
Abstract
We have previously demonstrated by immunohistochemistry the presence of secreted carbonic anhydrase (CA VI) in the acinar cells of the rat lacrimal glands. In this study we purified the sheep lacrimal gland CA VI to homogeneity and demonstrated by Western analysis that it has the same apparent subunit molecular weight (45 kD) as the enzyme isolated from saliva. RT-PCR analysis showed that CA VI mRNA from the lacrimal gland was identical to that of the parotid gland CA VI mRNA. An RIA specific for sheep CA VI showed the lacrimal gland tissue concentration of the enzyme to be 4.20 ± 2.60 ng/mg protein, or about 1/7000 of the level found in the parotid gland. Immunohistochemistry (IHC) and in situ hybridization (ISH) showed that lacrimal acinar cells expressed both immunoreactivity and mRNA for CA VI. Moreover, CA VI immunoreactivity was occasionally observed in the lumen of the ducts. Unlike the parotid gland, in which all acinar cells expressed CA VI immunoreactivity and mRNA, only some of the acinar cells of the lacrimal gland showed expression. These results indicate that the lacrimal gland synthesizes and secretes a very small amount of salivary CA VI. In tear fluid, CA VI is presumed to have a role in the maintenance of acid/base balance on the surface of the eye, akin to its role in the oral cavity.
Keywords
C
The secreted isozyme was first described in the saliva of sheep (Fernley et al. 1979). Since that time it has been isolated and characterized in a number of species, including sheep (Fernley et al. 1988a), rat (Feldstein and Silverman 1984), human (Kadoya et al. 1987; Murakami and Sly 1987), and cow (Hooper et al. 1995). The enzyme appears to form oligomers in solution, is a glycoprotein and, at least in the sheep, has a disulfide bond linking two cysteine residues (Fernley et al. 1988b). The cDNA sequence for human, bovine, and mouse CA VI includes a leader sequence typical of secreted proteins (Aldred et al. 1991; Jiang et al. 1996; Sok et al. 1999). This sequence is not present in the mature protein (Fernley et al. 1989). The enzyme is believed to have a role in the maintenance of oral pH (Feldstein and Silverman 1984). This concept may be supported by the recent observation that low salivary enzyme concentration is associated with increased susceptibility to dental caries (Kivelä et al. 1999).
It was reported previously that, in a screening of some 18 different tissues in the sheep, the enzyme was found only in the parotid and submandibular salivary glands (Fernley et al. 1989). The protein is known to be synthesized in the acinar cells of these serous salivary glands and secreted into saliva (Fernley et al. 1989; Parkkila et al. 1990; Aldred et al. 1991; Ogawa et al. 1992, 1993). On the other hand, Ogawa et al. (1995) detected a secreted form of CA in a minority of acinar cells of the rat lacrimal gland by IHC using a monoclonal antibody specific for rat CA VI. This may not be surprising because acinar cells of the lacrimal gland are similar in structure and function to the acinar cells of the salivary glands (Dartt 1989; Mircheff 1989). Indeed, a more recent study using RT-PCR and RT-PCR-Southern blotting analyses detected CA VI mRNA in human pancreas, which is another example of a serous gland that resembles the salivary gland (Fujikawa-Adachi et al. 1999a). However, the lacrimal gland CA VI has not been purified or characterized, probably because of the extremely small content in the gland. Therefore, conclusive evidence is lacking for the identity of lacrimal gland CA as CA VI.
In this study we have demonstrated by a number of techniques, including Western analysis, RT-PCR, IHC, and ISH, that CA VI is indeed synthesized by, stored in, and secreted from the sheep lacrimal gland.
Materials and Methods
Animals used were mature female sheep (Merino and Merino crossbred strains). CA VI was purified to homogeneity from parotid saliva as described previously (Fernley et al. 1988a). A specific polyclonal antibody to this protein had been raised and described (Fernley et al. 1988a). Lacrimal and parotid glands were excised immediately after death. For protein and RNA isolation, tissues were immediately frozen and stored at −80C until used. For IHC and ISH, tissues were fixed in Bouin's fixative and 4% paraformaldehyde-0.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.2), respectively, and embedded in paraffin. Sections (6 μm thick) were cut and mounted on silane-coated glass slides.
Western Analysis
Glands from three sheep were homogenized in 50 mM phosphate buffer (pH 7.4) containing 5 mM benzamidine, 1 mM phenylmethanesulfonyl fluoride, and 1 mg/ml aprotinin. The homogenate was centrifuged (20,000 × g for 15 min at 4C). Partially purified CA VI was prepared by ammonium sulfate precipitation, followed by sulfonamide-Sepharose affinity chromatography (Fernley et al. 1988a). The enzyme was further purified by anion-exchange chromatography (Mono Q; Pharmacia Fine Chemicals, Uppsala, Sweden). Enzyme activity was measured by the rate of pH change (7.4 to 7.0) in a buffered solution with saturating CO2 concentration. Protein samples were run on a 10.5% SDS-polyacrylamide gel and electroblotted onto Immobilon (Millipore; Bedford, MA). The blot was blocked with 10% skimmed milk powder in 20 mM Tris-HCl buffered saline (pH 7.5; TBS) and reacted with the anti-CA VI antibody (1:1000 in 3% skimmed milk powder in TBS). The blot was then reacted with peroxidase-conjugated goat anti-rabbit IgG (Bio-Rad Laboratories; Richmond, CA) at 1:2500 and the complex visualized by reaction with the Enhanced Chemiluminescence Detection System (Amersham International; Poole, UK) using the manufacturer's protocol.
Radioimmunoassay
Radioimmunoassay of sheep CA VI was carried out as described previously (Fernley et al. 1991). Lacrimal glands from 10 sheep were homogenized and centrifuged as described above and the supernatant was used for the analysis. The protein concentration was determined by a modified Lowry assay (Markwell et al. 1978). Samples were measured in at least two RIAs and the values averaged.
Immunohistochemistry
Deparaffinized sections were stained by an indirect immunoperoxidase technique (Ogawa et al. 1995) or an avidinbiotin-peroxidase technique. For the indirect immunoperoxidase technique, endogenous peroxidase activity was inactivated with 0.3% H2O2 and then nonspecific binding sites were blocked with normal swine serum (1:100 in 10 mM PSB, pH 7.2, containing 1% bovine serum albumin; PBS-BSA). The sections were reacted with the anti-CA VI antibody (1:100 in PBS-BSA). They were then reacted with peroxidase-conjugated swine anti-rabbit IgG (Dako; Glostrup, Denmark) at 1:200 in PBS-BSA containing 0.1% normal sheep serum. The immunoreaction sites were visualized by treatment with 3,3′-diaminobenzidine tetrahydrochloride-H2O2 solution (DAB-H2O2 solution). For the avidin-biotin-peroxidase technique, endogenous peroxidase and nonspecific binding sites were blocked with H2O2 and normal goat serum, respectively, as described above, and the sections were reacted with the primary antibody (1:1000 in PBS-BSA). Then the sections were sequentially reacted with biotinylated goat anti-rabbit IgG (1:200 in PBS-BSA containing 0.1% normal sheep serum; Vector Laboratories, Burlingame, CA) and avidin-biotinylated peroxidase complex (1:100 in PBS; Vector) for 45 min each. Visualization of the immunoreaction sites was carried out in DAB-H2O2 solution with or without nickel (Scopsi and Larsson 1986; Merchenthaler et al. 1989). Sections with or without methyl green counterstain were dehydrated and coverslipped with Permount. Negative controls were performed by substituting the primary antibody with normal rabbit IgG or primary antibody that had been pretreated for 24 hr at 4C with 0.125 or 0.25 mg/ml of CA VI.
RT-PCR Analysis
Total RNA was isolated from glands of two sheep using the Total RNA Isolation System (Promega; Madison, WI) and digested with RNase-free DNase I (Promega) for 15 min at 37C. Five μg of the denatured total RNA was reverse-transcribed in 20 μl of a reaction mixture containing 5 × RT buffer (375 mM KCl, 15 mM MgCl2, and 50 mM dithiothreitol in 250 mM Tris-HCl, pH 8.3), 0.5 mM each dNTP, 0.5 mg oligo(dT)15 primers, 100 U RNase inhibitor, and 200 U M-MLV reverse transcriptase (Wako Pure Chemicals; Osaka, Japan) for 1 hr at 37C and then heated for 10 min at 70C. After RT reaction, PCR was carried out in a total volume of 25 μl of a mixture containing 1 μl RT sample, 10 mM Tris-HCl (pH 8.3), 1.5 mM MgCl2, 50 mM KCl, 0.2 mM each dNTP, 0.4 mM each primer and 1.0 U AmpliTaq Gold DNA polymerase (Perkin-Elmer; Norwalk, CT). Forty cycles of denaturation (94C, 30 sec), annealing (60C, 1 min) and extension (72C, 1 min) were carried out in a DNA thermal cycler (Perkin-Elmer). The primer sequences for CA VI were 5′-GATACATAATTGAGATTCACGTCGTCCACTAC-3′ and 5′-GCTTGACAGTATCTGCTACCACAAACCA-3′. They were designed according to the nucleotide sequence of bovine submandibular CA VI (Jiang et al. 1996).
Aliquots (10 μl) of the PCR products were run on in a 2% agarose gel. The amplified products were excised from the gel, purified using Qiaex II (Qiagen; Hilden, Germany), and sequenced. The purified parotid product was subcloned into pT7Blue vector (Novagen; Madison, WI) using the Perfectly BluntTM Cloning Kit (Novagen).
In Situ Hybridization
Two types of recombinant pT7Blue vector, each containing a 261-bp coding region of sheep parotid CA VI with different orientations, were obtained by screening the pT7 library and were used as templates of antisense and sense ribo-probes. One mg of each plasmid was linearized with BamHI (Boehringer-Mannheim; Mannheim, Germany). In vitro transcription was performed with digoxygenin-11-UTP (Boehringer-Mannheim) using T7 RNA polymerase (Nip-pongene; Tokyo, Japan).
The following procedures were carried out at RT unless otherwise noted. Deparaffinized sections were washed in PBS, incubated in 1 μg/ml proteinase K (Nippongene) for 10 min, and washed in PBS. Nonspecific binding sites were blocked by acetylation with 0.25% acetic anhydride in 0.1 M triethanolamine (pH 8.0) for 15 min. The sections were washed in 4 × SSC (600 mM NaCl in 60 mM sodium citrate) and prehybridized in 50% formamide in 2 × SSC for 30 min at 42C. Hybridization was carried out in 40 μl of a solution containing 50% formamide, 2 × SSC, 1 mg/ml tRNA, 1 mg/ml salmon sperm DNA, 1 mg/ml BSA, 10% dextran sulfate, and 1 μg/ml probe for 16 hr at 42C. The sections were washed three times in 50% formamide in 2 × SSC for 20 min each at 42C, treated for 30 min at 37C with 20 μg/ml RNase A (Nippongene) in 10 mM Tris-HCl (pH 8.0) containing 0.5 M NaCl and 1 mM EDTA, and washed three times in 0.1 × SSC for 20 min each at 42C. Detection of the labelled probe was carried out using the DIG Nucleic Acid Detection Kit (Boehringer-Mannheim) according to the manufacturer's protocol with modifications. The modifications were as follows: (a) maleic acid buffer and detection buffer were substituted with 0.1 M Tris-HCl (pH 7.5) containing 0.15 M NaCl and 0.1 M Tris-HCl (pH 9.5) containing 0.1 M NaCl and 0.05 M MgCl2, respectively; (b) incubation in blocking solution and that in antibody solution were performed for 1 hr each. After washing in 10 mM Tris-HCl (pH 8.0) with 1 mM EDTA, the sections were coverslipped with glycerol gelatin (Sigma Diagnostics; St Louis, MO), with or without methyl green counterstain.
Results
Western Analysis and Radioimmunoassay
CA was isolated from lacrimal glands by affinity and ion-exchange chromatography, as for the parotid enzyme. However, the amount of enzyme recovered was very much smaller than that from the parotid. Fresh sulfonamide affinity resin was used for isolation of the lacrimal gland enzyme, and the Mono Q column was cleaned with 0.5 M NaOH and 1 M acetic acid before use to prevent contamination of the lacrimal gland preparation with the much more abundant salivary gland enzyme. Blank runs confirmed the absence of any contaminating salivary CA VI. Purified enzyme was run on a gel, blotted, and probed with the anti-CA VI antibody. The lacrimal gland CA had an apparent molecular weight of about 45 kD, the same as the salivary enzyme (Figure 1). The CA isolated from lacrimal glands appears to be the same as the salivary gland enzyme.
To measure the amount of CA VI present in the lacrimal gland, a specific RIA for sheep CA VI was used. Lacrimal glands from 10 sheep were homogenized separately and centrifuged, and the CA VI concentration was measured by this RIA. The protein concentrations of the supernatants were measured and the amount of enzyme/mg total protein calculated. The individual values along with age of the sheep are shown in Table 1. The CA VI content of the lacrimal gland was 0.35 ± 0.22 μg/g tissue (mean ± SD) and the concentration was 4.20 ± 2.60 ng/mg protein with a range of 1.46–9.12 ng/mg. The content and concentration for the parotid gland from sheep #10 were 1.55 mg/g tissue and 29.9 μg/mg protein. There is about 7000 times the concentration of CA VI in the parotid tissue as there is in the lacrimal tissue.
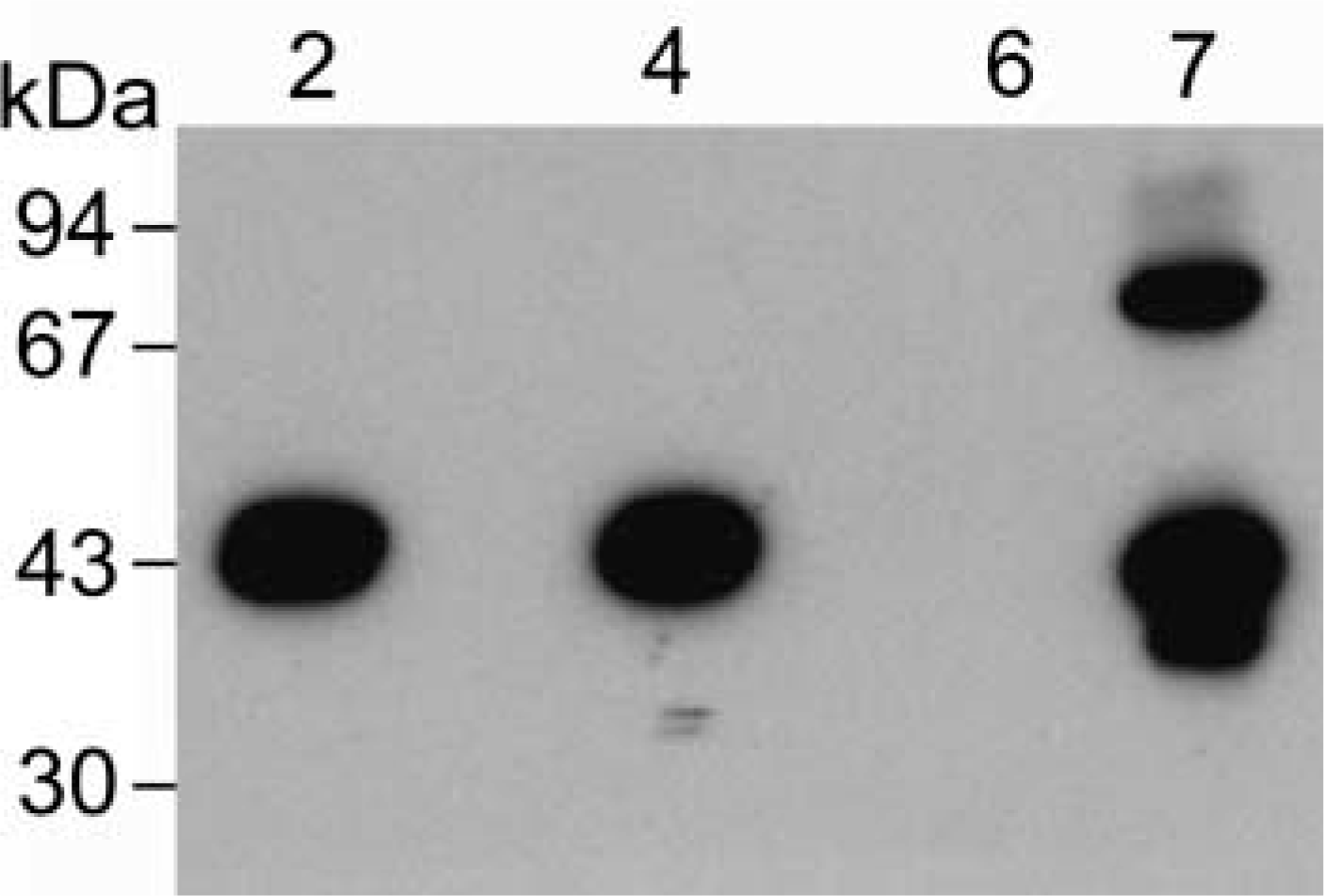
Western analysis of sheep lacrimal gland CA VI. Lane 2, purified sheep salivary CA VI (0.13 μg); Lane 4, sheep parotid homogenate (10 μl of a 1:100 dilution); Lane 6, sheep liver homogenate (10 μl); Lane 7, sheep lacrimal CA VI (2 μg). Molecular weights are indicated at left. The higher molecular weight band in the lacrimal CA VI lane may be a CAVI dimer.
CA VI content of sheep lacrimal glands
RT-PCR Analysis
RT-PCR was performed using 5′-primer and 3′-primer corresponding to positions 446–477 and positions 739–766, respectively, of the nucleotide sequence of bovine submandibular CA VI (Jiang et al. 1996). The amino acid sequences deduced from these primers are almost identical with positions 116–125 and positions 213–221 of the amino acid sequence of sheep salivary CA VI (Fernley et al. 1988b). Therefore, the expected size of the RT-PCR product is 321 bp, which is composed of a 261-bp sheep CA VI cDNA fragment and a total of 60 bp bovine primers. RT-PCR products of both the parotid gland and the lacrimal gland yielded a single band with the theoretically estimated size (Figure 2). These were purified and sequenced. The nucleotide sequences from the two products were identical to each other and the deduced amino acid sequences contained the amino acid sequence (positions 126–212) of sheep salivary CA VI (Figure 3).
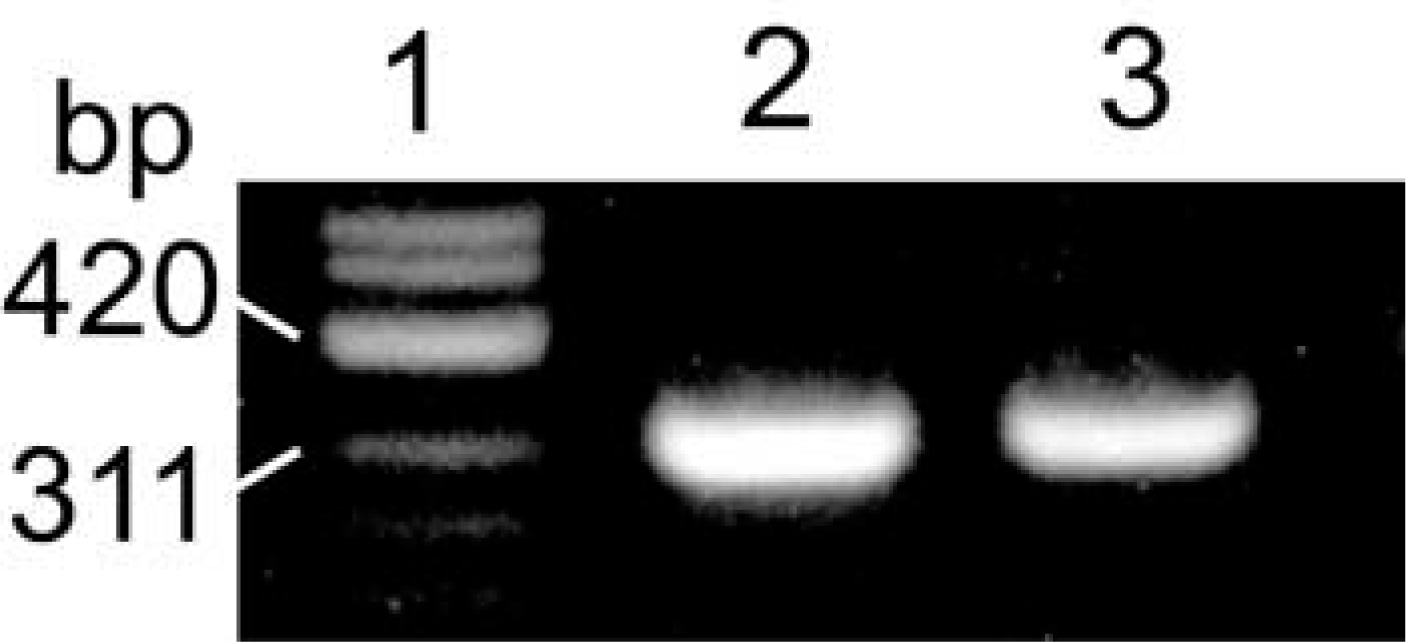
RT-PCR analysis of CA VI mRNA in sheep lacrimal gland. Lane 1, molecular weight marker (ϕX174/Hinf II); Lane 2, PCR product of sheep parotid gland as a positive control; Lane 3, PCR product of sheep lacrimal gland.
A cDNA probe was obtained from the plasmid containing the 261-bp fragment of sheep CA VI and was used for Northern analysis. This probe detected CA VI mRNA in the parotid gland but not the lacrimal gland (data not shown).
Immunohistochemistry and In Situ Hybridization
The polyclonal antibody specific for highly purified sheep secreted CA was found to label only occasional acinar cells in occasional lobules in the sheep lacrimal gland (cf. Figure 4D). Therefore, the number of cells containing the enzyme was far less than in the parotid gland, in which all acinar cells appeared to contain CA VI (Figure 4B). The immunostaining pattern was basically similar between the indirect immunoperoxidase technique and the avidin-biotin-peroxidase technique. Although the latter technique slightly enhanced the staining intensity and increased the number of stained cells, these were still a minority of the secretory cells. The avidin-biotin-peroxidase technique sometimes generated nonspecific staining such as those in the nuclei (Figure 4A, inset). Lacrimal glands are of the compound serous tubuloalveolar type. The secretory cells, which we refer to as “acinar cells” in this manuscript, can be divided into alveolar secretory cells and tubular secretory cells (Kühnel 1968; Orzalesi et al. 1971). Both cell types appeared to contain CA VI (Figure 4A). Cellular distribution patterns of CA VI mRNA in the lacrimal (Figure 4D) and parotid (Figure 4E) glands were similar to those of CA VI-like immunoreactivity.
In the salivary gland, the acinar immunostaining often took a granular configuration and the duct lumen sometimes contained positive staining (arrow in Figure 4B), indicating that the enzyme was stored in the secretory granules in the acinar cells and secreted into the duct lumen from these cells. In contrast, the intensity of the immunostaining, even by the avidin-biotin-peroxidase technique, was extremely weak in the lacrimal gland (compare Figures 4A and 4B). The weak immunostaining made it difficult to judge whether the staining had a granular configuration (Figure 4A). However, the duct luminal content was occasionally labeled by the avidin-biotin-peroxidase technique (Figure 4A, inset), indicating that the enzyme was secreted into the duct lumen. Such labeling was much less common in the lacrimal gland than in the salivary gland, and was not detected by the indirect immunoperoxidase technique.
The above specific staining was not seen when sections were reacted with normal rabbit IgG (Figure 4C), primary antibody pre-absorbed with CA VI, or a DIG-labeled sense probe (Figure 4F).
Discussion
In this study, Western and RT-PCR analyses have confirmed that sheep lacrimal gland synthesizes CA VI and that this enzyme appears to be identical to the salivary enzyme. An RIA specific for sheep CA VI has demonstrated that the concentration of CA VI in the lacrimal gland is very much less than that in the salivary gland. This would explain why the relatively insensitive Northern analysis of lacrimal gland mRNA failed to detect CA VI mRNA. From the IHC and ISH results, it appears that the amount of CA VI in the lacrimal gland, as compared to the parotid gland, is due to the fact that only a small minority of the lacrimal acinar cells synthesizes CA VI. A similar result was observed in rat lacrimal glands by IHC using a monoclonal antibody specific for rat CA VI (Ogawa et al. 1995). The secretion in the lacrimal duct was stained by the avidin-biotin-peroxidase technique but not by the indirect immunoperoxidase technique, indicating that, once secreted, CA VI was rapidly diluted to a level at which the less sensitive latter technique could not detect it.
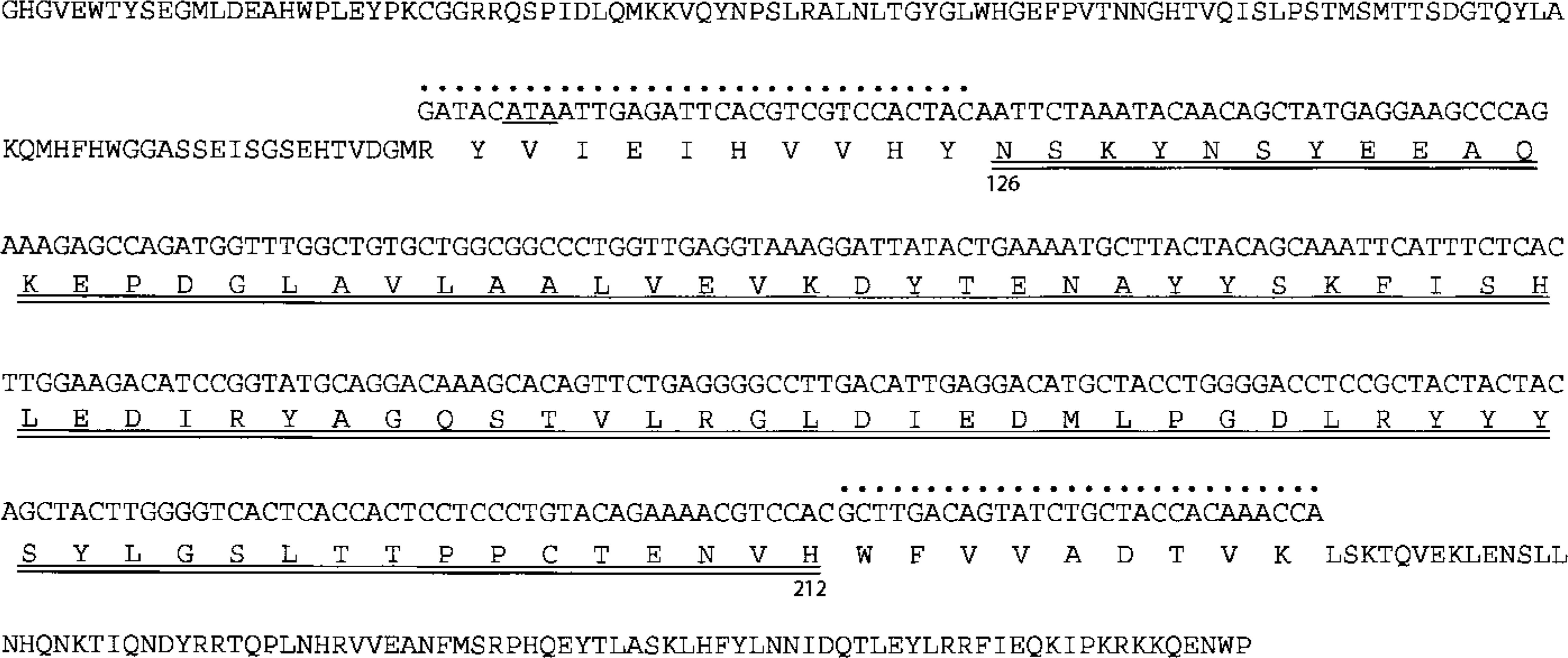
Nucleotide sequence of CA VI cDNA of sheep parotid and lacrimal glands with complete amino acid sequence of sheep salivary CA VI (Fernley et al. 1988b). The nucleotide sequence corresponds to positions 126–212 of the amino acid sequence (double underlined). The primer sequences are also shown (pointed). Because the primers were designed according to the bovine nucleotide sequence, an amino acid in the 5′-primer (the codon underlined) is isoleucine instead of valine in sheep CA VI.
The role of CA VI in tear fluid is presumably akin to its proposed role in saliva, i.e., regulation of pH through the reversible hydration of CO2. On the exposed surface of ocular globe, the concentration of CA VI increases by evaporation of water from tear fluid into the surrounding air between consecutive blinks (Tiffany 1985). Furthermore, tear flow rate is extremely low, about 1–4 μd/min (Schuller et al. 1996) compared to about 2 ml/min for parotid gland. Therefore, the actual concentration of CA VI in the fluid covering the corneal surface could be high enough to perform the physiological role. However, we did not measure the concentration on the corneal surface and it is not known if it is sufficient for this purpose. The acid/base balance of the fluid on the corneal surface may be maintained not only by CA VI but also by other CA(s). These may play a pivotal role, with the most plausible candidates being CA I, CA II, and possibly CA IV in the corneal epithelium. They are en-gaged in the transepithelial flux of CO2 and HCO3 - (Conroy et al. 1992; Candia 1996). Although the physiological significance is unknown, this flux inevitably affects acid/base status of corneal surface. Conroy et al. (1992) have measured the activity of CA I and CA II in the corneal epithelia of humans and several animal species. The highest level is found in the rabbit, followed by human, dog, sheep, and cat. Enzyme histochemistry has shown that rabbit tears do not express CA activity (Bromberg et al. 1993, 1994). Therefore, it is fair to conclude that, in rabbit, acid/base balance on the corneal surface is controlled only by the epithelial CAs. These enzymes in sheep may not have enough activity for pH regulation and, consequently, require some extraneous CA as a supplement. In this context, it is of interest to measure the amount of CA VI in the lacrimal glands of human, dog, and cat.
Previously, a range of tissues, including the pancreas, was examined for the presence of CA VI by Western analysis (Fernley et al. 1989) and by RIA (Fernley et al. 1991), but these methods failed to detect CA VI in these tissues. Recently, Fujikawa-Adachi et al. (1999a), using RT-PCR and RT-PCR Southern blotting analyses, have detected CA VI in the pancreas. The pancreas contains serous type cells that resemble those in the salivary glands, and it is not unexpected that they contain CA VI mRNA. More surprisingly, the protein has been detected in large amounts in human and rat milk (Karhumaa et al. 2001) and is presumably secreted from the mammary gland, which is not categorized as a serous-type gland. Therefore, it is possible that other cell types secrete CA VI, and a combination of sensitive molecular technique and protein biochemical techniques will be used to re-screen various exocrine glands.
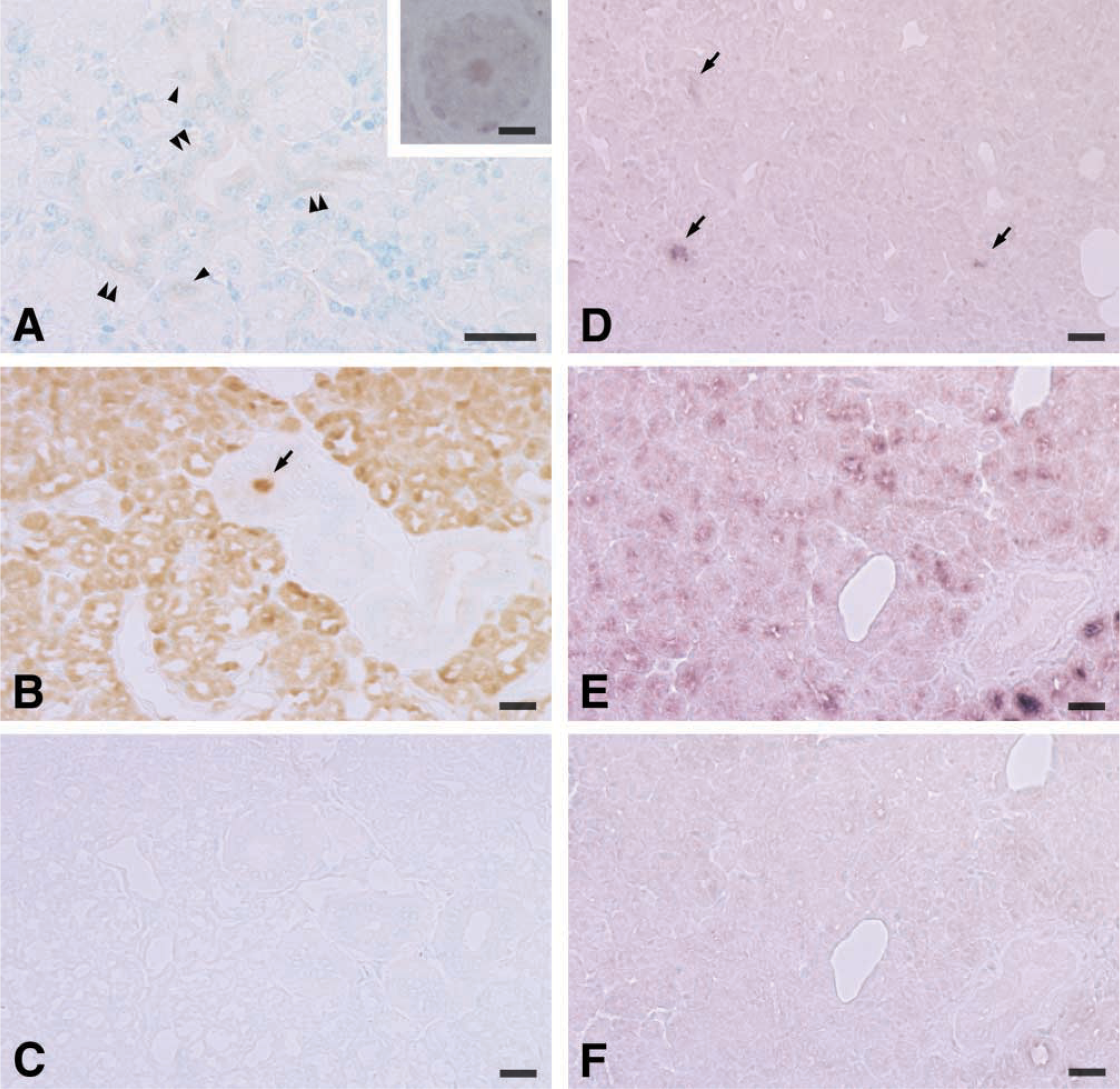
Immunohistochemistry of CA VI in sheep lacrimal (
Footnotes
Acknowledgments
Supported in part by an Institute grant from the National Health and Medical Research Council of Australia.
We thank Drs Haruhiko Takada, Yoshiaki Obara, and Takahiro Yamaguchi for their technical assistance.