
Abstract
Although considered a pericellular matrix component, hyaluronan was recently localized in the cytoplasm and nucleus of proliferating cells, supporting earlier reports that hyaluronan was present in locations such as the nucleus, rough endoplasmic reticulum, and caveolae. This suggests that it can play roles both inside and outside the cell. Hyaluronan metabolism is coupled to mitosis and cell motility, but it is not clear if intracellular hyaluronan associates with cytoskeletal elements or plays a structural role. Here we report the distribution of intracellular hyaluronan, microtubules, and RHAMM in arterial smooth muscle cells in vitro. The general distribution of intracellular hyaluronan more closely resembled microtubule staining rather than actin filaments. Hyaluronan was abundant in the perinuclear microtubule-rich areas and was present in lysosomes, other vesicular structures, and the nucleolus. Partially fragmented fluorescein-hyaluronan was preferentially translocated to the perinuclear area compared with high-molecular-weight hyaluronan. In the mitotic spindle, hyaluronan colocalized with tubulin and with the hyaladherin RHAMM, a cell surface receptor and microtubule-associated protein that interacts with dynein and maintains spindle pole stability. Internalized fluorescein-hyaluronan was also seen at the spindle. Following telophase, an abundance of hyaluronan near the midbody microtubules at the cleavage furrow was also noted. In permeabilized cells, fluorescein-hyaluronan bound to RHAMM-associated microtubules. These findings suggest novel functions for hyaluronan in cellular physiology.
I
Previous studies suggested that most of the hyaluronan in cells was small (<90 kD) and was found in vesicles of various sizes and shapes and/or other membranous compartments, and that smooth muscle cells prefer to take up low-molecular-weight hyaluronan (McGuire et al. 1987; Kan 1990; Tammi et al. 2001). The question remains, however, whether hyaluronan can serve an as-yet-undefined structural role within the cell. Recently the term “hyaluronasome” was proposed as a membrane-bound structure rich in hyaluronan in which coordinated synthetic and catabolic enzyme reactions occur in regulating the activities of the intracellular hyaladherins (Stern 2003). We previously found altered levels of intracellular hyaluronan-binding sites and their dramatic redistribution following serum stimulation of smooth muscle cells (Evanko and Wight 1999), but the identity of the binding sites was not clear. Various intracellular hyaladherins have been described, suggesting that intracellular hyaluronan may have important regulatory roles in such processes as cell cycle regulation, mitosis, cell motility, and RNA splicing (Hall et al. 1994; Grammatikakis et al. 1995; Deb and Datta 1996; Collis et al. 1998; Hofmann et al. 1998; Zhang et al. 1998; Assmann et al. 1999; Haddad and Turley 2000; Huang et al. 2000; Maxwell and Pilarski 2000; Hall et al. 2001). Intracellular hyaluronan may function in these processes in a transient fashion, either following intracellular synthesis or after uptake.
Given the structural role of hyaluronan in the pericellular matrix, it is possible that hyaluronan also serves a dynamic structural role inside cells or is part of a mechanotransduction mechanism. However, the relationship of intracellular hyaluronan with cytoskeletal elements has not yet been described. The hyaluronan receptors RHAMMIHABP (Assmann et al. 1999; Maxwell and Pilarski 2000) and CD44 (Underhill and Toole 1979) are both known to associate with the cytoskeleton. RHAMM was first described as a cell surface hyaluronan receptor that mediates hyaluronan-induced motility (Turley and Torrance 1984), In addition, RHAMM has also been described as a microtubule-associated protein that interacts with dynein and helps maintain spindle pole stability (Zhou et al. 2002; Maxwell et al. 2003). However, it is not clear whether hyaluronan is associated with RHAMM intracellularly. CD44, on the other hand, has been colocalized with hyaluronan within endosomes (Aguiar et al. 1999). Here we present evidence for a close relationship between intracellular hyaluronan, microtubules, and RHAMM in human arterial smooth muscle cells. Their distribution around the mitotic spindle and nucleolar area suggests novel functions for hyaluronan within cells.
Materials and Methods
Human SMCs were kindly provided by Elaine Raines (Department of Pathology, The University of Washington). Staining for intracellular hyaluronan was done as previously described using a biotinylated hyaluronan-binding protein specific for hyaluronan (Evanko and Wight 1999). Cells (1.5 × 105 per 35-mm dish) were plated and grown on 22-mm2 coverslips for 24 hr in DMEM containing 10% fetal bovine serum. Extracellular hyaluronan was removed prior to fixation by incubation of the cells with 2 U/ml Streptomyces hyaluronidase (Sigma; St Louis, MO) at 37C for 30 min in the culture medium. Cells were fixed in 2.5% formalin in PBS for 10 min, 22C and then permeabilized with 0.5% Triton X-100 for 10 min. The biotinylated probe was used at a concentration of 2.5 μg/ml in PBS containing 1% BSA, for 1 hr at 22C. The bound probe was localized either with streptavidin-horseradish peroxidase, FITC avidin, or Texas Red-streptavidin. Color for peroxidase labeling was developed with 3-amino-9-ethylcarbazole. The reaction was stopped after 5 min by rinsing in dH2O. Coverslips were mounted with Crystal Mount or Gel Mount (Biomeda; Foster City, CA). Brightfield images were taken with a light microscope (Axioscope, Carl Zeiss; Gottingen, Germany). Controls for specificity of intracellular hyaluronan staining included digestion of the cells with Streptomyces hyaluronidase after permeabilization (Figure 1D) and preincubation of the probe with an excess of hyaluronan (data not shown). All contrast in the brightfield micrographs arose from intracellular hyaluronan staining.
In some experiments, cells were also stained for tubulin using a monoclonal antibody to α-tubulin (Amersham Biosciences; Piscataway, NJ) or RHAMM using a polyclonal antibody generously provided by Volker Assmann (Richard Dimbleby Department of Cancer Research/ICRF Laboratory, St Thomas' Hospital, London, UK) and visualized with the appropriate Texas Red- or FITC-conjugated secondary antibodies. Staining for actin was done using bodipy-phalloidin, followed by hyaluronan staining. The antibody to the lysosomal marker, LAMP-1, comes from the Developmental Hybridoma Studies Bank at the University of Iowa. In some cases, cells were stained with fluorescein-hyaluronan after permeabilization to examine intracellular hyaluronan binding sites. Where noted, nuclei were stained with DAPI. Controls were stained with secondary antibody alone and were negative (Figure 1D). Fluorescent specimens were examined using either conventional fluorescence microscopy (Zeiss Axioscope, Carl Zeiss) or a confocal system (Leica TCS SP/NT on a Leica DMIRBE inverted microscope, Leica Microsystems Inc.; Bannockburn, IL). To examine uptake of exogenous hyaluronan, cells were incubated in the presence of fluorescein-labeled hyaluronan for 18 hr as described previously (Evanko and Wight 1999), and where appropriate, followed by staining for microtubules or RHAMM. In some cases, the fluorescein-hyaluronan (mol. wt. 1000 kD) was partially fragmented before incubation with the cells by brief digestion with Streptomyces hyaluronidase, 100 mU/ml, for 10 min at 37C or by brief sonication as previously described (McGuire et al. 1987). As determined by comparison on agarose gels, the sonicated material had a molecular weight of 300 kD and the enzyme-digested material was less than 50 kD. Hyaluronan binding sites were localized in fixed and permeabilized cells by incubation for 30 min with high-molecular-weight fluorescein-labeled hyaluronan (400 ng/ml), in PBS, 1% BSA. Negative controls included preincubation with excess unlabeled hyaluronan or exhaustive digestion of the fluorescein-hyaluronan with Streptomyces hyaluronidase (data not shown; see (Evanko and Wight 1999).
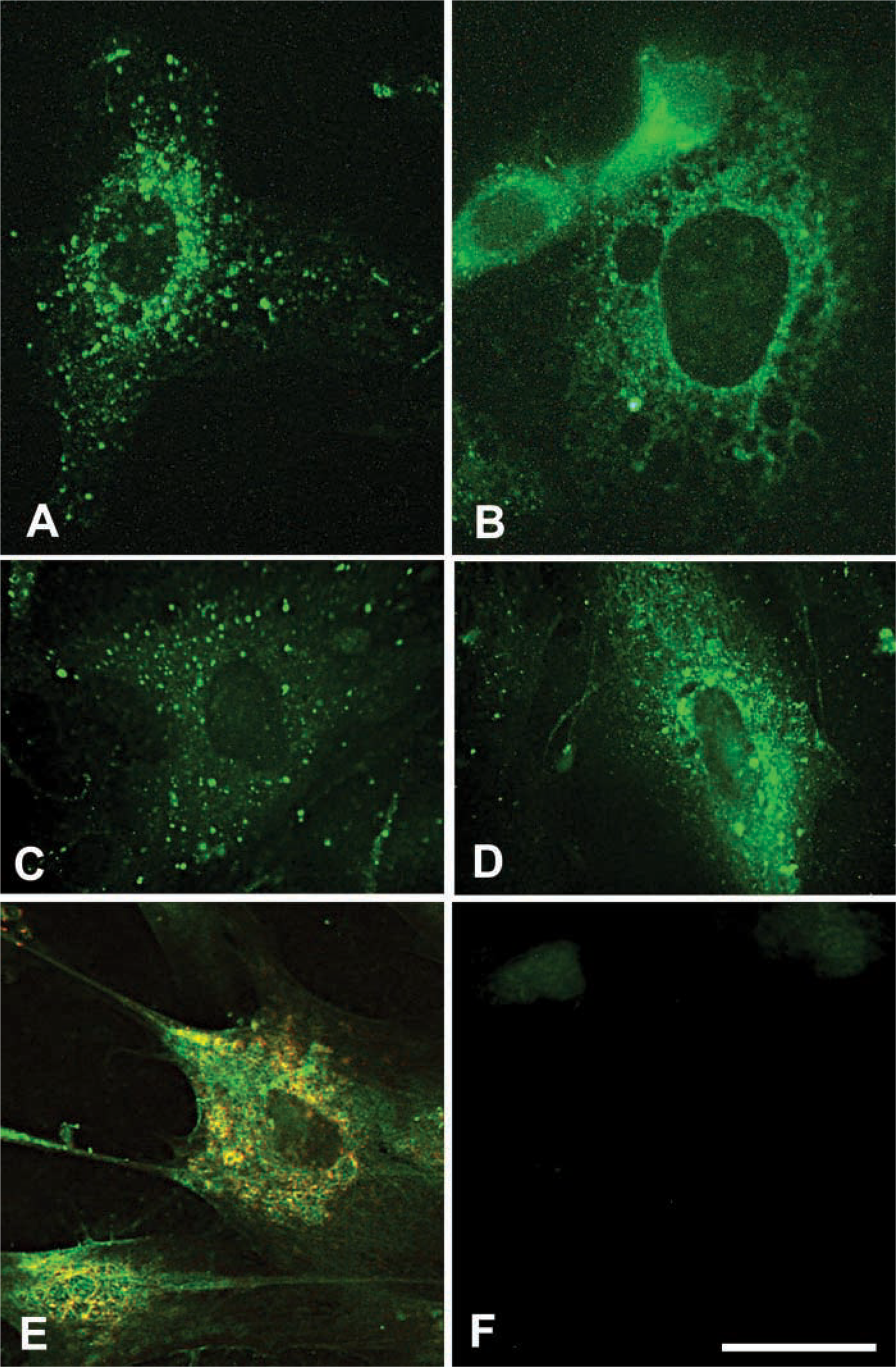
Size-dependent translocation of hyaluronan and partial lysosomal distribution. Smooth muscle cells were allowed to take up fluorescein-hyaluronan (fHA) of various molecular weights for 6 hr. (
Results
Size-dependent Intracellular Distribution of Hyaluronan
We previously found that hyaluronan was localized in vesicles of various sizes and was distributed in a diffuse network-like pattern in the perinuclear area as well as in the nucleolus in smooth muscle cells and fibroblasts (Evanko and Wight 1999). In that study, we also found that fluorescein-labeled hyaluronan was taken up more avidly by proliferating cells, but it appeared to be localized differently than the non-labeled intracellular hyaluronan, i.e., mostly restricted to large vesicles and much less prominent in the perinuclear area (Evanko and Wight 1999). This suggested that either there was an intracellular source of hyaluronan or that there was a difference in how the cells process their own hyaluronan vs the labeled material. Furthermore, another study indicated that smooth muscle cells prefer to take up hyaluronan of lower molecular weight (McGuire et al. 1987), while the hyaluronan used in our experiments was of high molecular weight (approx. 1000 kD).
Therefore, we examined the effect of hyaluronan size on the intracellular distribution following uptake by smooth muscle cells (Figures 1A and 1B). High molecular weight fluorescein-hyaluronan mostly remained in larger vesicles with some translocation to the perinuclear area, as seen previously (Figures 1A and 1C). In contrast, when the fluorescein-hyaluronan was partially fragmented to less than 50 kD by digestion with Streptomyces hyaluronidase (Figure 1B) or 300 kD by sonication (Figure 1D) before feeding to cells, it appeared more similar to the cell-derived hyaluronan, i.e., in a more diffuse network-like pattern throughout the cytoplasm and predominantly in the perinuclear area (compare Figures 1B and 1D with Figure 1E). This suggests that lower molecular weight hyaluronan may be more efficiently translocated to the perinuclear area and other cytoplasmic compartments. Translocation of tagged hyaluronan to the nucleus itself was not dramatic under these conditions, regardless of molecular weight. The signal within the nucleus was either weakly diffuse or faintly concentrated to the nucleoli.
Partial Lysosomal Distribution
Comparison of endogenous intracellular hyaluronan staining using a specific hyaluronan-binding probe with an antibody to the lysosomal marker LAMP-1 by confocal microscopy indicated that a substantial portion of the hyaluronan is in the lysosomal compartment in smooth muscle cells (Figure 1E). However, some hyaluronan did not colocalize with LAMP-1, such as that in the nucleolus (see below), early endosomes (data not shown) and some cytoplasmic regions near the nucleus.
Relationship of Hyaluronan with Microtubules and RHAMM
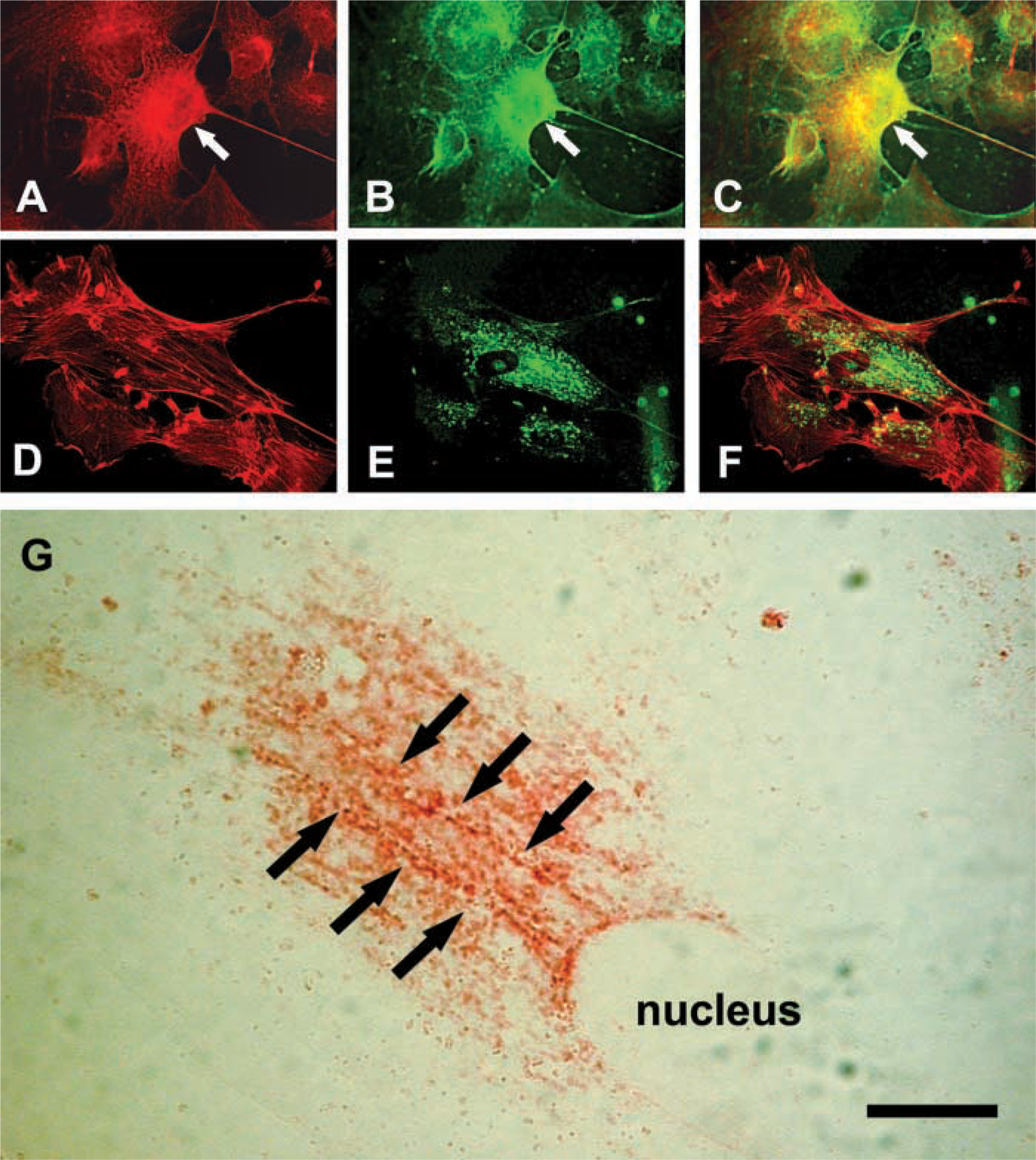
To compare the distribution of hyaluronan with cytoskeletal elements, permeabilized smooth muscle cells were double stained for endogenous hyaluronan and either f-actin or tubulin (Figure 2). The overall distribution of the hyaluronan staining within the cytoplasm most closely resembled microtubule organization (Figures 2A-2C). Hyaluronan codistributed with the microtubules in the perinuclear network and also more peripherally in the cytoplasm. In contrast, little or only occasional relationship between the intracellular hyaluronan and actin filaments was apparent (Figures 2D–2F). Hyaluronan-positive vesicles were frequently seen in distinct linear arrays or tracts that connected to the nuclear periphery (Figure 2G).
At higher magnification, extremely close apposition of hyaluronan staining and microtubules can be seen, both in the cytoplasm and adjacent to the nucleolar area (Figures 3A–3D). Notably, however, the hyaluronan was present immediately between and adjacent to the microtubules. Microtubules were seen in close association with both large and small hyaluronan-positive vesicle-like structures.
Cells were also stained using a polyclonal antibody specific for RHAMM, a microtubule-associated hyaluronan-binding protein. Although RHAMM binds to hyaluronan, to date there has been no visual evidence for an association of RHAMM with hyaluronan inside cells. In smooth muscle cells, RHAMM was present on the perinuclear microtubular network and linear arrays connecting to the nucleus (Figures 3E-3H), as well as the microtubules associated with nuclear clefts (data not shown). RHAMM-positive microtubules immediately surrounded hyaluronan-positive vesicle-like structures.
Using a different approach, intracellular hyaluronan-binding sites were localized by incubating fixed and permeabilized cells with fluorescein-hyaluronan and compared with the distribution of RHAMM (Figure 4). The fluorescein-hyaluronan bound directly to the microtubule structures in the perinuclear network that were stained with the anti-RHAMM antibody. This is in slight contrast to the distribution of endogenous hyaluronan and following uptake of fluorescein-hyaluronan by living cells, which localized between microtubules. Some RHAMM-positive structures, such as the thinner microtubules in the periphery of the cytoplasm, did not bind the fluorescein-hyaluronan. This suggests that some of the RHAMM associated with perinuclear microtubules is available for hyaluronan binding, i.e., not saturated. Binding of the labeled hyaluronan could be abolished by preincubation with excess unlabeled hyaluronan [data not shown, see (Evanko and Wight 1999)].
Involvement of Intracellular Hyaluronan in Mitosis
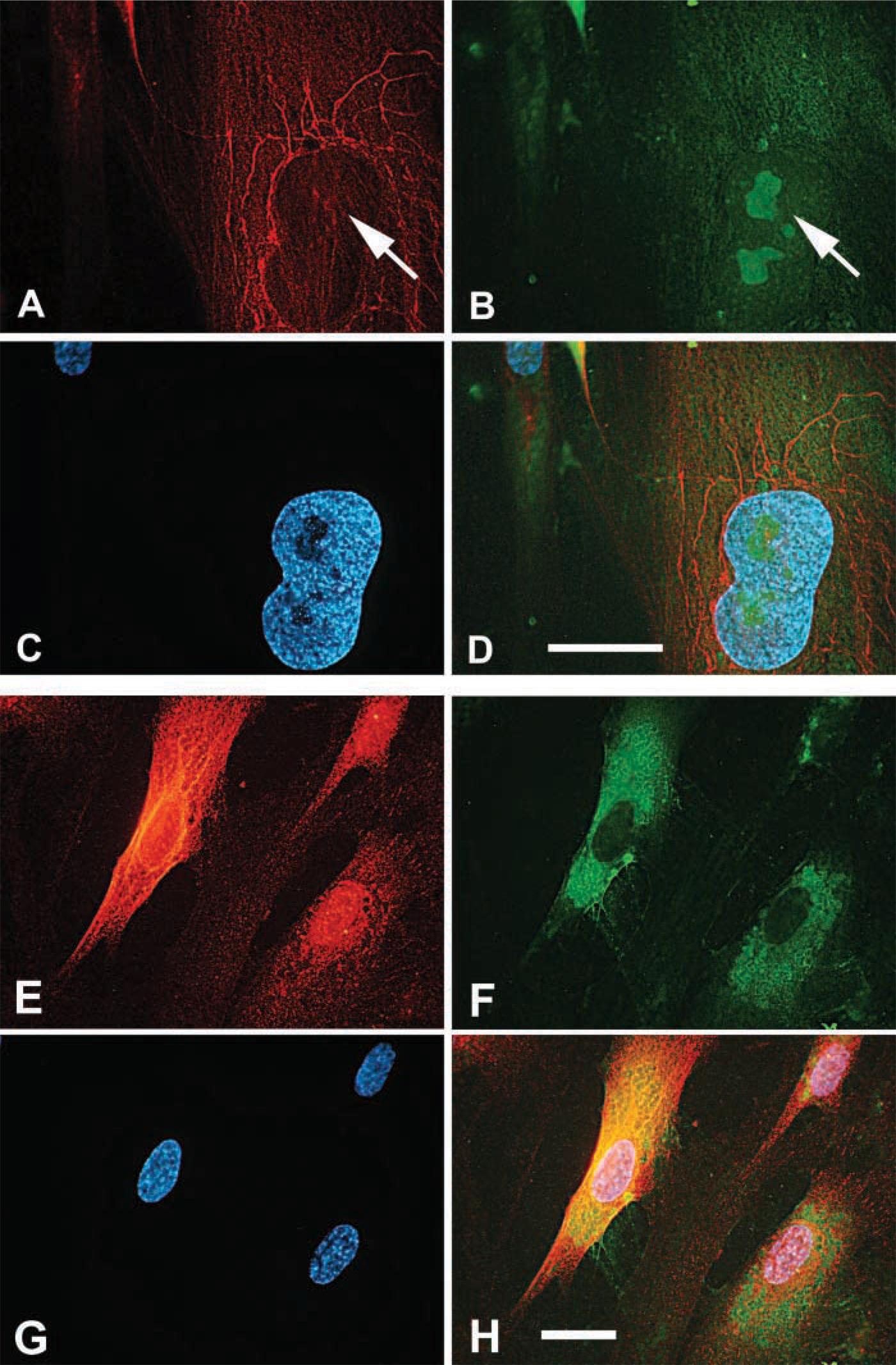
As previously described, intracellular hyaluronan was abundant in mitotic smooth muscle cells (Evanko and Wight 1999). Strong intracellular staining was seen during prophase (Figure 5A) during which cell detachment and rounding occur. This happens concurrently with the formation of a thick hyaluronan-rich pericellular matrix that involves both increased hyaluronan synthesis (Brecht et al. 1986) and traction of the hyaluronan and associated molecules into thick accumulations around the cell (Evanko et al. 1999). Intense hyaluronan staining persists through telophase (Figure 5B). In mitotic cells, staining for endogenous hyaluronan closely overlapped with RHAMM in and around the mitotic spindle (Figure 5C–5F), indicating that the hyaluronan is intimately associated with spindle and astral fibers during mitosis. Following uptake and translocation of exogenously added 1000 kD fluorescein-hyaluronan by living cells (Figure 5G), colocalization with RHAMM in mitotic spindles has also been seen. This suggests that some of the hyaluronan in this location does come from uptake. Late in mitosis, intense staining for hyaluronan was also seen around the microtubules of the midbody during cytokinesis (Figure 6A–6D), or occasionally in relatively large vesicles between the separating chromosomes (Figure 6E).
Similar distribution of hyaluronan and microtubules. (
Discussion
Intracellular Hyaluronan
The present observations add to a growing number of recent studies regarding intracellular hyaluronan. We noted here that intracellular hyaluronan staining generally resembled microtubule distribution, with only occasional codistribution with actin fibers. Much of the hyaluronan colocalized with a lysosomal marker, indicating that it may be destined for eventual degradation. However, some of the hyaluronan was in nonlysosomal vesicular compartments and in the nucleus. This is consistent with a previous study in which hyaluronan was present in endosomes and perinuclear tubular vesicles in epidermal keratinocytes (Tammi et al. 2001). Hyaluronan has also been localized to the rough endoplasmic reticulum (Londono and Bendayan 1988; Kan 1990; Evanko and Wight 1999), as well as nuclei and nucleoli (Londono and Bendayan 1988; Kan 1990; Evanko and Wight 1999). In smooth muscle cells and endothelial cells, hyaluronan was found in caveolae (Eggli and Graber 1995). Collis et al. (1998) showed that exogenous Texas Red-hyaluronan was rapidly translocated to the nucleus in 10T1/2 cells. In our experiments with human smooth muscle cells, translocation of fluorescein-hyaluronan to the nucleus was weakly evident and not as pronounced. This may be due to differences in the fluorescent label in the two preparations or the cells themselves. However, the presence of hyaluronan in such diverse locations suggests that it may have a wide array of intracellular functions.
Relationship of tubulin and RHAMM with endogenous hyaluronan. Cells were stained for (
Previous work has indicated that cells prefer to take up hyaluronan of low molecular weight (McGuire et al. 1987). Our results further these findings and suggest that the size of the hyaluronan may also influence its distribution intracellularly. These findings raise the question of whether the “hyaluronasome” (Stern 2003), or membranous organelle containing hyaluronan in various states of synthesis and degradation, has some as-yet-unknown intracellular function.
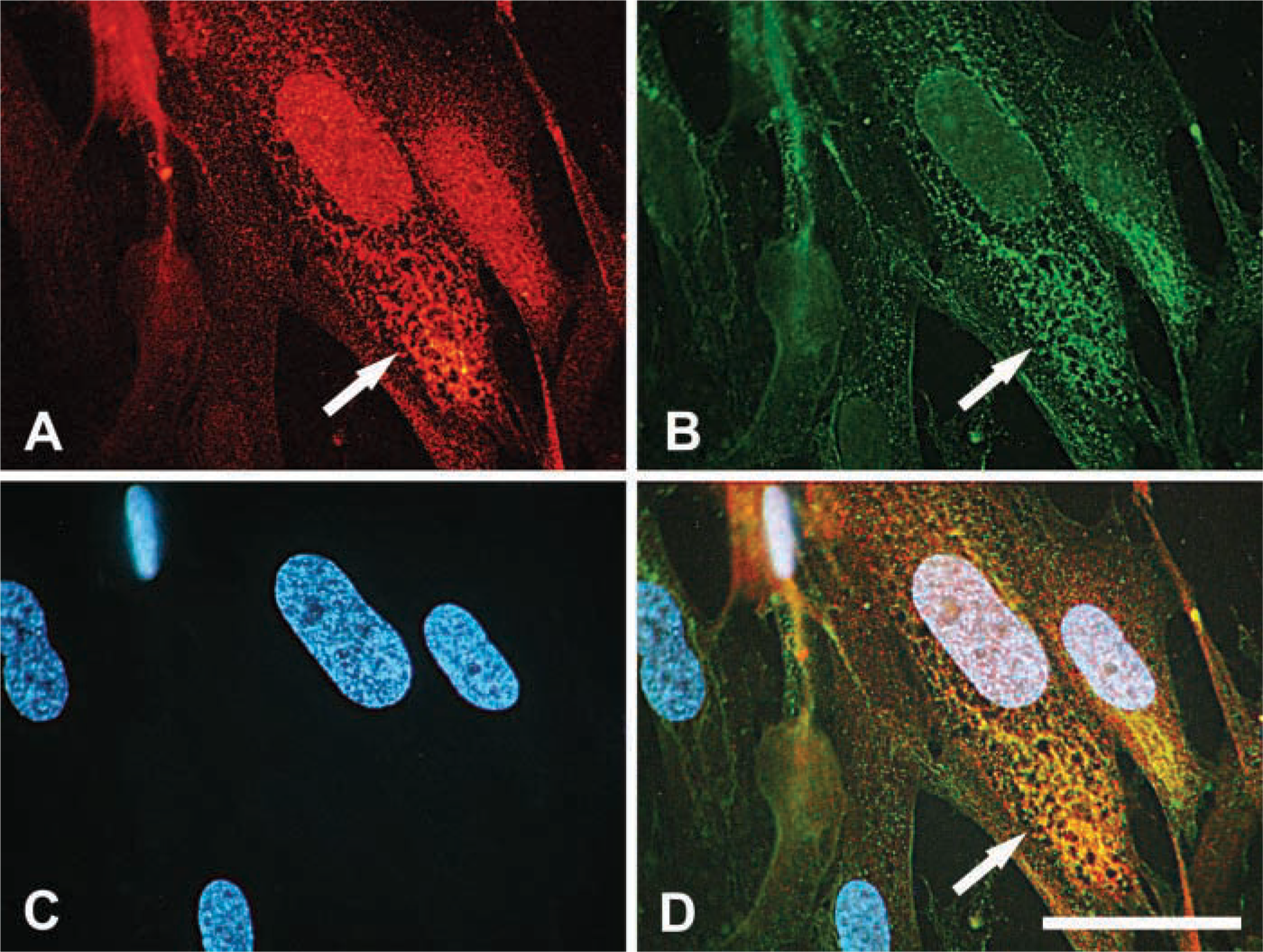
Fluorescein-hyaluronan binds to RHAMM-associated microtubules. Fixed and permeabilized cells were stained with (
Hyaluronan, Nuclear Architecture, Microtubule Function and Mitosis
A number of morphological observations suggest that there may be a relationship of intracellular hyaluronan to nuclear architecture and/or the mitotic process. These observations are as follows: (a) the presence of hyaluronan in association with heterochromatin and nucleoli (Londono and Bendayan 1988; Ripellino et al. 1989; Kan 1990; Evanko and Wight 1999); (b) the binding of fluorescein-labeled hyaluronan to the nuclear periphery and to nucleoli in fixed cells with a redistribution of binding sites following serum stimulation (Evanko and Wight 1999); (c) translocation of hyaluronan to the nucleus concurrently with stimulation of cell motility (Collis et al. 1998); (d) the association of intracellular hyaluronan with clefts and furrows in the nucleus of some cells (Evanko and Wight 1999); (e) elevated synthesis of hyaluronan during G2/M (Brecht et al. 1986), and the dynamic accumulation of hyaluronan in the pericellular matrix and intracellularly in mitotic cells (Evanko and Wight 1999; Evanko and Wight 2001); (f) colocalization of hyaluronan with microtubules and the microtubule-associated protein, RHAMM, at the mitotic spindle, and nucleolar area (present study); and (g) positive hyaluronan staining around microtubules at the location of the cleavage furrow [current study and (Evanko and Wight 2001)].
The abundance of a hyaluronan-positive compartment that is closely associated with the mitotic spindle and the rapid uptake that occurs at this time suggests that intracellular hyaluronan plays a role in the mitotic process. Microtubules are compression-bearing elements of the cytoskeleton that are thought to partly balance cell contraction (Stamenovic et al. 2002) and participate in key events in mitosis such as chromosome rearrangement and cleavage furrow formation. The microtubules most likely participate in the trans-location of the hyaluronan to lysosomes, the nucleus and other sites. However, hyaluronan is a well-known lubricant in synovial joints and may function similarly at the cell membrane/matrix interface (Evanko et al. 1999). This theme may also continue inside the cell. In other words, through an association with RHAMM, hyaluronan could help lubricate the intracellular motor machinery, influencing the viscosity and friction of the cellular microenvironment during microtubule-based movements (Figure 7). Alternatively, through its contribution to cytoplasmic swelling pressure, a hyaluronan-rich compartment may contribute to lateral stabilization or spacing of the microtubules and thus influence force balance within the cell during mitosis or other cell movements. In addition to a passive physical role, the hyaluronan may play an active role in modulating RHAMM function, a notion supported by an earlier study showing that blocking antibodies to RHAMM can block the binding of fluorescein-hyaluronan intracellularly (Pilarski et al. 1999).
At the other extreme, one could argue that the associations described here are simply the result of stochastic processes of organelle segregation during mitosis. However, this does not explain why the cell would need such a rapid increase in intracellular hyaluronan at this time. Furthermore, hyaluronan was also frequently found around the concentrated bundles of microtubules known as stem bodies (Rieder and Khodjakov 2003) that are present in the bridges between daughter cells in the area of the cleavage furrow (Figure 6), further suggesting that hyaluronan may be involved in microtubule function. Stem bodies are thought to be important in cytokinesis (Rieder and Khodjakov 2003). Previously, hyaluronan in the pericellular matrix at the cleavage furrow has been proposed to facilitate cytokinesis (Tammi and Tammi 1991). Thus, it is possible that the intracellular hyaluronan also serves some biophysical function in this location.

Colocalization of hyaluronan and RHAMM in the mitotic spindle. (
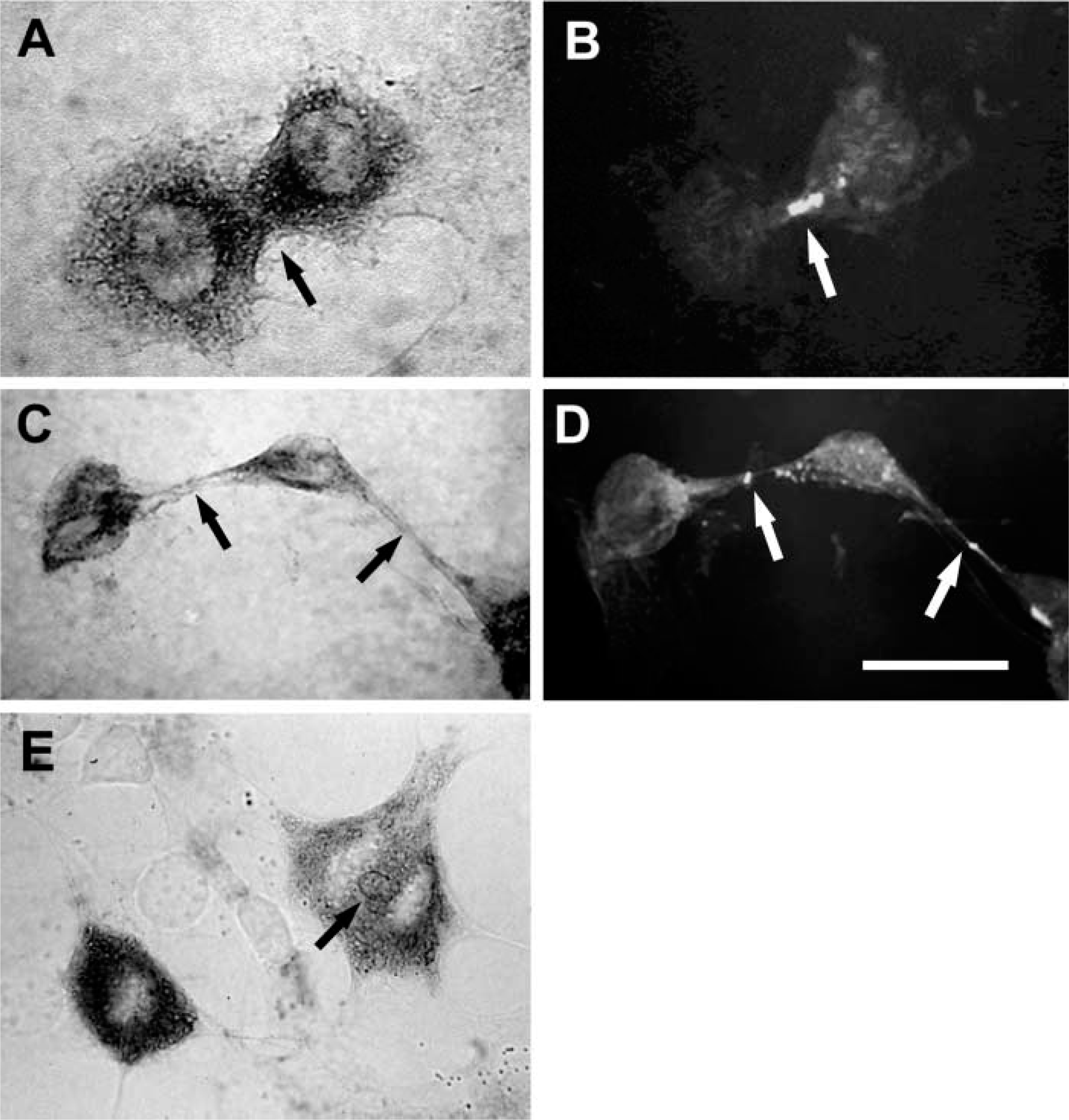
Hyaluronan is associated with midbody microtubules during cytokinesis. (
To our knowledge, this is the first report directly showing a close spatial relationship of intracellular hyaluronan with an intracellular hyaladherin and supports the idea that hyaluronan may influence the function of intracellular RHAMM (Hall et al. 1994; Collis et al. 1998; Hofmann et al. 1998; Pilarski et al. 1999). RHAMM was previously found in the perinuclear microtubule network of interphase cells and in the mitotic spindle (Assmann et al. 1999; Haddad and Turley 2000). We also found that microtubule-associated RHAMM is available for binding to fluorescein-hyaluronan, suggesting that RHAMM is not saturated with endogenous hyaluronan, consistent with an earlier study in hematopoietic cells (Pilarski et al. 1999). Thus, RHAMM may be an actual intracellular binding partner for the hyaluronan here. Hyaluronan interacts with phospholipid (Pasqualli-Ronchetti et al. 1997) and is extruded through the membrane during biosynthesis. Therefore, it is not inconceivable that hyaluronan within vesicles or other membranous compartment has the capacity to interact with RHAMM. The microtubule-binding domain of RHAMM is located in the N-terminal portion (Assmann et al. 1999) and the hyaluronan-binding sequence is in domain 5, closer to the C terminus (Yang et al. 1994). Thus, hyaluronan conceivably could bridge two RHAMM molecules while they are associated with interphase microtubules. On the other hand, the carboxy-terminal leucine zipper targets RHAMM to the centrosome (Maxwell et al. 2003) and overlaps with the hyaluronan-binding site, suggesting that hyaluronan could also play a competitive role in regulating mitosis. Hyaluronan staining has been previously noted to penetrate the nucleus in the prometaphase stage (Evanko and Wight 1999), where spindle formation and chromosome alignment occur. RHAMM is known to regulate ERK kinase activity (Zhang et al. 1998) and participates in hyaluronan-induced cell locomotion and the uptake and translocation of hyaluronan to the nucleus (Collis et al. 1998). RHAMM and hyaluronan promote focal adhesion turnover (Hall et al. 1994) and repeated contact of microtubules with focal adhesions promotes their disassembly (Kaverina et al. 2002). RHAMM was recently found to interact with dynein and help maintain spindle pole stability (Maxwell et al. 2003). Therefore, hyaluronan may regulate the effects of RHAMM on microtubule dynamics in various ways. RHAMM could mediate the binding of hyaluronan-containing vesicles to microtubules, and together they could serve in an as-yet-undefined biophysicalstructural role during the mitotic process. Alternatively, hyaluronan could regulate the amount of available RHAMM for interacting with the dynein motor complex or other signaling molecules such as ERK kinase.
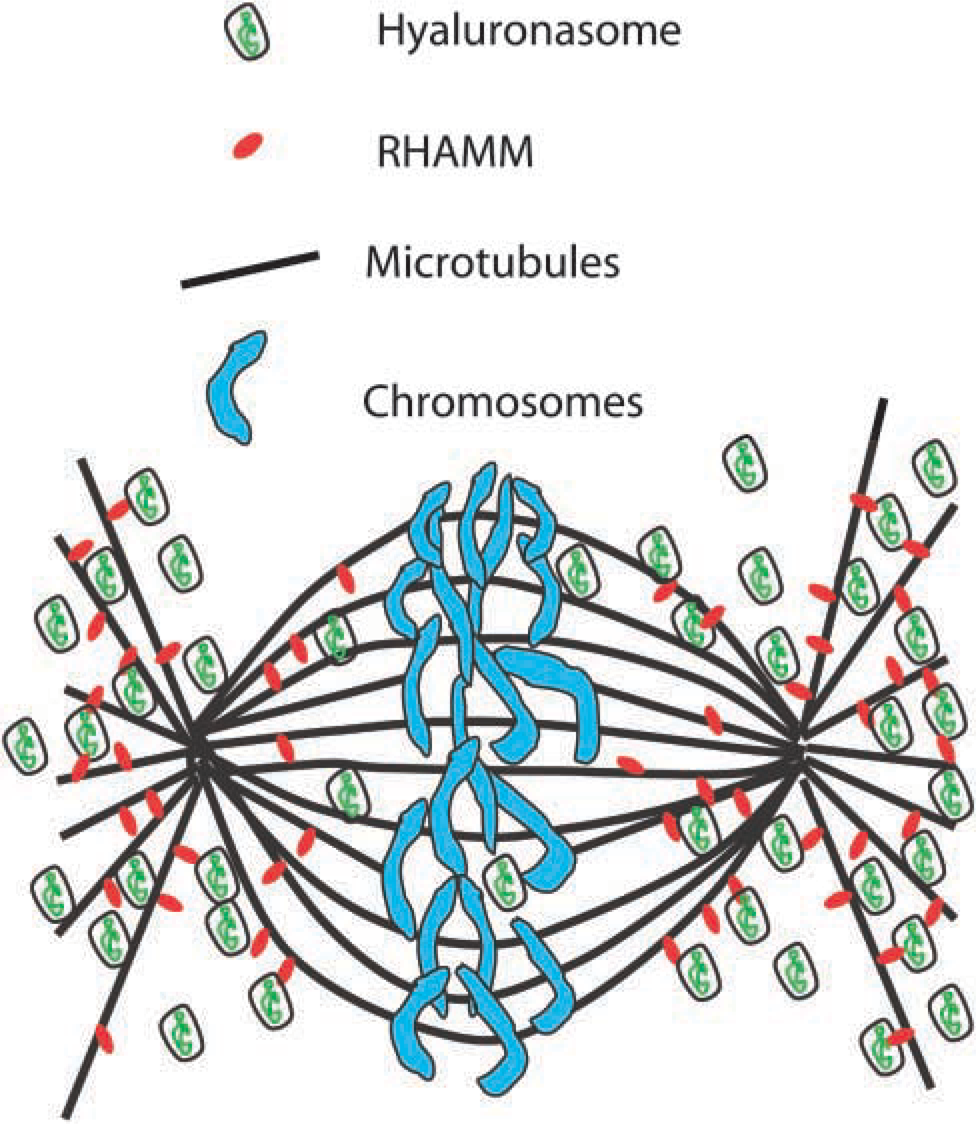
Model depicting the relationship of hyaluronan and RHAMM to the mitotic spindle. Through interaction with RHAMM, hyaluronan could affect microtubule spacing, lateral stability, or facilitate movements involving microtubule-based motor complexes. Hyaluronan may also influence RHAMM signaling events.
It is clear that the biological activity of hyaluronan is size dependent. We found that fragmented fluorescein-labeled hyaluronan was preferentially translocated to the perinuclear area (Figure 1). In contrast, although some high molecular weight fluorescein-hyaluronan that was taken up by cells could be seen in the perinuclear area (Figure 1A) and the mitotic spindle (Figure 5G), it was mostly present in large endosomes. This extends previous studies showing that most of the intracellular hyaluronan is of low molecular weight, is present in vesicles of varying size (100–1300 nm) and shape, and may be destined for eventual degradation (McGuire et al. 1987; Tammi et al. 2001). Various mammalian hyaluronidases and hyaluronidase inhibitors that may be central to regulating hyaluronan size and catabolism have been characterized (Stern and Csoka 2000). Given the discrete size of the hyaluronan inside cells, one can conceive of possible mechanisms whereby hyaluronan or the “hyaluronasomes” themselves could serve as novel structural/functional units. Clearly, the study of the functional and regulatory roles of intracellular hyaluronan is rapidly expanding and the testing of these and other ideas will be forthcoming. The present observations provide an additional basis for further investigation into this area.
Footnotes
Acknowledgements
This work was supported in part by NIH Grant HL-18645 (TNW).
We would like to thank Dr Volker Assmann (Richard Dimbleby Department of Cancer Research/ICRF Laboratory, St Thomas' Hospital, London, UK) for the anti-RHAMM antibody, and Dr Robert Stern (University of San Francisco) for helpful discussions.