-6-week-old), male Wistar rats weighing 100 ± 10 g (Charles Rivers Canada; St-Constant, QC, Canada) were anesthetized with an intraperitoneal injection of a 1:1:2 mixture of Hypnorm (fentanyl citrate and fluanison; Janssen Pharmaceutica, Beerse, Belgium), Versed (midazolam; Hoffmann-LaRoche, Mississauga, ON, Canada), and distilled water. An 8-mm incision was made through the skin following an imaginary line extending between the auditory meatus and the lip commisure (Figure 1A). To expose the hemimandible, the masseter muscle was separated along the length of the fibers with a scalpel surgical blade (No 15C; Almedic, Montreal, QC, Canada). A dental drill fitted first with a size 010 carbide round burr (Brassler; Montreal, QC, Canada), followed by a size 014, was used to make a hole in the alveolar wall on the bony elevation associated with the apical end of the incisor at ~2 mm from the posterior border of the ramus (Figure 1B). During drilling, the surgical site was irrigated with physiological saline. One- or 3-day Alzet osmotic minipumps [model 2001D for 1-day (8.0 μl/hr) and 1003D for 3-day (1.0 μl/hr); Alza, Palo Alto, CA] filled with complexes were slipped under the skin through a second incision made on the posterior region of the neck of the animal. A piece of vinyl tubing (size 0.72 × 1.22 mm; Scientific Commodities; Lake Havasu City, AZ) was hooked to the minipump and its free end passed through the neck area and underneath the masseter muscle. A metal catheter, made by bending a 20G1 needle (Becton-Dickinson; Rutherford, NJ), was used to connect the vinyl tubing to the bony hole. The metal catheter was immobilized against the bone surface with tissue adhesive (Indermil; Patterson Dental Supply, Montreal, QC, Canada) and bone cement (Zimmer; Warsaw, IN). The muscle was re-joined with 4-0 chromic gut sutures, and the skin was closed with 4-0 silk sutures (Patterson Dental Supply). The surgical site was cleaned and disinfected with 70% ethanol. The animals received an injection of Temgesic (buprenorphine HCl; Reckitt and Colman, Hull, UK) after surgery, and were fed with soft food containing Temgesic. X-rays, at 10 pulses/min, were taken to verify the positioning and the stability of the catheter (Figure 1C).
Six male Wistar rats weighing 100 ± 10 g (Charles River Canada) were anesthetized and injected through the jugular vein with DNP-tagged ALB or OPN.
The route of administration, concentrations, totals of amounts of complexes administered, and times of sacrifice of animals are summarized in Table 1. Negative control rats received only saline through the surgical window.
All animal procedures and experimental protocols described above were in accordance with guidelines of the Comité de Déontologie de l'Expérimentation sur les Animaux of the Université de Montréal.
(
Tissue Processing
The animals were anesthetized with 20% chloral hydrate solution (0.4 mg/g body weight; Fisher Scientific, Whitby, ON, Canada) and sacrificed by perfusion through the left ventricle with Ringer's lactate (Abbott Laboratories; Montreal, QC, Canada) for 30 sec, followed by a fixative solution consisting of 4% paraformaldehyde (BDH; Toronto, ON, Canada) and 0.1% glutaraldehyde (Electron Microscopy Sciences; Washington, PA) in 0.08 M sodium cacodylate (Electron Microscopy Sciences) buffer containing 0.05% calcium chloride (Sigma-Aldrich), pH 7.2, for 20 min. Treated and contralateral mandibles were taken, as well as the knees, and placed in the fixative solution for 24 hr at 4C. The hemimandibles and knees were washed with 0.1 M sodium cacodylate buffer, pH 7.2, and decalcified with 4.13% disodium ethylenediamine tetraacetic acid (Fisher Scientific) for 14 days at 4C (Warshawsky and Moore 1967). The decalcifying solution was changed every 2 days. Decalcified tissues were extensively washed in 0.1 M cacodylate buffer, pH 7.2, conventionally dehydrated in graded ethanols, and embedded in LR White resin (London Resin; Berkshire, UK) or osmicated with potassium ferrocyanide (Sigma-Aldrich)/reduced osmium tetroxide (Electron Microscopy Sciences) (Neiss 1984), dehydrated in acetone, and embedded in Taab 812 epoxy resin (Marivac; Halifax, NS, Canada). Both resins were polymerized at 58C for 48 hr. Some samples were left calcified and similarly processed for embedding.
Experimental outline
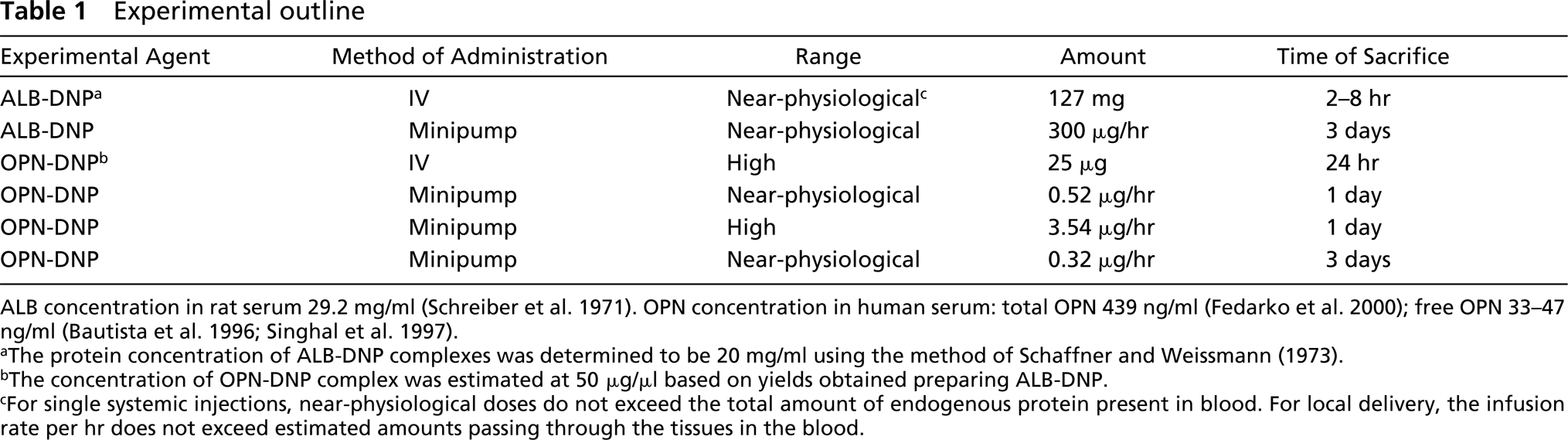
ALB concentration in rat serum 29.2 mg/ml (Schreiber et al. 1971). OPN concentration in human serum: total OPN 439 ng/ml (Fedarko et al. 2000); free OPN 33-47 ng/ml (Bautista et al. 1996; Singhal et al. 1997).
aThe protein concentration of ALB-DNP complexes was determined to be 20 mg/ml using the method of Schaffner and Weissmann (1973).
bThe concentration of OPN-DNP complex was estimated at 50 μg/μl based on yields obtained preparing ALB-DNP.
cFor single systemic injections, near-physiological doses do not exceed the total amount of endogenous protein present in blood. For local delivery, the infusion rate per hr does not exceed estimated amounts passing through the tissues in the blood.
Light microscope observations were made on 1-μm semithin sections obtained with glass knives on a Reichert Jung Ultracut E ultramicrotome and stained with toluidine blue. Ultrathin sections 80-100-nm thick were cut with a diamond knife and transferred on Formvar-coated (polyvinyl formate) 200-mesh nickel grids, and processed for postem-bedding colloidal gold immunolabeling.
Immunocytochemistry
Immunolocalization of proteins was done as previously described (Nanci et al. 1996) using the postembedding colloidal gold method (reviewed in Bendayan 1995). Briefly, gridmounted sections of osmicated tissues were first treated with a saturated aqueous solution of sodium metaperiodate (Fisher Scientific) (Bendayan and Zollinger 1983). All sections were placed for 15 min on blocking solution consisting of 0.01 M PBS, pH 7.2, containing 1% PBS-ovalbumin (Sigma-Aldrich) and then transferred onto a drop of anti-DNP antibody (1:200, 1 hr; DAKO, Carpinteria, CA) to reveal the DNP-protein complexes, anti-OPN (1:10, 2 hr; LF-123, courtesy of Dr. L.W. Fisher, NIDCR, NIH, Bethesda, MD), or rabbit anti-rat ALB (1:60, 2 hr; ICN Pharmaceutical, Aurora, OH) antibodies to immunodetect endogenous molecules. Following incubation with primary antibodies, the grids were rinsed with PBS and placed again on the blocking solution for 15 min. A protein A-gold complex with particle size of 10-12 nm (prepared in-house as described by Bendayan 1995) was used to reveal the site of antibody binding. Finally, the grids were washed with PBS, followed by distilled water. All grids were stained with 4% aqueous uranyl acetate for 6 min and with lead citrate for 2 min and were examined in a JEOL JEM-1200 operated at 60 kV or a JEOL JEM-2011 transmission electron microscope operated at 80 kV.
Results
Histological Observations
Light microscopic observations of semithin sections in the region of the surgical window of animals infused with complexes for 1 day showed the characteristics of an early tissue repair reaction (Figure 1D). Bone debris produced during drilling (Figure 1F), fibrin clot (Figure 1E), and a cellular infiltrate comprising inflammatory cells, some osteoclasts and/or multinucleated giant cells, and fibroblast-like cells (Figures 1D-1G) were present at the periphery of the hole. Although osteoclasts were seen on bone debris and in the bone at the periphery, their presence was not visibly increased in animals infused with dinitrophenylated conjugates (Figures 1E and 1G) compared with controls infused with saline only (data not shown). As early as 3 days after the start of infusion, evidence of new bone formation was observed in animals infused either with saline only or with DNP-protein complexes (Figures 1H and 1I).
Intravenous Injections of ALB-DNP
Endogenous ALB was immunodetected in the interstitial space between osteoblasts and in osteoid, but there was no significant accumulation in the bone matrix (Figure 2A). The presence of labeling between cells suggests that ALB can diffuse from the interstitial fluid, between cells and into osteoid. Tagged ALB was detected in the initial bone matrix deposited onto old bone (Figure 2B) and in osteoid (Figure 2C). Very few gold particles were found in association with cement lines, lamina limitans, or interfibrillar accumulations of non-collagenous matrix proteins. Calcified cartilage and bone exhibited almost no gold particles despite the occasional nearby presence of ALB-DNP in the tissue fluid (Figure 2D).
Intravenous Injections of OPN-DNP
At 24 hr following intravenous injections of OPN-DNP, tagged molecules were immunodetected on the surface of exposed calcified cartilage spicules in the primary spongiosa of the tibia (Figure 3A). In some mixed spicules, cement lines between calcified cartilage and bone or between adjacent layers of bone exhibited gold particles (Figure 3B). In general, bone was labeled along its surface but not in deeper regions, indicating that the unlabeled regions probably formed before the tracer was administered. Gold particles were found in mineralization foci in the osteoid seam and over accumulations of interfibrillar matrix in bone (Figure 3C). Endogenous OPN was immunodetected in these areas (Figure 3D).
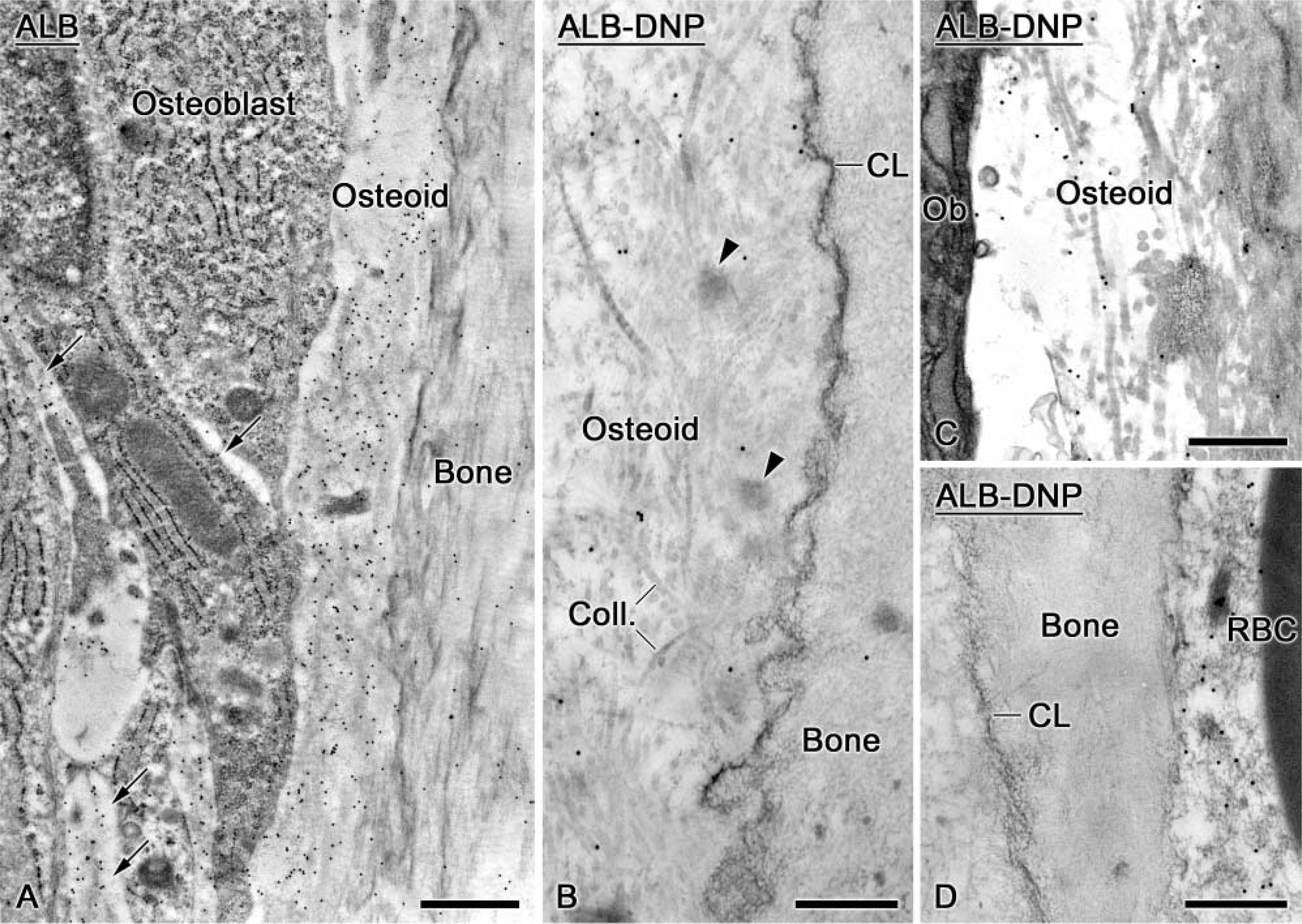
Electron micrographs illustrating the distribution of endogenous albumin (ALB) and of dinitrophenylated albumin (ALB-DNP) following intravenous injection. (
One-day Infusions of OPN-DNP
When high doses of OPN-DNP (Table 1) were infused over a 24-hr period, labeling was observed both at the drill site and at distant sites such as the growth plate. Gold particles were found in cement lines, interfibrillar matrix accumulations in bone (Figure 4A), and mineralization foci in osteoid (Figure 4B). At the drilling site, there was a concentration of tagged molecules on exposed bone surfaces, but many of them were also trapped in the adjacent fibrin clot (Figure 4C). Drilling caused microfractures and disrupted the collagen-packing network, resulting in diffusion of tracers into deeper bone (data not shown).
When near-physiological concentrations of OPN-DNP (Table 1) were infused for 24 hr, tagged proteins were found mainly in the surface layer of bone surrounding the hole and in the fibrin clot at the surgical site (Figure 4D). The density of labeling over these compartments appeared to be somewhat less than with the higher dose (compare Figures 4C and 4D). Some tracer molecules were found in deeper regions of bone along surfaces exposed during the surgical procedure, such as osteocyte canaliculi and split cement lines (Figure 4E). Very few gold particles were observed in deeper, undisturbed bone regions. Tagged molecules were not detected in tissue sections from tibia. With the tissue-processing and incubation conditions used in this study, no endogenous OPN was immunodetected in interstitial/circulating fluid compartments.
Three-day Infusions of ALB-DNP
Despite infusion of amounts of ALB-DNP many-fold larger than those for OPN, this tagged protein accumulated mainly in the fibrin clot in the hole region. However, some gold particles were also found over the bone matrix near surfaces exposed by the drilling (Figure 5A). The labeling associated with bone diminished away from the bony hole, and no apparent tagged ALB was observed at the surface of calcified bone matrix. Tagged molecules diffused through osteocyte canaliculi, but these were not incorporated into the bone matrix (Figure 5B). Some ALB-DNP was found among the collagen fibrils of osteoid situated in proximity to the surgical window (Figure 5C).
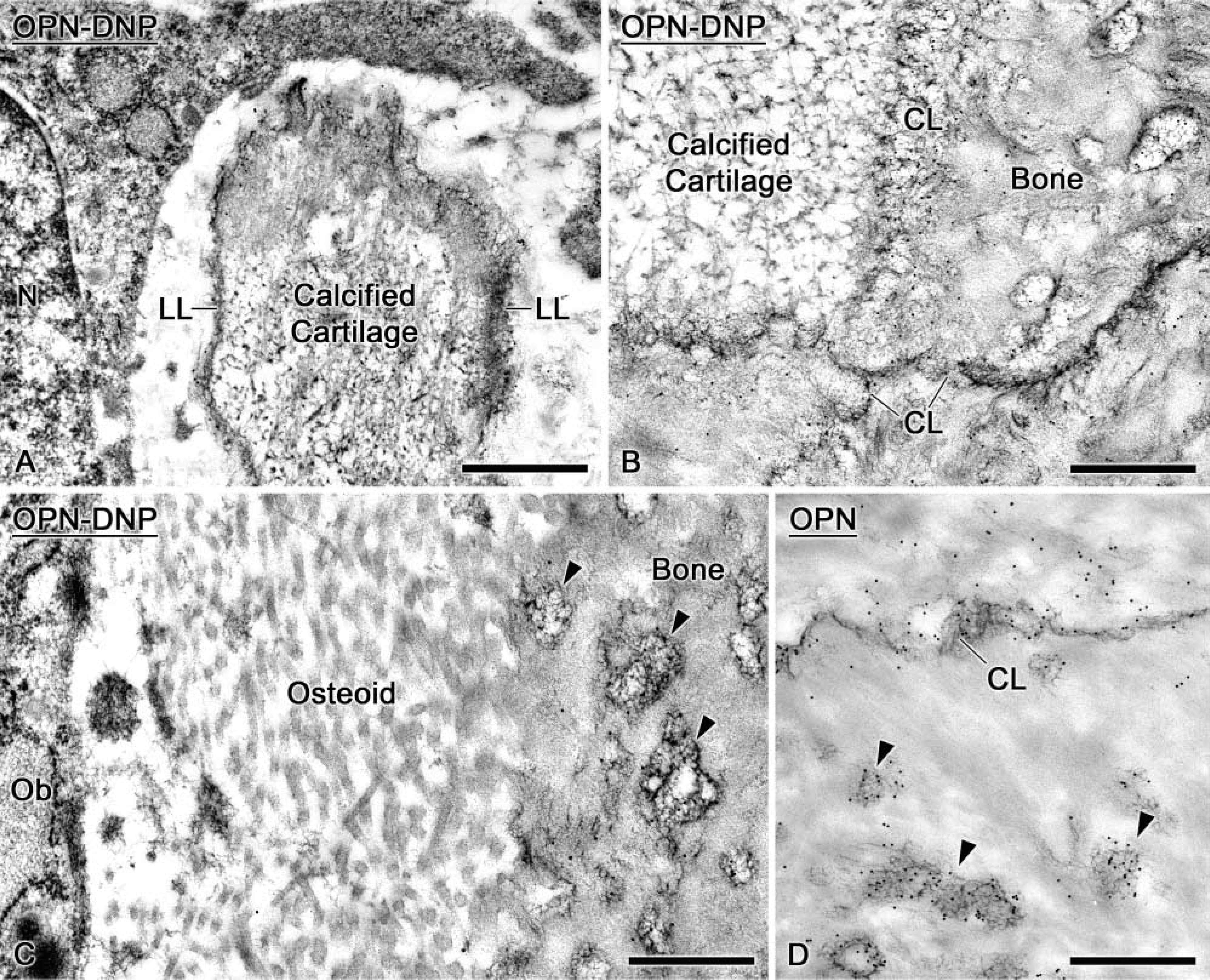
Immunocytochemical preparations of tibial bone (
Three-day Infusions of OPN-DNP
Some mineralization foci (Figure 6A) were labeled with OPN-DNP, and the tagged molecules integrated into both osteoid and newly formed bone (Figure 6B). OPN-DNP was also detected near the drill site along surfaces destined for resorption by osteoclasts or around small bone debris surrounded by macrophages (Figure 6C). Like endogenous OPN (Figure 6E), infused OPN-DNP was present in the lamina limitans, surrounding osteocytes and coating the bone surface near bone lining cells (Figure 6D).
Controls
Only a few randomly distributed gold particles were observed over tissue sections from animals injected with saline and incubated with anti-DNP antibody (data not shown). No significant immunolabeling was seen in contralateral hemimandible and tibial tissues. Incubation of tissue sections with protein A-gold likewise resulted in little background labeling.

Immunocytochemical preparations of (
Discussion
The results of this study demonstrate that OPN can be conjugated to DNP and that at least some OPN, transported via the blood circulation or diffusing locally in interstitial fluid, can integrate into bone under normal formative conditions or at sites of damage, a result consistent with the study of VandenBos et al. (1999).
Moreover, ultrastructural localizations of the tracer documented that exogenous OPN becomes incorporated into various compartments of bone in which endogenous OPN is believed to accumulate and act (Nanci 1999). One major advantage of the surgical window approach is that it allows the continuous administration of tracer in amounts that do not exceed the level of endogenous OPN constantly circulating through the tissues in the hemimandible.
ALB was employed as a control because this serum protein is found in bone but has a significantly lower inhibitory effect on hydroxyapatite formation than does OPN (Hunter et al. 1994). Some ALB-DNP is trapped in bone exposed during drilling, but the complexed protein, administered either systemically or locally, does not accumulate to any significant extent in non-collagenous matrix protein-enriched compartments. The relatively low affinity of this protein for bone is further demonstrated by the relatively modest labeling observed despite the fact that several-fold larger quantities of ALB-DNP were administered compared with OPN-DNP. Therefore, the behavior of ALB with respect to bone is not significantly changed by dinitrophenylation. This, together with the fact that OPN-DNP incorporates at sites in which endogenous molecules are believed to act, suggests that addition of DNP residues to a protein does not modify its affinity for bone.
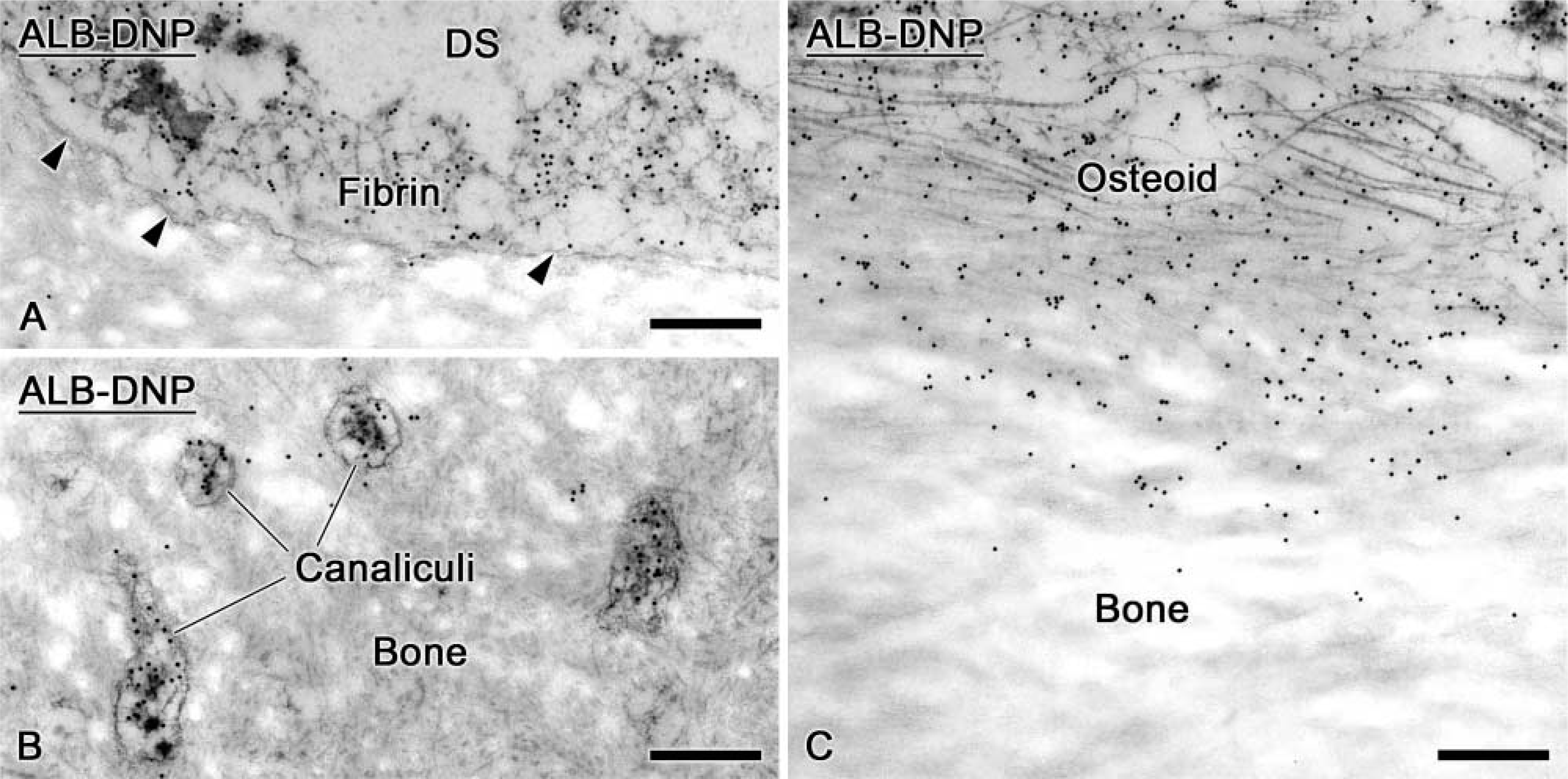
Immunocytochemical preparations of alveolar bone after a 3-day infusion of ALB-DNP. (
Although it would be anticipated that circulating OPN would be attracted to sites of mineralization, this may not necessarily be the case. It has been suggested that circulating OPN is strongly bound to complement factor H and thus is sequestered, and that its activities are limited to their functional ranges (Fedarko et al. 2000). Our results suggest that in our experimental model, binding of OPN to factor H must occur over a time frame that allows the molecules to be available for incorporation into bone and/or that OPN prefers a calcifying matrix to complement factor H.
In conclusion, this first ultrastructural study demonstrates that dinitrophenylated OPN can be traced following either systemic or local administration and that the surgical window in the rat hemimandible is an efficient system for investigating the fate of proteins administered at low concentrations per unit time. It also clearly shows that circulating OPN can integrate into bone compartments such as mineralization foci and cement lines. This suggests that the action of this matrix protein extends beyond its microenvironment. Circulating molecules may have an important impact on initial events of bone formation, for which few molecules are generally required. The surgical window approach allows investigation of the fate of non-collagenous matrix proteins over time after they are released from the cells that manufacture them, and is applicable to a number of functional studies, such as determination of the behavior of different isoforms and evaluation of the activity of predicted functional groups.
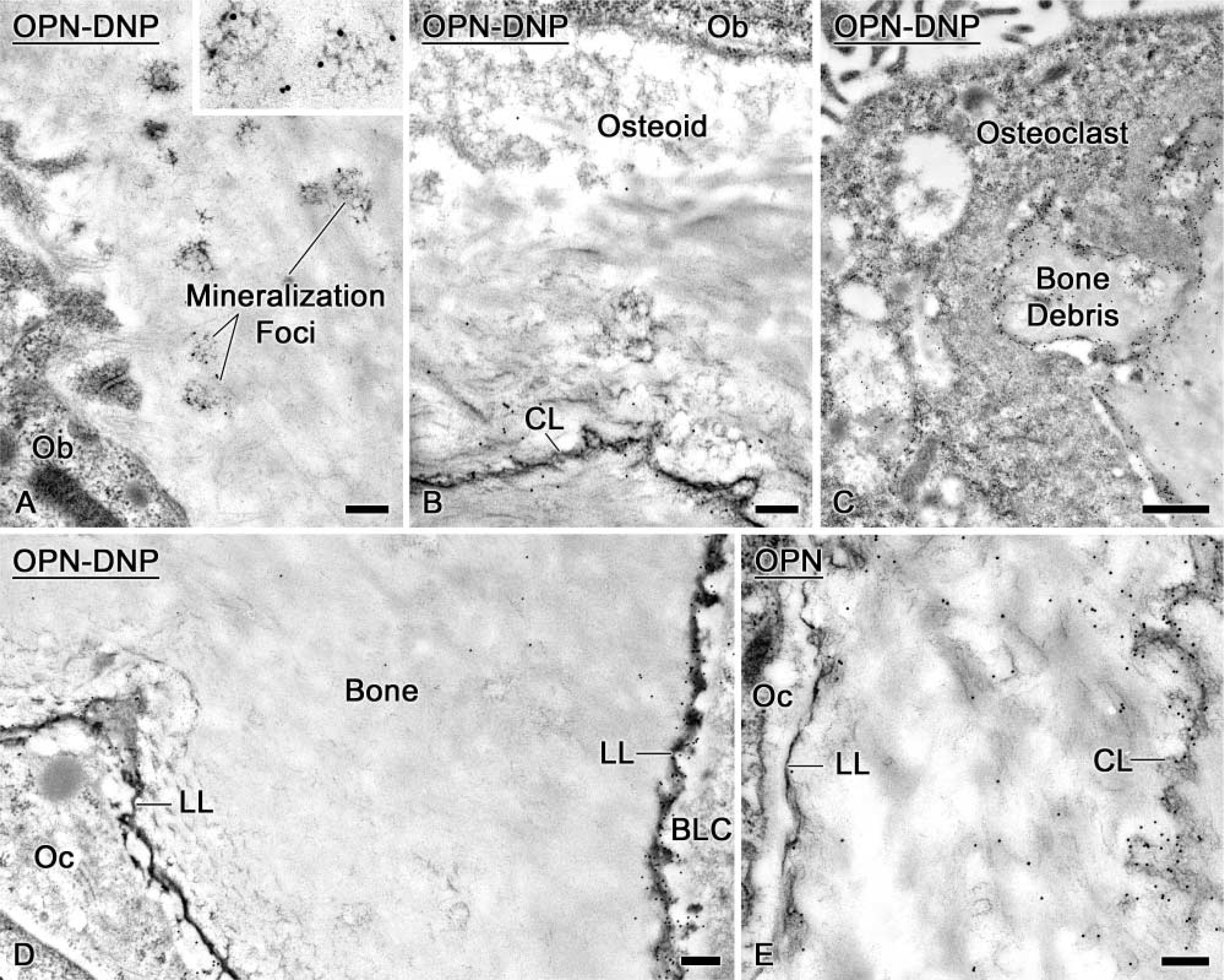
Electron micrographs of alveolar bone after 3-day infusions of near-physiological doses of dinitrophenylated osteopontin (
Footnotes
Acknowledgements
Supported by the Canadian Institutes of Health Research (CIHR).
We thank Dr Charles E. Smith for his comments and discussions on the manuscript.