
Abstract
Caspase-14 has been implicated in the formation of stratum corneum because of its specific expression and activation in terminally differentiating keratinocytes. However, its precise physiological role and its protein substrate are elusive. We studied the ultrastructural localization of caspase-14 in human epidermis to compare its distribution pattern with that of well-characterized differentiation markers. Immunogold cytochemistry confirmed that caspase-14 is nearly absent in basal and spinous layers. In the granular, layer nuclei and keratohyalin granules were labeled with increasing intensity towards the transitional layer. Particularly strong caspase-14 labeling was associated with areas known to be occupied by involucrin and loricrin, whereas F-granules, occupied by profilaggrin/filaggrin, were much less labeled. A high density of gold particles was also present at the forming cornified cell envelope, including desmosomes. In corneocytes, intense labeling was both cytoplasmic and associated with nuclear remnants and corneodesmosomes. These observations will allow focusing efforts of biochemical substrate screening on a subset of proteins localizing to distinct compartments of terminally differentiated keratinocytes.
M
Profilaggrin, a histidin-rich protein that aggregates keratins, is present in F-granules of composite keratohyalin (Fukuyama and Epstein 1986; Holbrook 1989; Steven et al. 1990; Manabe et al. 1991). F-granules have been described for both rodents and human keratohyalin (Yoneda et al. 1992; Ishida-Yamamoto et al. 1993). During terminal keratinocyte differentiation they are processed into mature filaggrin units and later degraded to form the so-called natural moisturizing factor of the stratum corneum, whereby only a small amount remains chemically associated with keratins 1 and 10 and corneous cell proteins (Resing and Dale 1991; Steinert and Marekov 1995; Ishida-Yamamoto et al. 1999, 2000).
Loricrin, the main sulfur-rich protein of keratohyalin, is instead localized in round or L-granules in rodents (especially mouse) keratohyalin (Steven et al. 1990; Hardman et al. 1998), or in amorphous material often associated with F-granules of the composite human keratohyalin (Yoneda et al. 1992; Ishida-Yamamoto et al. 1993, 1996, 2000). Another major protein of terminal differentiating keratinocytes, involucrin, is both cytoplasmic and associated with the amorphous material of composite keratohyalin granules (Warhol et al. 1985; Ishida-Yamamoto and Iizuka 1994). Proteins such as involucrin, small proline-rich proteins, sciellin, elafin, and eventually loricrin are deposited against the cytoplasmic side of the plasma membrane and desmosomes of terminally differentiating keratinocytes, while ceramides and waxes are deposited mainly on the external surface of the membrane (Kalinin et al. 2002; Menon and Norlen 2002).
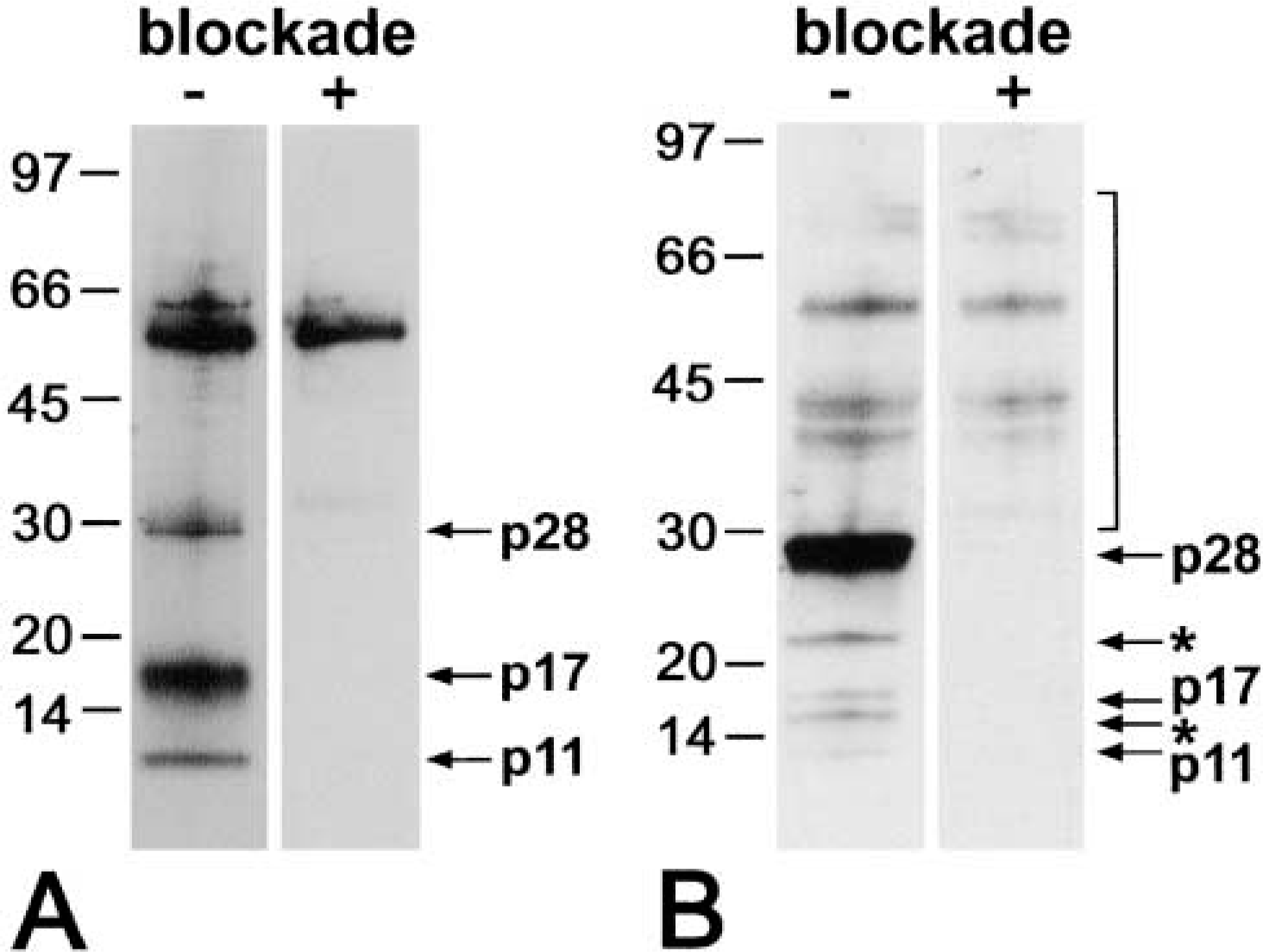
Specifity control of the anti-human caspase-14 antibody. Protein extracts from stratum corneum of psoriatic lesions (
Most transformation processes during keratinocyte cornification are mediated by specific enzymes. For example, the formation of the insoluble cornified cell envelope is achieved by transglutaminases and by sulfhydryl oxidase, which crosslink proteins via isopeptide and disulfide bonds, respectively (Polakowska and Goldsmith 1991; Hashimoto et al. 2000).
Because the transition of keratinocytes of the granular layer into enucleate corneocytes represents a form of cell death, we recently screened the caspase family of pro-apoptotic proteases for members specifically expressed in these cells. One caspase, caspase-14, was found to be almost exclusively present in differentiating keratinocytes (Eckhart et al. 2000a, b; Lippens et al. 2003). Like other caspases, caspase-14 is expressed as a proenzyme that is activated by cleavage into a large and a small subunit.
The protein substrate of caspase-14 is unknown at present. Activation of caspase-14 was found to be associated with the formation of the stratum corneum in epidermal equivalent models in vitro, and processed caspase-14 was detected in the stratum corneum of human skin (Eckhart et al. 2000a). Immunohistochemical (IHC) analysis revealed that caspase-14 is also present in the upper layers of the epidermis, in sebaceous glands and hair follicles. Immunoreactivity was found primarily in the cytosol but occasionally also in the nuclei of keratinocytes (Eckhart et al. 2000a; Lippens et al. 2003). Knowledge of the detailed cytological localization of casp-14 is desirable because this may be a valuable guide in the search for the protein substrate. Therefore, we determined the subcellular localization of caspase-14 in human epidermis by immunoelectron microscopy.
Materials and Methods
Antibodies
A rabbit antiserum was raised against the human caspase-14 proenzyme produced by recombinant DNA technology in E. coli. The specificity of the antibody was confirmed by Western blotting analysis of protein lysates from stratum corneum squames and normal epidermis, as described below. Experiments were also performed with a rabbit antiserum raised against murine caspase-14 [kindly provided by Wim Declercq, Flanders Interuniversity Institute for Biotechnology (VIB) and Ghent University, Ghent, Belgium] (Lippens et al. 2000). The latter antibody recognized the same bands as the anti-human caspase-14 antibody on Western blots and crossreacted with human caspase-14 in IHC (Eckhart et al. 2000a; Lippens et al. 2000, 2003). As control, the anti-caspase-14 antibody was preabsorbed with the antigen by incubating a 1:10 dilution of the antiserum with 3 μg recombinant human caspase-14 for 1 hr at room temperature.
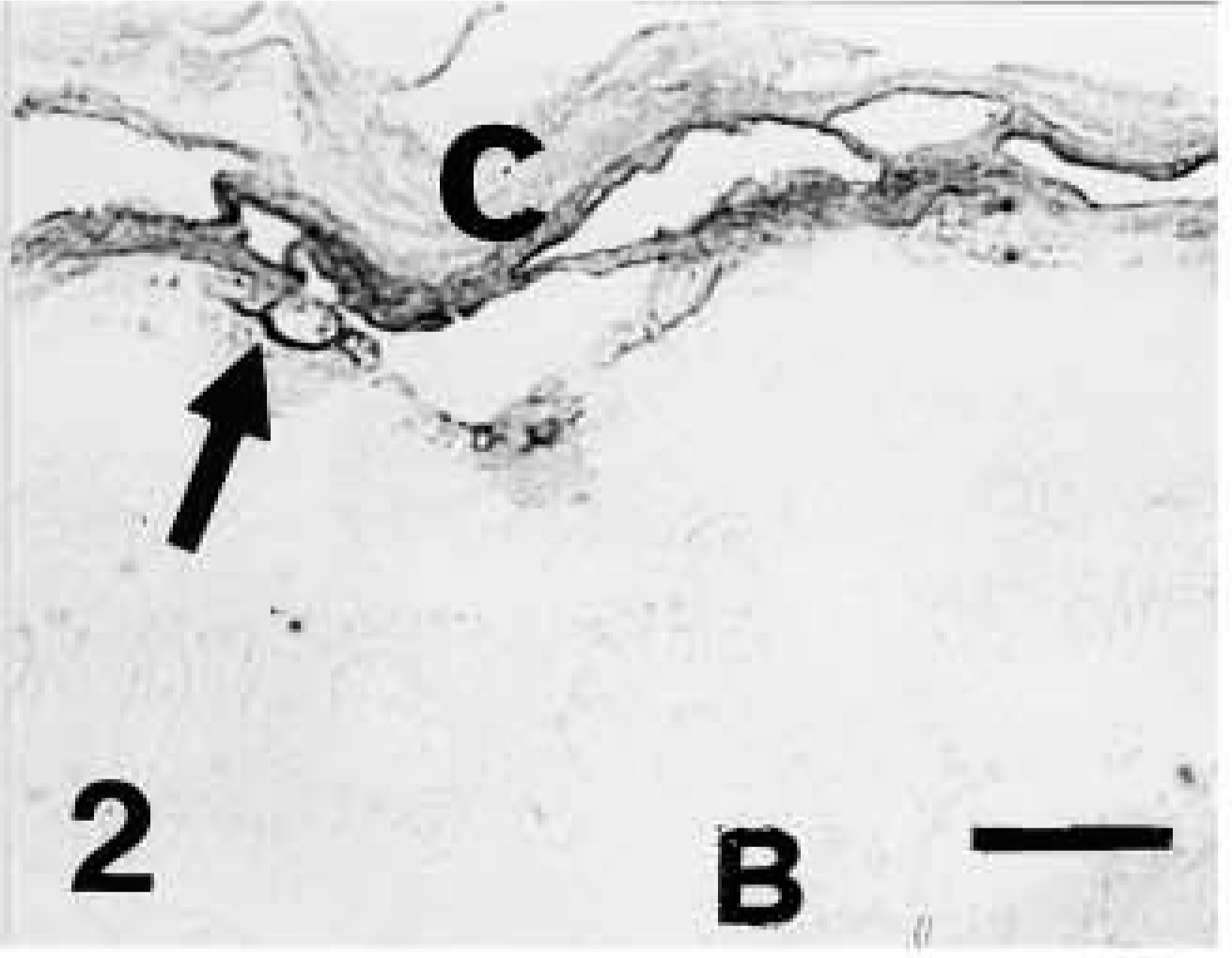
Light microscopic investigation of caspase-14 in human skin. Anti-human caspase-14 antibody. Caspase-14 immunolabeling present only in granular (arrow) and corneous (c) layer of human epidermis. B, basal layer. Bar = 10 μm.
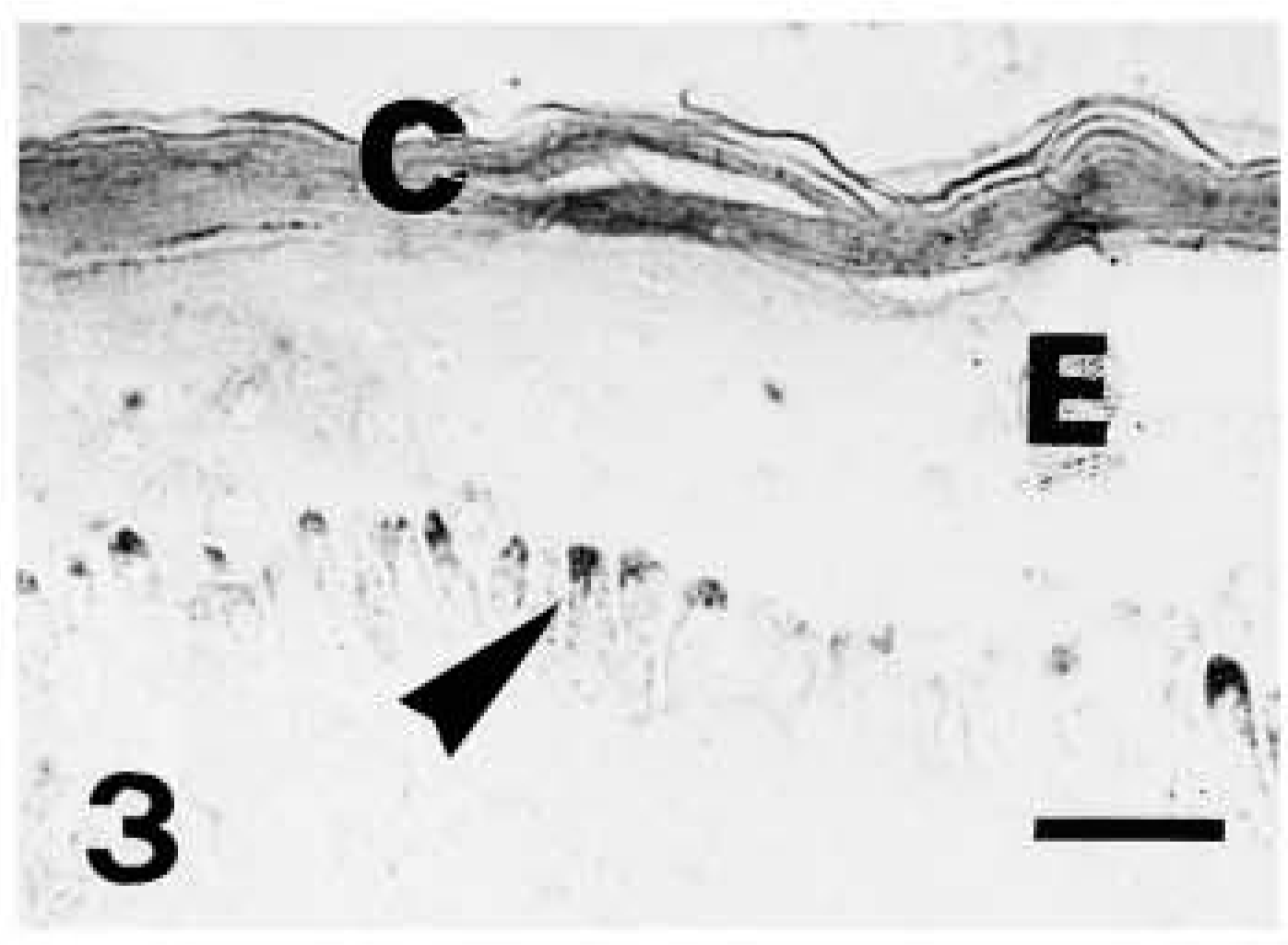
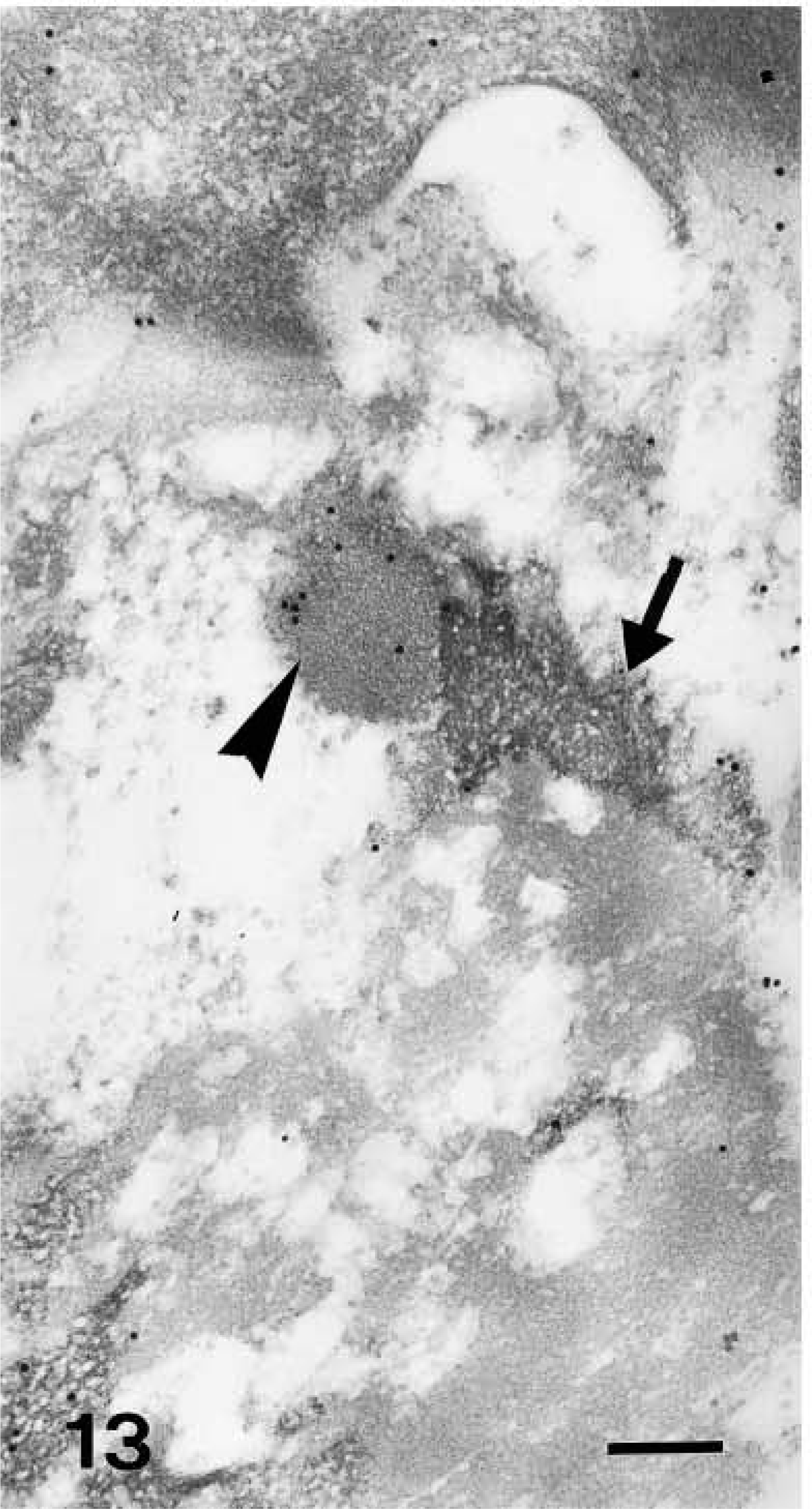
Light microscopic investigation of caspase-14 in human skin. Anti-human caspase-14 antibody. Strong caspase-14 labeling restricted to the corneous layer (C) of another piece of epidermis (E). The arrowheads point to pigmented but unstained melanocytes in the basal layer. Bar = 10 μm.
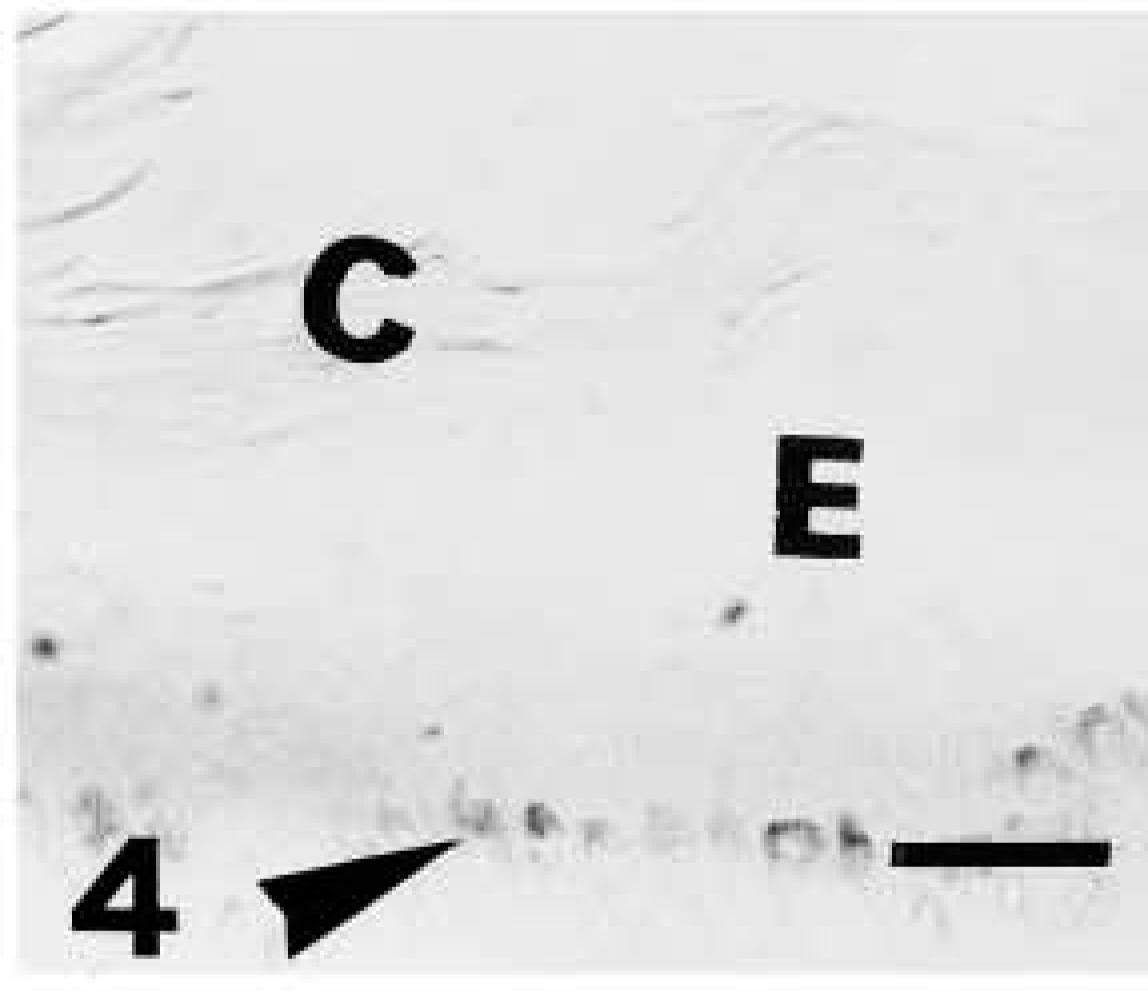
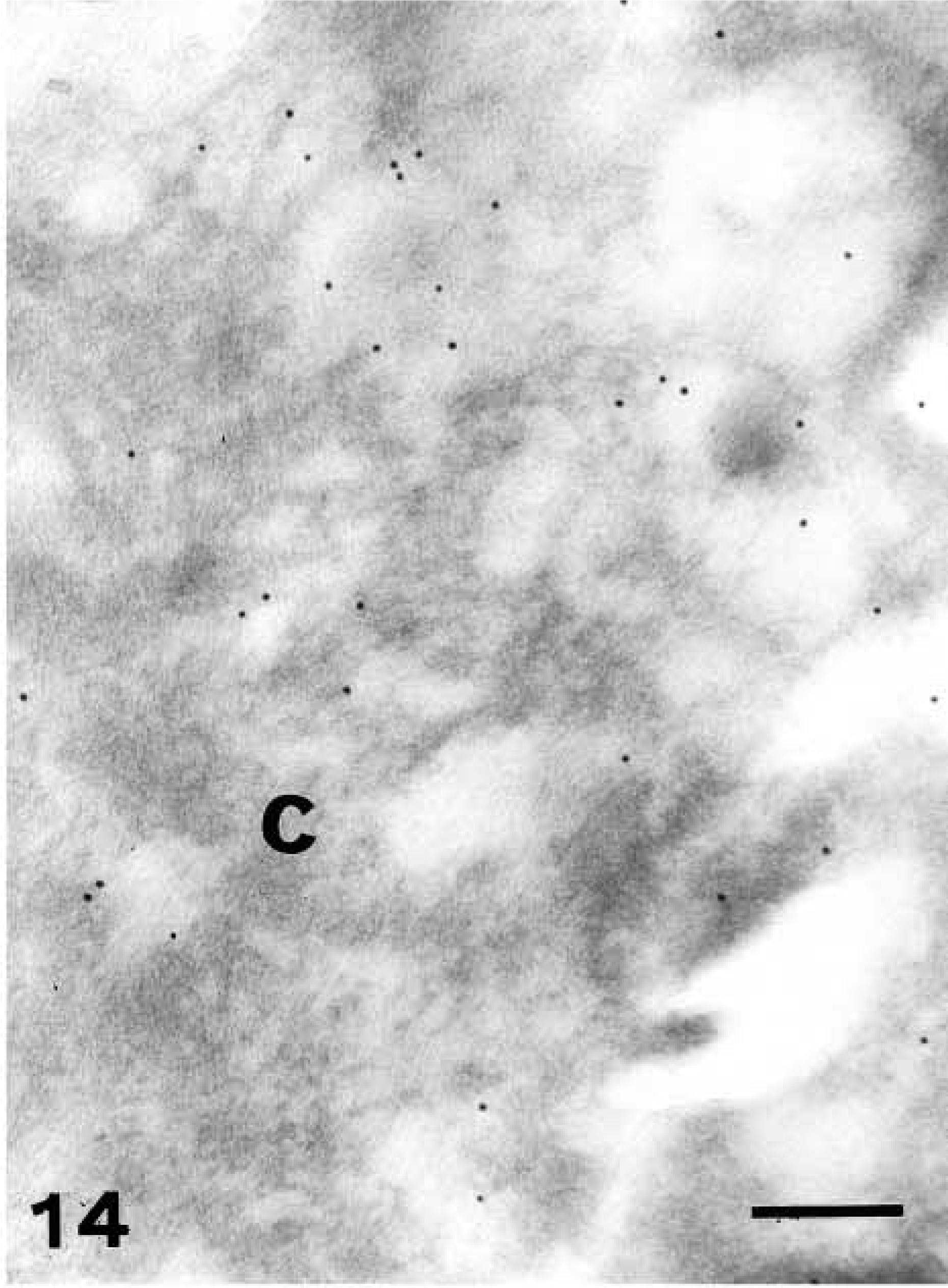
Light microscopic investigation of caspase-14 in human skin. Anti-human caspase-14 antibody. Negative control (omitting the anti-human caspase 14 antibody) of previous section showing immunonegative epidermis (E) and corneous layer (C), while pigmentation in basal melanocytes is still present (arrowhead). Bar = 10 μm.
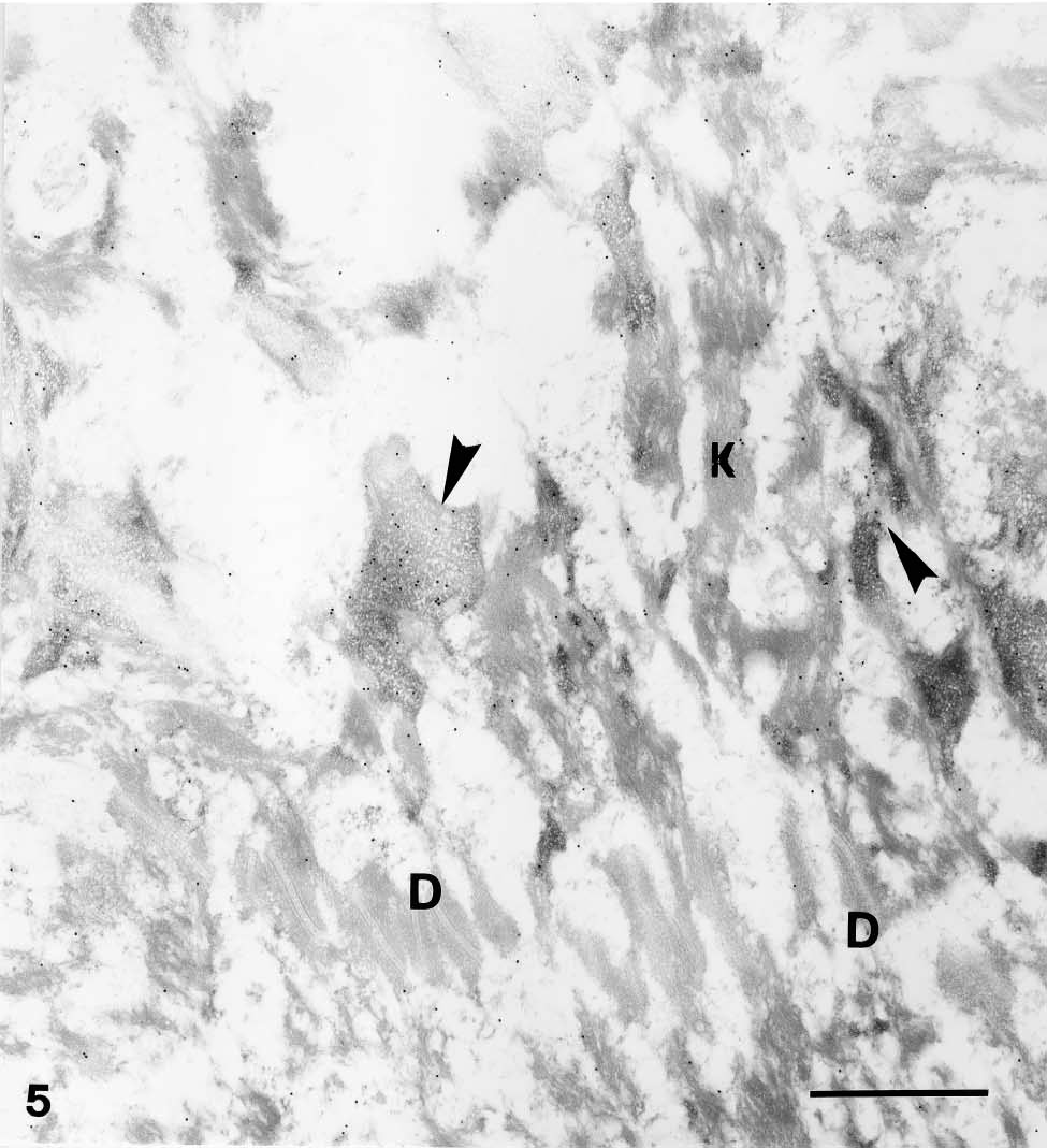
Immunogold electron microscopic overview of granular and corneous layers of human epidermis. Anti-mouse caspase-14 antibody. Distribution in granular layer as detected by 20-nm immunogold electron microscopy at low magnification. Most immunogold labeling is associated with keratohyalin granules (arrowheads), while keratin bundles (K) and desmosomes (D) are negative. Bar = 1 μm.
Immunogold electron microscopic overview of granular and corneous layers of human epidermis. Anti-mouse caspase-14 antibody. Low-magnification view of upper granular and corneous layer as detected by 20-nm gold particles. The labeling is associated with composite keratohyalin granules (arrowheads) and the corneous layer (C) but not with keratin bundles (
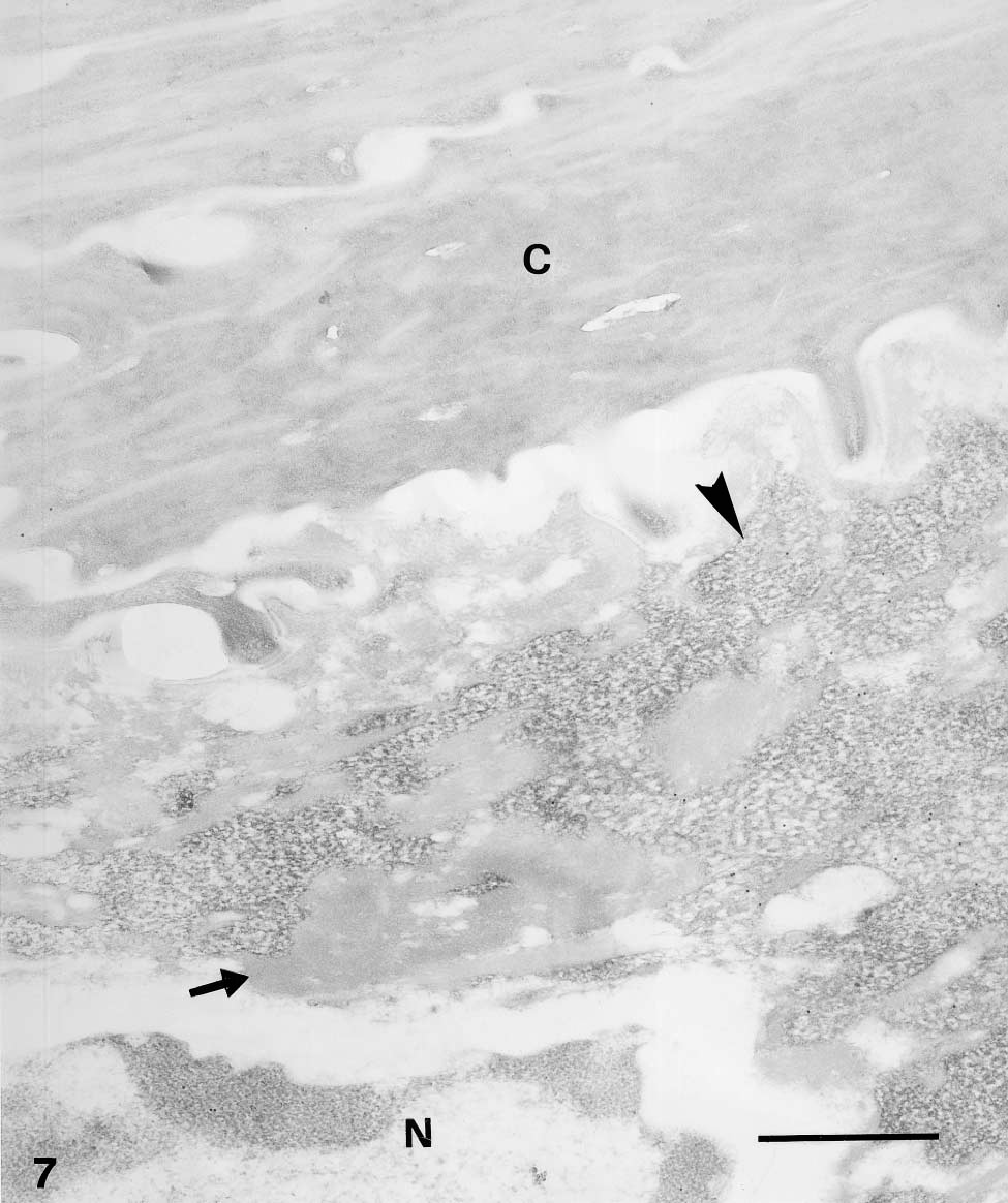
Immunogold electron microscopic overview of granular and corneous layers of human epidermis. Anti-mouse caspase-14 antibody. Low-magnification (the same as previous figure) of upper granular and corneous layer after preabsorption of the mouse caspase-14 antibody with the antigen (human caspase-14). Very few gold particles are seen over F-granules (arrowhead), associated electron-pale material (arrow), and the corneous layer (C).
Skin Samples
For Western blot analysis, skin samples were obtained from mammary reduction plastic surgery and stratum corneum squames were collected from psoriasis patients after obtaining their informed consent. For immunolocalization of caspase-14, samples of normal human skin (hospital biopsies), collected from the arm and finger of three individuals, were used. Samples (2 × 4 mm) were collected under ethical and legal approval.
Electrophoresis and Immunoblotting
Epidermal proteins and proteins from stratum corneum squames were prepared as described previously and subjected to Western blotting analysis according to a published protocol (Eckhart et al. 2000a). The antibody raised against human caspase-14 was used at a dilution of 1:1000. As control, the anti-caspase-14 antibody was preincubated with 3 μg/μl recombinant human caspase-14 for 2 hr at RT.
Tissue Preparation
The tissues were immediately fixed at 0-4C for 4-6 hr with in a modified Carnoy's fluid (9 parts 90% ethanol and 1 part acetic acid). In addition, some pieces from the last sample were fixed in 4% paraformaldehyde in 0.1 M phosphate buffer at pH 7.4. Tissues were dehydrated at 0-4C and embedded in Bioacryl resin under UV polymerization at 0-4C (Scala et al. 1992). Using an ultramicrotome, tissues were sectioned at 1-2-μm thickness for light microscopic observations. Parallel sections were collected over chromoalum/albumin-coated slides for the following immunocytochemical reaction. From tissues fixed in 4% paraformaldehyde, thin sections at 40-80-nm thickness were collected on nickel grids for ultrastructural immunogold cytochemistry.
IHC and Electron Microscopy
The dilution was 1:500-1000 for the anti-human caspase-14 or 1:1500 for the mouse caspase-14 antibodies in 0.05 M Tris-HCl at pH 7.6 with 2% BSA for light immunocytochemistry, or with 1% coldwater fish gelatin for ultrastructural immunocytochemistry. In controls the primary antibody was preabsorbed with the antigen by mixing the antibody solution with a solution of recombinant human caspase-14 (2 μg/μl) at a ratio of 1:1 for 1 hr at RT. In other negative controls the primary antibody was omitted from the incubating solution. Light immunocytochemistry was performed using a secondary HRP-conjugated anti-rabbit IgG antibody (Biorad, Richmond, CA, or Sigma, St Louis, MO) at a dilution of 1:50, which was revealed by a diaminobenzidine (DAB) reaction. For transmission electron microscopy (TEM) immunocytochemistry the secondary (anti-rabbit IgG) antibody was conjugated to 10-nm gold (Sigma). To check for co-localization of involucrin and caspase-14 in the epidermis, a double-labeling immunostaining was performed with a monoclonal anti-human involucrin antibody (clone SY5; Sigma) that is specific for human epidermis (Alibardi and Maderson 2003) and with the anti-murine caspase-14 antibody. The secondary antibodies were a 10-nm gold-conjugated goat anti-mouse IgG (for involucrin) and a 20-nm gold-conjugated goat anti-rabbit IgG (for caspase-14). Thin sections were lightly stained in 4% uranyl acetate. The ultrastructural immunocytochemical observation was done using a Philips CM-100 electron microscope operating at 80 kV.
Results
In the present study an anti-caspase-14 antibody described previously (Eckhart et al. 2000a; Lippens et al. 2000, 2003) and a novel antibody against human caspase-14 were used. The anti-human caspase-14 antibody recognized procaspase-14 as well as the catalytically active caspase-14 subunits, as revealed by Western blotting of stratum corneum and total epidermal proteins (Figure 1). Specificity of binding was confirmed by antigen preabsorption, which blocked reactivity towards caspase-14 bands, whereas unspecific binding to high molecular weight proteins was unaffected (Figure 1).
Light microscopic observations with the anti-human caspase-14 or the anti-mouse caspase-14 (Lippens et al. 2000) antibody showed a similar pattern, although 10-nm gold was used for human caspase-14 and 20-nm gold was used for mouse caspase-14. Unless specifically reported, the following description refers to the labeling pattern observed with both the antibodies.
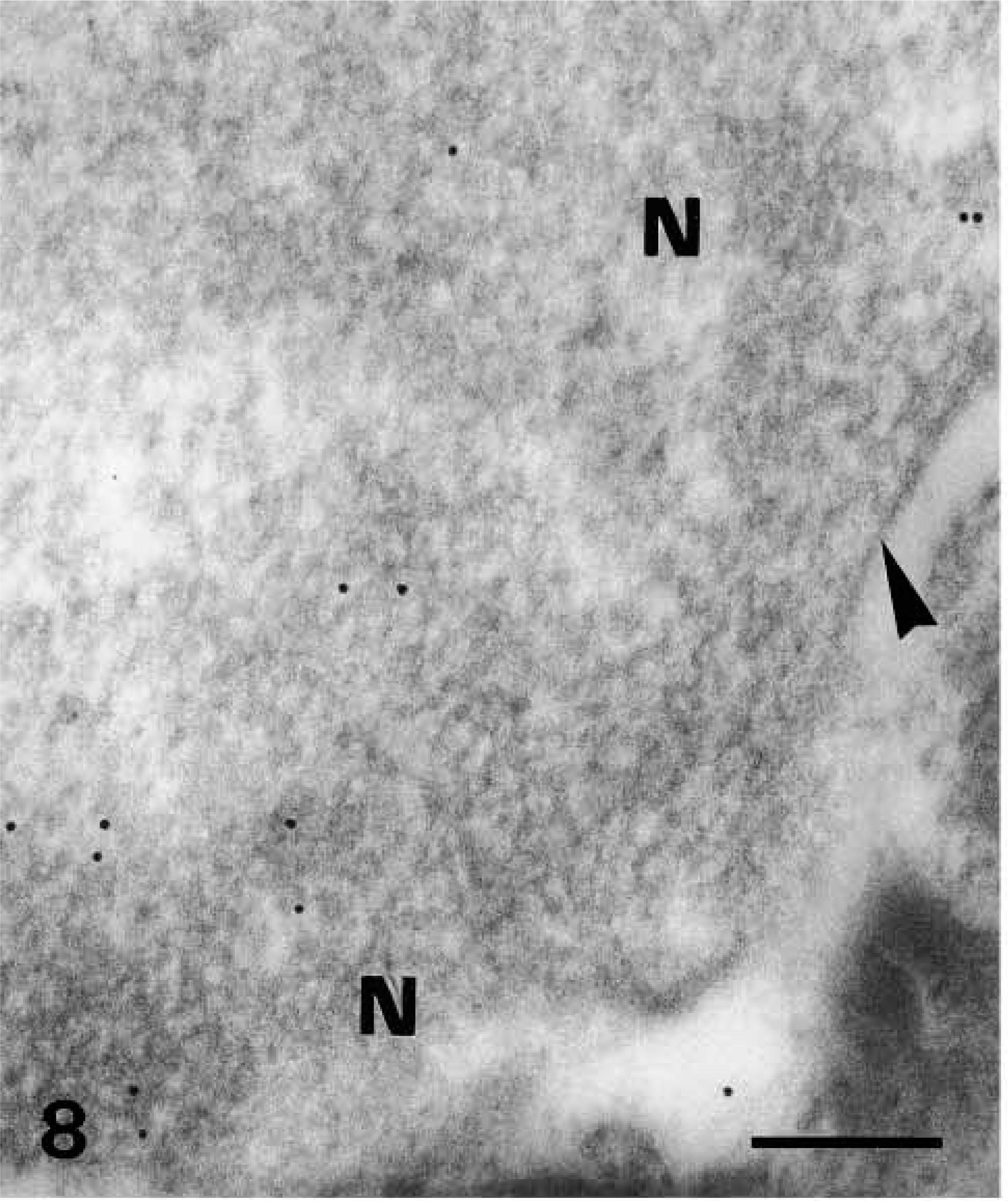
Ultrastructural distribution of caspase-14 in upper granular layer as detected by 10-nm gold particles. Anti-human caspase-14 antibody. Detail of labeling of nuclear heterochromatin (
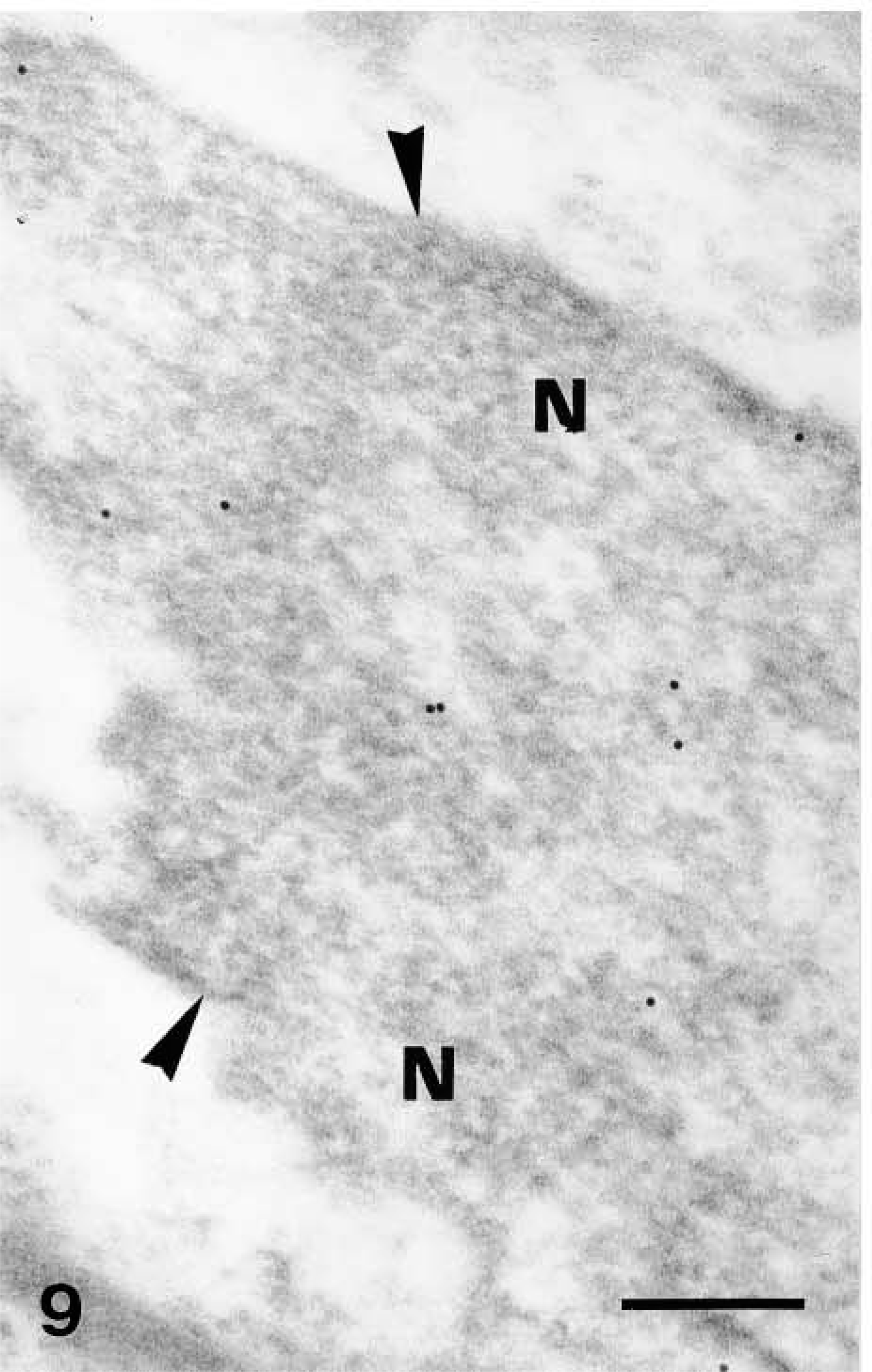
Ultrastructural distribution of caspase-14 in upper granular layer as detected by 10-nm gold particles. Anti-human caspase-14 antibody. Other diffusely labeled nuclei (
Ultrastructural distribution of caspase-14 in upper granular layer as detected by 10-nm gold particles. Anti-human caspase-14 antibody. Reticular appearance of F-granule of keratohyalin free in the cytoplasm of an upper granular cell. The arrow points to a filament of the network that makes the granule. Bar = 200 nm.
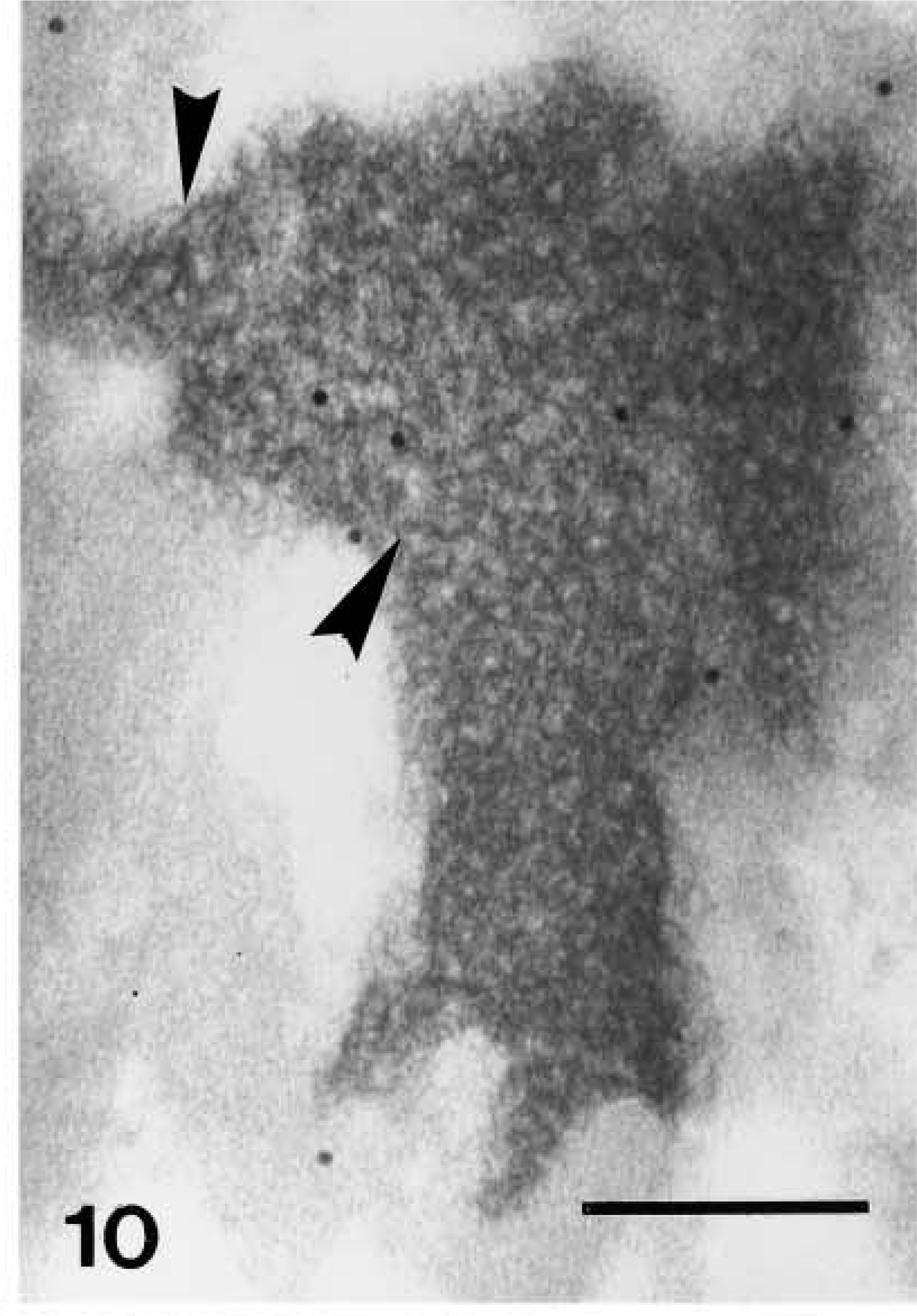
Ultrastructural distribution of caspase-14 in upper granular layer as detected by 10-nm gold particles. Anti-human caspase-14 antibody. Structure of composite keratohyalin in upper granular cells with F-granules (arrows) and intensely labeled amorphous component (arrowhead). D, desmosome. Bar = 200 nm.
Caspase-14 immunoreactivity was restricted to the granular and mainly to the transitional and corneous layers of human epidermis (Figures 2 and 3). Basal and spinous layers were only weakly stained or were immunonegative. Labeling was seen over keratohyalin granules of the upper granular layer, as detailed below in the ultrastructural description. Negative controls performed by preabsorption of the primary antibody with recombinant antigen or by omitting the primary antibody yielded greatly reduced or no labeling (Figure 4).
The ultrastructural study showed absence of caspase-14 or occasional labeling over organelles, keratin bundles, or nuclei in cells of the basal and spinous layers (not shown). In the lowermost granular layer, characterized by small and sparse keratohyalin granules with a reticular structure, a few organelles labeled with gold particles were seen near the cell membrane. Neither keratin bundles nor desmosomes were labeled (Figure 5). Most of the labeling for caspase-14 was seen in the granular and corneous layers. In the granular layer the labeling was localized in the composite keratohyalin granules associated with the typical reticular F-granules of keratohyalin (Figure 5). A few gold particles were present over clumps of heterochromatin in nuclei of lower granular layer cells, but not over keratin bundles.
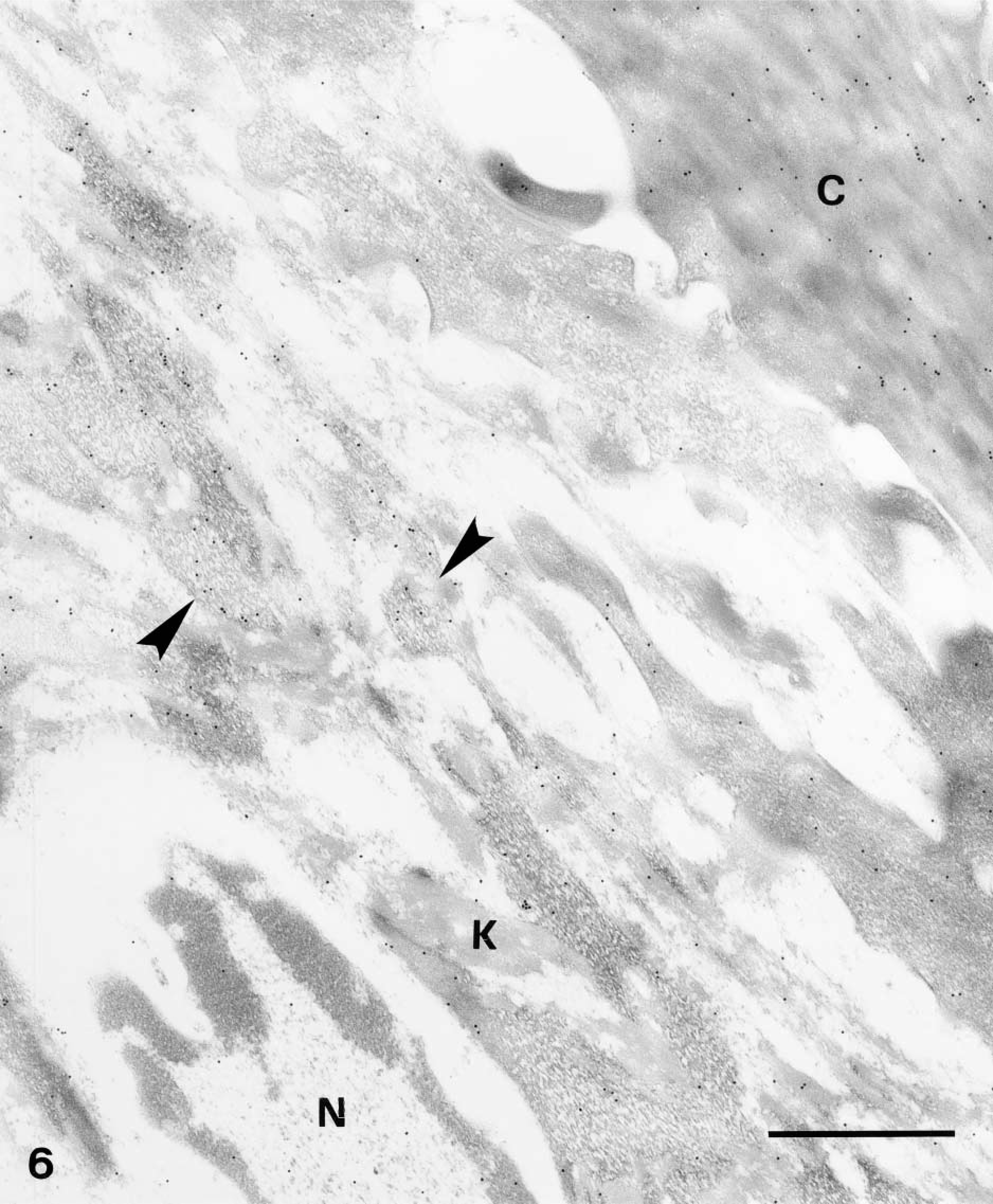
In cells of the upper granular and transitional layers, diffuse but consistent labeling was seen among keratin bundles and over keratohyalin composite granules but not along the plasma membrane (Figure 6). In controls, either negative (omitting the primary anti-human or anti-mouse antibodies), no, or little labeling was seen, including the labeling over the corneous layer that was almost completely abolished (Figure 7).
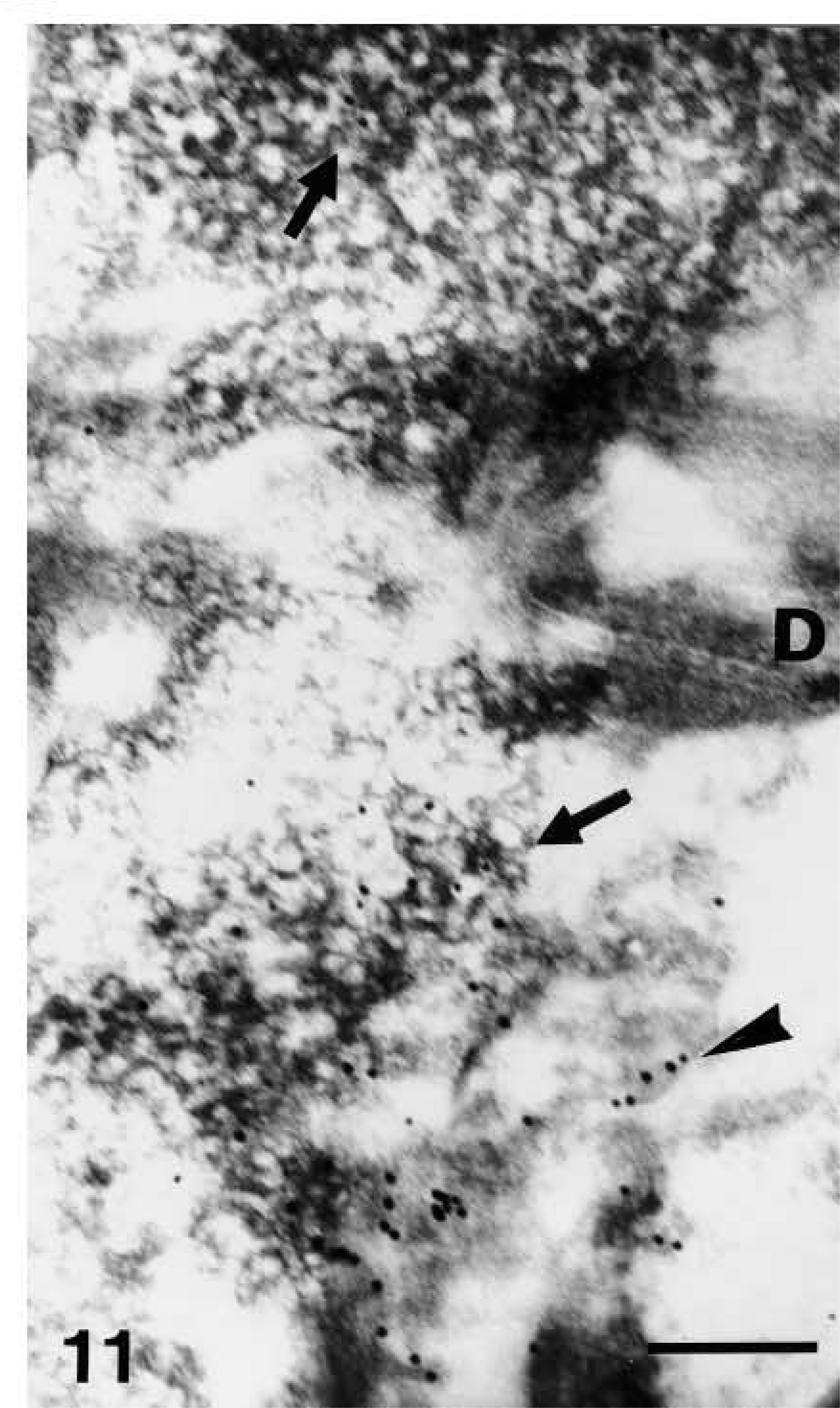
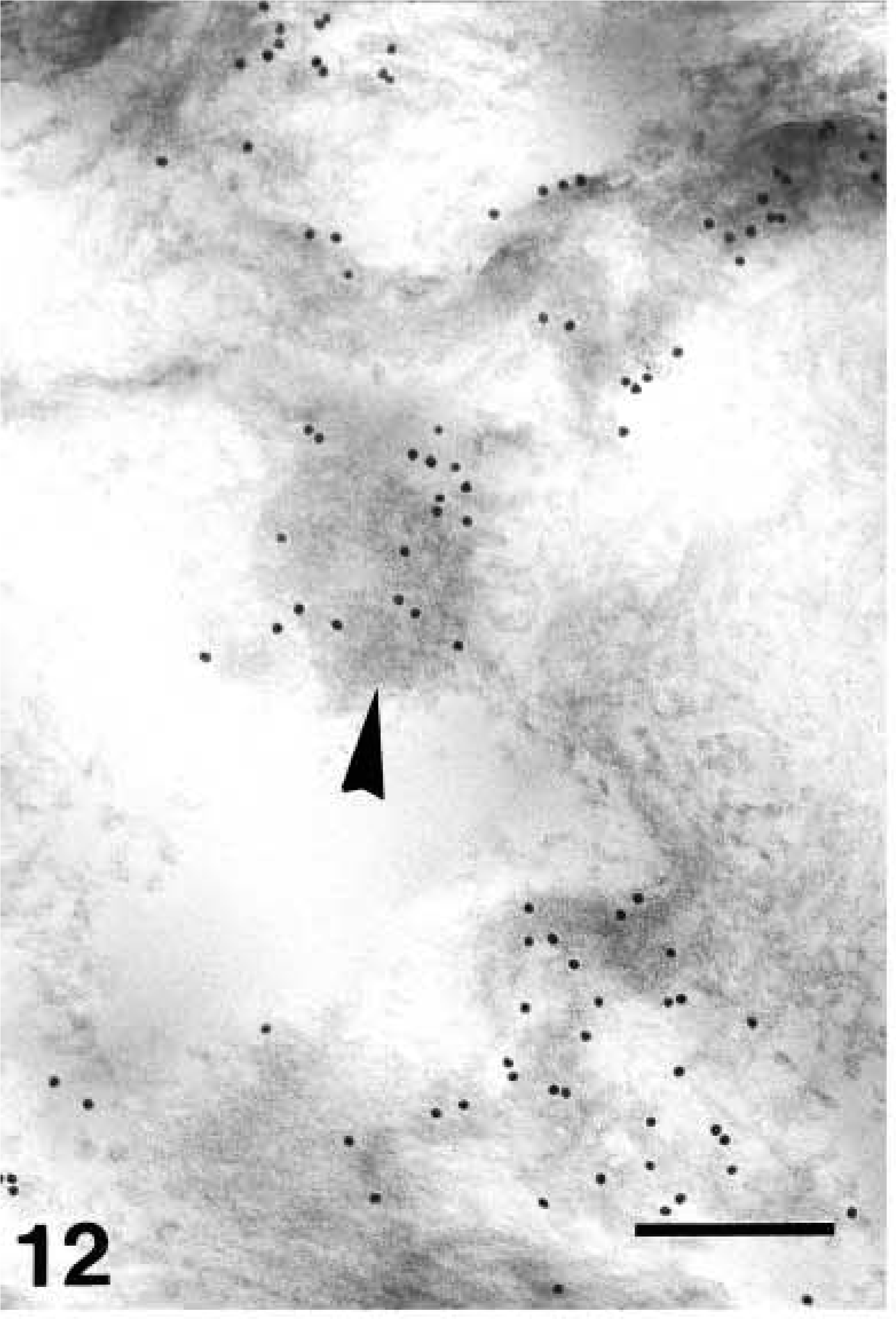
Nuclei of upper granular cells showed a distinct although still diffuse immunolabeling (Figures 8 and 9). Detailed study of composite large masses of keratohyalin showed an intense labeling (either with the anti-human or the anti-mouse antibody) mainly localized over the amorphous electron-pale material between F-granules (Figures 10, 11, 12, and 13). The latter appeared as a network of ~10-nm-thick filaments. The labeling also appeared among keratin bundles mixed with keratohyalin granules (Figures 12 and 13). No labeling was present in the extracellular spaces.
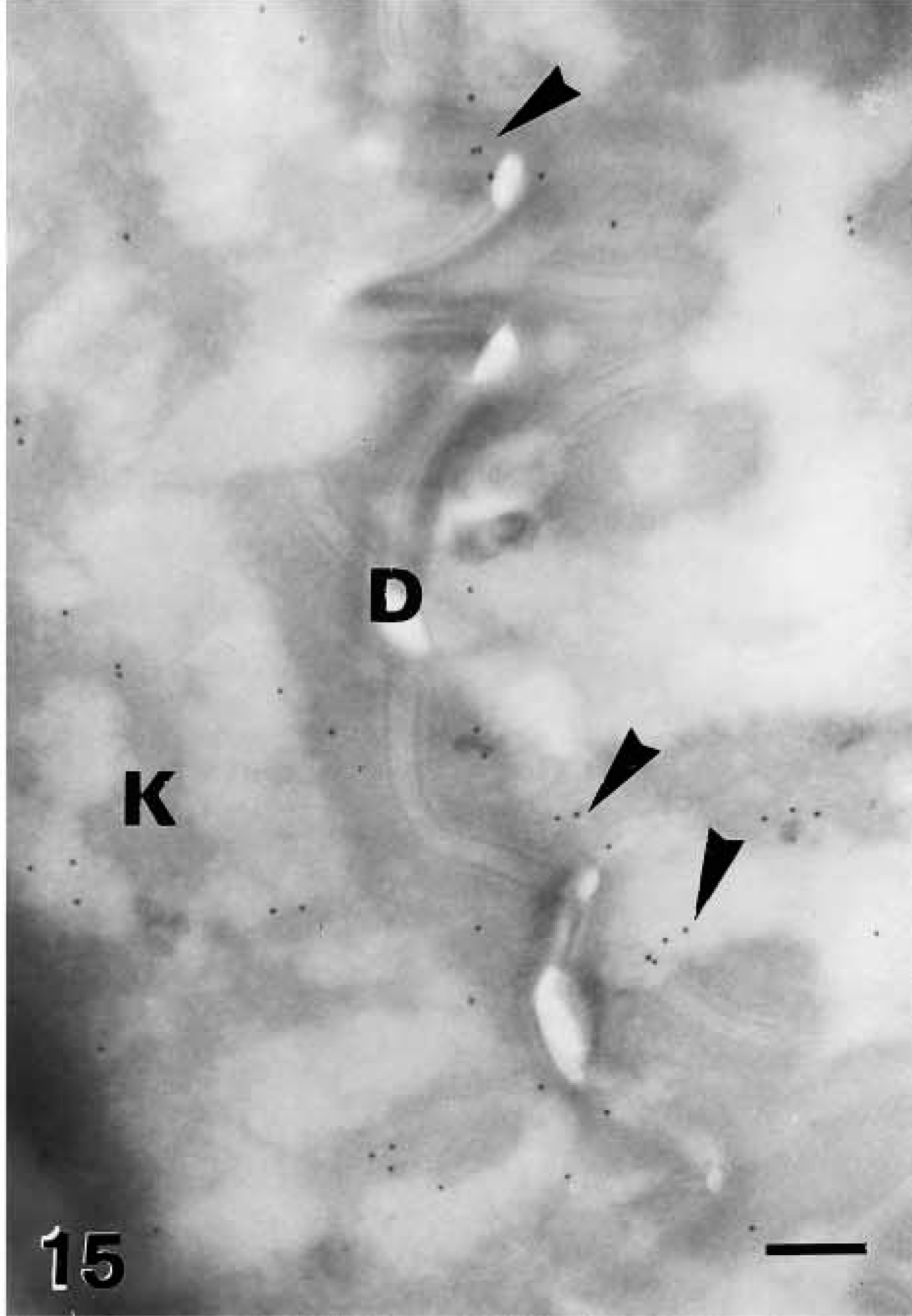
In partially cornified cells, most of the organelles disappeared, including keratohyalin, while a diffuse immunolabeling became visible over condensed amorphous material derived from the degradation of keratin mixed with keratohyalin and nuclear remnants (Figure 14). Although most gold particles were cytoplasmic, some particles also decorated areas along the plasma membrane and desmosomes of transitional cells (Figure 15).
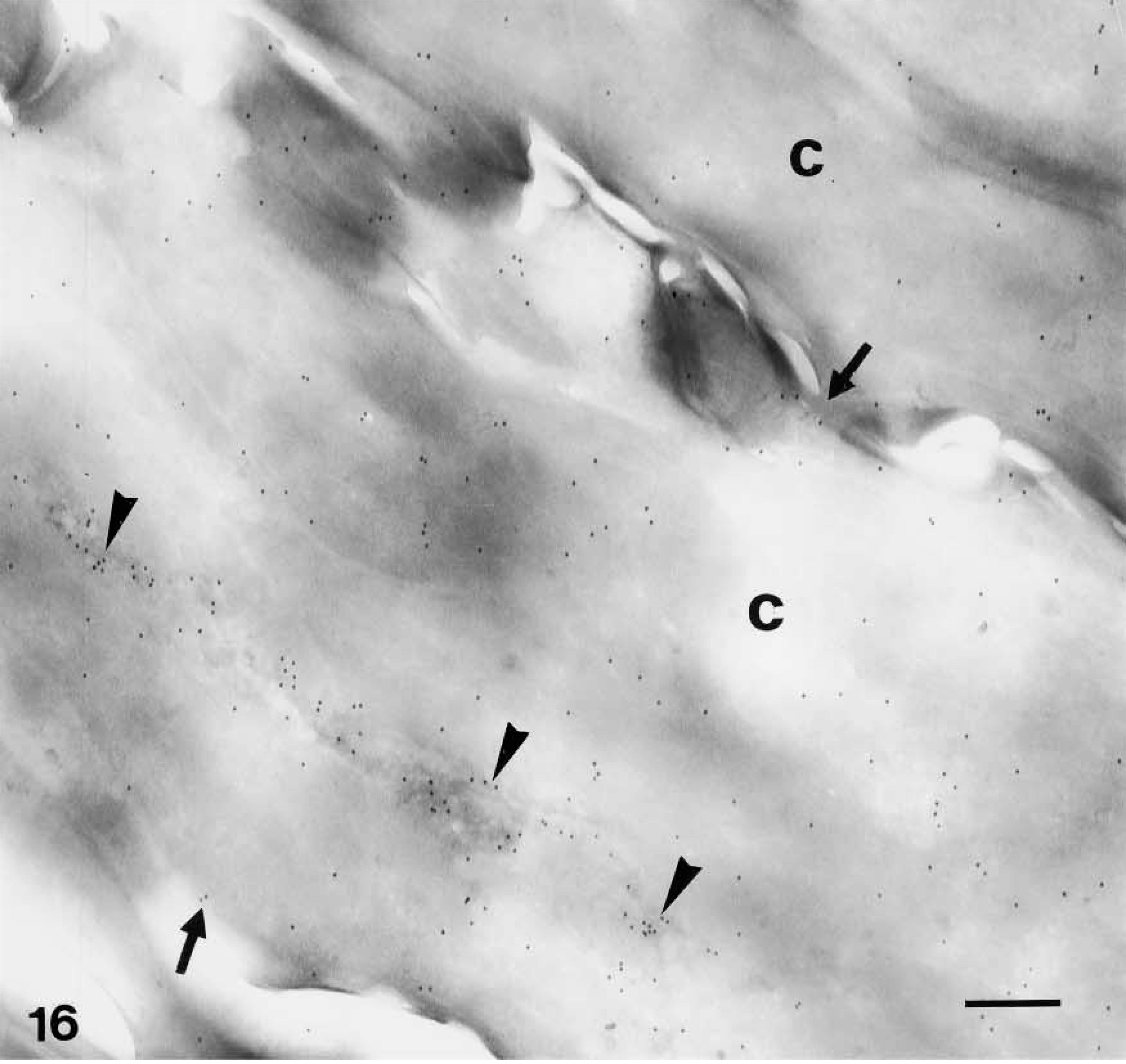
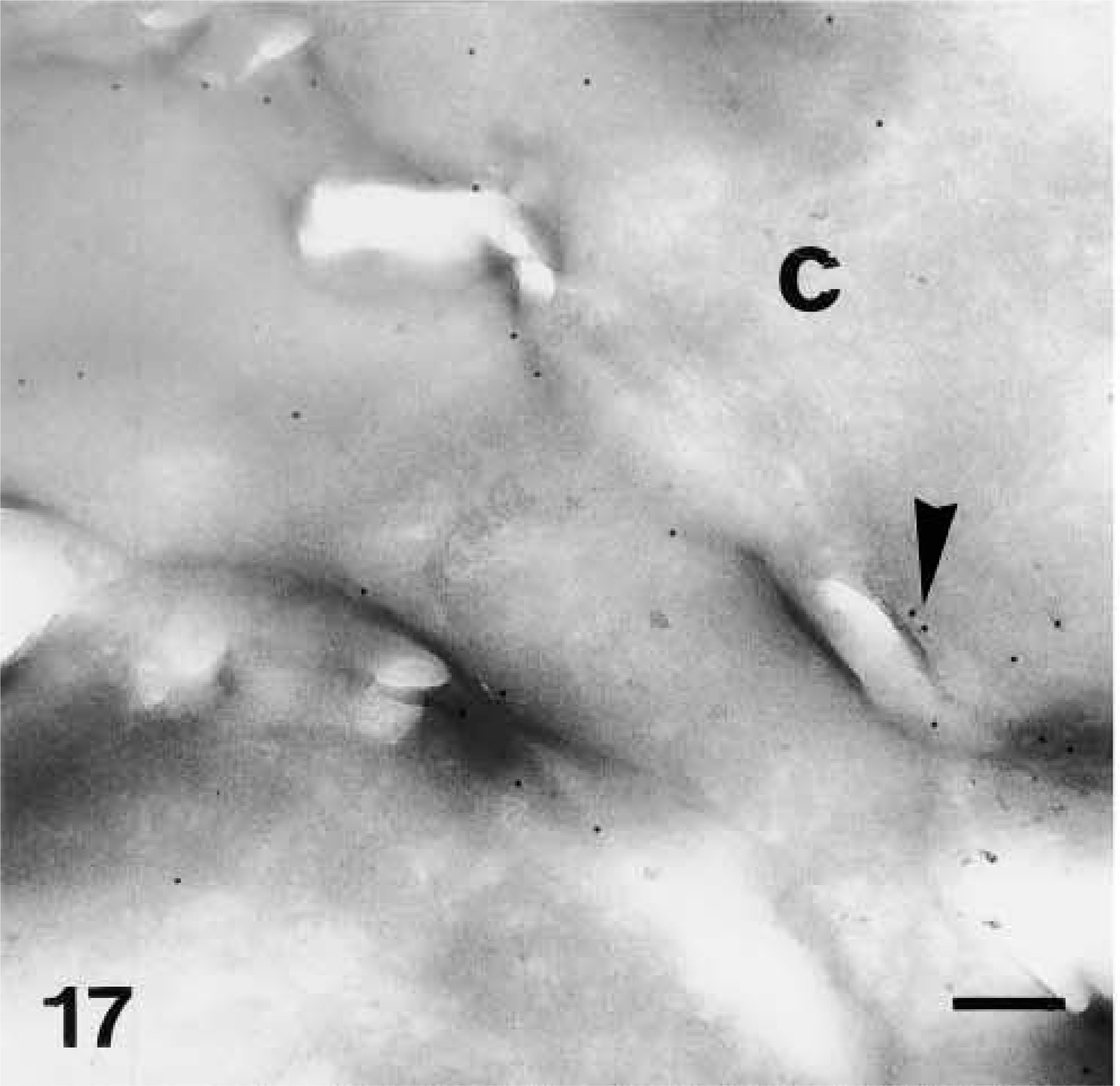
Very strong immunolabeling was found in partially or completely mature corneocytes of the lowermost corneous layer, both in the corneous cytoplasm and, at highest intensity, in areas occupied by seemingly nuclear remnants (Figures 5 and 16). Some gold particles were consistently observed near the cornified envelope or were more clearly associated with corneodesmosomes present along the irregular surface and interlocking spines of mature corneocytes (Figures 17 and 18). The labeling was intracellular and was nearly absent extracellularly among corneocytes.
Because the distribution pattern of caspase-14 resembled that of involucrin, we performed double labeling to substantiate the data on co-localization. In these experiments, caspase-14 was detected with a polyclonal rabbit antibody that had been raised against murine caspase-14 but that crossreacted strongly with human caspase-14 (Lippens et al. 2000). This antibody yielded a very similar pattern of immunolabeling as the one specific for human caspase-14 and could be blocked almost completely by preabsorption with recombinant human caspase-14 (Figure 7; see Lippens et al. 2000). Involucrin, labeled with a monoclonal mouse antibody, was diffusely distributed in the cytoplasm of the upper spinous layer, while caspase-14 labeling was absent or very diffuse (Figure 19). In granular cells the double labeling was diffuse or associated with composite granules, but not over keratin bundles or desmosomes (Figure 20). In keratohyalin granules of the granular and transitional layers, both involucrin and caspase-14 immunolabeling were especially associated with the amorphous material around the network of F-granules (Figure 21). In stratum corneum, involucrin labeling decreased towards the outer layers while caspase-14 labeling remained strong (Figure 22).
Ultrastructural distribution of caspase-14 in upper granular layer as detected by 10-nm gold particles. Anti-human caspase-14 antibody. Intense immunolabeling over mixed keratin and keratohyalin (arrowhead) of a transitional layer cell. Bar = 200 nm.
Caspase-14 immunolabeling over mixed pale amorphous material (arrowhead) and F-granules (arrow) of keratohyalin of a transitional cell. Anti-mouse caspase-14 antibody. Bar = 200 nm.
Caspase-14 immunolabeling in transitional cells. Anti-human caspase-14 antibody. Ultrastructural distribution of labeling among the condensing corneous material (C) in transitional layer. Bar = 200 nm.
Caspase-14 immunolabeling in transitional cells. Anti-human caspase-14 antibody. Condensing keratin material (
Discussion
The present investigation of the ultrastructural localization of caspase-14 in human epidermis extends previous studies on such distribution (Eckhart et al. 2000a, b; Lippens et al. 2000, 2003). With a new antibody specific for human caspase-14, a strong increase of caspase-14 abundance during keratinocyte differentiation was observed, in agreement with former studies that had been performed with an anti-murine caspase-14 antibody that crossreacted with the orthologous human protein (Eckhart et al. 2000a, b; Lippens et al. 2000; Rendl et al. 2001; and present observations). Also at the ultrastructural level, both antibodies yielded the same pattern of labeling, as presented for the first time in our study.
By high-resolution electron microscopy caspase-14 was found to be mainly cytosolic, but distinct labeling was also detected in the nucleus and at desmosomes (see below). The cytosolic labeling increased from the lower to the upper layer of the stratum granulosum, where it was preferentially detected on the electronpale areas of composite keratohyalin. Double-labeling experiments revealed that caspase-14 and involucrin are present at the same sites in these cells. Because previous reports have shown that loricrin and other minor proteins destined to form the cornified envelope co-localize with involucrin (Warhol et al. 1985; Steven et al. 1990; Manabe et al. 1991; Ishida-Yamamoto et al. 1996, 2000; Presland et al. 1997; Ishida-Yamamoto and Iizuka 1998), our findings suggest that all these components of keratohyalin are potential substrates for caspase-14. By contrast, the weak labeling intensity of F-granules suggests that filaggrin and its binding partners are less likely candidates. Interestingly, co-localization of caspase-14 and involucrin was not detectable in the stratum corneum. Whereas caspase-14 reactivity increased and remained at least partially cytosolic, the involucrin labeling decreased towards the mature stratum corneum. The low involucrin labeling is probably caused by crosslinking through transglutaminase in the course of its integration into the cornified cell envelope (Ishida-Yamamoto and Iizuka 1998; Ishida-Yamamoto et al. 2000; Kalinin et al. 2002). Caspase-14 seemed to remain soluble, making it plausible that caspase-14 plays a role in the mature corneocyte. Consistent with such a possible role, preparations of caspase-14 from the stratum corneum contained caspase-14 fragments of the size of mature subunits, suggesting that the enzyme is in its active state (Eckhart et al. 2000a).
In addition to the cytoplasm, caspase-14 was also detected in the nucleus of granular layer cells as well as in nuclear remnants within cells of the transitional layer and corneus layer. It is therefore possible that caspase-14 is involved in regulation of the breakdown of the nucleus in terminally differentiating keratino-cytes (Fukuyama et al. 1984; Karasek 1988; Holbrook 1989). A closely related protease, caspase-3, is known to cleave ICAD, the inhibitor caspase-activated DNase (CAD) during regular apoptosis (Enari et al. 1998). Whether caspase-14 initiates an analogous degradation pathway in transitional layer keratinocytes remains to be determined by molecular biological studies. Alternatively, it is conceivable that caspase-14 targets structural proteins, such as periphilin, which localize to the nucleus before being transferred to the cell surface (Kazerounian and Aho 2003). Strikingly, periphilin was found to be susceptible to processing by caspase-5 in vitro. We will investigate whether this potential substrate can also be cleaved by caspase-14.
The finding that some immunolabeling for caspase-14 was associated with desmosomes and the cornified envelope may support the hypothesis that proteins eventually constituting these structures are substrates for this enzyme (Warhol et al. 1985; Steven et al. 1990; Ishida-Yamamoto and Iizuka 1994, 1998). The detection of caspase-14 near corneodesmosomes may also suggest that this enzyme intervenes with desquamation of corneocytes.
Ultrastructural distribution of caspase-14 in the corneous layer. Anti-human caspase-14 antibody. Corneocytes (C) of the low stratum corneum with a diffuse immunolabeling, also present along the corneous membrane (arrows) but more intensely over a structure that appears as nuclear remnants (arrowheads). Bar = 200 nm.
Ultrastructural distribution of caspase-14 in the corneous layer. Anti-human caspase-14 antibody. Detail of labeling (arrowhead) in corneocytes (C) near the corneous membrane. Bar = 200 nm.
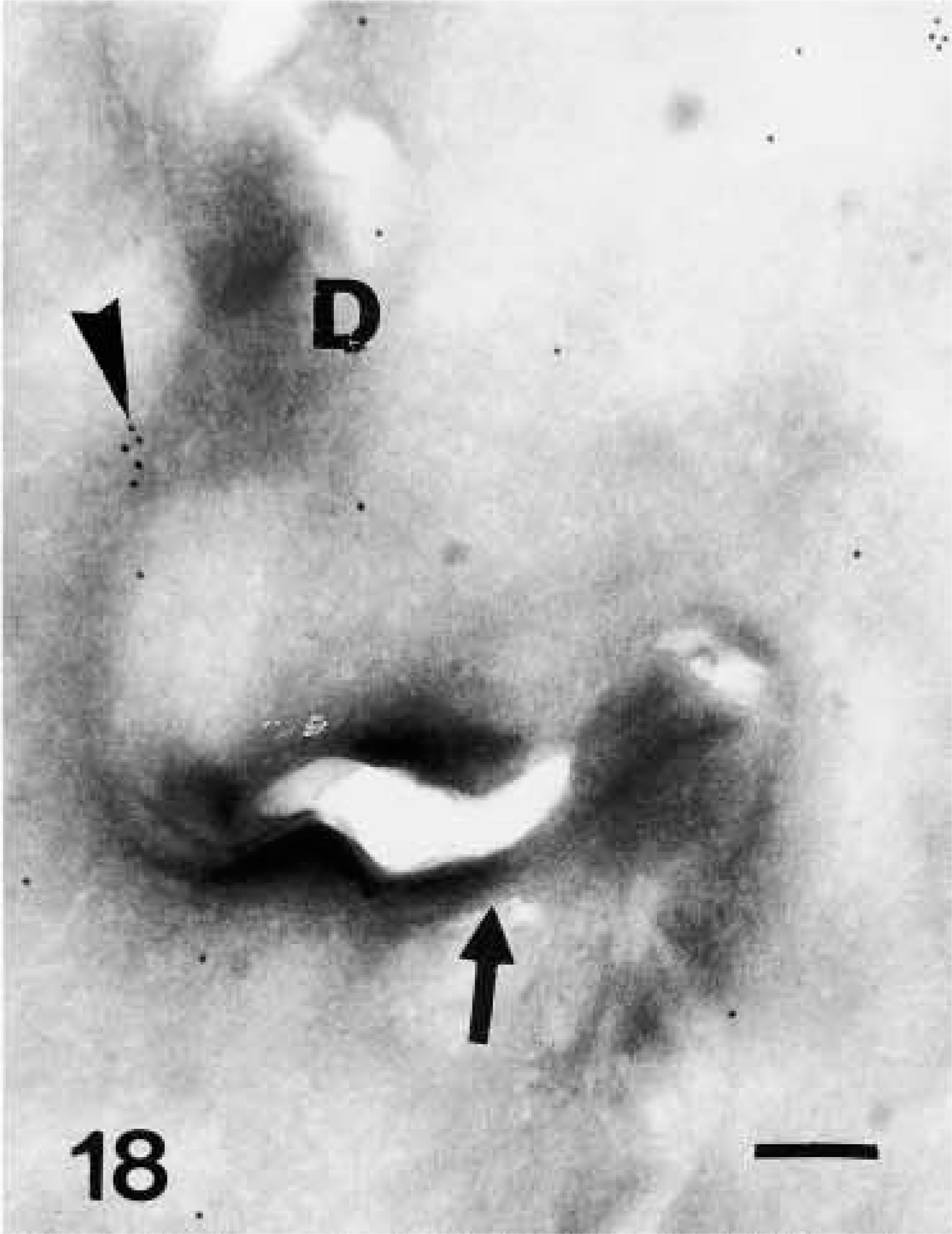
Ultrastructural distribution of caspase-14 in the corneous layer. Anti-human caspase-14 antibody. Detail of labeling (arrowhead) near a corneodesmosome (D) along the tortuous perimeter (arrow) of mature corneocytes. Bar = 200 nm.
The following technical issues must be considered when the data presented here are interpreted. First, it must be noted that our investigation of Bioacryl-embedded epidermis did not permit analysis of lamellar bodies in the granular layers. These organelles can be revealed only by standard osmium staining or by specific staining with ruthenium tetroxide, whereby both techniques destroy the immunoreactivity of typical antigens (Menon and Norlen 2002). Although lamellar bodies contain a large number of hydrolytic enzymes (Holbrook 1989; Menon and Norlen 2002), they are considered more likely to be involved in the lipid metabolism of corneocytes than in protein degradation during cornification. Second, the precise localization of involucrin along the cornified envelope can be revealed only by demasking agents such as proteases, which were not used in our study. Our interpretation that caspase-14 and involucrin do not colocalize in the stratum corneum is based on published studies on involucrin in which this methodology was used (Ishida-Yamamoto and Iizuka 1994, 1998; Ishida-Yamamoto et al. 1996, 2000). Third, when tissues are embedded in Bioacryl it is difficult to differentiate between F-keratohyalin granules and fragments of nuclear material that may even be part of composite keratohyalin. In our investigations, some components of composite keratohyalin granules (F-granules) appeared as reticular organelles made up of coarse filaments of ~10-nm diameter. These data are in agreement with earlier reports on the ultrastructure of keratohyalin in mammalian epidermis (Fukuyama and Epstein 1986; Holbrook 1989; Steven et al. 1990; Manabe et al. 1991).
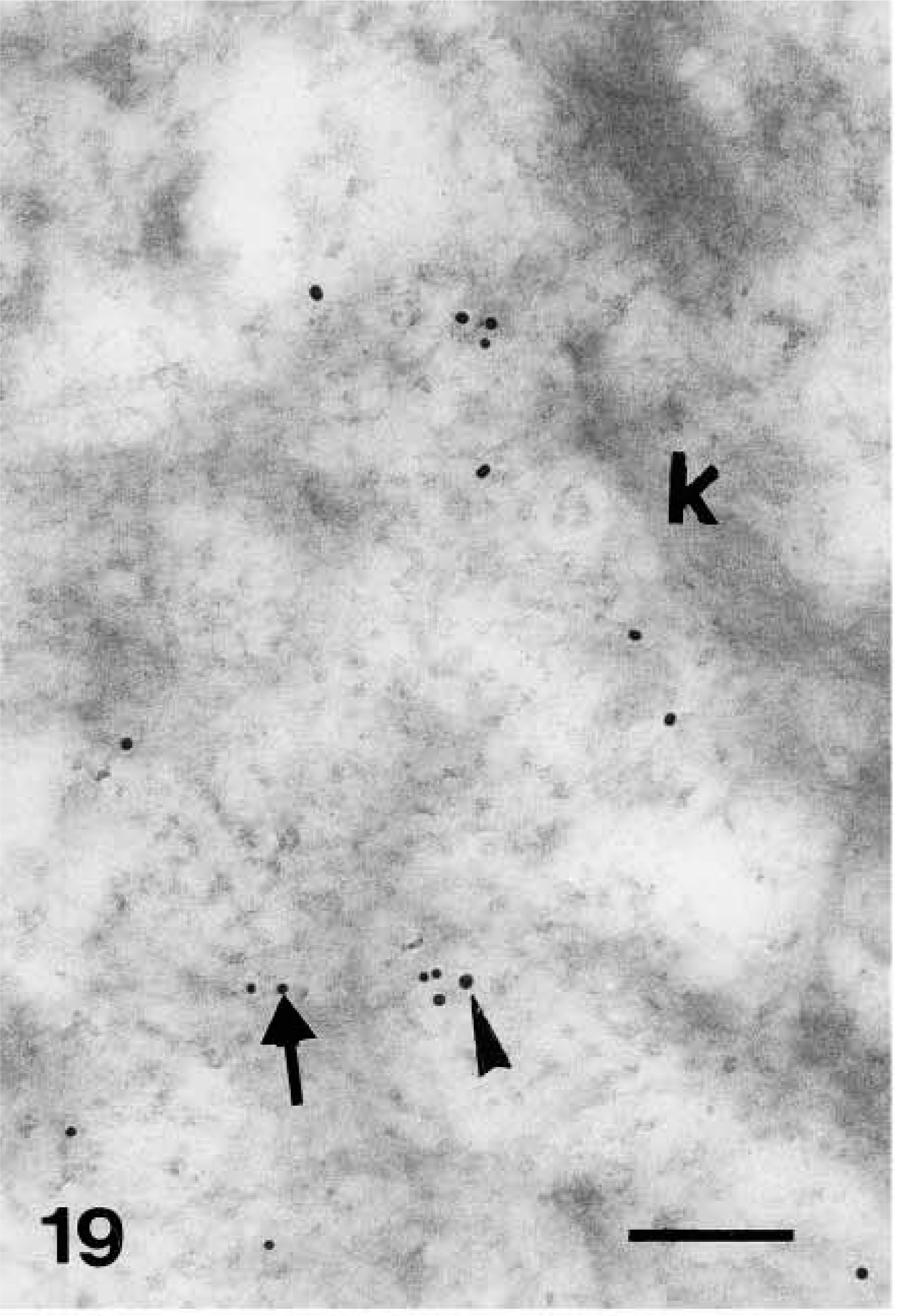
Double-labeling localization of involucrin (10-nm large gold particles) and caspase-14 (anti-mouse caspase-14 antibody, 20-nm large gold particles) in human epidermis. Detail of cytoplasm of an upper spinous layer cell double-labeled against involucrin (arrow on smaller gold particles) and caspase-14 (arrowhead on larger gold particles). Bar = 200 nm.
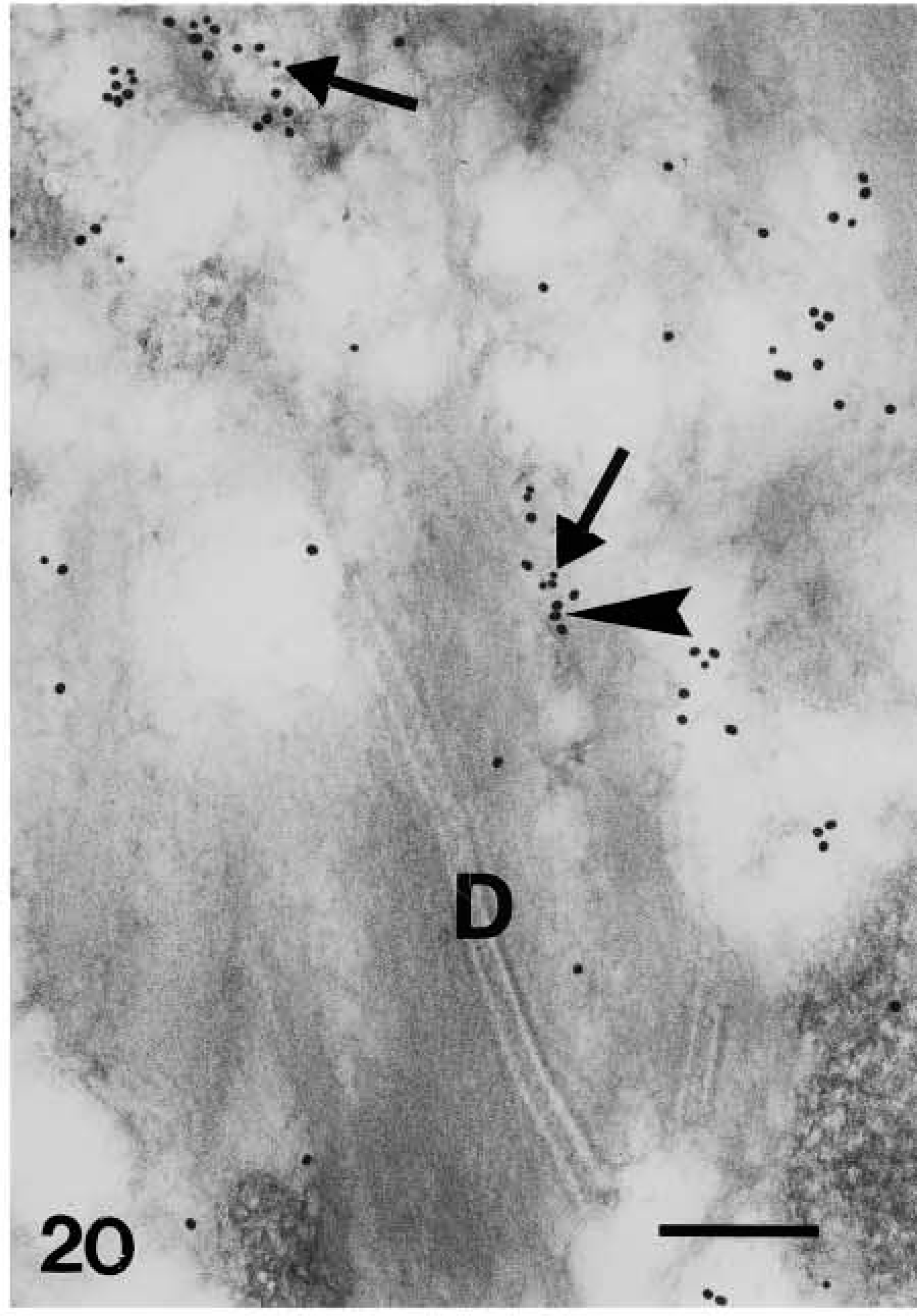
Double-labeling localization of involucrin (10-nm large gold particles) and caspase-14 (anti-mouse caspase-14 antibody, 20-nm large gold particles) in human epidermis. Detail of desmosome (D) in upper granular cell joined to keratin bundles associated with double labeling for caspase-14 (arrowhead on larger gold particles) and involucrin (arrow on smaller gold particles). Bar = 200 nm.
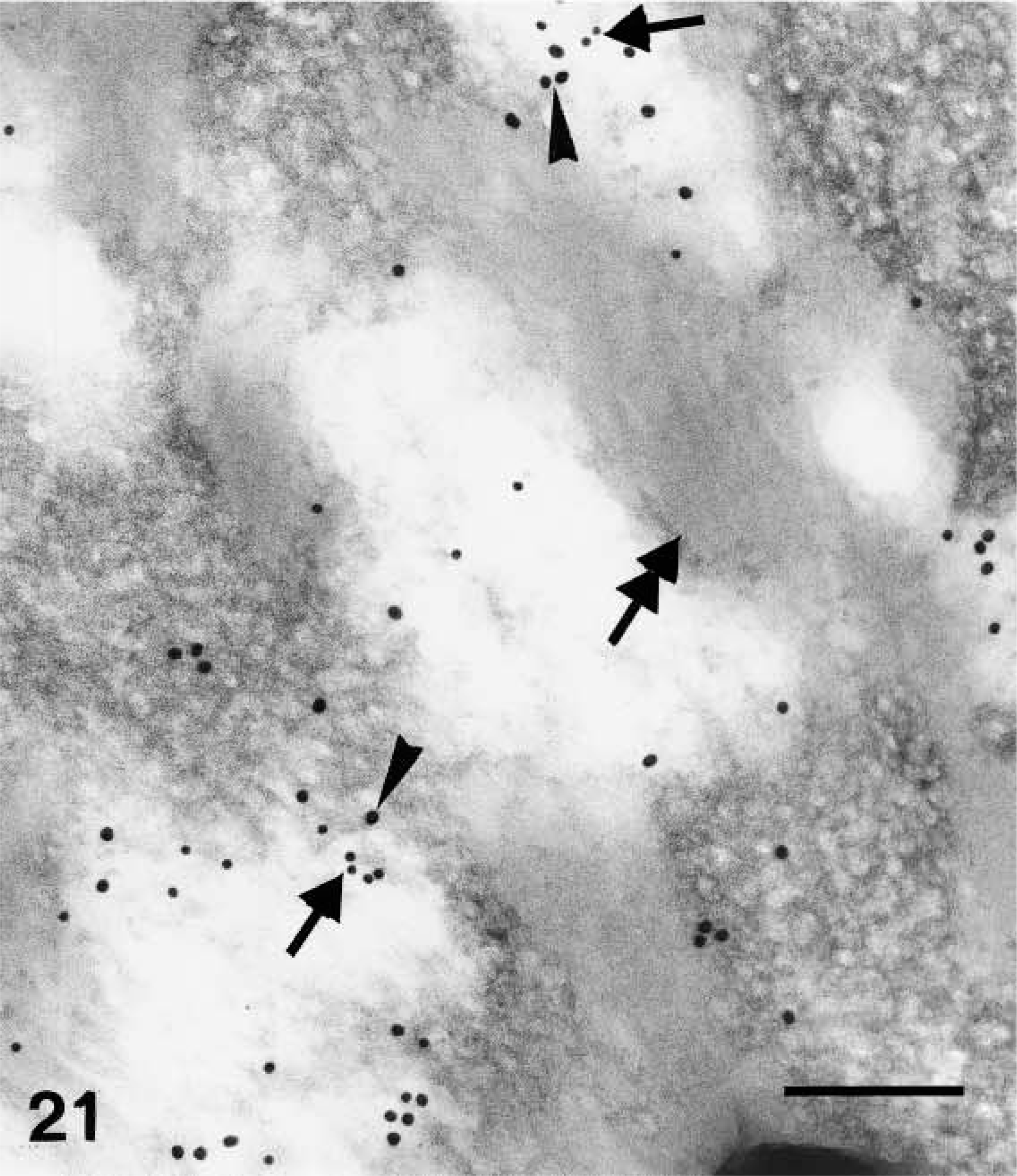
Double-labeling localization of involucrin (10-nm large gold particles) and caspase-14 (anti-mouse caspase-14 antibody, 20-nm large gold particles) in human epidermis. Double-labeling of keratohyalin showing involucrin (arrows on smaller gold particles) and caspase-14 (arrowhead on larger gold particles) especially localized near the amorphous material (double arrow) associated with the central network of F-granules. Bar =200 nm.
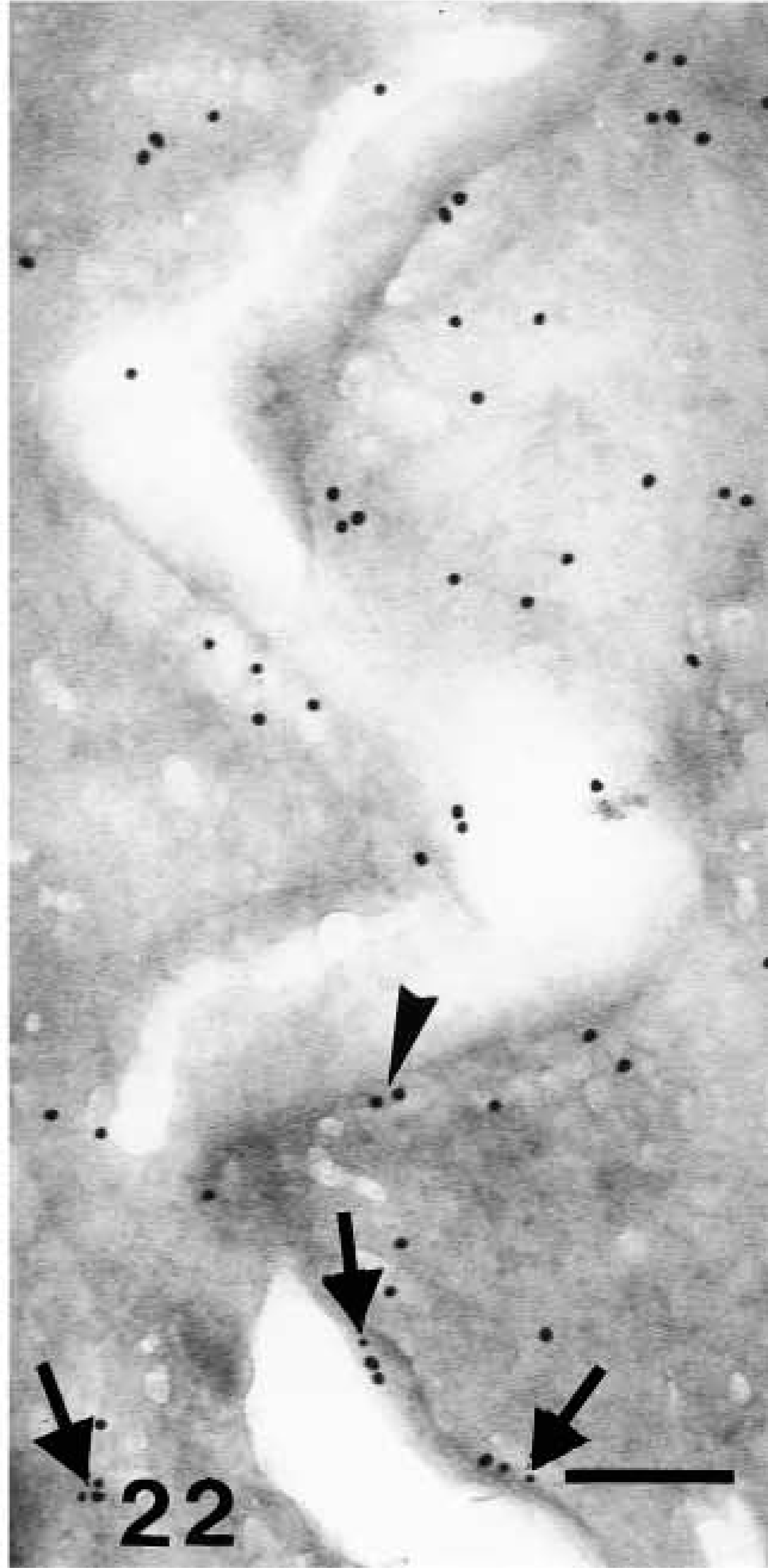
Double-labeling localization of involucrin (10-nm large gold particles) and caspase-14 (anti-mouse caspase-14 antibody, 20-nm large gold particles) in human epidermis. Detail of caspase-14 labeling (arrowhead on larger gold particles) along the cornified envelope of mature corneocytes, while little involucrin labeling is detectable (arrows on smaller gold particles). Bar = 200 nm.
Because the available anti-caspase-14 antibodies do not discriminate between proenzyme and catalytically active subunits in IHC, our data on the intracellular distribution of caspase-14 do not reveal the site of initial procaspase-14 activation. Previously we have shown by Western blotting analysis that caspase-14 subunits appear concomitant with cornification and that almost all caspase-14 proenzyme is converted to the subunits in mature stratum corneum (Eckhart et al. 2000a). Ongoing studies in our laboratories are aimed at the identification of caspase-14 protein species in preparations of cytoplasmic and nuclear proteins of terminally differentiating keratinocytes. The specific degradative substrates (Chien et al. 2002; Kuechle et al. 2001) of caspase-14 will be identified through biochemical and molecular biological studies. With the information provided here, the screening of substrates can be focused on a subset of proteins localizing to certain compartments of terminally differentiated keratinocytes.
Footnotes
Acknowledgements
Supported by a 60% University of Bologna Grant.
We thank Martin Stichenwirth, Heinz Fischer, Daniela Burtscher, and Andreas Kapetanopoulos for technical support and helpful discussions, and Wim Declercq (Flanders Interuniversity Institute for Biotechnology and Ghent University; Ghent, Belgium) for providing the antibody against murine caspase-14.