
Abstract
Hyaluronan is a high molecular weight glycosaminoglycan found in the extracellular matrix of many tissues, where it is believed to promote cell migration and proliferation. It was recently shown that hyaluronan-dependent pericellular matrix formation is a rapid process that occurs as cells detach during mitosis. Growing evidence for intracellular hyaluronan in tissues in vivo, together with evidence of intracellular hyaluronan binding molecules, prompted us to examine hyaluronan distribution and uptake as well as hyaluronan binding sites in cells and their relationship to cell proliferation in vitro, using a biotinylated hyaluronan binding protein and fluorescein-labeled hyaluronan. In permeabilized smooth muscle cells and fibroblasts, hyaluronan staining was seen in the cytoplasm in a diffuse, network-like pattern and in vesicles. Nuclear hyaluronan staining was observed and confirmed by confocal microscopy and was often associated with nucleoli and nuclear clefts. After serum stimulation of 3T3 cells, there was a dramatic increase in cytoplasmic hyaluronan staining, especially during late prophase/early prometaphase of mitosis. In contrast, unstimulated cells were negative. There was a pronounced alteration in the amount and distribution of hyaluronan binding sites, from a mostly nucleolar distribution in unstimulated cells to one throughout the cytoplasm and nucleus after stimulation. Exogenous fluorescein-labeled hyaluronan was taken up avidly into vesicles in growing cells but was localized distinctly compared to endogenous hyaluronan, suggesting that hyaluronan in cells may be derived from an intracellular source. These data indicate that intracellular hyaluronan may be involved in nucleolar function, chromosomal rearrangement, or other events in proliferating cells.
H
In addition to its role in the extracellular matrix, evidence is growing that hyaluronan is also present in the cytoplasm and nuclei of cells in a number of tissues in vivo (Margolis et al. 1976; Furukawa and Terayama 1977, 1979; Londono and Bendayan 1988; Ripellino et al. 1989, 1989; Kan 1990; Eggli and Graber 1995). Furthermore, recent studies have shown that there are a number of intracellular hyaluronan binding proteins that may be important in regulation of the cell cycle or in gene transcription (Grammatikakis et al. 1995; Deb and Datta 1996; Hofmann et al. 1998; Zhang et al. 1998). For example, a vertebrate homologue of the cell cycle control protein CDC37 was recently cloned and found to bind hyaluronan (Grammatikakis et al. 1995), and an intracellular form of the hyaluronan receptor, RHAMM, was shown to regulate erk kinase activity (Zhang et al. 1998).
This study examined the intracellular localization of hyaluronan and hyaluronan binding sites, as well as hyaluronan uptake, and their relationship to cell proliferation in vitro. The results show that there is an increase in the amount of intracellular hyaluronan and a redistribution of hyaluronan binding sites that are associated specifically with mitosis, and provide evidence that cytoplasmic and nuclear hyaluronan may be derived from intracellular sources.
Materials and Methods
Cell Culture
Human arterial smooth muscle cells (SMCs) derived from explants of human aortic tissue were kindly provided by Dr. Russell Ross (Department of Pathology, University of Washington). Human skin fibroblasts were provided by Dr. Peter Byers (Department of Pathology, University of Washington). Swiss 3T3 D1 cells were provided by Dr. Daniel Bowen-Pope (Department of Pathology, University of Washington). All cells were maintained in DMEM containing 10% fetal bovine serum. For cytochemical localization, cells were seeded on coverslips in 35-mm tissue culture dishes at 2 × 105 cells per dish. In certain experiments, 3T3 cells or SMCs were made quiescent by switching the medium to DMEM containing 0.5% fetal bovine serum for 48 hr and were then stimulated by readdition of serum to 10% or addition of 10 ng/ml PDGF AB. Cells were prepared for hyaluronan localization as described below at 12 hr, 24 hr, and 27 hr after serum addition to correlate with the times that cells are expected to be in G1, S, and G2/M of the cell cycle (not shown). To examine hyaluronan internalization, fluorescein-labeled hyaluronan (1.65 µg/ml) was added to the medium of stimulated and unstimulated cells after serum addition and was incubated for 24 hr. Cells were then stained for endogenous hyaluronan as described below.
Localization of Hyaluronan and Hyaluronan Binding Sites
Cells on coverslips were digested with 2 U/ml Streptomyces hyaluronidase (Sigma; St Louis, MO) in DMEM at 37C for 1 hr before fixation to remove pericellular hyaluronan, which interfered with the visualization of intracellular hyaluronan. After rinsing with PBS, the cultures were fixed by addition of 10% neutral buffered formalin directly to the medium to give a final concentration of 2.5% formalin, for 10 min at 22C. Cells were rinsed three times with PBS and permeabilized with 0.5% Triton X-100 in PBS for 10 min.
To localize hyaluronan, cells were stained with a biotinylated probe consisting of a mixture of cartilage proteoglycan core protein and link protein (kindly provided by Charles Underhill; Department of Anatomy and Cell Biology, Georgetown University) at a concentration of 2 μg/ml. This is a commonly used probe that specifically recognizes hyaluronan in tissues and cultured cells (Underhill et al. 1993). Controls for intracellular hyaluronan staining included staining of nonpermeabilized cells, digestion of the cells with Streptomyces hyaluronidase after permeabilization (4 U/ml in DMEM, 1 hr, 37C), and preincubation of the biotinylated probe with 100 μg of hyaluronan. The biotinylated probe was applied in PBS containing 1% BSA for 1 hr at 22C. After rinses, the samples were incubated with a 1:500 dilution of streptavidin-horseradish peroxidase conjugate (Kirkegaard & Perry; Gaithersburg, MD). Color was developed for 10 min using 3-amino-9-ethylcarbazole (AEC) in 0.05 M sodium acetate, pH 5.0. The substrate solution was freshly prepared by dissolving 2 mg AEC in 0.5 ml of DMSO and adding 10 ml of 0.05 M sodium acetate, pH 5.0. Ten µl of 30% H2O2 was added just before use. Cells were mounted with Crystal Mount (Biomeda; Foster City, CA). Alternatively, the biotinylated probe was detected with a 1:500 dilution of streptavidin conjugated to either Texas Red or FITC (Zymed; South San Francisco, CA) and mounted with Gel Mount (Biomeda). For confocal analysis, nuclei were counterstained with DAPI at 1 μg/ml in H2O for 10 min at 22C (see Figure 4).
To compare the distribution of endogenous hyaluronan with exogenous hyaluronan taken up by cells by endocytosis, 3T3 cells on coverslips were made quiescent by incubation in medium containing 0.2% fetal bovine serum for 48 hr, and then some cultures were stimulated to proliferate by addition of serum to 10%. Fluorescein-labeled hyaluronan was added (5 μg/ml) and cells were allowed to take up the labeled hyaluronan for 24 hr. Cells were then fixed and stained for endogenous intracellular hyaluronan using the biotinylated probe and streptavidin-Texas Red.
To localize intracellular hyaluronan binding sites, fixed and permeabilized cells were incubated with fluorescein-labeled hyaluronan (5 μg/ml) for 10 min. after having stained the endogenous intracellular hyaluronan with Texas Red. As a control, binding of the labeled hyaluronan was blocked by preincubation of the cells with excess unlabeled hyaluronan (3 mg/ml). Additional controls included digestion of the fluorescein-hyaluronan with Streptomyces hyaluronidase (20 U/ml in PBS, 37C, 18 hr), chondroitinase ABC (10 U/ml, 37C, 18 hr), or pronase (200 μg/ml, 37C, 18 hr), preincubation and staining of the cells in the presence of chondroitin sulfate (10 mg/ml), and staining of the cells with free fluoresceinamine (see Figure 7). Cells were examined with a Zeiss Axioskop photomicroscope equipped for epifluorescence or with an ACAS Ultima confocal laser scanning microscope with UV and 488-nm excitation to view DAPI and FITC, respectively.
Preparation of Fluorescein-labeled Hyaluronan
Hyaluronan was labeled with fluorescein according to published methods (de Belder and Wik 1975). Hyaluronan (10 mg, Grade I; Sigma) was dissolved in 8 ml dH2O and added to 4 ml DMSO. A mixture of acetaldehyde (5 μl)/cyclohexylisocyanide (5 μl)/fluoresceinamine (5 mg; all from Aldrich, Milwaukee, WI) in 0.3 ml DMSO was added to the hyaluronan and the mixture was incubated at room temperature with stirring for 5 hr. This was added to 160 ml cold 100% ethanol and precipitation was performed with ∼2 ml saturated NaCl. Precipitates were centrifuged at 3000 rpm for 2 min, pooled in 10 ml dH2O, and added to 100 ml cold 100% ethanol. Centrifugation was repeated, and precipitates were resuspended in 5 ml dH2O and dialyzed exhaustively against PBS, 0.2% sodium azide. The final preparation had an average molecular weight of approximately 500 kD and contained approximately 0.8 μg fluorescein/μg of uronate.
Results
Intracellular Localization of Hyaluronan
A biotinylated hyaluronan binding probe was used for localization of hyaluronan in human aortic smooth muscle cells and skin fibroblasts in vitro. In nonpermeabilized cells, hyaluronan staining was prominent on the cell surface and in the pericellular matrix as a tangled flocculent network (Figure 1A). Digestion of the cells with Streptomyces hyaluronidase before fixation abolished the pericellular matrix and cell surface staining (Figure 1B). Occasionally, a small amount of residual punctate staining of the cell surface was visible. Cells that had been pretreated with Streptomyces hyaluronidase to remove extracellular hyaluronan were permeabilized by treatment with 0.5% Triton X-100 and then stained with the hyaluronan probe. In the permeabilized cells, there was prominent cytoplasmic staining, which appeared both in round vesicular structures, probably representing endocytotic vesicles (see below), and also in a diffuse pattern throughout the cytoplasm (Figure 1C). The cytoplasmic staining was often particularly prominent in the perinuclear region. In SMC cultures, approximately 75% of the cells exhibited positive cytoplasmic staining. However, there were marked variations in the intensity of staining among hyaluronan-positive cells. Interestingly, cells having similar staining intensities were often seen in small groupings (not shown). Digestion of the cells with hyaluronidase a second time, after permeabilization, entirely abolished the intracellular staining, confirming the specificity of the hyaluronan probe (Figure 1D). Preincubation of the hyaluronan binding probe with excess hyaluronan also abolished staining (not shown).
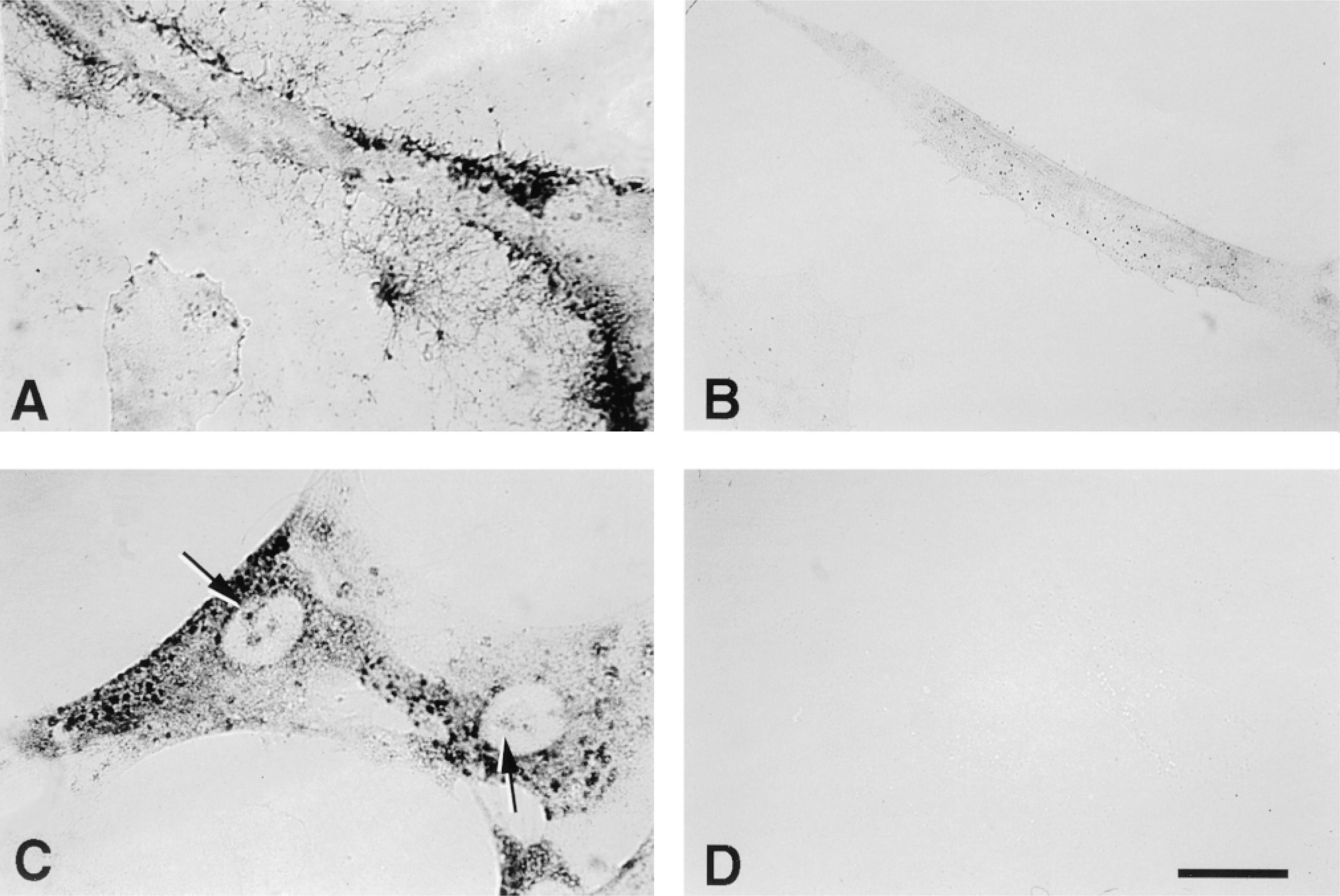
Intracellular and nuclear hyaluronan. Histochemical staining for hyaluronan in human SMCs using the biotinylated hyaluronan binding region of aggrecan and the peroxidase substrate AEC in (A) intact cells, (B) cells treated with Streptomyces hyaluronidase before fixation, (C) cells treated with hyaluronidase and then permeabilized, and (D) cells treated again with hyaluronidase after permeabilization. Arrows in C indicate nucleolar staining. Bar = 25 μm.
In addition to cytoplasmic staining, apparent nuclear localization was also frequently seen. In SMCs, the nuclear staining often appeared to be associated with nucleoli (Figure 1?; see also below). Hyaluronan staining occasionally could also be seen as fine filamentous material throughout the nuclei, but this was difficult to resolve on photomicrographs.
The hyaluronan localization described above employed a diffusable chromogenic substrate. One concern was that the diffuse staining in the cytoplasm, as well as the perinuclear and nucleolar staining, resulted from diffusion of the chromogen away from the cytoplasmic vesicles. Therefore, we used streptavidin conjugated to Texas Red or FITC as a nondiffusable marker to localize the biotinylated hyaluronan binding probe. An identical hyaluronidase-sensitive pattern was observed with the fluorescent markers (Figure 2), indicating that the fine network-like cytoplasmic staining observed with the peroxidase substrate did not result from diffusion.
In permeabilized human skin fibroblasts, hyaluronan staining in the cytoplasm was seen in a similar pattern as in the SMCs, i.e., both vesicular and diffuse cytoplasmic staining (Figure 3). However, there was a noticeably different pattern of nuclear staining in the fibroblasts. In addition to occasional nucleolar staining, there was a clear association of hyaluronan staining with distinct clefts and furrows in the fibroblast nuclei.
Although many examples of distinct nucleolar staining were observed, it was possible that this was actually hyaluronan in vesicles positioned over or under the nucleus. However, confocal microscopy, which resolves the fluorescent signal in adjacent 1.0-μm-thick optical sections, demonstrated that the hyaluronan staining (green label) was in the same focal plane as the nuclear DAPI staining, confirming the nuclear localization (Figure 4).
Possible Intracellular Source of Hyaluronan
To determine whether the cytoplasmic and/or nuclear hyaluronan resulted from translocation of extracellular hyaluronan or if it might be derived from intracellular sources, either quiescent, serum-starved 3T3 cells or cells stimulated with 10% fetal bovine serum were allowed to take up fluorescein-labeled hyaluronan for 24 hr and then were fixed, permeabilized, and stained for intracellular hyaluronan with Texas Red-streptavidin. In the quiescent control cells, uptake of fluorescein-labeled hyaluronan was minimally detectable, was in small vesicles, and was seen only in a few scattered cells (Figure 5A). Interestingly, hyaluronan staining with the hyaluronan binding probe in the same control cells was only occasionally detectable and was confined to the same vesicles that contained the fluorescent hyaluronan, but was negative in the cytoplasm and nuclei. This indicates that fluorescein remains associated with hyaluronan after internalization and further confirms the specificity of the hyaluronan binding probe. This also supports the idea that the positively stained vesicular structures shown in Figure 1 contain internalized endogenous hyaluronan. By contrast, in the serum-stimulated cells there were both prominent cytoplasmic staining for hyaluronan and increased uptake of fluorescein-labeled hyaluronan into endosomal vesicles (Figure 5B). However, the exogenous fluoresceinated hyaluronan was confined to the endosomes and did not co-localize with the endogenous hyaluronan network in the cytoplasm, suggesting that some of the cytoplasmic hyaluronan may be derived from intracellular sources and not from translocation of extracellular hyaluronan. Interestingly, uptake of the fluoresceinated hyaluronan occurred to the greatest extent in cells with a stellate or motile fusiform morphology, consistent with a recent study showing rapid uptake of hyaluronan in motile cells (Collis et al. 1998). Previous studies have shown that hyaluronan-dependent pericellular coat formation was also more prominent in cells with this morphology (Evanko et al. 1999). In contrast, the highly spread and flattened cells did not take up much hyaluronan (present study) and they do not form hyaluronan coats. In cells that appeared to be in mitosis, the green endosomal vesicles containing exogenous hyaluronan appeared to be distributed relatively equally to the daughter cells after division (not shown).
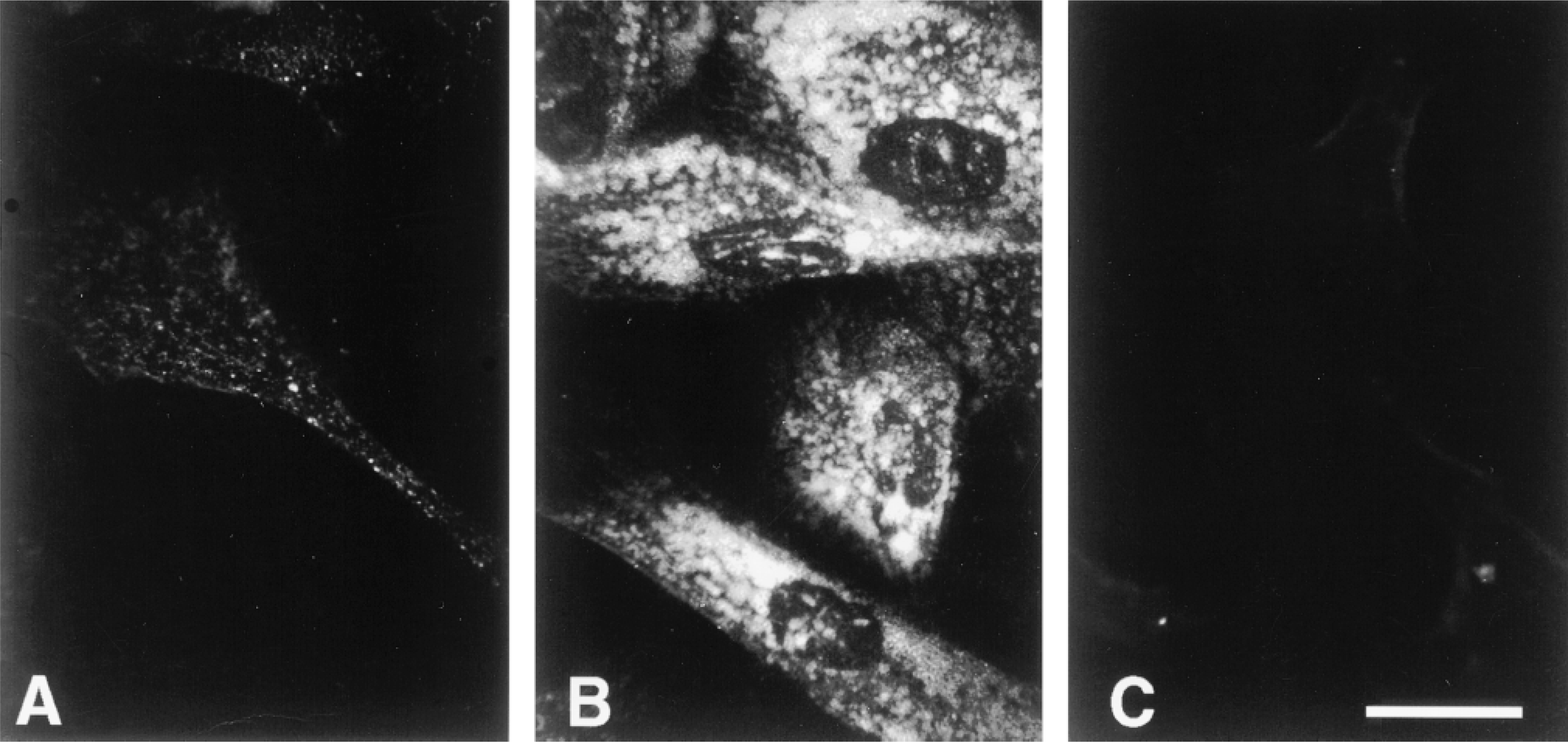
Intracellular hyaluronan in human skin fibroblasts. Histochemical staining for hyaluronan using Texas Red-streptavidin to detect the hyaluronan probe in (A) cells treated with Streptomyces hyaluronidase before fixation, (B) cells treated with hyaluronidase and then permeabilized, and (C) cells treated again with hyaluronidase after permeabilization. Bar = 25 μm.
Distribution of Hyaluronan Binding Sites
In a related experiment, the fluorescein-labeled hyaluronan was used as a probe to localize intracellular hyaluronan binding sites in permeabilized cells. In this case, fixed and permeabilized cells were first stained for endogenous intracellular hyaluronan using the biotinylated hyaluronan binding probe and Texas Red-streptavidin, and then incubated with fluorescein-labeled hyaluronan. In the quiescent control cells, fluorescein-labeled hyaluronan bound to a fine, lace-like network in the cytoplasm and, most prominently, to nucleoli in the nucleus (Figures 5C and 5G). As above, there was little or no staining for endogenous hyaluronan. Incubation of the cells with excess unlabeled hyaluronan (3 mg/ml) dramatically diminished binding of the labeled hyaluronan (Figures 5D and 6C). Additional controls confirmed the specificity of this binding (Figure 6). For example, chondroitin sulfate had negligible effect on the binding, even at a threefold higher concentration (10 mg/ml). In addition, exhaustive digestion of the labeled hyaluronan with Streptomyces hyaluronidase (Figure 6) or chondroitinase ABC (not shown) dramatically diminished the binding to permeabilized cells. Pronase digestion of the fluorescein-hyaluronan preparation had no effect, indicating that the binding was not due to any protein contaminants in the preparation. In addition, no binding was seen with free fluorescein.
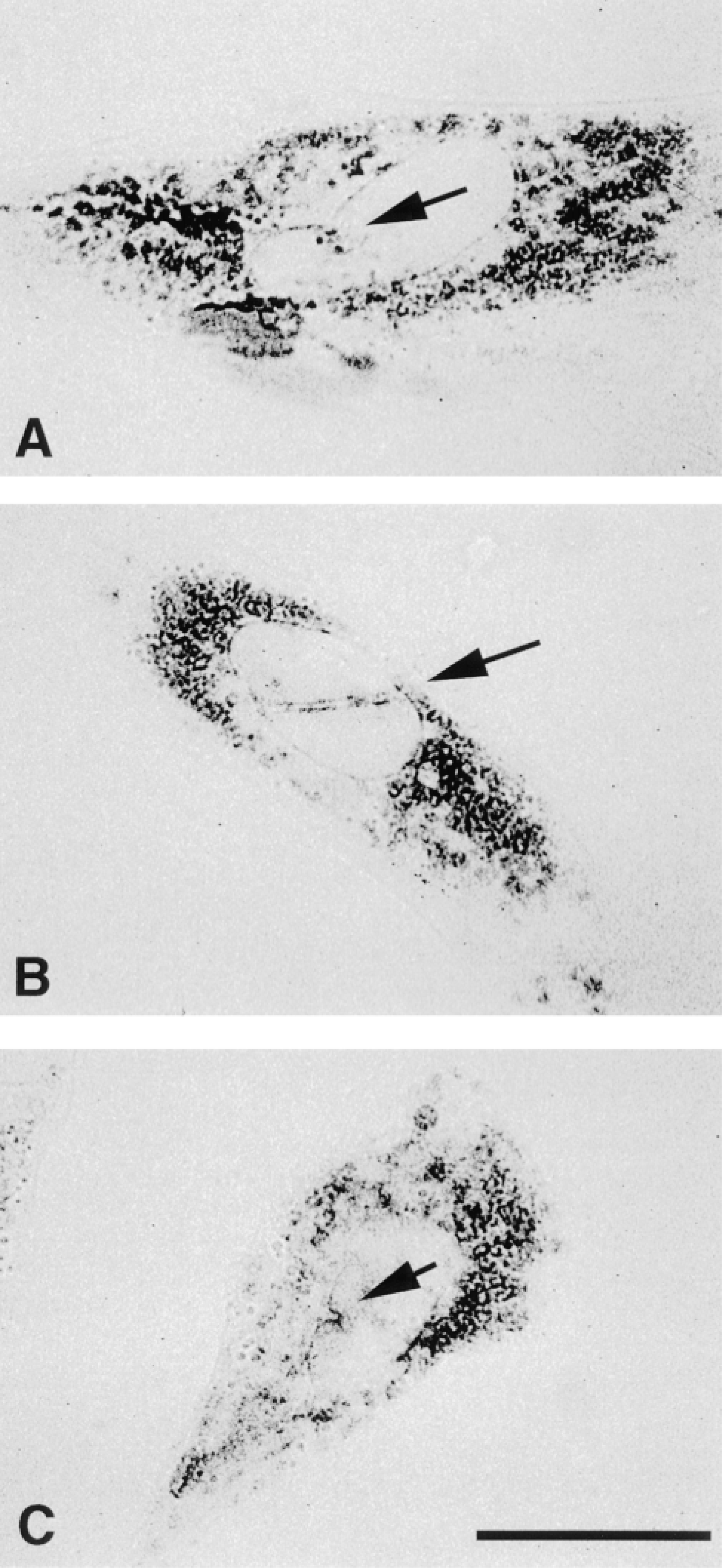
Hyaluronan is associated with nuclear clefts in fibroblasts. (A-C) Representative examples of human skin fibroblasts stained for intracellular hyaluronan using the red chromogen AEC. Arrows indicate clefts and furrows in the nuclei with associated hyaluronan staining. Bar = 25 μm.
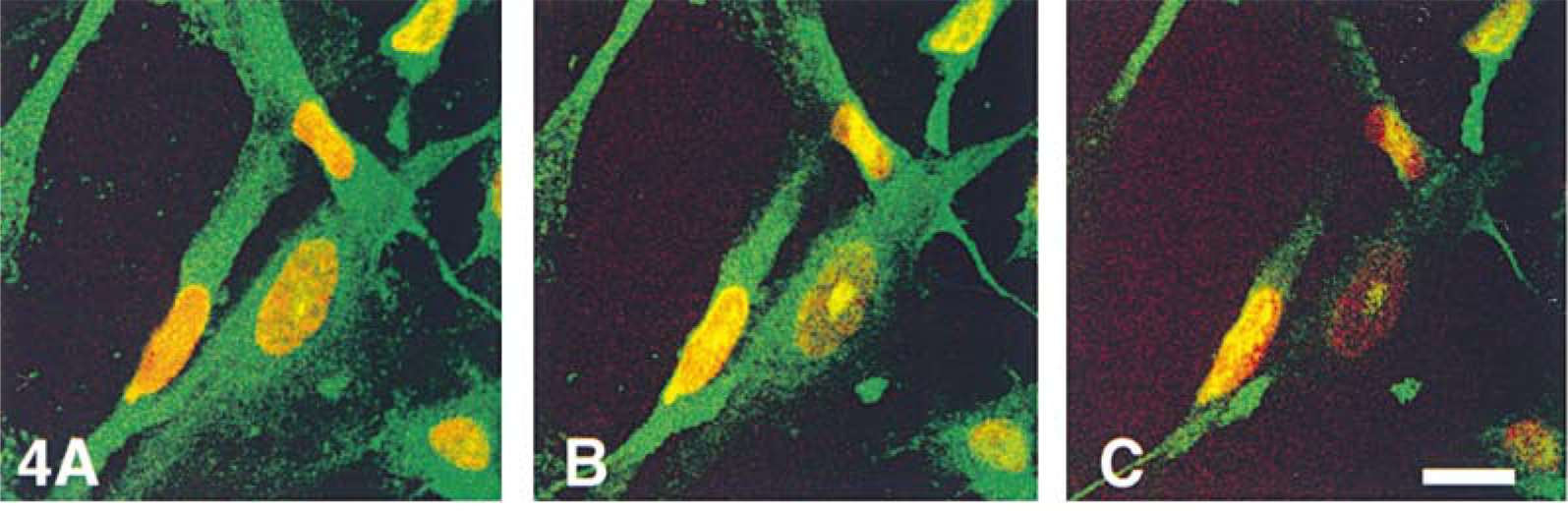
Hyaluronan is present in the nucleus of smooth muscle cells. (A-C) Confocal microscopic “z series” of three adjacent optical sections of SMCs stained for intracellular hyaluronan. Hyaluronan was detected with FITC-avidin (green); the nuclei were stained with DAPI. The images were pseudo-colored using Adobe Photoshop, and the DAPI-stained nuclei (originally blue) were converted to red to enhance the contrast with the green hyaluronan. Yellow areas indicating co-localization of hyaluronan and nuclear staining in the same optical section (1 μm thick) can be seen both centrally and peripherally in some nuclei. In this figure endogenous hyaluronan is green. Bars = 25 μm.
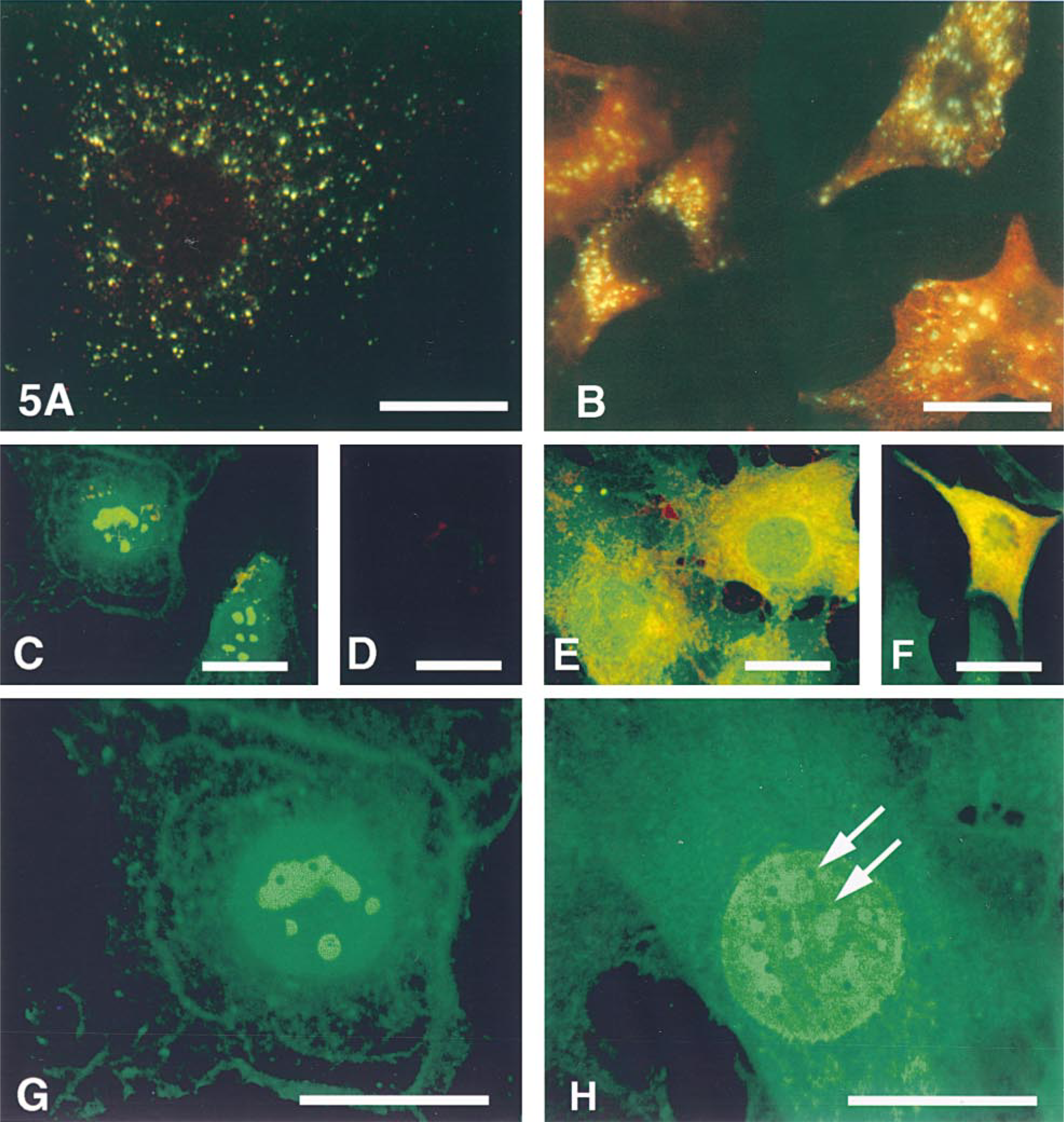
Hyaluronan uptake, intracellular hyaluronan, and hyaluronan binding sites are altered after serum stimulation. Control and serum-stimulated 3T3 cells were allowed to take up fluorescein-labeled hyaluronan (green) for 24 hr and then were fixed and stained for endogenous intracellular hyaluronan (red). There is only a small amount of uptake of the fluorescein-labeled hyaluronan into vesicles in quiescent cells, and no cytoplasmic staining for endogenous hyaluronan (A), whereas the stimulated cells (B) show both enhanced uptake of fluorescein-labeled hyaluronan and intense hyaluronan staining in the cytoplasm. The localization of endogenous hyaluronan is distinct from that in endosomal vesicles. (C-F) Intracellular hyaluronan binding sites were detected with fluorescein-labeled hyaluronan (green) after staining of permeabilized cells for endogenous hyaluronan (red). In control cells (C), the fluorescein-labeled hyaluronan bound primarily to nucleoli and slightly to a cytoplasmic network. Preincubation of cells with unlabeled hyaluronan abolished staining with the labeled hyaluronan (D). In serum-stimulated cells (E,F), the fluorescein-labeled hyaluronan bound extensively in the cytoplasm, throughout the nucleus, and to the nuclear periphery, and co-localized with endogenous cytoplasmic hyaluronan. Compare the mitotic cell in prometaphase (F) with Figure 7C. Higher magnification shows more detail of the binding of fluorescein-hyaluronan to nucleoli in control cells (G) and the increased binding to the cytoplasmic network and nucleus in serum-stimulated cells (H). Only the fluorescein label is shown in G and H. Arrows indicate possible nuclear pores, negatively stained by fluorescein-hyaluronan at the nuclear periphery. Endogenous hyaluronan is red. Bars = 25 μm.
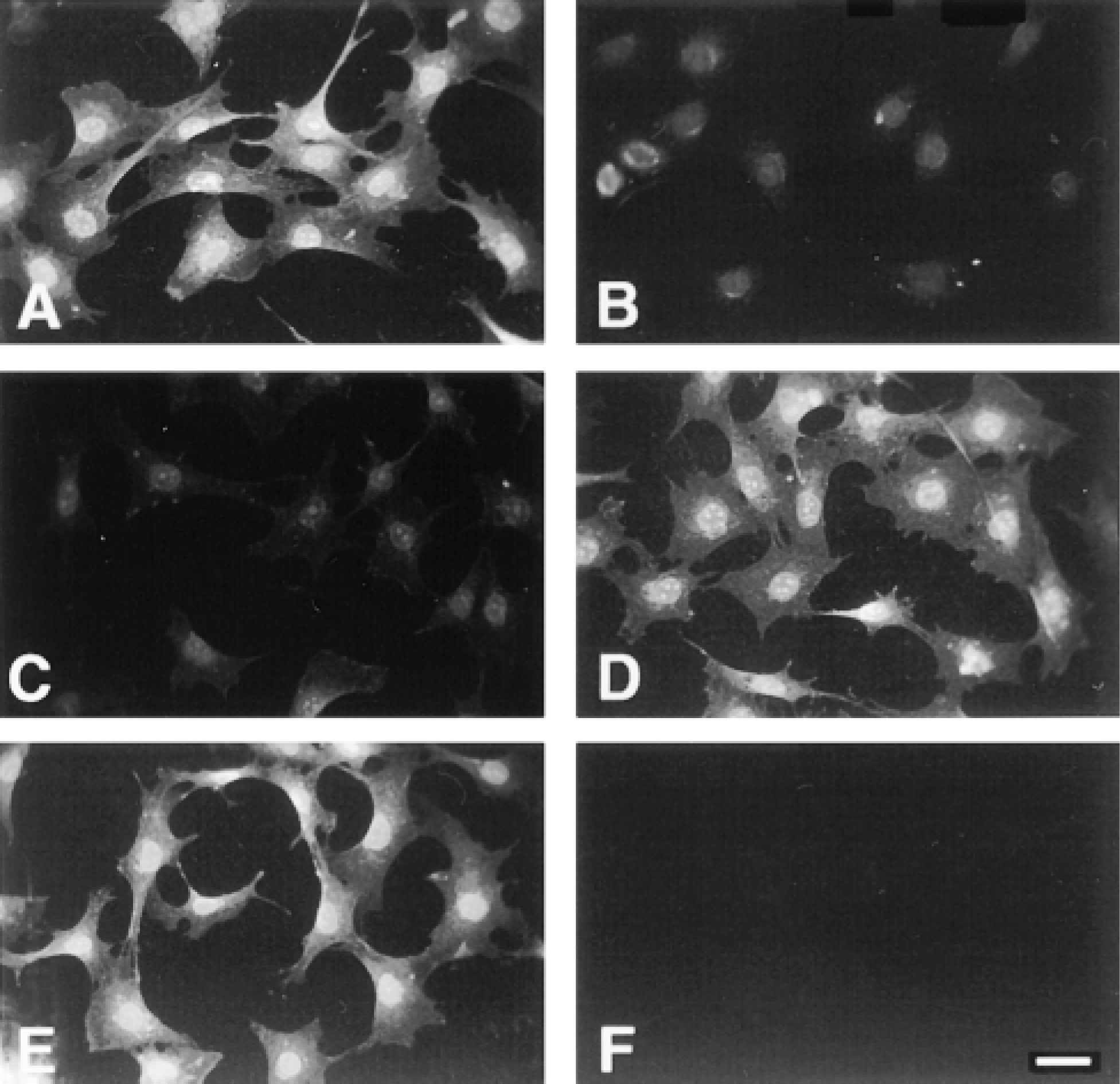
Specificity of hyaluronan binding to intracellular sites. Staining of permeabilized 3T3 cells with (A) fluorescein-labeled hyaluronan alone, (B) fluorescein-labeled hyaluronan previously digested with Streptomyces hyaluronidase, (C) fluorescein-labeled hyaluronan after preincubation of the cells with unlabeled hyaluronan (3 mg/ml) or (D) chondroitin sulfate (10 mg/ml), (E) fluorescein-labeled hyaluronan digested with pronase or (F) free fluoresceinamine. Bar = 50 μm.
In contrast to quiescent cells, there was a pronounced increase in the binding of fluorescein-labeled hyaluronan to the reticular network in the cytoplasm in serum-stimulated cells, indicating an increase in the amount of available binding sites (Figures 5E, 5F, and 5H). In stimulated cells, the bound fluorescein-hyaluronan (green) co-localized to a large extent with the staining of the endogenous hyaluronan network (red label) in the cytoplasm of both premitotic (Figure 5E) and mitotic cells (Figure 5F), but not in the endosomal vesicles. Similar results were seen in fibroblasts and SMCs (not shown). Interestingly, in mitotic cells the fluorescent hyaluronan bound to sites between the chromosomes and was similar to the distribution of endogenous hyaluronan (also see below; compare cells in prometaphase in Figures 5F and 7C). Futhermore, in marked contrast to the control cells, in which the the fluorescein-labeled hyaluronan bound primarily to nucleoli, the fluorescent hyaluronan bound extensively throughout the nucleus and/or to the nuclear periphery in the stimulated cells. A higher-magnification view, shown in Figures 5G and 5H, compares the extent to which fluorescein-labeled hyaluronan bound to the reticular network in the cytoplasm and to the nucleus in control vs stimulated cells. Interestingly, in many of the nuclei in the stimulated cells there were many round, uniformly sized dark spots, perhaps representing nuclear pores, that presumably were made apparent by negative staining with the fluoresceinated hyaluronan which bound to the nuclear periphery. This may represent binding to areas of condensed chromatin (Kan 1990), which is excluded from the nuclear pore region (Alberts et al. 1989). These data suggest that, together with an increase in cytoplasmic hyaluronan, there is a pronounced shift in the amount and distribution of hyaluronan binding sites in cells after mitogenic stimulation by serum.
Association of Intracellular Hyaluronan with Mitosis
As described above, there appeared to be a striking increase in the amount of intracellular hyaluronan and hyaluronan binding sites in serum-stimulated cells. It was previously demonstrated that formation of hyaluronan-dependent extracellular matrix occurs in the pericellular space just before the detachment phase of mitotic cell rounding in vitro (Evanko et al. 1999). Similar association of pericellular hyaluronan with mitosis has been observed in vivo (Tammi and Tammi 1991), and others have shown that the sythesis of hyaluronan is elevated during G1, and especially G2, of the cell cycle (Brecht et al. 1986). Therefore, we wondered if intracellular hyaluronan was also associated with particular stages of the cell cycle. 3T3 cells were used because of the degree to which they can be synchronized by serum deprivation.
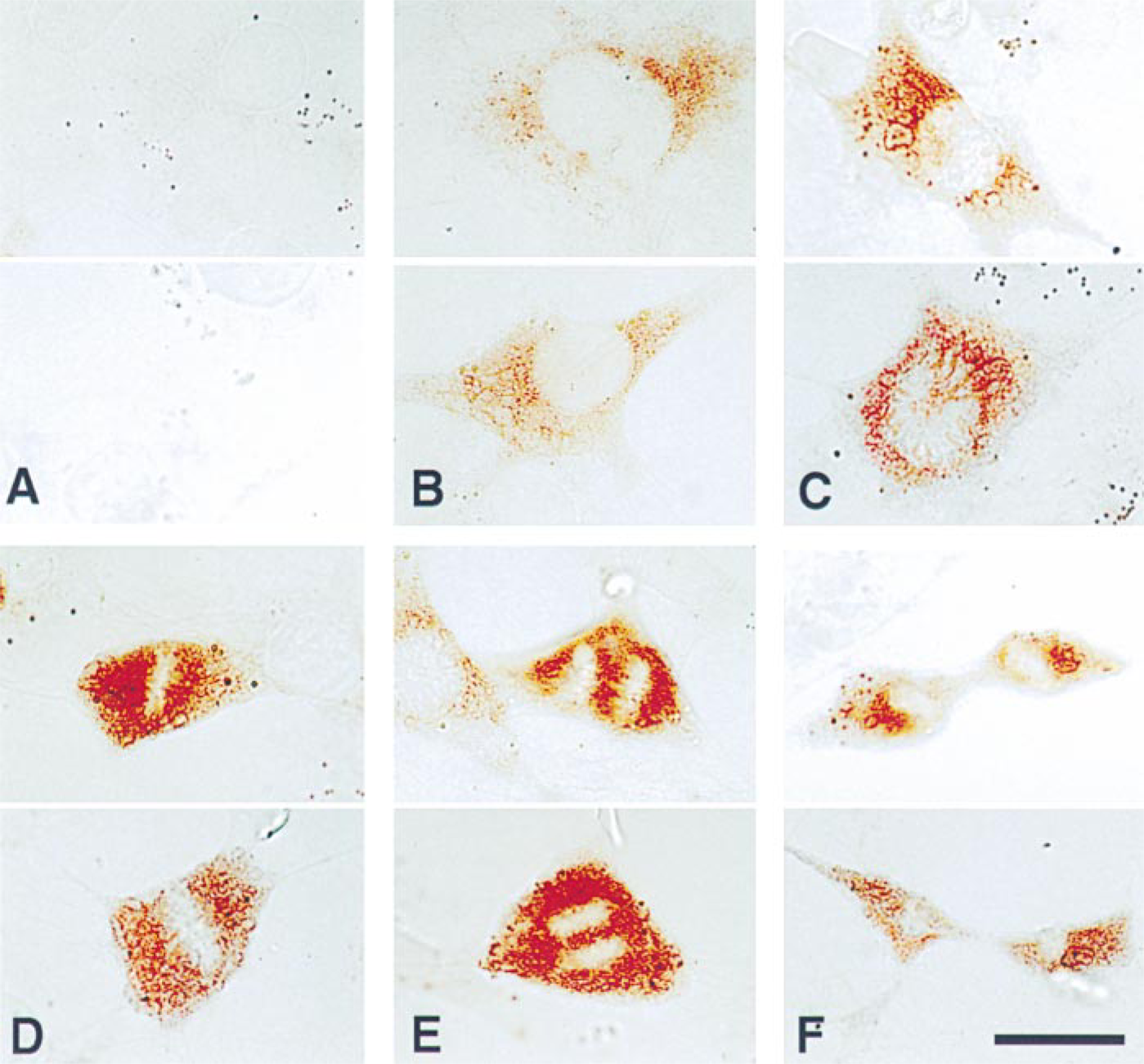
Intracellular hyaluronan is associated with mitosis. Examples of hyaluronan staining in permeablized 3T3 cells after serum stimulation. (A) In control interphase cells, no staining is present. (B) At 24 hr after serum stimulation, hyaluronan has accumulated in the cytoplasm and is concentrated near the nucleus. (C-F) Intense staining is seen in all mitotic cells at 27 hr after serum stimulation. In late prophase/early prometaphase (C), chromosomes are arranged radially around a central area and staining can be seen between chromosomes. In metaphase (D), hyaluronan fills the cell and surrounds the chromosomes at the metaphase plate. In anaphase (E), staining is present between the chromosomes as they move to opposite poles of the cell. Strong staining persists through telophase (F). Bar = 25 μm.
Virtually no intracellular hyaluronan staining was observed in serum-starved quiescent cells, i.e., G0/interphase (Figure 7A). After serum stimulation, hyaluronan staining was detectable by 12 hr, when the cells are expected to be in G1 of the cell cycle, and was seen as a diffuse network throughout the cytoplasm and concentrated around the nucleus (not shown). Staining increased gradually in almost all cells and, compared to 12 hr, was clearly stronger at 24 hr (Figure 7B), which is just after the peak of DNA synthesis in these cells. Most notably, at 27 hr there was a striking increase in the intensity of cytoplasmic hyaluronan staining specifically in the mitotic cells. In late pro-phase/early prometaphase, when individual chomosomes become visible and the nuclear membrane breaks down, hyaluronan staining was strong and, in several cases, appeared to extend from the periphery of the nucleus into the space between chromosomes (Figure 7C). Hyaluronan filled the cell and completely surrounded the chromosomes at the metaphase plate (Figure 7D) and then filled the space between the separating chromosomes during anaphase (Figure 7E). Strong hyaluronan staining persisted through telophase (Figure 7F). Intense staining of mitotic SMCs after stimulation by PDGF was also seen (not shown), indicating that this is a general effect of mitogenic stimulation and not an undefined serum effect. These observations indicate that hyaluronan may have a functional role on the inside of the cells as well as in the pericellular matrix, specifically during mitosis.
Discussion
Hyaluronan is a structurally simple yet evolutionarily ancient molecule, which has a number of diverse functions (Laurent and Fraser 1992). From previous studies, it is known that synthesis and organization of hyaluronan in the pericellular matrix increases specifically during mitosis in fibroblasts, SMCs, and other cells, suggesting that it aids in cell detachment and rounding (Brecht et al. 1986; Tammi and Tammi 1991; Evanko et al. 1999). The present study indicates that there is also a concomitant increase in the amount of intracellular hyaluronan and the uptake of extracellular hyaluronan in cells that have been stimulated to proliferate. Therefore, the metabolism of hyaluronan, in general, is increased in proliferating cells, especially during mitosis. Furthermore, these data demonstrate that the distribution and numbers of hyaluronan binding sites in cells are also altered in reponse to mitogenic stimulation.
The presence of intracellular hyaluronan may be a general phenomenon because we have also found it in bovine endothelial cells, human mammary epithelial cells, and mammary tumor cells (unpublished observations.) However, the source of cytoplasmic and nuclear hyaluronan is not clear. The present results suggest that in 3T3 cells, fibroblasts, and SMCs, some of the cytoplasmic and nuclear hyaluronan may be derived from an intracellular source. However, this is not yet fully resolved. One recent study reported rapid uptake and translocation of labeled hyaluronan to the nucleus and cytoplasm in association with increased cell motility (Collis et al. 1998). The probe used here requires at least a decasaccharide for binding to hyaluronan (Hascall and Heinegard 1974), and the network appearance of the intracellular staining suggests that the cytoplasmic hyaluronan may be relatively large and not in the form of small oligosaccharides that might be expected from a degradation pathway. Although hyaluronan synthases are believed to be located exclusively in the plasma membrane (Laurent and Fraser 1992), it is possible that there is a cytoplasmic enzyme or, more likely, a synthase on an intracellular membrane that is oriented to secrete hyaluronan into the cytoplasm or nucleus. It is also conceivable that the hyaluronan could be synthesized within a vesicle that is on its way to or from the cell surface but makes its way into the cytoplasm. Support for this idea may come from studies showing that hyaluronan can interact with phospholipid under certain conditions to create perforated membrane-like structures (Pasqualli-Ronchetti et al. 1997), which could allow long, rapidly growing chains of hyaluronan within small vesicles into the cytoplasm. These possibilities warrant further investigation.
There is increasing evidence for the presence of intracellular glycosaminoglycans, and the localization of hyaluronan in the cytoplasm, nucleus, and caveolae in tissues such as brain, liver, artery, cumulus cells, and oocytes has been reported previously (Stein et al. 1975; Fromme et al. 1976; Margolis et al. 1976; Furukawa and Terayama 1977,1979; Londono and Bendayan 1988; Ripellino et al. 1988,1989; Kan 1990; Eggli and Graber 1995). Therefore, hyaluronan may be a component of the nuclear matrix and cytoskeleton in some cells. Although found in a few tissues in vivo using similar histochemical techniques, intracellular hyaluronan may have been overlooked in other tissues because of the overwhelming amounts in the extracellular matrix. An advantage of the cell culture system is that the extracellular hyaluronan can be removed before permeabilization, which facilitates detection of the intracellular material.
The intracellular function(s) of hyaluronan is not yet clear, however. The observations of nucleolar hyaluronan staining and the binding of exogenous hyaluronan to nucleoli and the nuclear periphery are consistent with a study in which hyaluronan was localized to nucleoli and to areas of condensed chroma-tin in the nuclear periphery in oocytes and cumulus cells (Kan 1990). Kan also found hyaluronan to be associated with the rough endoplasmic reticulum. The cytoplasmic staining of hyaluronan and hyaluronan binding sites observed in the present study would be consistent with a distribution in the rough endoplasmic reticulum. A pronounced shift in the binding of labeled hyaluronan from a predominantly nucleolar location in quiescent cells to a distribustion throughout the nucleus and nuclear periphery in growing cells was also noted here. During mitosis, both hyaluronan and hyaluronan binding molecules completely filled the cytoplasm and surrounded the chromosomes during arrangement at the metaphase plate and during separation of the chromosomes in anaphase. Interestingly, this pattern is highly similar to the distribution of lamins during mitosis (Chaly et al. 1984). The processes of chromosomal condensation and rearrangement in mitosis remain poorly understood (Alberts et al. 1989). The present data, along with previous evidence that hyaluronan is associated with chromatin (Furukawa and Terayama 1977, 1979; Kan 1990), raise the possibility that hyaluronan might somehow play a role in chromosome condensation in some cells, perhaps through interactions with histones or lamins. The steric exclusion properties of hyaluronan may also be involved in such a function, as has been proposed for its regulation of cell-cell and cell-substrate adhesion (Toole 1982; Lee et al. 1993; Evanko et al. 1999). The ability of hyaluronan to form networks and organize water within the cell could also have some functional role in mitosis.
On the other hand, it is known that nucleolar components are located at the tips of certain chromosomes and that nucleolar proteins disperse to surround all of the chromosomes during mitosis, and then help reestablish the nucleoli in the daughter cells during telophase (Alberts et al. 1989), which is also consistent with the observations of hyaluronan binding sites. Nucleolar targeting sequences are highly basic (Henderson 1997), and proteins containing these sequences may be responsible for binding hyaluronan in the nucleolus in our experiments. It is tempting to speculate that hyaluronan and hyaluronan binding molecules may have some role in nucleolar function, perhaps being involved in transport of proteins targeted for the nucleus, ribosomal production and trafficking, or mRNA processing. The reported association of hyaluronan with the rough endoplasmic reticulum (Kan 1990) and a recent report of a cytoplasmic hyaluronan binding protein, P32, which has been co-purified with the splicing factor SF2, lend some support to the latter notion (Deb and Datta 1996). Evidence that hyaluronan is found in caveolae is of interest (Eggli and Graber 1995) given that caveolae may be an important cellular location for signaling and regulatory events. In addition, there was an apparent association of hyaluronan with nuclear clefts in fibroblasts. This is of interest because there is emerging recognition that gene regulatory mechanisms are functionally linked to nuclear architecture (Stein 1998); hyaluronan could therefore be involved in some way in this process.
Precedent for regulatory roles of intracellular glycosaminoglycans comes from studies showing that nuclear heparan sulfate is involved in growth control and transcriptional regulation (Fedarko and Conrad 1986; Fedarko et al. 1989; Ishihara and Conrad 1989; Busch et al. 1992). Reports of other hyaluronan binding proteins suggest that this glycosaminoglycan may also have multiple roles inside the cell. For example, the mammalian homologue of the cell cycle control protein CDC37 was recently cloned and shown to bind hyaluronan (Grammatikakis et al. 1995). Second, a putative intracellular isoform of the hyaluronan receptor RHAMM was recently shown to alter erk kinase activity (Zhang et al. 1998). Most recently, Hofmann et al. (1998) reported the cloning of IHABP, an intracellular hyaluronan binding protein which, they suggest, represents the full-length version of RHAMM. IHABP had a remarkably similar cytoplasmic distribution in 3T3 cells to that observed for hyaluronan and hyaluronan binding sites, indicating that it may be one important candidate molecule for further study.
Therefore, hyaluronan now appears to be among a growing list of secreted molecules that also have an intracellular or nuclear function (Henderson 1997; Gooden et al. in press). Clearly, this field is in its infancy, and more studies are necessary to elucidate the role(s) of hyaluronan inside the cell.
Footnotes
Acknowledgements
We would like to thank Russell Ross, Peter Byers, and Daniel Bowen-Pope (all of the Department of Pathology, University of Washington) for providing human smooth muscle cells, human skin fibroblasts, and 3T3 cells, respectively, and Charles Underhill (Department of Anatomy and Cell Biology, Georgetown University) for providing the hyaluronan binding probe. We also thank Terry Kavanaugh (Department of Environmental Health, University of Washington) and Stephanie Lara (Department of Pathology, University of Washington) for assistance with the confocal microscopy.