
Abstract
This article describes new ultrastructural staining methods for osmicated tissues based on the incubation of sections with sodium metaperiodate and sodium borohydride solutions before uranyl/lead staining. Sections incubated with sodium metaperiodate and sodium borohydride, treated with Triton X-100, and stained with ethanolic uranyl acetate/lead citrate showed a good contrast for the nucleolus and the interchromatin region, whereas the chromatin masses were bleached. Chromatin bleaching depended on the incubation with these oxidizing (metaperiodate) and reducing (borohydride) agents. Other factors that influenced the staining of the chromatin masses were the en bloc staining with uranyl acetate, the incubation of sections with Triton X-100, and the staining with aqueous or ethanolic uranyl acetate. The combination of these factors on sections treated with metaperiodate/borohydride provided a different appearance to the chromatin, from bleached to highly contrasted. Most cytoplasmic organelles showed a similar appearance with these procedures than with conventional uranyl/lead staining. However, when sections were incubated with metaperiodate/borohydride and Triton X-100 before uranyl/lead staining, the collagen fibers, and the glycocalix and zymogen granules of pancreatic acinar cells, appeared bleached. The possible combination of these methods with the immunolocalization of the amino acid taurine was also analyzed.
C
Treating ultrathin sections with sodium metaperiodate (oxidizing agent) or sodium borohydride (reducing agent) is a common procedure in some immuno-cytochemical methods (reviewed in Merighi 1992; Priestley et al. 1992), and the sequential use of sodium metaperiodate and tritiated sodium borohydride specifically radiolabels glycoproteins (Hindenburg et al. 1985; Kress et al. 1991). However, the effects of the sequential incubation with these reagents on the staining of sections have not been previously examined. We have observed that some postembedding immunocytochemical methods, which involve the incubation of sections in sodium metaperiodate and sodium borohydride solutions before immunolabeling (Lobo et al. 2000a), result in chromatin bleaching. However, the immunogold methods used in other studies that included the use of sodium metaperiodate but not sodium borohydride (Galán et al. 1996; Lobo et al. 1997; Goldstein et al. 1999) provide an electron-dense contrast to chromatin masses. This observation led us to develop new procedures as simple and reproducible staining methods for the cell nucleus and other cellular structures. Here we describe these procedures.
Materials and Methods
Sprague-Dawley and albino Wistar rats (250 g) were anesthetized with a mixture of ketamine, xylocaine, and atropine and perfused through the ascending aorta with 100 ml PBS, followed by the fixative solution: 4% paraformaldehyde- 0.5% glutaraldehyde in PBS, pH 7.4. The brain and pancreas were dissected and immersed in the same fixative used during perfusion for 6–24 hr at 4C. Samples from a dog stomach were removed under sodium pentobarbital anesthesia and immersed in the above-mentioned fixative for 6–24 hr at 4C. After rinsing in PBS, some tissue samples (1 mm3) were postfixed in buffered 1% osmium tetroxide at 4C for 2 hr and other samples were not osmium-fixed. All samples were dehydrated through a graded acetone series. To analyze the effects of the en bloc staining, some samples were stained during dehydration with a saturated solution of uranyl acetate in 70% acetone and other samples were not stained. Moreover, some samples were partially dehydrated (up to 80% acetone) and embedded at 55C in a glycol methacrylate-derived resin, Unicryl (British Biocell International; Cardiff, UK), and other samples were completely dehydrated and embedded in araldite according to conventional methods (Gonzalez-Santander et al. 1997). Tissue blocks were sectioned and serial ultrathin sections of similar thickness, giving gold interference color, were collected on nickel grids. Grids were incubated in aqueous 5% sodium metaperiodate for 5 min (Unicryl sections) or 15–20 min (araldite sections) and rinsed in distilled water and in Tris-buffered saline (TBS), pH 7.6. Grids were then incubated in freshly prepared 0.2% sodium borohydride in TBS, pH 7.6, for 10 min. During incubation a gentle shaking of grids was performed and/or the grids were raised from the solution drops and placed again on the drops. This was done to release the bubbles formed because they could prevent the contact between the sections and the sodium borohydride solution. The effects of detergent (Triton X-100) treatment before staining were also analyzed. Some grids were rinsed in TBS, pH 7.5, for 10 min, incubated in TBS with 0.05% Triton X-100 for 30 min, and washed again in TBS and in distilled water. Other grids were rinsed in TBS for 10 min and in distilled water for 5 min. Staining was carried out at room temperature (RT) with saturated solutions of uranyl acetate in 70% ethanol or in distilled water for 20 min, followed by Reynold's lead citrate for 15 min. Sections were observed in a Zeiss EM10 transmission electron microscope (TEM). Some grids were not incubated with sodium metaperiodate and/or sodium borohydride solutions. These sections were stained in the same way for comparative purposes and used as controls.
The oxidation/reduction step with metaperiodate/borohydride has been used in some immunocytochemical procedures to analyze the cellular localization of the amino acid taurine (Lobo et al. 2000a) and the translational factor eukaryotic initiation 2α (Lobo et al. 2000c). To illustrate the possible combination of the staining procedures shown here with immunocytochemical methods, some sections from osmicated samples were immunostained for taurine as previously reported (Lobo et al. 2000a). Briefly, grids were incubated in sodium metaperiodate and sodium borohydride solutions as described above. They were then incubated for 30 min at RT in 3% normal goat serum with 0.05% Triton X-100 in TBS, pH 7.4, followed by incubation at 4C for 12 hr with the rabbit anti-taurine antibody (Chemicon International; Temecula, CA) diluted 1:200 in TBS, pH 7.5. Afterwards the grids were rinsed in TBS (pH 7.5 and 8.2) for 20 min and incubated at RT for 2 hr in 15-nm gold-conjugated goat anti-rabbit IgG (British Biocell) diluted 1:40 in TBS, pH 8.2. After extensive rinsing in TBS (pH 7.6 and 7.4) sections were fixed in 1% glutaraldehyde at RT for 10 min and washed in distilled water. Finally, the sections were counter-stained as described above. Controls to demonstrate the specificity of the immunolabeling were performed by omitting the primary antibody or using this antibody preabsorbed with an excess of antigen. The specificity of the anti-taurine antibody used here has been extensively demonstrated by ELISA (Campistron et al. 1986). Sections untreated with metaperiodate or borohydride solutions were immunolabeled as above and used for comparative purposes. For quantitative analysis, the number of gold particles per square micrometer in different cell types and in empty resin (to evaluate background) was calculated according to the morphometric technique described by Bendayan et al. (1980). For each group of sections (those treated with metaperiodate/borohydride and those incubated only with metaperiodate or borohydride), 30 micrographs of each cell type (vascular endothelial cells, neurons, and astrocytes) were analyzed. Values obtained were expressed as mean ± SD.
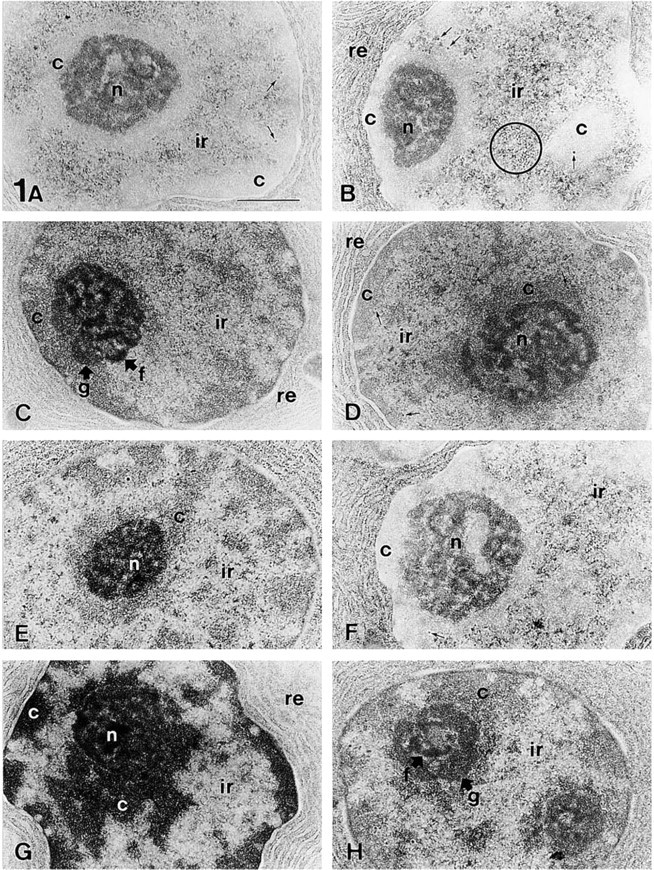
Acinar cells of the rat exocrine pancreas. All samples were fixed with glutaraldehyde/paraformaldehyde/osmium and embedded in araldite. (
Results
The staining procedures described here were tested in several tissues, such as the pancreas (Figure 1 and Figure 2A–2D), stomach (Figures 2E–2G), and brain (Figure 3). Sections from osmicated samples incubated with sodium metaperiodate and sodium borohydride solutions, washed with solutions containing Triton X-100, and stained with ethanolic uranyl acetate/lead citrate, showed a good contrast for the nucleolus and the interchromatin region, whereas the condensed chromatin masses appeared bleached (Figures 1A, 1B, 2D, 3A, and 3C). Therefore, this procedure provided different results than conventional uranyl/lead staining, which led to highly contrasted chromatin masses (Figure 2B). Using this method in sections from samples not fixed with osmium, we did not observe chromatin bleaching and the results were similar to those obtained using conventional TEM staining (Figure 2E). In sections from osmicated samples, chromatin bleaching depended on the incubation with these oxidizing and reducing agents because when any of them was not used chromatin bleaching was not observed (Figure 3B). Moreover, when sections were incubated with sodium borohydride before incubation with sodium metaperiodate, chromatin bleaching was not observed. Several solutions and incubation times were tested, the best results being obtained using 5% sodium metaperiodate for 5 min (Unicryl sections) or 15–20 min (araldite sections), and 0.2% sodium borohydride in TBS for 10 min, as described in Materials and Methods. Shorter incubation time (e.g., araldite sections incubated for 5 min in each of the above-mentioned solutions) did not result in chromatin bleaching. Using the procedure recommended here in some tissue samples, such as the lamina propria of the dog stomach, some cells showed chromatin bleaching, whereas adjacent cells did not show chromatin bleaching (Figure 2F). However, by increasing the incubation time or the concentration of the metaperiodate/borohydride solutions, the chromatin of all cells became bleached. Other factors influenced the staining of the chromatin masses, such as (a) the en bloc staining with uranyl acetate, (b) the incubation with Triton X-100 before uranyl/lead staining, and (c) the staining with aqueous or ethanolic uranyl acetate (Figure 1). En bloc staining resulted in increased chromatin bleaching compared with samples that were not en bloc-stained (compare Figure 1F with 1E and Figure 1H with 1G). The incubation with solutions containing Triton X-100 also resulted in increased chromatin bleaching compared to sections untreated with detergent (compare Figure 1A with 1E and Figure 1C with 1G). Moreover, aqueous uranyl acetate gives higher electron opacity to the chromatin masses than ethanolic uranyl acetate (compare Figure 1C with 1A, Figure 1D with 1B, Figure 1G with 1E, and Figure 1H with 1F). Each possible combination of these factors on sections treated with metaperiodate/borohydride solutions provided a different appearance to the cell nucleus, mainly to the chromatin masses (Figure 1). With all these methods, the components of the interchromatin region, such as the perichromatin and interchromatin granules and fibers, were well contrasted. With all the methods used, the nucleolus exhibited high contrast and three main domains were clearly visible in this nuclear compartment: the dense fibrillar component, the granular component, and the fibrillar centers (Figure 1). Most cytoplasmic organelles (such as the endoplasmic reticulum, Golgi complex, and mitochondria) showed a similar appearance using these procedures than with conventional uranyl/lead staining (Figures 1–3). However, when sections were incubated with metaperiodate/borohydride and Triton X-100 before staining with ethanolic uranyl acetate/lead citrate, the collagen fibers (Figure 2F) and the glycocalix and mature zymogen granules of pancreatic acinar cells (Figure 2A) appeared bleached. The bleached glycocalix was difficult to visualize. However, when the secretory products present in the lumen were highly stained, the bleached glycocalix became visible against a dark background (Figure 2A). In the exocrine pancreas, the zymogen granules of acinar cells were bleached (Figure 2A), whereas in the pancreatic islets, the secretory granules of endocrine cells were highly contrasted (Figure 2C). The bleaching of these structures also depended on the incubation with the oxidizing and reducing agents because when any of them was not used
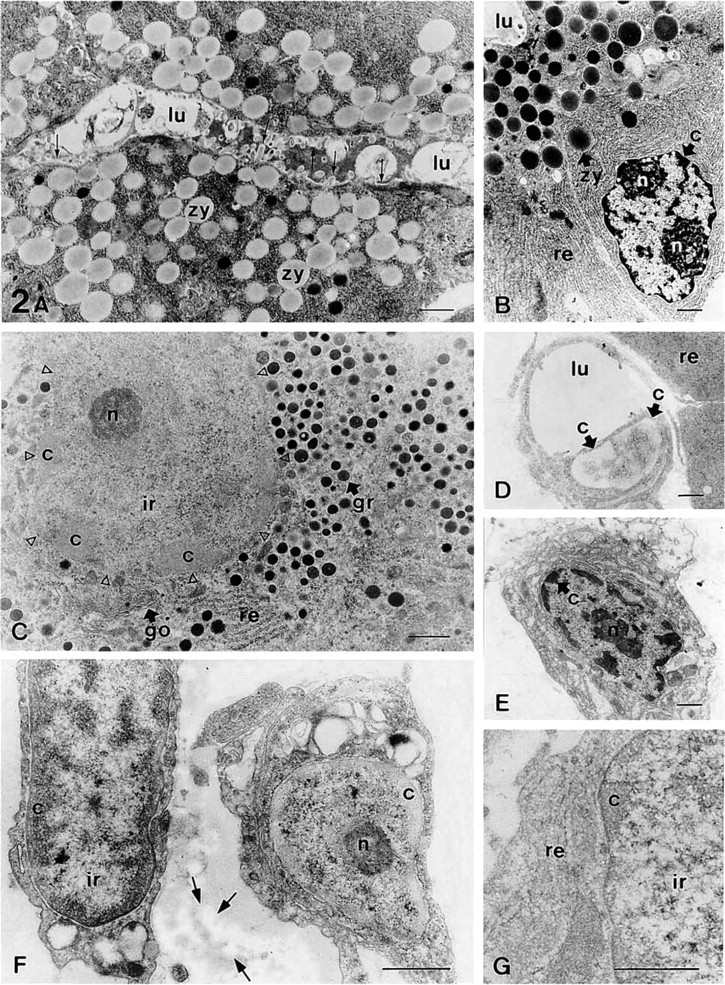
Sections of the rat pancreas (
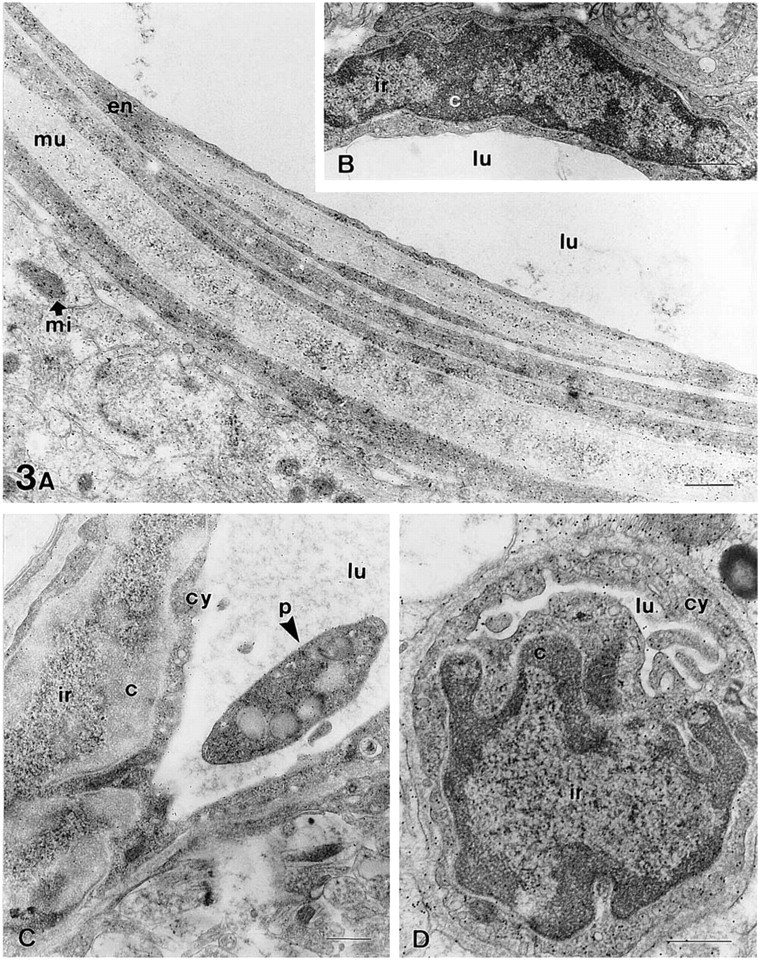
Arteriole (
they were well contrasted. Furthermore, these methods provided similar results in sections from samples embedded either in araldite or Unicryl resin [compare the bleached chromatin shown on araldite sections (Figures 1A, 1B, 3A, and 3C) and on Unicryl sections (Figure 2C)].
In sections from osmicated samples, the combination of the oxidation/reduction treatment with the immunolocalization of the amino acid taurine was analyzed and the results compared with the corresponding immunogold assays on sections untreated with metaperiodate or borohydride. The immunolabeling pattern on treated sections (Figures 2F, 3A, 3C, and 3D) was similar to that found on sections untreated with sodium borohydride (Figure 3B). No staining was found in sections untreated with sodium metaperiodate. This is consistent with the notion that etching sections with metaperiodate is necessary to restore antigenicity on osmicated samples (Merighi 1992). In general, taurine appeared randomly distributed throughout the cell; in the nucleus, cytoplasm and mitochondria (Figure2Fand3). In some cell types (such as vascular endothelial cells) a variable degree of taurine staining was observed. Thus, some cells were intensely immunoreactive, whereas others were not stained. This pattern of distribution is similar to that reported for endothelial cells in other tissues (Lobo et al. 2000b). In the samples from the rat hippocampus (CA1 region) treated with the oxidation/reduction method (Figure 3A, 3C, and 3D), the stained endothelial cells contained a mean density of 42.1 ± 1.21 particles/μm2 in the cytoplasm and 44.3 ± 2.1 in the nucleus. Similar results were found in serial sections treated only with metaperiodate, 39.8 ± 1.57 for the cytoplasm and 41.2 ± 1.9 for the nucleus (Figure 3B). In sections processed with the metaperiodate/borohydride treatment, neuronal perikarya showed a mean of 19.3 ± 0.85 particles/μm2 in the cytoplasm and 19.8 ± 1.17 in the nucleus, and astrocytes contained 22.9 ± 0.73 in the cytoplasm and 21.9 ± 1.14 in the nucleus. Similar values were found in sections treated only with metaperiodate (neuron/cytoplasm 18.9 ± 0.3, neuron/nucleus 18.3 ± 0.5, astrocyte/cytoplasm 24.4 ± 1.3, astrocyte/nucleus 23.7 ± 0.2). Background values were very low (1.38 ± 0.2 in sections treated with metaperiodate/borohydride and 1.8 ± 0.9 in those treated with metaperiodate). These values are similar to those reported, using different immunocytochemical procedures, in the rat spinal cord (Lee et al. 1992). These results demonstrate that neither the pattern nor the intensity of taurine labeling was different in sections treated with the oxidation/reduction step and in those treated only with metaperiodate. No staining was found in the negative controls performed (Figure 2G). Internal positive controls for taurine immunolabeling consisted of the vascular smooth muscle cells (Figure 3A) and the platelets (Figure 3C) found in some vessels. In the smooth muscle cells (Figure 3A), taurine was detected as previously described (Lobo et al. 2000a). Although there are no previous studies about the distribution of taurine in platelets, platelets are highly enriched in taurine, as demonstrated by chromatography (reviewed in Wright et al. 1986), and we found a high labeling density in these cells (Figure 3C).
Discussion
This article describes new staining methods based on the sequential incubation of ultrathin sections with sodium metaperiodate and sodium borohydride solutions before uranyl/lead staining. These methods provide similar and reproducible results in different tissue samples. The staining patterns obtained with some of these procedures are similar to those achieved by the EDTA regressive staining for ribonucleoproteins (Bernhard 1969) because both result in chromatin bleaching, whereas the nucleolus and the interchromatin structures appear well contrasted. Other methods tested result in highly contrasted chromatin masses, showing an ultrastructural appearance for the cell nucleus similar to that obtained using conventional TEM methods. Therefore, these methods provide a different appearance to the chromatin masses, from bleached to highly contrasted, whereas the nucleolus and the interchromatin region are well contrasted. Some of these methods also modify the staining properties of other cellular and extracellular structures such as the zymogen granules, the glycocalix, and the collagen fibers. Thus, using the metaperiodate/borohydride/Triton X-100 treatment before uranyl/lead staining these structures appear bleached, whereas with conventional TEM methods they are well contrasted.
The interactions between the salts of heavy metals employed for cytological staining and components of biological tissues are not fully understood (reviewed in Robards and Wilson 1993). However, uranyl acetate is known to stain nucleic acids selectively, probably by ionic bonding of the positively charged uranyl ions to negatively charged phosphate groups. It also stains proteins, probably by bonding to their free carboxyl and amino groups. Secondary staining with lead salts, which have an affinity for other cell components such as lipids and glycogen, further improves the overall contrast of sections. In addition, alcoholic staining solutions penetrate more rapidly and produce more intense staining than aqueous solutions with the same concentration. No other differences between the staining properties of aqueous uranyl solutions and the alcoholic ones have been reported. Moreover, etching of sections with Triton X-100 before immunolabeling and staining is a common procedure used to enhance penetration of immunoreagents. These methods (uranyl/lead staining with or without detergent pretreatment) result in highly contrasted chromatin masses as well as other cell structures. Therefore, the main difference between the procedures presented here and those previously reported for electron microscopy is that we introduce an oxidation/reduction step before immunolabeling and staining. This step would determine some chemical modifications of specific cellular components that modify their staining properties. In this sense, it has been reported that the use of sodium metaperiodate and tritiated sodium borohydride specifically radiolabels glycoproteins (Hindenburg et al. 1985; Kress et al. 1991). Moreover, sodium metaperiodate has been used in several staining techniques for polysaccharides and glycoproteins, which are derived from the PAS method (periodic acid-Schiff) of classical histology (Thiéry 1967; Pearse 1980). According to the well-established chemical reactions of these procedures, sodium metaperiodate acts in the oxidation of certain alcohol groups (glycols) of polysaccharides into aldehyde groups. These aldehydes could then react with sodium borohydride. Therefore, this oxidation/reduction step probably acts on glycoproteins. The bleaching of the zymogen granules and the glycocalix (structures rich in glycoproteins and muco-polysaccharides) observed with some of the procedures described here appears to support this notion. However, the mechanism of these staining methods is thus far unclear.
Etching ultrathin sections with sodium metaperiodate is a critical step in some antigen retrieval methods (Merighi 1992; Stirling and Graff 1995). However, it is a highly aggressive procedure because sections treated with this solution for too long a time show clefts and holes that make them unstable under the electron beam (Merighi 1992). Therefore, the duration of the metaperiodate treatment is important to avoid damage to the sections (Merighi et al. 1989; Merighi 1992). We tested several incubation times of the sodium metaperiodate and sodium borohydride solutions and the best results were obtained using the procedures described above.
The performance of immunocytochemical procedures requires that the antigenic properties of the samples should not be affected by the processing techniques. This fact often leads to the choice of a compromise between good ultrastructural preservation and maintenance of the tissue antigenic properties. In this sense, it has been shown that, although the use of osmium tetroxide provides better ultrastructural morphology, antigenicity may be extensively reduced by osmium postfixation (Merighi 1992). Some oxidizers have been used to restore antigenicity on ultrathin sections from osmium-fixed samples, but sodium metaperiodate appears to provide the most satisfactory results (Pelletier et al. 1981; Bendayan and Zollinger 1983; Merighi 1992). Sodium metaperiodate appears to restore tissue antigenicity by removing osmium from the sections (Bendayan and Zollinger 1983) and enhancing the hydrophilic nature of the embedding resin (Causton 1984), hydrophilicity allowing an easier antigen-antibody interaction. Moreover, sodium borohydride solutions have been used in several immunocytochemical procedures, mainly when tissue samples are fixed using glutaraldehyde or glutaraldehyde-paraformaldehyde mixtures (Kosaka et al. 1986; Priestley et al. 1992). Treating sections with this solution appears to diminish nonspecific immunostaining by reducing unreacted aldehyde groups. It also reduces glutaraldehyde-created double bonds to single bonds, which partially restore the tertiary structure of proteins (restoring antigenicity) and increase permeability and reagent penetration (Kosaka et al. 1986; Priestley et al. 1992). Thus, etching the sections by immersion in these solutions is not only an oxidation/reduction step of the staining procedures presented here but also could act as an antigen retrieval procedure.