
Abstract
Connexin43 (Cx43) is one of the most predominant gap junction proteins found in the testis. We used in situ hybridization and indirect immunofluorescence to study the distribution of Cx43 mRNA and protein in the rodent seminiferous epithelium. During mouse testis maturation, Cx43 mRNA and its corresponding protein were first detected in the adluminal compartment of the growing seminiferous tubules (early postnatal age: Day 12) to become progressively located in the basal compartment at later ages (Days 16, 19, 27). In seminiferous tubules of sexually mature animals, the intensity of the hybridization signal was stage-dependent, with a maximum at Stage VII compared with Stages V and IX of the spermatogenic cycle (p<0.05). The highest expression of Cx43 mRNA was observed in the supporting Sertoli cells and, to a lesser extent, in the most basally located and less mature germ cells (spermatogonia and spermatocytes). Consistent with these observations, in situ dye coupling was observed between Sertoli cells and basal germ cells. In a mutant mouse deficient for the retinoid X receptor β, which exhibited abnormal spermatogenesis due to altered Sertoli cell function, Cx43 transcripts were markedly decreased in the seminiferous epithelium (p<0.01). The immunoreactive signal for Cx43 was significantly reduced in seminiferous tubules of the 3-month-old mutant mice (p<0.05) and undetectable in older animals. These data provide new information about the precise localization of Cx43 mRNA and protein in seminiferous tubules of immature and mature rodent testes. Moreover, they suggest that retinoids, through the RXRβ receptors, could be involved in the control of Cx43 gene expression in Sertoli cells.
M
It is now established that Cx can be modulated at the transcriptional and translational levels or by alterations in the gating channels (for review see Bruzzone et al. 1996). Recent data have demonstrated that in seminiferous tubules Cx43 protein expression is age- and stage-dependent (Risley et al. 1992; Pelletier 1995; Tan et al. 1996; Lablack et al. 1998) and that the presence of germinal cells is a prerequisite for tubule Cx43 expression (Batias et al. 1999). However, the variations of Cx43 protein expression under various endocrine conditions remain unexplained. Because changes in the amount of Cx gene products have been reported in the ovary and myometrium, but not in the heart, differential regulation appears likely in different tissues (Risek et al. 1990).
Immunoreactive Cx43 has been mainly detected in the basal compartment of the seminiferous tubules (Risley et al. 1992; Tan et al. 1996; Lablack et al. 1998) at the same layers of cells as the tight junction-associated protein ZO-1, which is implicated in the blood-testis barrier (Byers et al. 1991; Bourdon et al. 1998; Batias et al. 1999). These findings confirm previous ultrastructural observations showing a predominant localization in the region of Sertoli-Sertoli occluding junctions (Dym and Fawcett 1970; Gilula et al. 1976). The presence of gap junctions between Sertoli cells and specific germ cells was also reported by freeze-fracture (McGinley et al. 1979; Pelletier and Friend 1983). This was recently confirmed by immunolocalization of the gap junction protein Cx43, the predominant Cx in the testis (Pelletier 1995). The Cx43 mRNA transcripts were shown to be detected in the testis (Kadle et al. 1991; Risley et al. 1992). However, these latter did not study the pattern of temporal mRNA expression, and cell types that express Cx43 mRNA in the seminiferous epithelium were not clearly determined.
In this study, the cellular localization of Cx43 was determined in the developing and functional mature testis at the mRNA and protein level by in situ hybridization (ISH) and indirect immunofluorescence, respectively. In addition, the presence of Cx43 transcripts and Cx43 protein was examined during the normal spermatogenic cycle and in mutant mice deficient for the retinoid X receptor β with abnormal spermatogenesis. These results point to the essential roles of gap junction protein in the maturation of fertile male germ cells.
Materials and Methods
Animals
Male Long-Evans rats and male Swiss mice were housed in a day-night-cycled room, with food and water provided ad libitum. Mice were sacrificed at 4, 8, 12, 16, 19, 27, or 60 days postpartum and rats at 3 months. RXRβ−/− mice were obtained by homologous recombination (Kastner et al. 1996). Testes from 3-and 11-month-old wild-type mice and from RXRβ−/−males were generously provided by Pr. P. Chambon (Institut de Génétique et de Biologie Moléculaire et Cellulaire, Strasbourg, France).
Indirect Immunofluorescence
Testes were frozen in OCT embedding compound (Tissue Tek; Miles, Naperville, IL) and cryosectioned. Testicular sections (5 μm) were applied to 3-amino-propyltriethoxysilane-coated slides and treated as previously reported (Bourdon et al. 1998; Lablack et al. 1998). Briefly, slides were incubated for 2 hr with primary antibody to Cx43 (Transduction Laboratories; Lexington, KY) diluted at 1:200 in PBS containing 3% BSA. After several rinses, slides were then incubated for 1 hr with secondary antibody diluted at 1:500: a rhodamine (TRITC)-conjugated goat anti-mouse immunoglobulins (Jackson Immunoresearch Laboratories; Baltimore, UK) in PBS containing 3% BSA. Controls were performed by omission of the primary antibody or by replacement of the Cx43 antibody by nonimmune IgGs. Stage identification of the seminiferous epithelium was made after incubating slides for 1 min in a solution composed of 2 μg/ml DAPI (Sigma; Saint Quentin Fallavier, France) for nuclear staining. Stages of the seminiferous epithelium were identified according to morphological criteria, including the depth and shape of elongate spermatid nuclei in Sertoli cells, the presence or absence of two generations of spermatids (round and elongate spermatids), and the size of the acrosome surmounting the round spermatid nucleus (Leblond and Clermont 1952; Oakberg 1956). Slides were examinated with a Nikon fluorescence microscope and photographed with T-Max 400 film (Kodak) set at 800 ASA.
In Situ Hybridization
Testes were dissected out and immediately fixed in 4% (w/v) paraformaldehyde in PBS (pH 7.4) for 4 hr before standard paraffin embedding. Tissue sections (5 μm thick) were collected on 3-aminopropyltri-ethoxysilane-coated slides and ISH was performed as previously described (Sibony et al. 1995). The rat Cx43 cDNA (gift from Dr M. Mesnil; IARC, Lyon, France) was subcloned into the EcoR1-restriction site of Bluescript M13 vector. Cx43 antisense and Cx43 sense were linearized with Hind III (Boehringer Mannheim; Meylan, France) and with Bam H1 (Boehringer Mannheim), respectively. The radiolabeled probes were generated by in vitro transcription and incorporation of [35S]-uridine triphosphate (specific ratio activity 1000 Ci/mmol; Amersham, Poole, UK) as previously reported (Sibony et al. 1995). Two serial testicular sections were mounted on the same histological slide. One received the antisense RNA probe and the other received the RNA sense probe as negative control. Heart tissue sections prepared as described above were used as positive control tissue. Tissue sections were covered with 30-100 hybridization buffer [50% formamide, 10% dextran sulfate, 1 mg/ml salmon sperm DNA, 70 mM DTT and 20 × SSC (3 M sodium chloride, 0.3 M sodium citrate)] containing the radiolabeled probe at a concentration of 10-15 × 103 cpm/μl. After the posthybridization washings, the slides were dipped in NTB2 emulsion (Kodak; Rochester, NY) and exposed at 4C in light-tight boxes for 15 days. Slides were developed, fixed, and counterstained with Toluidine blue. At least three testicular sections from three different testes were processed. Slides were examined by two observers and areas to be photographed were selected as representative. Photographs were taken on a Nikon microscope with Ilford PANF 50 ASA film. The stages of the cyle of the seminiferous epithelium were identified according to the morphological criteria for rat (Leblond and Clermont 1952) and mouse (Oakberg 1956) spermatogenesis.
In Situ Dye Coupling Experiments
Cell-to-cell coupling in seminiferous tubules was measured by the cut end-loading method originally described by Tsien and Weingart (1976). Freshly dissected testes were cut in half transversely and incubated in PBS containing 0.5% (w/v) Lucifer yellow (MW 475.2; Sigma) and 0.5% (w/v) dextran coupled to rhodamine (Sigma) for 10 min at 32C. The dextran-rhodamine (MW 10,000) was used as a negative control because this dye molecule is too large to pass through gap junctions. The testis fragments were rinsed five times in PBS, fixed in 4% paraformaldehyde for 48 hr at room temperature, and then embedded by the standard paraffin procedure. Sections (5 μm) were cut in plane perpendicular to that of the razor blade cutting. The sections were incubated for 1 min in a solution composed of 2 μg/ml of DAPI to stain nuclei and examined by fluorescence microscopy to determine the cellular distribution of the fluorescent dyes and to identify dye coupled cells. About 100 sections from three different testes were examined. Sections in which cells injured by cutting were loaded with both Lucifer yellow and dextran-rhodamine were selected. To verify that dye coupling is gap junction-mediated, testis fragments from three different testes were preincubated in the presence of 3 mM heptanol, an inhibitor of the gap junctional coupling, and processed as described above. As for controls, about 100 sections in which cells were loaded with both Lucifer yellow and dextran-rhodamine were selected.
Semiquantitative Evaluation
Hybridization intensity was quantified using a computer-assisted image analysis system Visilog 4. 15 (Noesis; Les Ulis, France), consisting of an IMC 500 camera and an IMC 500 digitizer. The intensity of hybridization was expressed as percentage of pixels within a marked area occupied by silver grains that was above a set gray threshold level. Measurements were taken from 10 seminiferous tubules from three different testis sections for each animal group. For each group, two serial sections were analyzed. One was probed with the antisense RNA probe and the remaining section was probed with the sense probe. Both serial sections were analyzed for quantitation. Semiquantitative evaluation of Cx43 immunostaining was performed by use of a computerized image analysis system as previously described (Bourdon et al. 1998; Batias et al. 1999). All the measurements were carried out using the same parameters as the monitor screen and were taken from 10 seminiferous tubules from three different testis sections. Values are expressed as percentage of immunostaining in seminiferous tubules of wild-type mice. Data are expressed as means ± SEM.
Statistical Analysis
A one-way analysis of variance (ANOVA) followed by a Student-Newman-Keuls post hoc test was used to test for differences among groups. Values were considered statistically different when p<0.05.
Results
Localization of Cx43 mRNA and Protein in Mature Testis
In the mature testis, darkfield images reveal a high accumulation of silver grains at the base of seminiferous tubules and in the interstitial compartment of both rat (Figure 1A) and mouse (Figure 2A). Brightfield examinations indicate that the intensity of the ISH signal was changed throughout the spermatogenic cycle of rat (Figures 1D, 1F, and 1H) and mouse (Figures 2D, 2F, and 2H) testes, suggesting that the accumulation of Cx43 transcripts might be dependent on the stage of the seminiferous epithelium cycle. The seminiferous tubules corresponding to Stage VII appeared the most strongly labeled (Figures 1F and 2F). Semiquantitative analysis indicated that the intensity of the signal, measured with the antisense probe, was significantly greater (p<0.05) at Stage VII compared to Stages V and IX in the rat seminiferous epithelium (Figure 3). Control sections incubated with the sense probe did not reveal any specific hybridization. Similar results were obtained in the mouse (not shown).
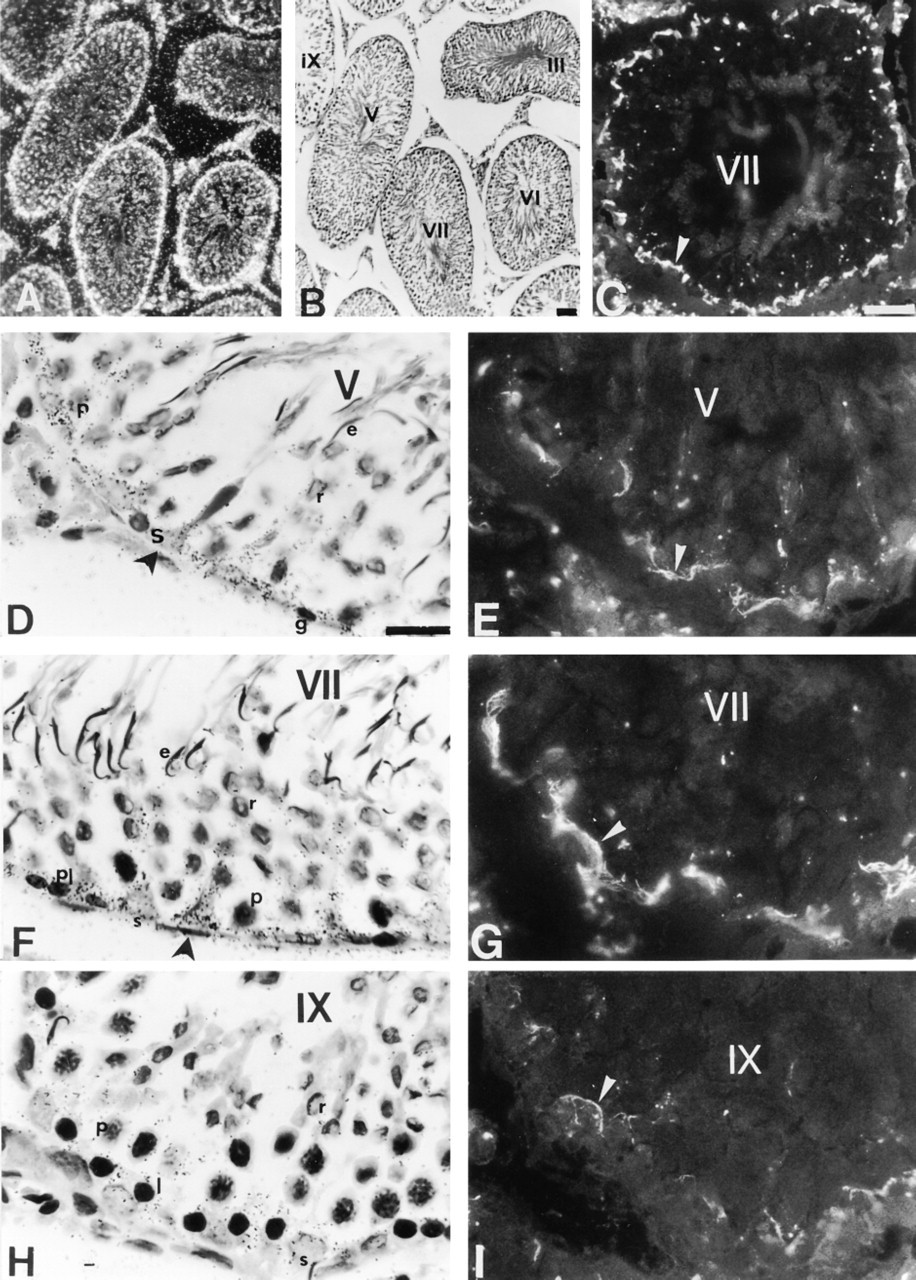
Localization of Cx43 mRNA and Cx43 protein in mature rat testis. (
Immunofluorescence studies showed that Cx43 signal was detected in the basal compartment of seminiferous tubules, and in interstitial cells of both species (Figures 1C and 2C). As observed for Cx43 mRNA, the pattern of Cx43 protein distribution appeared stage-dependent. From Stage I to Stage V of spermatogenesis, faint immunostaining for Cx43 was detected in the basal compartment of rat (Figure 1E) and mouse (Figure 2E) seminiferous tubules. At Stage VII of spermatogenesis, the intensity of the immunostaining, localized in the basal compartment of seminiferous tubules was increased (Figures 1G and 2G). Finally, at Stage IX, the basal immunoreactivity was decreased in both species (Figures 1I and 2I). In controls, no immunoreactivity could be detected (not shown).
The cellular distribution of Cx43 mRNA was examined at the same stages. Transcripts for Cx43 were mainly expressed in the cytoplasm of Sertoli cells (Figures 1D, 1F, 1H, 2D, 2F, and 2H). This localization was clearly evidenced by the columnar distribution of silver grains, a pattern typical for Sertoli cells (Figures 1F and 2F). In addition to Sertoli cells, ISH signals were also present in the basally located germ cells of rat and mouse at all stages considered (Figures 1D, 1F, 1H, 2D, 2F, and 2H). Higher magnification of the basally located ISH signal in both species indicates that silver grains were detected in spermatogonia (Figure 4A), preleptotene spermatocytes (Figures 4B and 4C), and in leptotene and zygotene spermatocytes (not shown). In pachytene spermatocytes, accumulation of Cx43 transcripts appeared to be correlated with their position in the seminiferous epithelium (Figures 4A and 4D). No distinct signal was observed in round and elongated spermatids at any stage of the mouse and rat seminiferous epithelium cycle (Figures 1D, 1F, 2D, and 2F).
In Situ Dye Coupling in Mature Rat Seminiferous Tubule
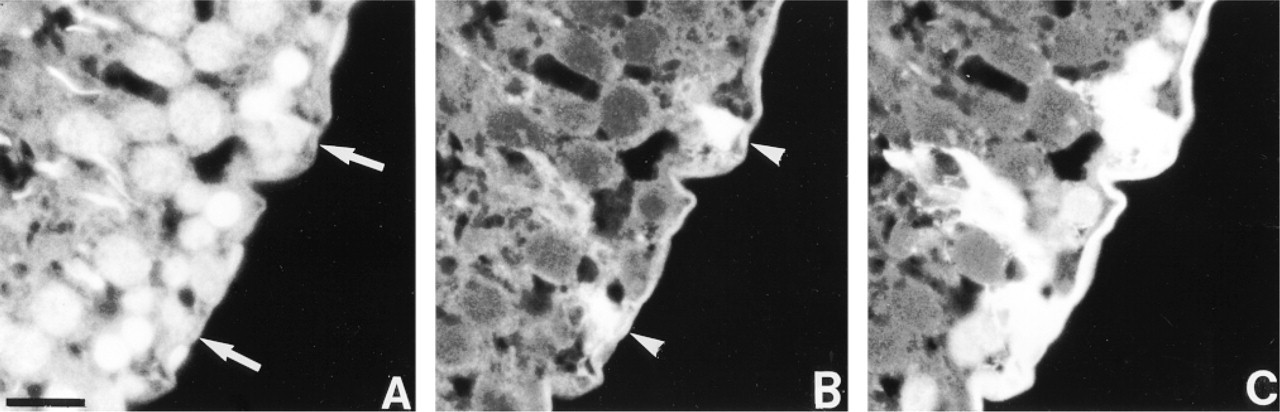
Dye coupling experiments were developed to determine if functional coupling correlates with the distribution of Cx43 mRNA and of the protein within the seminiferous epithelium. As shown in Figure 5, a selective intercellular transfer of Lucifer yellow was observed between cells basally located in the seminiferous tubules and identified by DAPI staining (Figure 5A). In the seminiferous tubule section selected, representative of about 20% of sections examined, dextran-rhodamine fluorescence was detected in Sertoli cells, identified by the specific columnar distribution of the fluorescence (Figure 5B). No transfer of this dye molecule through germ cells could be observed. In contrast, diffusion of the Lucifer yellow occurred from Sertoli cells to spermatogonia and early spermatocytes (Figure 5C). In the other sections analyzed, dye coupling from germ cell to Sertoli cell was never observed. Preincubation of testes in the presence of 3 mM heptanol reduced dye coupling between Sertoli and basal germ cells by 90%, indicating that diffusion of the Lucifer yellow is gap junction-mediated (not shown).
Localization of Cx43 mRNA and Protein in Developing Mouse Testis
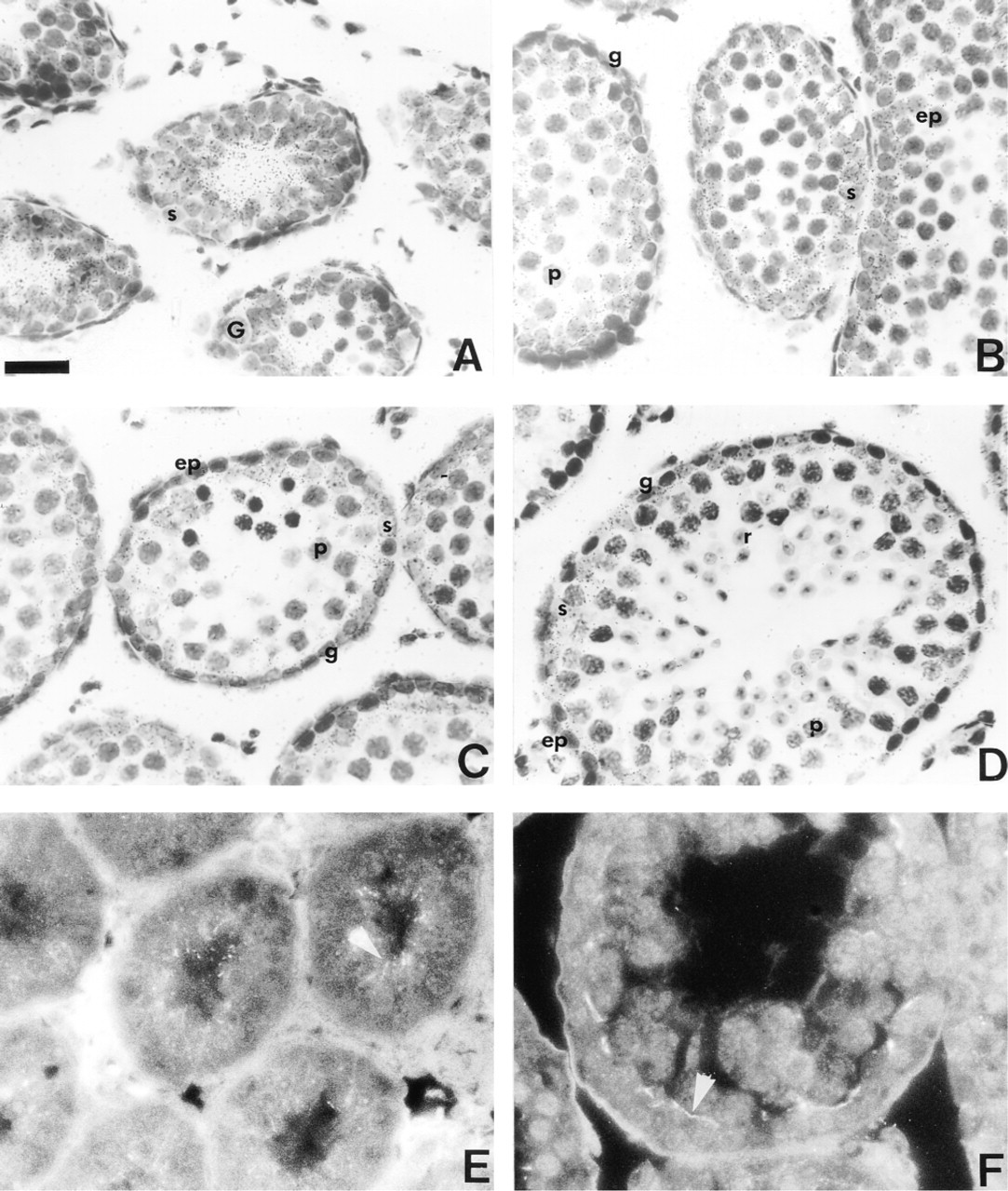
During the first wave of mouse spermatogenesis, Cx43 mRNAs were detected at all postnatal ages studied (Days 4, 8, 12, 16, 19, and 27). Because the localization of Cx43 mRNA was similar from Day 4 to Day 12, only the more representative data are presented here. At Day 12, Cx43 mRNA and Cx43 immunostaining were localized in the adluminal region of all seminiferous tubules (Figures 6A and 6E). At Day 16, Cx43 transcripts became progressively present in the basal region of the tubules (Figure 6B). At this stage, Cx43 immunoreactivity was located in the basal third of the seminiferous tubules (not shown). At Days 19 and 27, Cx43 mRNA (Figures 6C and 6D) and Cx43 immunoreactivity (Figure 6F) were located in the basal compartment of the seminiferous tubules. As in the mature seminiferous epithelium, silver grains were detected in cytoplasm of Sertoli cells and in basally located germ cells (spermatogonia and spermatocytes). For Sertoli cells, the ISH signal was detected at any postnatal age (Figures 6A-6D), whereas for germ cells few silver grains were observed at Day 19 (Figure 6C) and Day 27 (Figure 6D). Cx43 mRNA and Cx43 protein were also present in interstitial and peritubular cells at all stages studied (not shown).
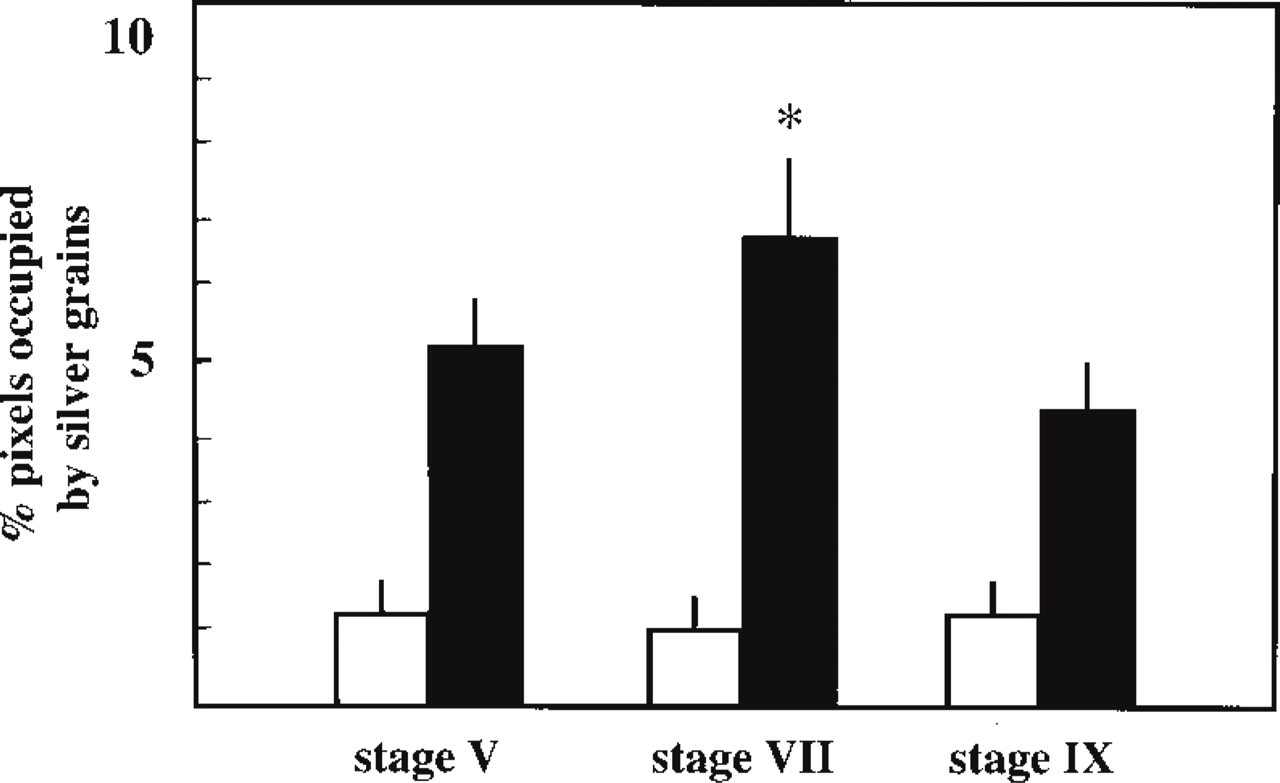
Quantification of Cx43 ISH in rat seminiferous tubules sections probed with the antisense probe (black columns) or with the sense probe (open columns). The intensity of hybridization was quantified as percentage pixels occupied by silver grains. Spermatogenesis stages were identified according to the morphological criteria established by Leblond and Clermont (1952). Values represent the means ± SEM (n=10). ∗Significantly different (p<0.05) from Stage V and Stage IX.
Localization of Cx43 mRNA and Protein in the RXRβ−/− Mutant Mouse Testis
In RXRβ−/− mutant mice, darkfield illumination revealed faint ISH signal for Cx43 mRNA in the seminiferous tubules and in the interstitial compartment of 3- (Figure 7A) and 11-month-old animals (Figure 7C). The accumulation of silver grains at the periphery of the seminiferous tubules was markedly reduced in the 11-month-old mutant (Figure 7C) compared to the mutant at 3 months of age (Figure 7A) and to wildtype mice (Figure 7A). Brightfield illumination at 3 months showed, various germ cell types in the seminiferous tubules (Figure 7B), whereas at 11 months, different degrees of germ cell loss, specifically the elongated spermatids, could be observed (Figure 7D). The intensity of the hybridization signal was significantly reduced in seminiferous tubules of the 3-month-old (p<0.01) and 11-month-old (p<0.01) mutants without significant modification of the background levels revealed with the sense probe (Figure 8). A large decrease in the ISH signal was observed in Sertoli cells of the mutant at both ages studied (Figures 7E and 7G, insets). In 3-month-old mutant mice, the immunoreactive Cx43 signal localized in the basal compartment of seminiferous tubules was reduced (compare Figures 7F and 2G). Cx43 expression decreased by 47 ± 9% compared to wild-type mice (p<0.05). In the 11-month-old mutant, only a few silver grains were detected, but no distinct Cx43 immunostaining was observed (Figure 7H).
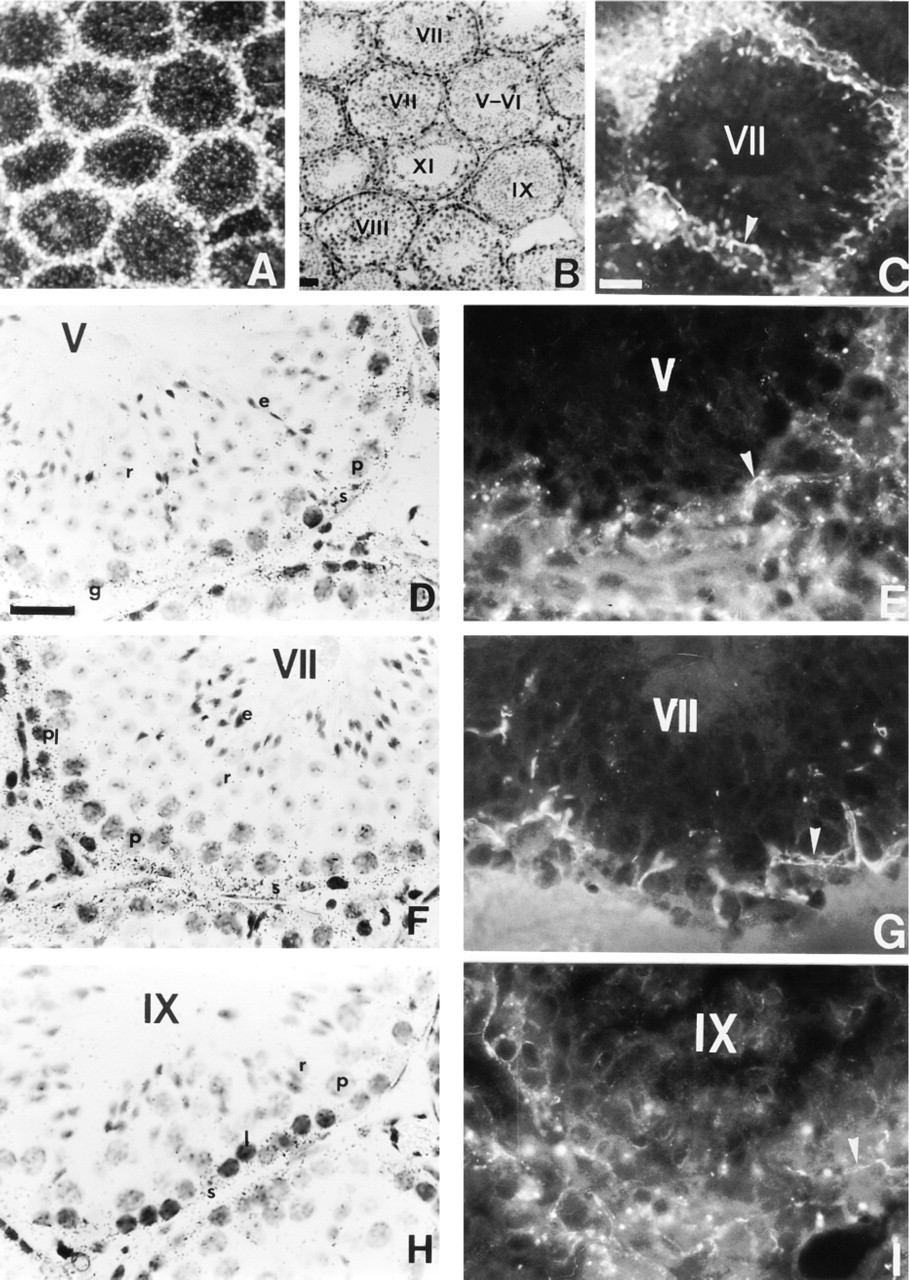
Localization of Cx43 mRNA and Cx43 in mature mouse testis. (
Discussion
Previous studies by Northern blotting analysis have reported the presence in rat testes of mRNA for Cx43, the predominant gap junction protein in this organ (Kadle et al. 1991; Risley et al. 1992). In the present study we show for the first time the exact location of these transcripts in the seminiferous epithelium. Cx43 mRNA is detected in Sertoli cells and in basally located germ cells: spermatogonia and spermatocytes.
The presence of Cx43 transcripts in Sertoli cells is supported by the columnar distribution of the ISH signal detected at the base of the seminiferous tubules which is typical of Sertoli cell localization. An identical distribution has been already reported for mRNA transcripts of Sertoli cell-specific genes (Morales et al. 1987; Kaipia et al. 1992; Millar et al. 1994). The presence of Cx43 mRNA in spermatogonia and in spermatocytes in mature wild-type animals is in agreement with previous freeze-fracture studies from several laboratories, which reported the presence of small gap junctions between these germ cells and Sertoli cells (for review see Enders 1993). In contrast, immunofluorescence studies on testis sections were unable to show Cx43 signal at germ cell surfaces (Risley et al. 1992; Lablack et al. 1998). However, we recently reported that immunofluorescence localization of gap junctions analyzed by confocal microscopy appeared both as continuous and as anastomosed ribbons between Sertoli cells and also as thin dots (Batias et al. 1999). On the basis of these data, we have proposed that the dotted pattern could result from Cx43 associated within the gap junctions between Sertoli cells and germ cells. Despite the use of confocal microscopy, it was not possible to attribute the Cx43 fluorescence signal to a specific type of germ cell. The current observation that ISH signals are present in spermatogonia and spermatocytes enables us to hypothesize that these basally located spermatogenic cells assemble Cx43 into gap junctions with Sertoli cells. This possibility is strongly supported by our current in situ dye coupling experiments, which show functional communication between supporting cells, i.e., Sertoli cells, and germ cells at the base of the seminiferous tubule. The latter observation does not preclude the possibility that other Cx could be implicated in these gap junction channels, because it was reported in a recent study that Cx31 mRNAs are also present in spermatogonia and early spermatocytes (Mok et al. 1999). However, these authors were unable to detect transcripts for Cx31 and immunoreactive Cx31 in Sertoli cells, suggesting that Cx31 is probably not associated with functional gap junction channels between Sertoli cells and germ cells.
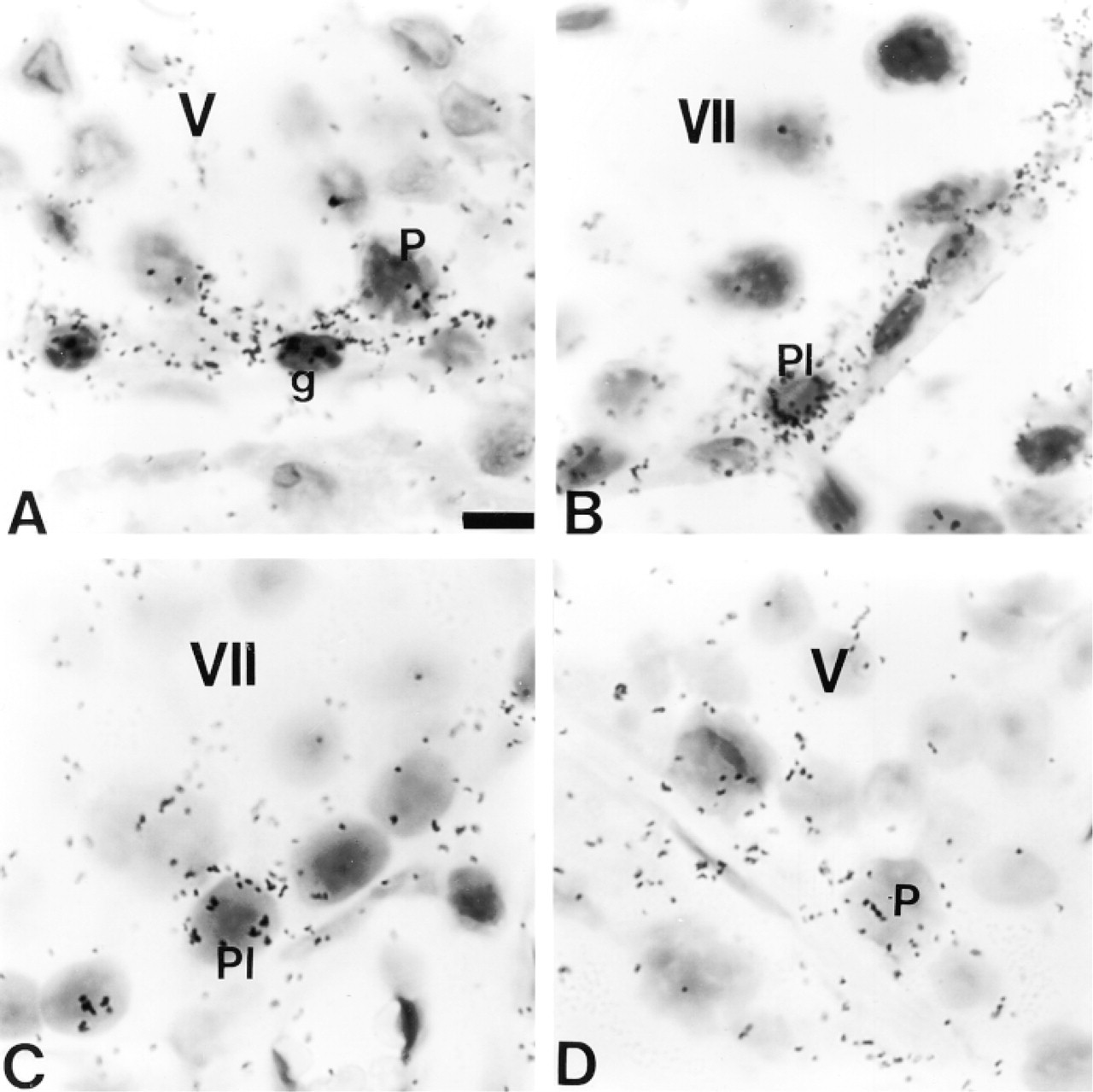
Localization of Cx43 mRNA in germ cells of adult rat (
In situ dye coupling in seminiferous tubules after bulk loading of fluorescent dyes in mature rat testis. (
Developmental expression of Cx43 mRNA (
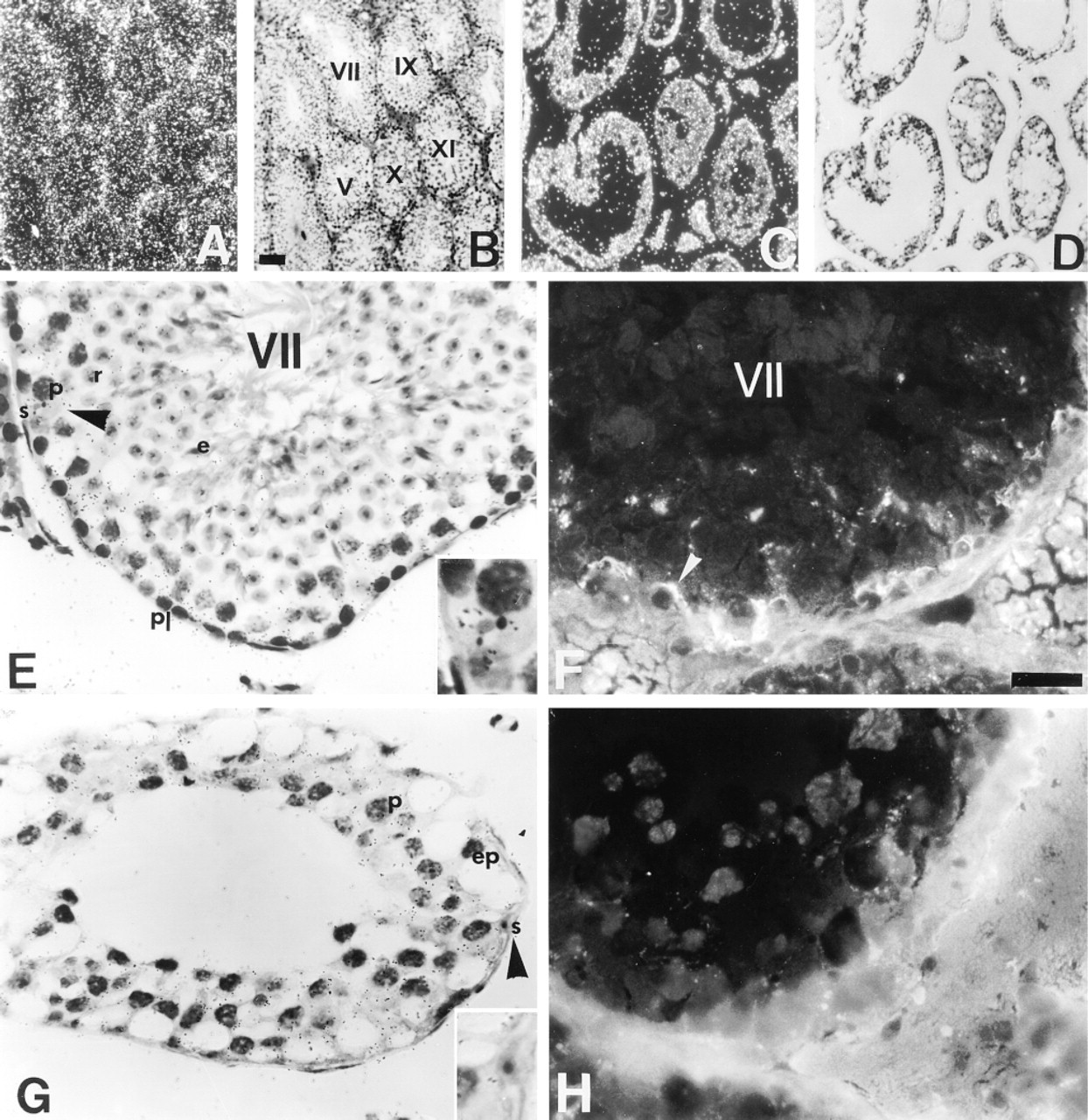
Expression of Cx43 mRNA and Cx43 protein in the RXRβ−/− mutant mouse testis. (
Our present observations also reveal that Cx43 mRNA and protein expression is dependent on the stage of the spermatogenic cycle in the mature testis. In accordance with the results obtained in the protein analysis, which showed a strong immunoreactive signal at Stage VII (Risley et al. 1992; Tan et al. 1996; Lablack et al. 1998; and this study), we observed an increase in Cx43 mRNA level at this stage of spermatogenesis. These data indicate the existence of a direct correlation between the level of Cx43 protein and its gene expression in the seminiferous tubules, and suggest that the higher expression of Cx43 protein at Stage VII of spermatogenesis may be related to an increase in the amount of Cx43 mRNA. However, the mechanisms that drive Cx43 mRNA expression remain to be elucidated.
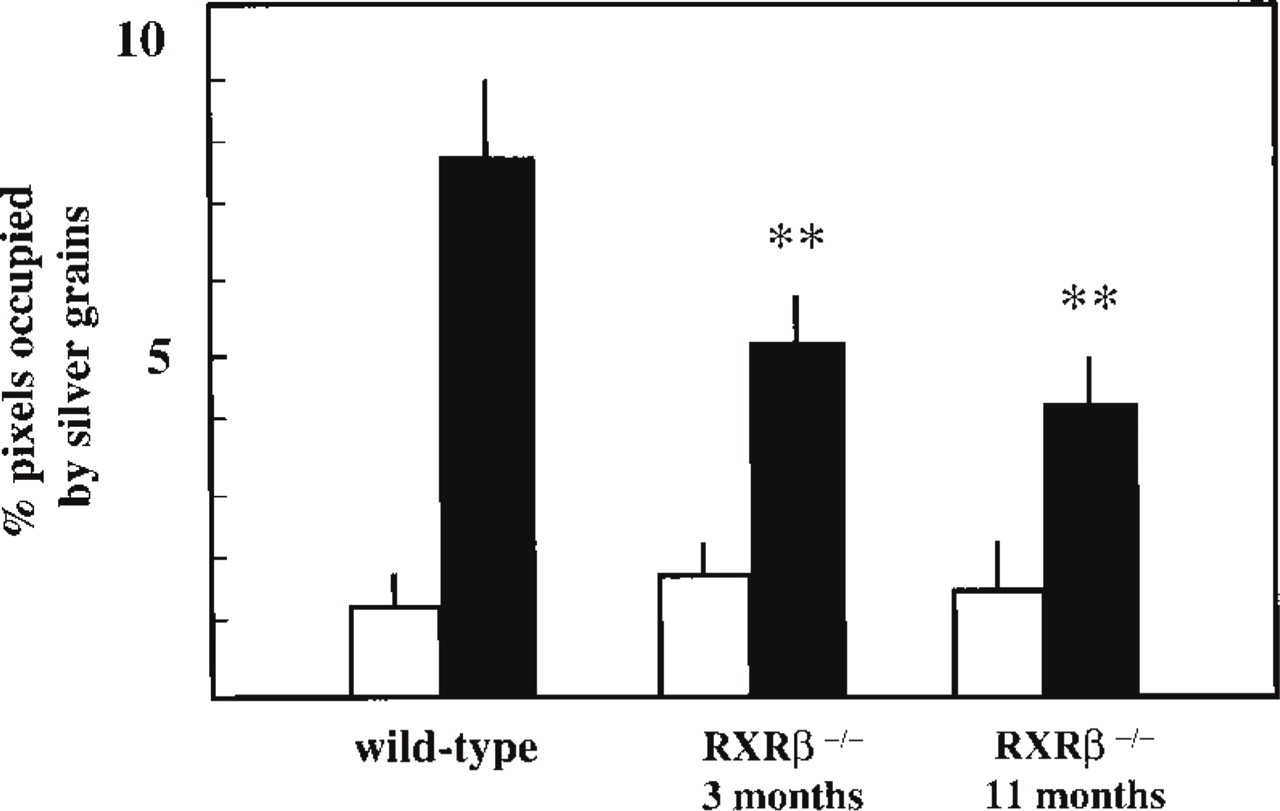
Quantification of Cx43 ISH in seminiferous epithelium of RXRβ−/− mutant testis at 3 and 11 months of age. The intensity of hybridization was quantified as percentage pixels occupied by silver grains. Black columns represent analysis of antisense hybridization. Open columns represent analysis of sense hybridization. Values presented are means ± SEM; n=10. ∗∗ p<0.01 compared to wild-type mice.
In the developing mouse testis, Cx43 mRNAs were detected in seminiferous tubules at all postnatal ages, from Day 4 to Day 27. The distribution of Cx43 transcripts, first in the adluminal region of the seminiferous tubules (Days 4-12) and then in the basal region (later phases) probably result from Sertoli cell cytoplasm organization during development. At the earlier phase, the cytoplasm of palisaded Sertoli cells filled most of the lumen of the growing seminiferous tubules, whereas at the later phase it occupied a larger portion in the basal region of the tubules. This specific distribution of Cx43 mRNA during development coincided with the localization of immunodetectable Cx43 protein as previously observed (Pelletier 1995), and was consistent with the distribution of the gap junctions during mouse testis maturation (Nagano and Suzuki 1976). Although the major ISH signal was found in Sertoli cells during postnatal development of the mouse seminiferous epithelium, the current data indicate that Cx43 mRNAs were also present in basally located germ cells (spermatogonia and spermatocytes). Such observations suggest that gap junctions containing Cx43 occur between these germ cells and adjacent Sertoli cells and could participate in the onset of spermatogenesis.
Another interesting observation is that the expression of Cx may vary in response to various natural factors, such as toxic agents and cell proliferation and transformation (Bruzzone et al. 1996). Among the candidates in the first group are retinoids (vitamin A and its derivatives), which have been shown, through specific nuclear receptors, to upregulate in vitro junctional communications and expression of Cx43 mRNA and protein in mouse fibroblasts (Rogers et al. 1990), in human and rat epidermis (Guo et al. 1992; Risek et al. 1998), and in mouse teratocarcinoma cells (Clairmont et al. 1996). Recently, a mutant mouse deficient in the retinoid X receptor β gene was generated; mutant males that survived exhibited abnormal germ cell maturation due to altered Sertoli cell function (Kastner et al. 1996). The current study clearly shows a reduced expression of Cx43 mRNA and protein in the disorganized seminiferous tubules of this mutant. Interestingly, our observations strongly suggest that the low level of Cx43 mRNA in the retinoid receptor mutant testis appears mainly to result from a reduced number of transcripts in the Sertoli cells. These results are in agreement with the previous observation that the Sertoli cell is the primarily affected target cell in the RXRβ−/− mutant (Kastner et al. 1996). Therefore, altered Cx43 gene expression in Sertoli cells of the mutant mice could affect the gap junction communication between Sertoli cells and between Sertoli and germ cells, thus resulting in abnormal germ cell maturation. Retinoids are essential for normal growth and differentiation of germ cells and for the maintenance of normal spermatogenesis in the mammalian testis, because in the vitamin A-deficient seminiferous epithelium meiotic and postmeiotic germ cells degenerate (for review see Eskild and Hansson 1994). However, little is known about the mechanisms by which retinoids control spermatogenesis. The present findings suggest a possible implication of Sertoli-to-Sertoli and Sertoli-to-germ cell gap junction communication in the control of spermatogenesis by retinoids.
In conclusion, the present data provide evidence that Cx43 mRNA and Cx43 protein, the predominant gap junction protein in the testis, are present in the supporting Sertoli cells and in specific basally located germ cells, spermatogonia and spermatocytes. This cell type-specific expression of Cx43 mRNA and the stage-specific expression of Cx43 protein and its messengers at restricted periods of spermatogenesis suggest that this gap junction protein is involved in a specific function during male germ cell differentiation. Intercellular communication via gap junctions is believed to be involved in the control of cell growth (Loewenstein and Rose 1992). Whether this gap junctional coupling via Cx43 between Sertoli and basally located germ cells is a determinant of spermatogonia multiplication and maturation remains to be elucidated.
Footnotes
Acknowledgements
Supported by INSERM, Ministère de la Recherche et de l'Enseignement Supérieur, and by grants from ARC (no. 98-80). CB was supported by grants from Hoechst Marion Roussel.
We thank Dr J-M Gasc and M-T Morin for their help in in situ hybridization experiments, N. Parseghian, F. Carpentier, and B. Decrossas for technical assistance, J.M. Lepecq for photographic illustrations, and Dr F. Brucker-Davis for revising the English.