
Abstract
Ovarian follicular atresia in all vertebrates is mediated via apoptosis that is initiated in the granulosa cell layer. Here we investigated the relation between connexin expression, cell coupling, and apoptosis in avian granulosa cells. Results from qualitative and quantitative immunocytochemical analysis and Western blotting of connexin43 (Cx43) and electron microscopic observations of gap junctions were compared with functional data on gap junctional coupling obtained by fluorescence recovery after photobleaching in four experimental groups: a control group of freshly isolated granulosa cells, 24-hr serum-free cultures as the apoptosis-inducing condition, and two other groups in which apoptosis was inhibited by either hormone substitution or exposure to elevated extracellular calcium. Our work shows that apoptosis induction in granulosa cells is accompanied by an increased level of cell coupling and that decreasing cell coupling with the gap junction blocker α-glycyrrhetinic acid dose-dependently inhibits apoptosis. The level of Cx43 expression was inversely related to the apoptotic index, suggesting that Cx43 expression plays a role in granulosa cell survival. Our study supports the hypothesis that gap junctional coupling plays a role in propagating a cell death message and suggests a role for Cx43 expression per se in granulosa cell survival.
T
Most studies on apoptosis regulation, including ours, are focusing predominantly on intracellular signaling pathways (D'Herde et al. 2000; Krysko et al. 2001), while tissue function obviously involves cell-to-cell communication. Granulosa cells can communicate either through local production of intraovarian factors such as cytokines (Adashi 1992) and growth factors (Adashi et al. 1991), which act as paracrine and/or autocrine modulators, or through gap junctions. Gap junctions are channels formed by two hemichannels, each formed by six connexin subunits contributed by each cell. All known connexins are transmembrane proteins with four transmembrane domains, two extracellular loops, one intracellular loop, and cytoplasmic amino and carboxyl termini (Bruzzone et al. 1996; Munari-Silem and Rousset 1996; Trosko and Ruch 1998). Cx43 is the major gap junctional protein expressed in granulosa cells that makes a significant contribution to intercellular coupling (Dekel 1987; Grazul-Bilska et al. 1997; Farioli-Vecchioli et al. 2000; Sommersberg et al. 2000; Rosenfeld et al. 2001; Gittens et al. 2003). It has been shown that gap junctions play an important role in granulosa cell development, differentiation, and luteinization (Kidder and Mhawi 2002). Classically, it is believed that gap junctions provide a way for cell-to-cell diffusion of hydrophilic molecules with a molecular mass of less than 1 kD, such as cAMP, Ca2+, IP3, and ATP (Kumar and Gilula 1996). More recently, however, several studies demonstrated that Cx43 could affect signaling pathways independently of its contribution to gap junction channels (Moorby and Patel 2001; Plotkin and Bellido 2001). Despite the increased interest of research in this field, little is known regarding the involvement of gap junctional intercellular communication and Cx43 expression during apoptosis. In a previous study, we observed by electron microscopy (EM) in 24-hr serum-free cultured granulosa cell explants, in which apoptosis is elicited by gonadotropin withdrawal, an increased number of huge gap junctions in comparison with uncultured granulosa cells (D'Herde and Leybaert 1997). This observation prompted us to investigate the relation between connexin expression, cell coupling, apoptosis, and survival of the granulosa cell explants.
Materials and Methods
Isolation and Culture of Granulosa Cell Sheets
Granulosa cell sheets were prepared from ovarian follicles of regularly laying adult Japanese quails (Coturnix coturnix japonica). The animals were reared under an artificial light cycle of 18 hr per day, with food (fresh lettuce and complete breeding food; Biofor AVEVE, Belgium) and water ad libitum. Animal care procedures were conducted in accordance with the guidelines set by the local ethical committee ECP 00/05 (2000). The monolayered granulosa epithelium of the largest preovulatory follicle (F1) was isolated in Krebs-Ringer solution as previously described (Gilbert et al. 1977). This method provides large sheets of vital granulosa cells sandwiched between their basement and vitelline membranes (Mussche et al. 2000; Krysko et al. 2001). The granulosa explants are cultured in suspension in phenol red-free M199 medium (Sigma; Bornem, Belgium) supplemented with 0.1% w/v bovine serum albumin (BSA) fraction V (Sigma), 6.0 g/liter HEPES (Acros; Geel, Belgium), 1% v/v penicillin-streptomycin (GIBCO; Paisley, UK) at pH 7.4. To inhibit the apoptotic process, the culture medium of controls was supplemented either with a combination of luteinizing hormone (LH, 100 ng/ml; Sigma-Aldrich, Bornem, Belgium) and insulin-like growth factor-I (IGF-I, 10 ng/ml; Sigma-Aldrich; (Onagbesan and Peddie 1988; Krysko et al. 2001) or with a continuous load of increased extracellular Ca2+, a protocol that promotes survival through activation of a calcium sensing receptor (Mussche et al. 2000; and Mussche et al. unpublished data). The concentration of all vehicle solutions was always below 0.1% and had no effects on the viability of granulosa cell explants (data not shown).
Quantification of Apoptosis by DAPI Staining
Granulosa cell explants were rinsed in 10 mM PBS, pH 7.4, and fixed in 4% buffered formaldehyde. Thereafter, granulosa cell explants were rinsed again in PBS and stained with a 1:1000 solution of DAPI (2',6'-diamidino-2-phenylindole; Sigma-Aldrich) in PBS. Mounted granulosa cell explants were examined in a Leica DM IRB/E inverted microscope and apoptotic cells were identified by their characteristic fragmented and condensed chromatin masses. Small groups of apoptotic bodies were counted as remnants of one apoptotic cell. Apoptotic indexes (AIs) are expressed as the number of apoptotic nuclei per number of total nuclei counted in the same microscopic field. This AI was averaged for 10 fields, giving a total number of ≃1500 cells counted per experiment.
Antibodies
A mouse monoclonal antibody (IgG1) raised against the peptide sequence corresponding to the amino acid residues 252–270 of the native rat Cx43 was used for immunocytochemistry (ICC) and Western blotting and was purchased from ProBio (Poole, UK) and BD Biosciences (Erembodegem, Belgium), respectively. These monoclonal antibodies were reported to crossreact with avian Cx43 (Beyer 1990; Musil et al. 1990). Affinity-purified secondary goat anti-mouse (IgG) antibody conjugated to horseradish peroxidase (HRP) were obtained from Cell Signaling Technology (Westburg, Leusden, The Netherlands) and secondary rabbit-FITC anti-mouse (IgG) antibody was purchased from DAKO (Glostrup, Denmark).
Immunocytochemistry of Connexin43
After rinsing of granulosa cell explants with PBS, they were fixed for 20 min in 3% PBS-buffered paraformaldehyde. Next, explants were incubated for 20 min in 3% BSA and then for 10 min in 1% glycine. After 45-min incubation in 10% normal rabbit serum, the explants were incubated, directly and without a rinsing step, overnight at 4C with mouse monoclonal anti-Cx43 antibody (1:400 dilution of 1 mg/ml stock; or for the controls the corresponding IgG1 at the same concentration; DAKO). Then the granulosa explants were incubated for 45 min with rabbit-FITC anti-mouse IgG (1:40). For double staining, the granulosa explants were stained with DAPI as described above, rinsed with 0.9% NaCl, and mounted with fluorescent mounting medium. Unless otherwise described, explants were rinsed thoroughly with PBS between all steps.
Quantification of Cx43 Immunofluorescence
Quantitative measurements of Cx43 immunofluorescence (Laing et al. 1997) were performed by determining the average fluorescence intensity in images obtained by confocal laser scanning microscopy (CLSM) of granulosa cell sheets or explants immunostained as described above. Images were acquired with a custom-made video-rate CLSM (Sanderson and Parker 2003) built around a Nikon Eclipse TE300 (Analis; Ghent, Belgium) equipped with a ×40 oil immersion objective (CFI Plan Fluor; Nikon) and with 488-nm excitation from an argon laser.
Protein Extraction and Western Blotting Analysis of Cx43 Expression
Granulosa cell explants were lysed in 50 μl of ice-cold modified RIPA buffer (50 mM NaCl, 0.5% NP40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate, 250 mM α-glycerophosphate, 25 mM Tris, pH 8.2) containing a standard phosphatase inhibitor cocktail 2 (20 μl/ml; Sigma-Aldrich) and complete EDTA-free protease inhibitor tablet (20 μl/ml; Roche Diagnostics, Brussels, Belgium) for 10 min and sonicated on ice three times for 10 sec using a Vibracell VC-130 sonicator. The protein concentration of each sample was measured according to Bradford (1976) using the BioRad protein assay kit (BioRad Laboratories; Nazareth/Eke, Belgium) and a microtiter plate reader model 550 (Bio-Rad Laboratories). Protein samples were solubilized in Laemlli sample buffer and were heated at 100C for 10 min and centrifuged at 13,000 × g for 7 min to remove cell debris. Rat brain lysate was obtained from BD Biosciences (Erembodegem, Belgium) and was used as a positive control for Cx43 protein expression on Western blots. Approximately 55 μg of total protein from each sample was separated on 12% Tris-HCl precast gels (BioRad Laboratories). After separation, proteins were transferred to a nitrocellulose membrane (VWR; Zaventem, Belgium), blocked for 1 hr in TBS containing 5% non-fat dried milk and 0.1% Tween (blocking buffer), and incubated overnight at 4C with monoclonal antibodies to Cx43 diluted 1:250 in blocking buffer. After extensive washes in blocking buffer, membranes were probed for 1 hr with HRP-conjugated secondary goat anti-mouse IgG antibody diluted in blocking buffer (1:2000) and developed by enhanced chemiluminescence (Cell Signaling Technology; Westburg) according to the manufacturer's instructions. The time of exposure (typically 10 min) to X-ray films (Kodak Biomax Light-1; Sigma-Aldrich) was identical for all experimental conditions. The intensity of the bands after Western blotting was determined by scanning of the X-ray films followed by quantitative analysis using Total-lab software (Phoretix).
Transmission Electron Microscopy
The granulosa explants were fixed by immersion for 2 hr in 2% glutaraldehyde buffered with 0.1 M Na-cacodylate containing 1 mM CaCl2 (pH 7.4). After a rinse in 0.1 M Nacacodylate containing 7.5% sucrose, the explants were osmicated in 2% OsO4 in 0.1 M Na-cacodylate and embedded in LX medium (Ladd; Burlington, VT). Ultrathin sections were stained with uranyl acetate and lead citrate and were analyzed on a Jeol 1200 EXII electron microscope.
Fluorescence Recovery After Photobleaching (FRAP)
Gap junctional coupling was investigated in granulosa cell sheets or explants using the FRAP technique (Braet et al. 2003). For this purpose, the sheets or explants were loaded with the gap junction-permeable fluorescent tracer 5-carboxy-fluorescein diacetate (CFDA; Molecular Probes, Leiden, The Netherlands) by incubation in Hank's buffered salt solution (HBSS-HEPES) containing 50 μM CFDA for 60 min at room temperature and under continuous shaking. The preparations were then rinsed three times with HBSS-HEPES and transferred to the microscope stage of the CLSM described above. The loading and rinsing solutions contained in addition 1 mM probenecid (Sigma) to prevent rapid dye extrusion from the cells (Di Virgilio et al. 1988; D'Herde and Leybaert 1997). Fluorescence within a single cell was photobleached by spot exposure to the 488-nm line of an argon laser over a period of 10 sec. Spot exposure was done by reducing the scan amplitude of the horizontal and vertical scanning mirrors. CFDA fluorescence before and after photobleaching was recorded in confocal imaging mode with normal scanning mirror settings. Recovery of fluorescence because of dye influx from neighboring cells was quantified at 4 min after the start of photobleaching. A correction was made for the baseline bleaching effect associated with repeated exposure to the 488-nm imaging light. This was done by recording the fluorescence profile in cells not exposed to the spot bleach and correcting the recovery signal in proportion to the baseline fluorescence decay.
Gap Junction Blocking by 18 α-Glycyrrhetinic Acid
Eighteen α-glycyrrhetinic acid (3 α -hydroxy-11-oxo-18 α, 20 α -olean-12-en-29-oic acid, AGA; Sigma) was dissolved in dimethylsulfoxide (DMSO; UCB, Leuven, Belgium). To study the effect of gap junction blocking on cell survival, granulosa cell explants were incubated for 24 hr with AGA doses ranging from 25 μM to 250 μM and AIs were calculated on DAPI- stained granulosa cell explants as described above. Cell coupling after treatment with AGA (250 μM, 24 hr) was investigated using FRAP.
Statistical Analysis
The data are expressed as mean ± SEM, with n denoting the number of experiments. Statistical significance was tested using a t-test for unpaired observations and using a p value of less than 0.05. Multiple groups were compared using variance analysis, followed by the Dunnett test for multiple comparisons to a control group or the Student-Newman-Keuls test for comparison of all groups among each other. Curve fittings for dose–response relations were performed with non-linear least-square procedures available in the program Inplot.
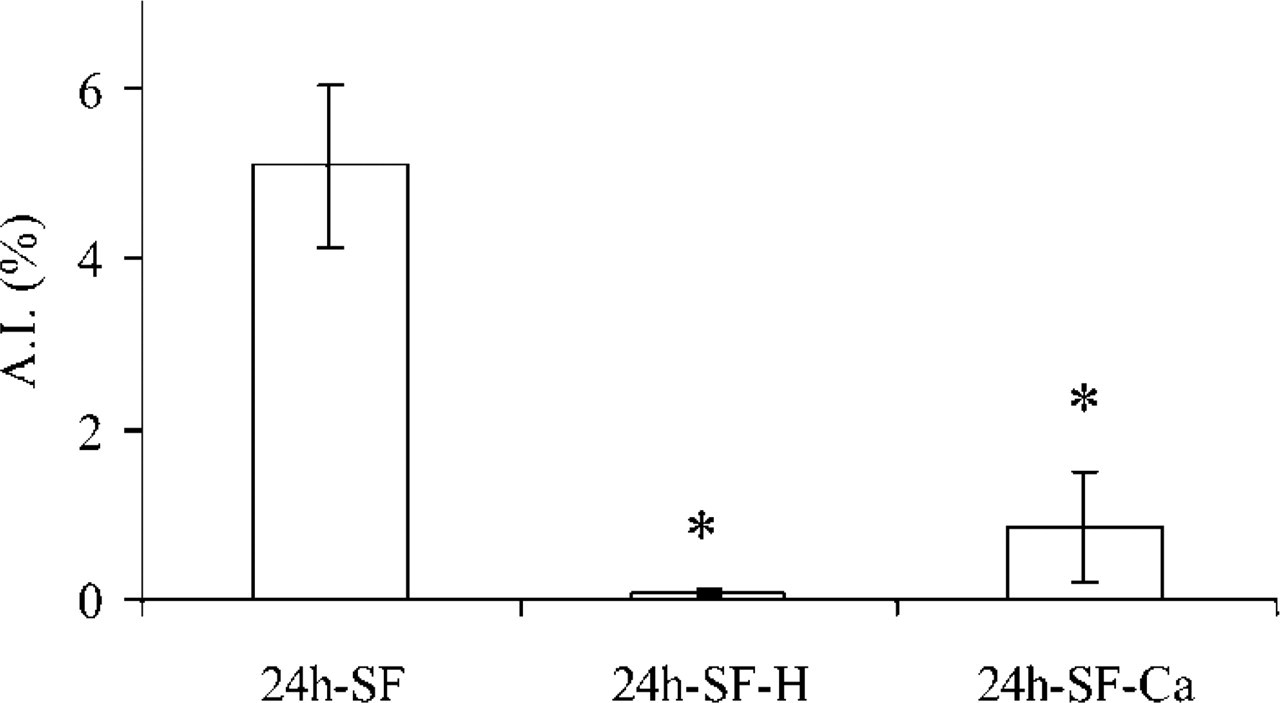
Effect of the culture conditions on the AI in granulosa cell explants. Culturing granulosa cell explants under 24h-SF condition induced apoptosis, while culturing granulosa cell explants for 24h-SF-H or 24h-SF-Ca supplemented conditions inhibited this induced apoptosis. The AI in fresh granulosa cell sheets was negligible and therefore is not shown. ∗p<0.05 compared with 24h-SF; n = 60 in six different animals.
Results
Apoptosis and Cx43 ICC
Freshly isolated preparations were used for control purposes, 24-hr serum-free (24h-SF) cultures as the apoptosis-inducing condition (D'Herde and Leybaert 1997), and 24h-SF cultures supplemented either with hormones (IGF-I and LH; 24h-SF-H) (Onagbesan and Peddie 1988; Krysko et al. 2001) or with an increased concentration of extracellular calcium (24h-SF-Ca) (Mussche et al. 2000; and Mussche et al. unpublished data) as the apoptosis-inhibiting conditions. The AIs observed under these experimental conditions are illustrated in Figure 1.
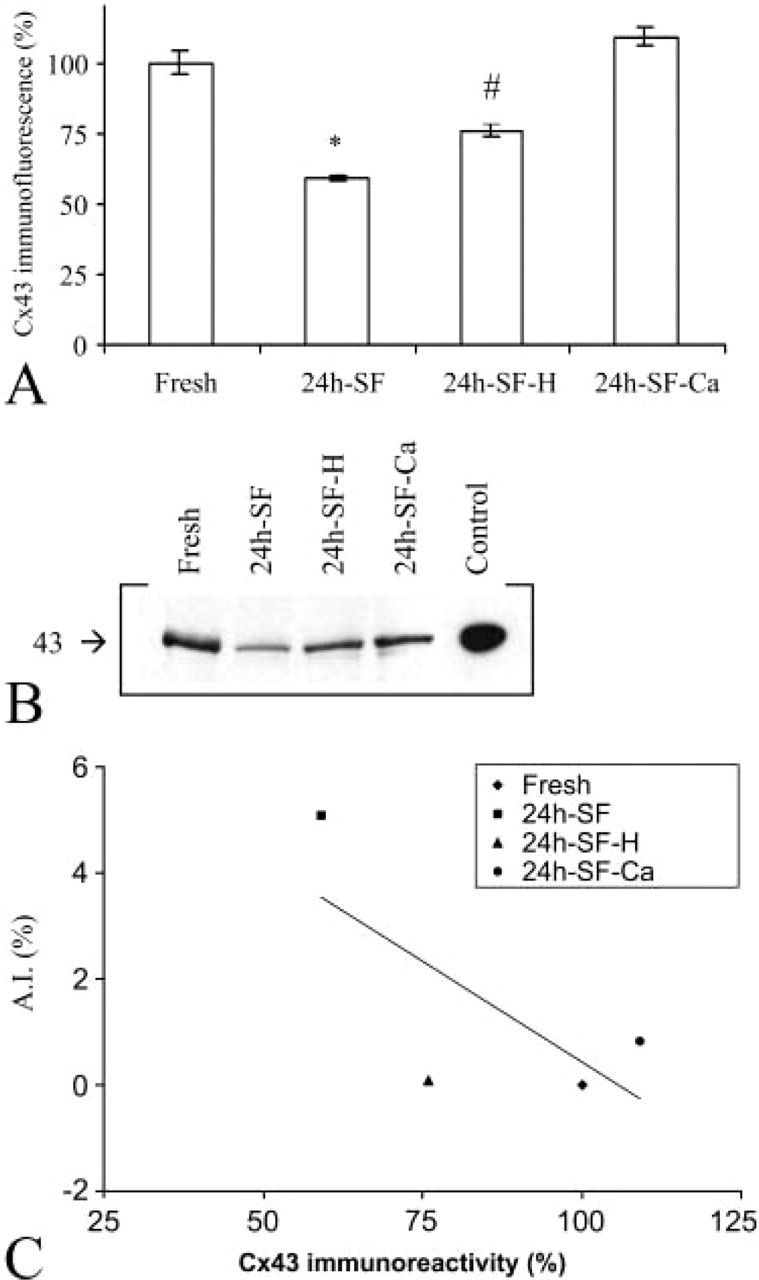
Staining of freshly isolated granulosa cell sheets with anti-Cx43 antibodies demonstrated immunoreactivity appearing as a fine and continuous trace, lining the cell borders and involving both places of cell-cell contact as well as non-junctional regions (Figures 2A and 2E). Granulosa cell explants in 24h-SF cultures displayed gap junctional plaques in the form of large dots or lines at cell-cell interfaces (Figures 2B, 2F, and 2I). On average, each cell contained 5.24 ± 0.11 (n=100 cells) of these immunopositive dots or lines. Apoptotic cells, easily recognized by their nuclear morphology by DAPI staining, did not show Cx43 immunolabeling at their plasma membrane (Figure 2I). However, occasionally apoptotic bodies demonstrated very weak dot-like cytoplasmic staining (Figure 2I). Interestingly, DAPI staining revealed apoptotic granulosa cells that were lying in clusters or streaks (Figure 2J). Granulosa cell explants in 24h-SF-H culture displayed a staining pattern that was regular and more intense than in 24h-SF culture condition. The gap junctions appeared to be huge as well (Figures 2C and 2G) with on average 6.30 ± 0.14 (n=100 cells) of these gap junctional immunopositivities per cell (significantly higher compared with 24h-SF granulosa cell explants). In granulosa cell explants cultured under 24h-SF-Ca conditions, the immunostaining appeared as a dotted pattern of many short gap junctions surrounding each cell (Figures 2D and 2H). Quantification of the level of Cx43 immunoreactivity, determined from the average intensity of confocal images acquired in the four conditions, demonstrated that the immunoreactivity in 24h-SF granulosa cell explants was significantly below the level observed in freshly isolated granulosa cells. The Cx43 immunoreactivity was significantly higher under the 24h-SF-H and 24h-SF-Ca, i.e., apoptosis-inhibiting conditions, compared with 24h-SF (Figure 3A).
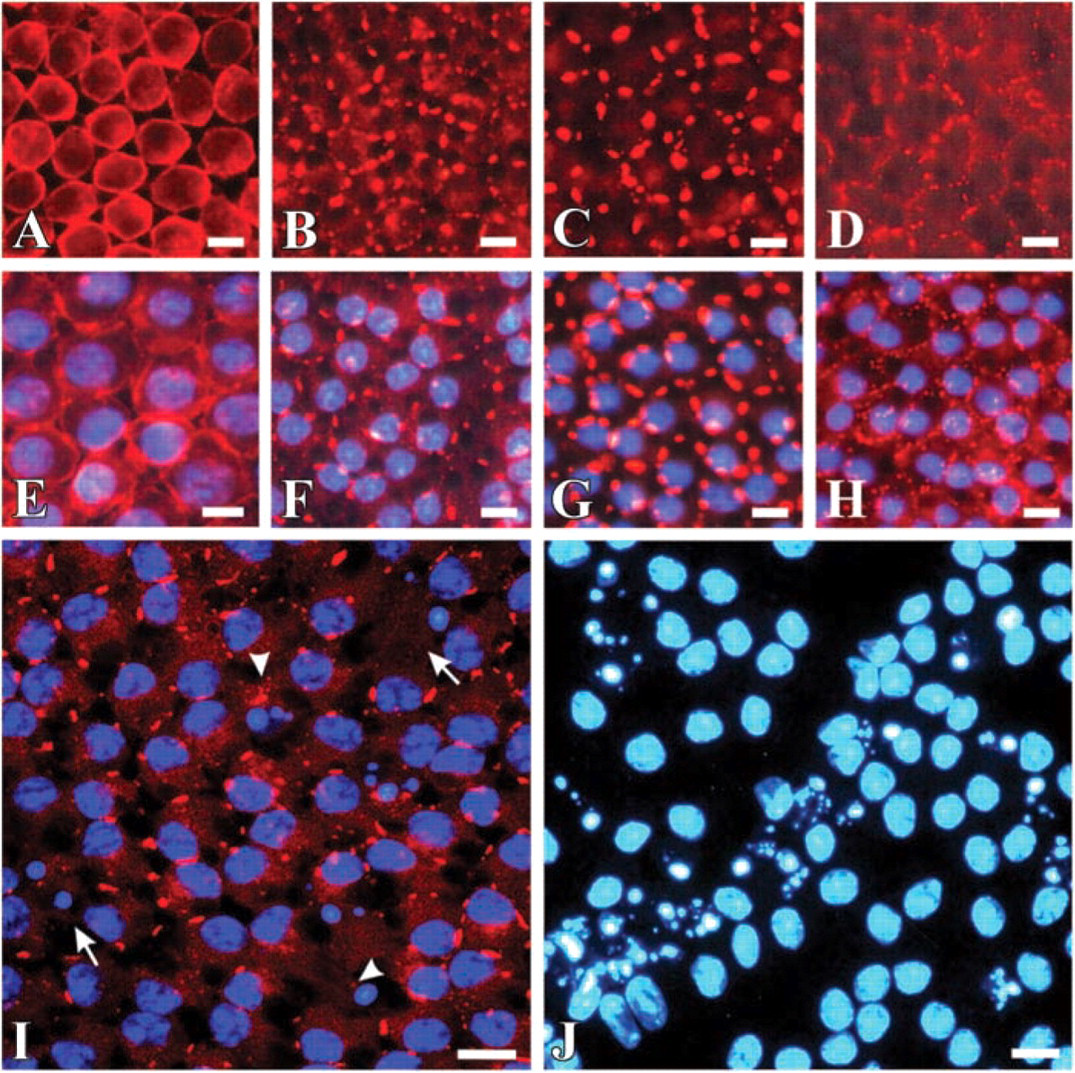
Epifluorescence micrographs of granulosa cells. (
(
Level of Cx43 Expression as Studied by Western Blotting
Granulosa cell explants, both cultured and uncultured, express a 43-kD protein that reacts with Cx43-specific antibodies on Western blots. Immunoblots of whole cell lysates showed that 24h-SF granulosa cells explants contained only 39% of what is expressed in freshly isolated granulosa cells sheets, while the level of Cx43 expression recovered to 74% and 76% under the 24h-SF-H and 24h-SF-Ca conditions, respectively (Figure 3B), which is in accordance with the data obtained by ICC analysis. Combination of the quantitative data from ICC analysis and Western blotting showed that Cx43 expression was inversely related to the AIs (Figure 3C).
Electron Microscopy
Electron microscopic analysis of granulosa cells demonstrated rare gap junctions in freshly isolated granulosa cells despite the positive Cx43 immunoreactivity described above, and showed huge gap junctions in 24h-SF and 24h-SF-H conditions (Figure 4). In 24h-SF-Ca conditions, many short gap junctions were observed in accordance with the ICC punctate pattern (Figure 4). Internalization of gap junctions did not occur, as evidenced by the absence of annular junctions.
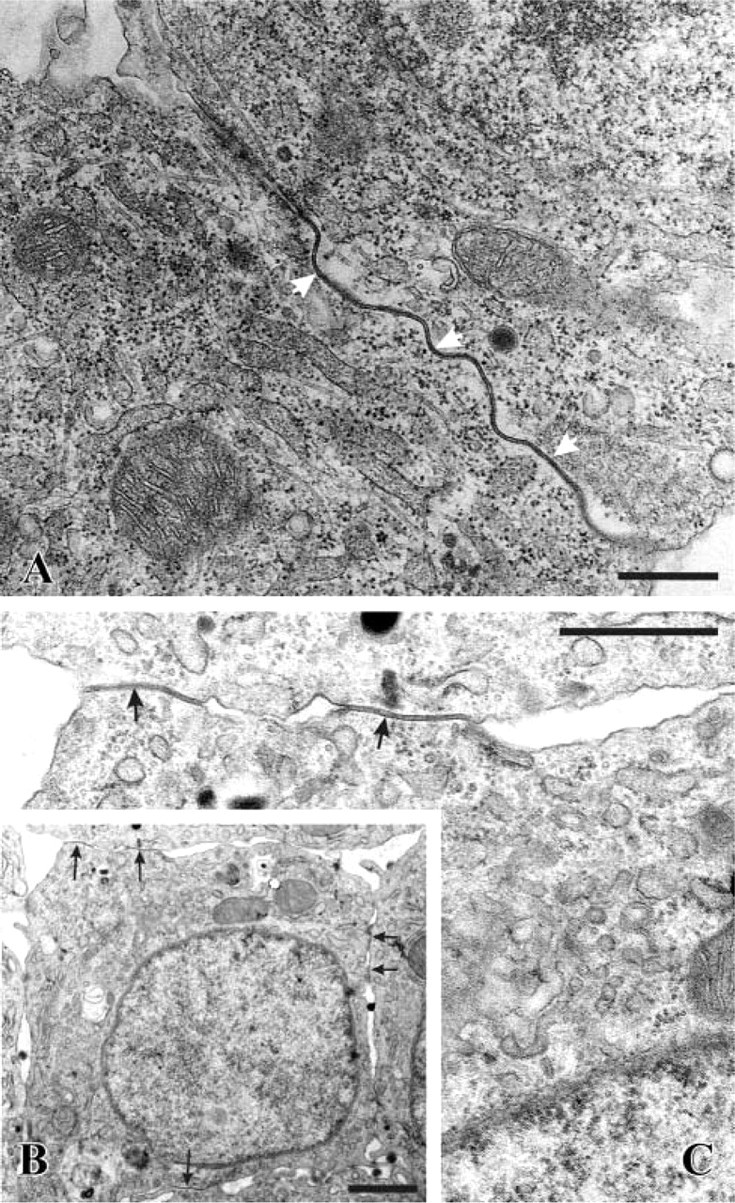
Electron microscopic micrographs. (
Functional Coupling as Studied by FRAP
The level of gap junctional coupling in granulosa cell explants cultured in 24h-SF conditions was significantly higher compared with the freshly isolated granulosa cell sheets (Figure 5). In 24h-SF-H treated granulosa cell explants, the level of gap junctional coupling was comparable to the freshly isolated conditions, while granulosa cell explants kept under 24h-SF-Ca displayed the highest level of coupling (Figure 5).
Effect of Gap Junction Blockade Experiments on Apoptotic Indexes
To investigate the effect of gap junction blocking on cell survival, granulosa cell explants were incubated for 24 hr in serum-free condition with increasing doses of AGA (25 μM to 250 μM range) and AIs were evaluated by DAPI staining. These results showed that the AI was dose-dependently decreased by AGA, with a half-maximal effect occurring at ≈40 μM (Figure 6). FRAP experiments showed that 24 hr exposure of the granulosa cell explants to AGA caused a drastic reduction of gap junctional coupling, with 250 μM AGA decreasing the fluorescence recovery from a control level of 45.8 ± 2.6% to 3.2 ± 0.6% (n=8 in one animal; p<0.00001).
Discussion
The purpose of the present study was to investigate the role of cell coupling and level of connexin43 expression in apoptosis and survival of granulosa cell ex-plants. We demonstrated that apoptosis induction in granulosa cell explants is accompanied by an increased level of gap junctional coupling, that apoptosis was dose-dependently inhibited by the gap junction blocker AGA, and that apoptosis induction is accompanied by a decreased level of Cx43 expression, which is regained by the two applied survival protocols.
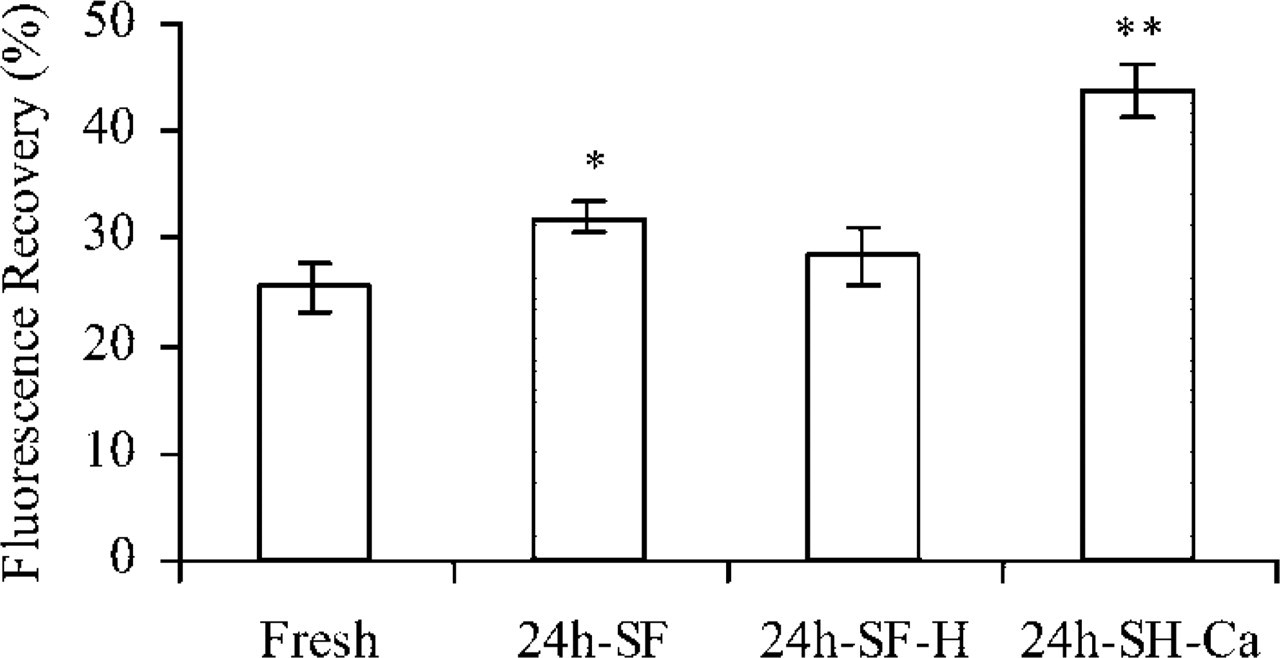
Gap junctional coupling as studied with FRAP. Gap junctional coupling was more prominent (larger extent of fluorescence recovery) in 24h-SF conditions compared with freshly isolated conditions (Fresh). Under 24h-SF-H conditions, gap junctional coupling was no different from the freshly isolated condition, whereas the 24h-SF-Ca condition was associated with a very large coupling degree. ∗p<0.05, ∗∗p<0.01 compared with the fresh condition; n = 25 in at least three different animals.
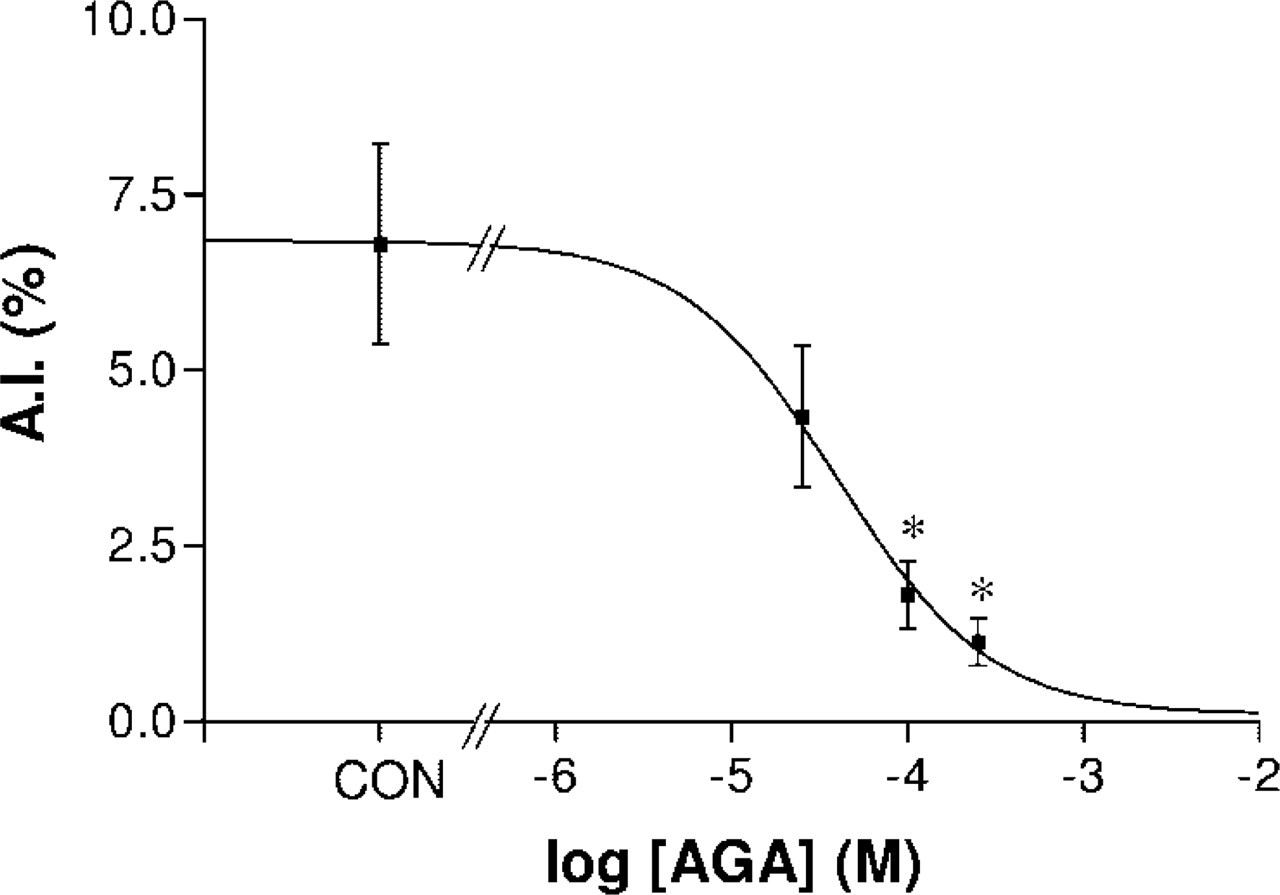
Dose-response relation between inhibition of gap junctional coupling with increasing doses of AGA and the resulting effect on AI. AGA dose-dependently inhibited apoptosis, with half-maximal inhibition occurring with concentrations in the order of 40 μM. ∗p<0.05 compared with control (con).
Granulosa cell explants provide a model system closely mimicking the apoptotic process that occurs in vivo during spontaneous follicular atresia (D'Herde and Leybaert 1997). Contributing factors to explain the relatively low apoptotic rates in 24h-SF granulosa cell explants might be the presence of native extracellular matrix, i.e., the vitelline and basement membranes, which are still present in the used model system and which are known to promote survival (Amsterdam et al. 1989; Boudreau et al. 1995). Second, as reviewed by Johnson (2000), death-suppressing members of the Bcl-2 family (Bcl-xLong, Bcl-2) are expressed in granulosa cells from preovulatory follicles. Another contributing factor is the expression of IAP-1, an inhibitor of apoptosis protein found in preovulatory hen granulosa cells (Johnson et al. 1998).
The data of the present study demonstrated that, in freshly isolated granulosa cells, Cx43 immunolabeling was observed all around the cells, even in those regions of the plasma membrane where visible contact with any apposing plasma membrane of neighboring cells was missing. In the absence of any apposing cells, immunolabeling in the non-junctional plasma membrane regions is assumed to represent hemichannels of Cx43 (Musil and Goodenough 1991; Zampighi et al. 1999; Quist et al. 2000). Quantification of the Cx43 immunoreactivity (as revealed by ICC and Western blotting) in freshly isolated granulosa cell sheets showed a higher amount of Cx43 protein compared with 24h-SF or 24h-SF-H supplemented cultures in which at EM level long gap junctions between neighboring cells are easily detected. Despite the detected Cx43 immunolabeling and coupling of the cells as assessed by FRAP, extremely rare gap junctions were detected by EM analysis in these freshly isolated granulosa cells. This discrepancy between ultrastructural detection of gap junctions and ICC detection of their major component Cx43 might be explained by the fact that immunoreactivity is not confined only to gap junctions but also reveals Cx43 present in the cytoplasm or organized as connexin hemichannels. Moreover, one should consider the fact that clustering of gap junctions into plaques is the primary basis for their ultrastructural identification (Goodenough and Revel 1970; McNutt and Weinstein 1970).
In 24h-SF cultured granulosa cell explants, the apoptosis-inducing condition, well-organized gap junctions were observed (as revealed by ICC and EM), providing a better coupling of the granulosa cells than in freshly isolated granulosa cells as measured by FRAP analysis. However, a lower immunoreactive signal was detected in these 24h-SF granulosa cells (as revealed by quantitative analysis of both the confocal images and immunoblots) compared with freshly isolated granulosa cells. These apparently inconsistent findings underscore the fact that the abundance of Cx43 does not necessarily correlate with the amount of intracellular communication (Lau et al. 1992; Kurata and Lau 1994; Laing et al. 1997; Sia et al. 1999).
In agreement with the data from the present study, Wilson et al. (2000) found an average increase of gap junctional coupling during initiation of apoptosis in serum-deprived (24-hr) cultures of a normal diploid epithelial cell line (WB-F344), assessed by the scrape load/dye transfer method. In the same study, individual cells were analyzed by the FRAP assay to compare nuclear morphology to differences in dye transfer during serum deprivation. The authors demonstrated that most apoptotic bodies did not communicate with neighboring cells. This is in line with the present ICC findings revealing that granulosa cells with clear apoptotic nuclear morphology do not show Cx43 immunolabeling at their plasma membrane. Kalvelyte et al. (2003) reported that expression of Cx43 in HeLa-transfected cells accelerates the transition of cells from an early phase of apoptosis to the late phase.
To verify whether cell coupling is indeed a means of propagating a cell death message, experiments using the gap junctional intercellular communication inhibitor AGA were carried out. This compound was shown to block cell coupling (Davidson et al. 1986), presumably by disassembling gap junctional channels and affecting their dephosphorylation (Guan et al. 1996). In the present model system, gap junctional coupling was significantly inhibited after long-term AGA exposure. Moreover, it was demonstrated that, by blocking gap junctional coupling with AGA, AIs were dose-dependently decreased (Figure 6). In agreement with the present data, Krutovskikh et al. (2002) showed that in BC31 cells (a rat bladder carcinoma cell line) cell coupling and apoptosis were significantly inhibited by treatment with AGA. In addition, Lin et al. (1998) reported that gap junctions achieved by transfection of Cx43 could mediate the propagation of a death signal between dying and healthy glial cells in a co-culture system.
A further argument in support of a role of cell coupling in the propagation of a death signal among granulosa cells comes from DAPI-stained granulosa explants in the 24h-SF condition (Figure 2J), which demonstrates that granulosa cells did not die randomly but, rather, in tracks of closely associated cells. This finding supports the observation described by Cotrina et al. (1998) and Krutovskikh et al. (2002) that communicating cells die by clusters. Although apoptosis in granulosa cell explants is associated with increased cell coupling (as discussed above), it is important to note that increased cell coupling does not necessarily induce apoptosis per se, as illustrated in the Ca2+-rescue protocol. Therefore, our findings and those of others (Lin et al. 1998; Sai et al. 2001; Krutovskikh et al. 2002; Kalvelyte et al. 2003) are consistent with the hypothesis that gap junctional intercellular communication is indeed needed for cell-cell transmission of a death signal and thus may contribute to the regulation of the apoptotic process in diverse model systems.
It is known that gap junctions are permeable to several second messengers, such as inositol-1,4,5-triphosphate, ATP, cAMP, and calcium.
In a rat bladder carcinoma line, it has been demonstrated that a reduction of gap junction permeability with oleamide, which appears not to affect the passage of Ca2+ ions, did not abrogate coordinated cell death by clusters, suggesting that Ca2+ ions are involved as the signal propagating cell death (Krutovskikh et al. 2002).
Interestingly, Cx43 immunoreactivity (as studied by ICC and immunoblotting) was inversely correlated with apoptosis (Figure 3C), suggesting that Cx43 expression plays a role in the survival process. Sasson and Amsterdam (2002) reported that apoptosis induction by LH and forskolin was accompanied by increased expression of Cx43 in human luteinized granulosa cells, a discrepancy that might be related to the use of different cell models and apoptosis-inducing stimuli. Several reports point to the fact that Cx43 plays a role in promoting survival in response to extracellular cues independently of gap junctional intercellular communication (Plotkin and Bellido 2001; Plotkin et al. 2002). Huang et al. (2001) demonstrated that regulation of apoptosis by Cx43 in human glioblastoma cells is mediated by downregulation of the apoptosis inhibitor protein Bcl-2 and hence is not linked to gap junction channel function. Recently, Lin et al. (2003) demonstrated that forced expression of Cx43 as well as two other members of the connexin family (Cx32 and Cx40) increased the resistance of astrocytes to injury and that the anti-death activity of connexin proteins was independent of the functional status of gap junctions.
In conclusion, the present study suggests that gap junctions mediate the propagation of a cell death message, while independently Cx43 expression is needed for survival. Future studies will focus on the experimental modulation of Cx43 expression and its effects on the major pro- and anti-apoptotic signaling pathways.
Footnotes
Acknowledgements
Supported by Ghent University (GOA grant no. 12050502 to KD) and the Fund for Scientific Research Flanders, Belgium (FWO grant nos. 3G023599, 3G001201, and G.0335.03 to LL).
We are very grateful to Barbara De Bondt, Dominique Jacobus, and Hubert Stevens for excellent technical assistance.