
Abstract
The placenta has a dynamic and continuous capacity for self-renewal. The molecular mechanisms responsible for controlling trophoblast proliferation are still unclear. It is generally accepted that the simultaneous activity of proteins involved in cell proliferation, apoptosis, and extracellular matrix degradation plays an important role in correct placental development. We investigated in depth the expression of the serine protease HtrA1 during pregnancy in human placenta by in situ hybridization and immunohistochemistry, we demonstrated that HtrA1 displayed a low level of expression in the first trimester of gestation and a strong increase of HtrA1 expression in the third trimester. Finally, by electron microscopy, we demonstrated that HtrA1 was localized either in the cytoplasm of placental cells, especially close to microvilli that characterized the plasma membrane of syncytiotrophoblast cells, or in the extracytoplasmic space of the stroma of placental villi, particularly in the spaces between collagen fibers and on collagen fibers themselves. The expression pattern of HtrA1 in human placentas strongly suggests a role for this protein in placental development and function. Moreover, on the basis of its subcellular distribution it can be postulated that HtrA1 acts on different targets, such as intracellular growth factors or extracellular matrix proteins, to favor the correct formation/function of the placenta.
C
The bacterial serine protease HtrA, also known as DegP, is a heat shock-induced envelope-associated serine protease (Pallen and Wren 1997). Its chief role is recognition and degradation of misfolded proteins in the periplasm, combining a dual activity of chaper-one and protease (Spiess et al. 1999; Krojer et al. 2002). Recently, three human homologues of HtrA have been identified and named human HtrA1 (PRSS11 or L56) (Zumbrunn and Trueb 1996; Hu et al. 1998), HtrA2 (Omi) (Faccio et al. 2000; Gray et al. 2000), and HtrA3 (PRSP) (Nie et al. 2003a). These proteases appear to be involved in diverse functions, such as cell growth, apoptosis, and inflammation (Clausen et al. 2002). In particular, HtrA2 plays an important role in programmed cell death, eliminating the caspase-inhibitory activity of inhibitor of apoptosis proteins (IAPs) and thus promoting cell death (Martins et al. 2002; van Loo et al. 2002; Verhagen et al. 2002). The precise function of HtrA3 is still largely unknown. Given the high structural similarity with HtrA1, it has been suggested that HtrA3 could display functions similar to HtrA1 (Nie et al. 2003a). Therefore, it has been recently demonstrated that HtrA3 is dramatically increased during placentation (Nie et al. 2003b). HtrA1 is a secreted protein, which is upregulated during osteoarthritis and is significantly downregulated in transformed fibroblasts, in ovarian cancer and in melanoma (Zumbrunn and Trueb 1996; Hu et al. 1998; Shirdar et al. 2002; Baldi et al. 2002, 2003). Moreover, its overexpression inhibits cell growth and proliferation in vitro and in vivo, thus suggesting a possible role as a tumor suppressor (Baldi et al. 2002). The differential expression of HtrA1 in human tissues (De Luca et al. 2003) and the observation that the transcription of this gene is highly regulated between fetal liver and postnatal liver (Nagata et al. 2003) strongly suggest that HtrA1 exerts its function of control on cell growth not only in neoplastic cells but also under physiological conditions.
HtrA1 is a protein with two distinct domains of homology. The N-terminal domain is homologous to mac25, a recently characterized gene product related to insulin-related growth factor-binding protein (Swisshelm et al. 1995) and follistatin (Kato et al. 1996). The second domain is more than 40% identical to bacterial HtrA serine protease (Hu et al. 1998). It is therefore conceivable that HtrA1 may interact with IGF-I or IGF-II (Zumbrunn and Trueb 1996). Moreover, the end of the IGF-binding domain and the beginning of the protease domain resemble the Kazal type of protease inhibitor (Zumbrunn and Trueb 1996). The presence of this protease inhibitor motif suggests that the human HtrA1 serine protease may be a self-regulating enzyme or that it may regulate other serine proteases (Hu et al. 1998). During pregnancy, IGFs are important for growth of fetal and maternal tissues. Therefore, IGF-II is a major modulator of placental and fetal growth (Nie et al. 2003b). It is well documented that the activity of IGF-I and IGF-II is modulated by IGF-binding proteins (IGFBPs) that control the availability of these growth factors for their receptors (Zumbrunn and Trueb 1996). The IGFs are liberated from the complexes by specific proteases belonging to the family of the serine proteases (Zumbrunn and Trueb 1996). In particular, the protease specific for IGFBP-4 is most similar to HtrA1 (Zumbrunn and Trueb 1996). IGFBP-4 is the second most abundant IGFBP in the maternal decidua (Han et al. 1996) and is a potent inhibitor of IGF actions (Rajaram et al. 1997).
Drawing on this background, in this study we investigated in depth the expression of HtrA1in the human placenta.
Materials and Methods
Samples
Specimens from first trimester placentas were obtained from uterine evacuations (n = 15) and samples from third trimester placentas were obtained from cesarean sections or after spontaneous delivery at term (n = 15). Indications for cesarean sections and evacuations were always elective. The gestational age of the first trimester samples ranged from 5 to 14 weeks and the gestational age of the third trimester specimens ranged from 28 to 40 weeks. The collected specimens were immediately fixed in formalin, 4% paraformaldehyde, or 2.5% glutaraldehyde for immunohistochemistry (IHC), in situ hybridization (ISH), and electron microscopy, respectively. Representative sections of each specimen were stained with hematoxylin-eosin and examined by a pathologist to confirm histological preservation of the microanatomic structure. The absence of any placental pathology was confirmed by microscopic observation.
In Situ Hybridization
The specimens collected for ISH were immediately fixed in 4% paraformaldehyde and left to stand overnight at 4C. They were then stored in 75% ethanol, passed through an increasing concentration gradient of ethanol, and embedded in paraffin. Sections from each specimen were cut at 5–7 μm and mounted on baked glass slides precoated with a 2% solution of TESPA (3-aminopropyl triethoxysilane; Sigma, St Louis, MO) in acetone. Slides were then dried at 37C overnight to allow sections to adhere to the slides. All solutions were made using bidistilled water treated with 0.1% DEPC (diethylpyrocarbonate; Sigma) to inactivate RNases and then autoclaved. The cDNA corresponding to the C-terminal domain of the human HtrA1 protein (aa 363–480) was cloned into pGEX-2Tk vector (Pharmacia; Piscataway, NJ) between the BamHI and EcoRI sites. After digestion, a fragment of 350 bp was generated, sense and antisense digoxigenin (DIG)-labeled RNA probes were obtained using the DIG RNA Labeling kit (Boehringer Mannheim; Mannheim, Germany), and the concentrations determined according to the manufacturer's instructions.
Sections after dewaxing were washed in PBS with 0.3% Triton X-100, permeabilized in Tris-EDTA containing 10 μg/ml Rnase-free proteinase K at 37C for 10 min, and post fixed in 4% paraformaldehyde in PBS, pH 7.4, for 30 min. Prehybridization was performed for 30 min in 2 × SSC with Tris/glycine. Hybridization was performed in the following buffer: 40% formamide, 5 × SSC, 1× Denhardt's solution, 100 μg/ml yeast tRNA, 100 μg/ml denatured salmon sperm DNA, 100 ng DIG-labeled RNA probe. Sections were then incubated overnight at 60C. Slides were washed in 5 × SSC at RT for 20 min, then washed in 0.5 × SSC/20% formamide at 60C for 40 min. Finally, sections were incubated in NTE buffer (0.5 M NaCl, 10 mM Tris, pH 7.0, 0.5 M EDTA) at 37C for 30 min. Then background blockade and DIG revelation were performed using a DIG Detection Kit (Boehringer Mannheim) and developed with BM purple (Boehringer Mannheim).
Immunohistochemistry
IHC was carried out essentially as described previously (Baldi et al. 2002). Briefly, sections from each specimen embedded in paraffin were cut at 5–7 μm, mounted on glass, and dried overnight at 37C. Sections were then deparaffinized in xylene, rehydrated through a graded alcohol series, and washed in PBS. PBS was used for all subsequent washes and for antiserum dilution. Tissue sections were quenched sequentially in 3% hydrogen peroxide in aqueous solution and blocked with PBS-6% nonfat dry milk (BioRad; Hercules, CA) for 1 hr at RT. Slides were then incubated at 4C overnight with affinity-purified rabbit polyclonal antiserum raised against a purified bacterially expressed glutathione-S-transferase (GST)-HtrA1 (aa 363–480) human fusion protein (Baldi et al. 2002) at a 1:10 dilution. After several washes to remove excess antiserum, the slides were incubated with diluted goat anti-rabbit biotinylated antibody (Vector Laboratories; Burlingame, CA) at a 1:200 dilution for 1 hr. Slides then were processed by the ABC method (Vector Laboratories) for 30 min at RT. Novared (Vector Laboratories) was used as the final chromogen and hematoxylin was used as the nuclear counterstain. Negative controls for each tissue section were prepared by substituting the primary antiserum with the isotype-matched nonimmune IgG. All samples were processed under the same conditions. The expression level of HtrA1-stained cells per field (×250) at light microscopy was calculated and compared in different specimens by two separate observers (A.B. and F.B.) in a double-blind fashion and was described as score 0 (less than 1% positive cells), score 1 (1–10% positive cells), score 2 (10–20% positive cells), and score 3 (more than 20% positive cells).
An average of 22 fields was observed for each specimen. All values were expressed as mean ± SEM and differences were compared using Student's t-test.
Immunoelectron Microscopy
Placentas were collected, cut into pieces less than 1 mm3, and fixed in 2.5% glutaraldehyde in phosphate buffer (pH 7.3) for 2 hr. Tissues were postfixed in osmium tetroxide (4%), dehydrated through graded concentrations of ethanol and propylene oxide, and embedded in Epon 812. For the immunogold technique, tissues were fixed only in 2.5% glutaraldehyde in phosphate buffer (pH 7.3). Ultrathin sections were cut from blocks and mounted on copper grids for morphology or on nickel grids for immunogold. The immunogold technique was carried out essentially as described previously (Polak and Van Noorden 1987). All sections were etched on drops of 3% hydrogen peroxidase for 10 min to permeabilize resin. Tissue sections were washed in microfiltered distilled water and floated for 30 min on a drop of background blocking solution (0.05 M Tris-HCl, pH 7.4, 0.1% BSA). Grids then were incubated at 4C overnight with the rabbit polyclonal antiserum raised against HtrA1 (Baldi et al. 2002) at a 1:10 dilution. After several rinses to remove excess antiserum, the grids were incubated with anti-rabbit antibody conjugated with 10-nm gold particles used as a secondary antibody at a dilution of 1:100 in diluent for gold-adsorbed antibody (0.05 M Tris-HCl buffer, pH 8.2, 1% BSA) for 1 hr. The grids were counterstained with lead citrate and uranyl acetate as for conventional electron microscopy. A preimmune serum was used as a negative control. The sections were observed with a Siemens Elmiskop I A electron microscope.
Results
Expression of HtrA1 mRNA in Human Placenta
The cell types that express HtrA1 mRNA in the human placenta were determined by ISH using riboprobes specific for HtrA1. In the first trimester of gestation, a very low level of expression was detected in the cells surrounding villi both in the inner layer of the cytotrophoblast and in the outer layer of the syncytiotrophoblast (Figure 1a). The intensity of placental staining for HtrA1 mRNA was clearly increased in the third trimester of gestation. At this time a very strong signal was especially detected in the outer layer forming the syncytiotrophoblast (Figure 1b). This expression pattern was confirmed in four independent specimens taken from first trimester placentas and four from third trimester placentas.
Localization of HtrA1 Protein in Human Placenta
The localization of HtrA1 protein in the human placenta was performed by IHC using a rabbit polyclonal antiserum produced by immunizing rabbits with a chimeric GST-HtrA1 (aa 363–480) protein. The specificity of this antiserum and its suitability for IHC were previously validated by several experiments (Baldi et al. 2002). In the first trimester human placenta, a low level of HtrA1 staining was observed in both layers surrounding placenta villi, even if a high percentage of cells of the cytotrophoblast compartment appeared positive for HtrA1 protein compared to the syncytiotrophoblast (Figure 1c). In the third trimester we observed a strong increase of HtrA1 staining in accordance with expression of HtrA1 mRNA. At this time we observed elevated expression of HtrA1. This expression was higher in the syncytiotrophoblast than in the cytotrophoblast (Figure 1d). Moreover, we also observed positive expression in the stromal components of the placental villus in accordance with its secretory properties. These results were obtained on a total of 30 samples, 15 full-term placentas and 15 chorionic villi from the first trimester. Figure 2 shows the peculiar expression pattern of HtrA1 during pregnancy as determined by IHC analysis. Specifically, we observed an increase in HtrA1 expression from the first to the third trimester of gestation, in which its distribution was more elevated in the syncytiotrophoblast than in the cytotrophoblast.
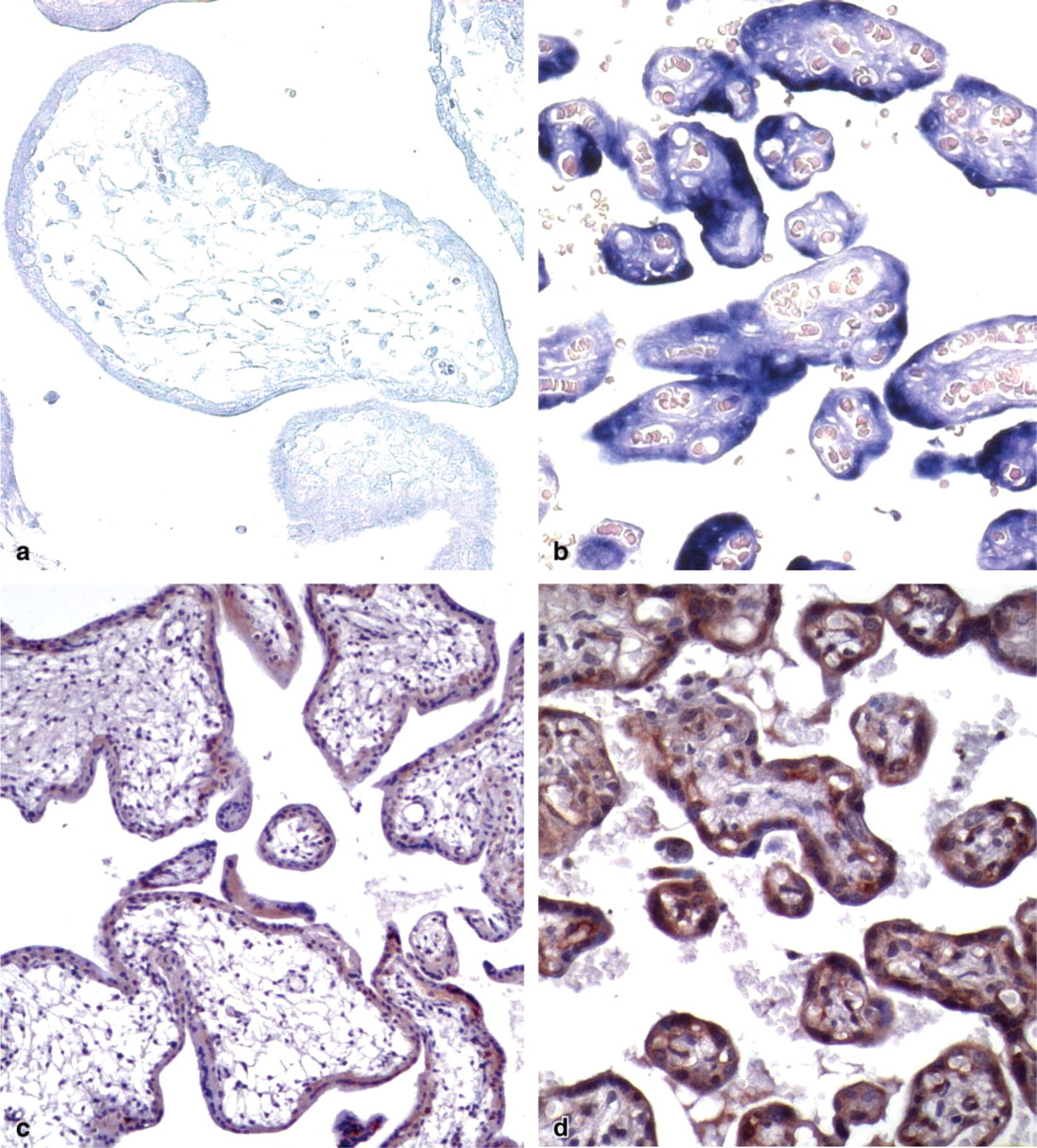
Pattern of expression of HtrA1 in human placenta during pregnancy. Human placentas of first (
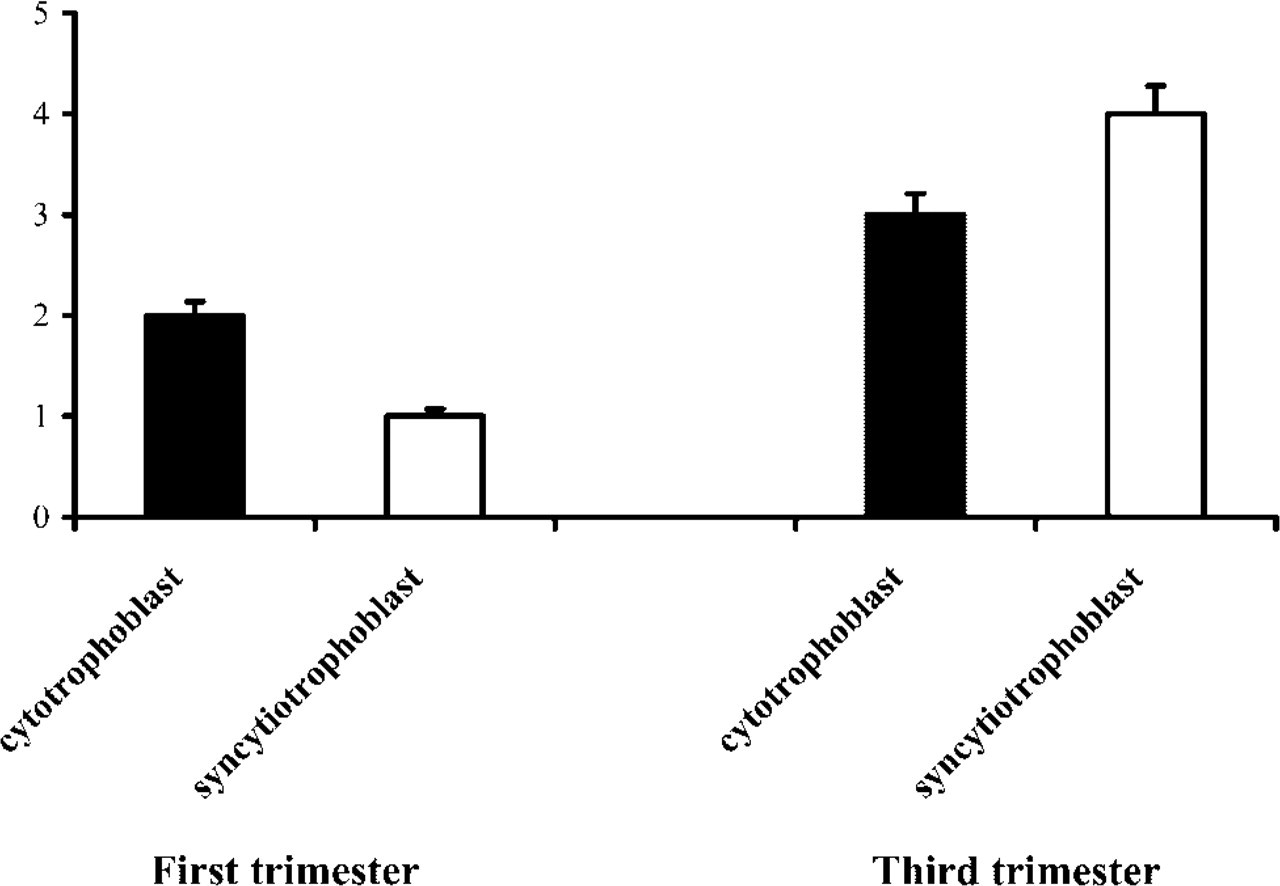
Gestational time course of syncytiotrophoblast and cytotrophoblast immunoreactivity for HtrA1. Ordinate: immunoreactivity scored as described in Materials and Methods. Abscissa: trimester of gestation. Vertical lines show SEM.
Subcellular Localization of HtrA1 Protein in Human Placenta
Subcellular localization of HtrA1 protein in the human placenta was performed by the immunogold technique using a rabbit polyclonal antiserum against HtrA1 (Baldi et al. 2002). We observed that HtrA1 was dispersed in the cytosol of placental cells (Figure 3a). Moreover, we observed clear positivity for HtrA1 close to the microvilli that characterize the plasma membrane of syncytiotrophoblast cells, in accordance with the fact that HtrA1 can be a secreted protein (Figure 3b). In addition, we observed a peculiar localization of HtrA1 protein in extracytoplasmic space of the stroma of placenta villi, consistent with a secreted protein. In particular, positivity for HtrA1 was localized in the spaces between collagen fibers (Figure 3c). Intriguingly, we also observed several gold particles that represent HtrA1 positivity on the collagen fibers themselves (Figure 3d). These results were confirmed in four different specimens of placenta, without significant differences between full-term placentas and chorionic villi from first trimester.
Discussion
The placenta has a dynamic and continuous capacity for self-renewal. Throughout gestation the placenta shows a high cell proliferation rate and lack of cell contact inhibition. For these morphological characteristics, the term “pseudo-malignancy” has been used to describe this tissue (Ohlsson et al. 1993). Most of the continuous remodeling of this tissue is the result of apoptotic events. Therefore, regulation of the balance between cell proliferation and cell death in the placenta is essential to allow the necessary morphological and functional changes (Smith et al. 1997; Lea et al. 1999; Levy and Nelson 2000). Abnormalities in these processes may lead to gestational abnormalities such as miscarriage, fetal growth restriction, and preeclampsia (Cross 1996). Moreover, the stepwise progression starting from the villous cytotrophoblast to the invasive extravillous cytotrophoblast is characterized by dramatic change in expression of cell adhesion molecules and proteinases that degrade the extracellular matrix (Fisher and Damsky 1993).
We have investigated the expression pattern of the serine protease HtrA1 during pregnancy in humans. Data at both the RNA and the protein levels clearly showed a strong increase in HtrA1 expression from the first to the third trimester of gestation, being higher in the syncytiotrophoblast than in the cytotrophoblast. This behavior is very similar to that already described for HtrA3, the third member of the human HtrA family of serine proteases, and is consistent with the hypothesis that these two proteins display similar functions (Nie et al. 2003b). Analysis of the subcellular localization of HtrA1 protein by electron microscopy demonstrated that this protein was localized either in the cytoplasm, in association with plasma membrane microvilli, or in the extracytoplasmic space of the stroma of placenta villi between collagen fibers or on the collagen fibers themselves. This expression pattern suggests that HtrA1 may have different protein targets in the cytoplasm and in the extracytoplasmic space.
These data are consistent with the fact that HtrA1 is considered a secreted protein (Hu et al. 1998). Moreover, Hu and co-workers (1998) suggest a possible involvement of HtrA1 in the complex molecular network responsible for placentation, particularly in the orchestrated balance of cytotrophoblast cell proliferation and differentiation into either of the differentiated cell types, syncytiotrophoblast or invasive cytotrophoblast (Cross et al. 1994). Interestingly, analysis of HtrA1 protein sequence (Hu et al. 1998) has revealed that the N-terminal region of human HtrA1 closely resembles that of mac25, a putative growth suppressor considered to act as an activated follistatin (Kato et al. 1996). It is well known that follistatin plays an important role in the negative regulation of activin A, a glycoprotein belonging to the transforming growth factor beta (TGFβ) superfamily (Massague 1990). Activin A is one of the many growth factors produced by placental and fetal membranes and is considered a local regulator of human cytotrophoblast cell differentiation (Caniggia et al. 1997). The observation that HtrA1 is highly expressed in the epithelium of proliferative endometrium, in contrast to epithelium of secretory endometrium, where the expression of HtrA1 is very low (De Luca et al. 2003), further supports the hypothesis that HtrA1 takes part in the complex pathway of local and systemic growth factors, such as sex steroid hormones, that modulates physiological and neoplastic tissue remodeling and neovascularization of uterus and placenta (Petraglia et al. 1997). Nevertheless, a follistatin-like molecule is typical of some extracellular matrix proteins, such as BM-40, and is partially responsible for enhancing collagen affinity (Sasaki et al. 1997). The extracellular localization of HtrA1 in the spaces between collagen fibers or on the fibers themselves strongly suggests a direct action of this protein on collagen structures, perhaps in the regulation of extracellular matrix components.
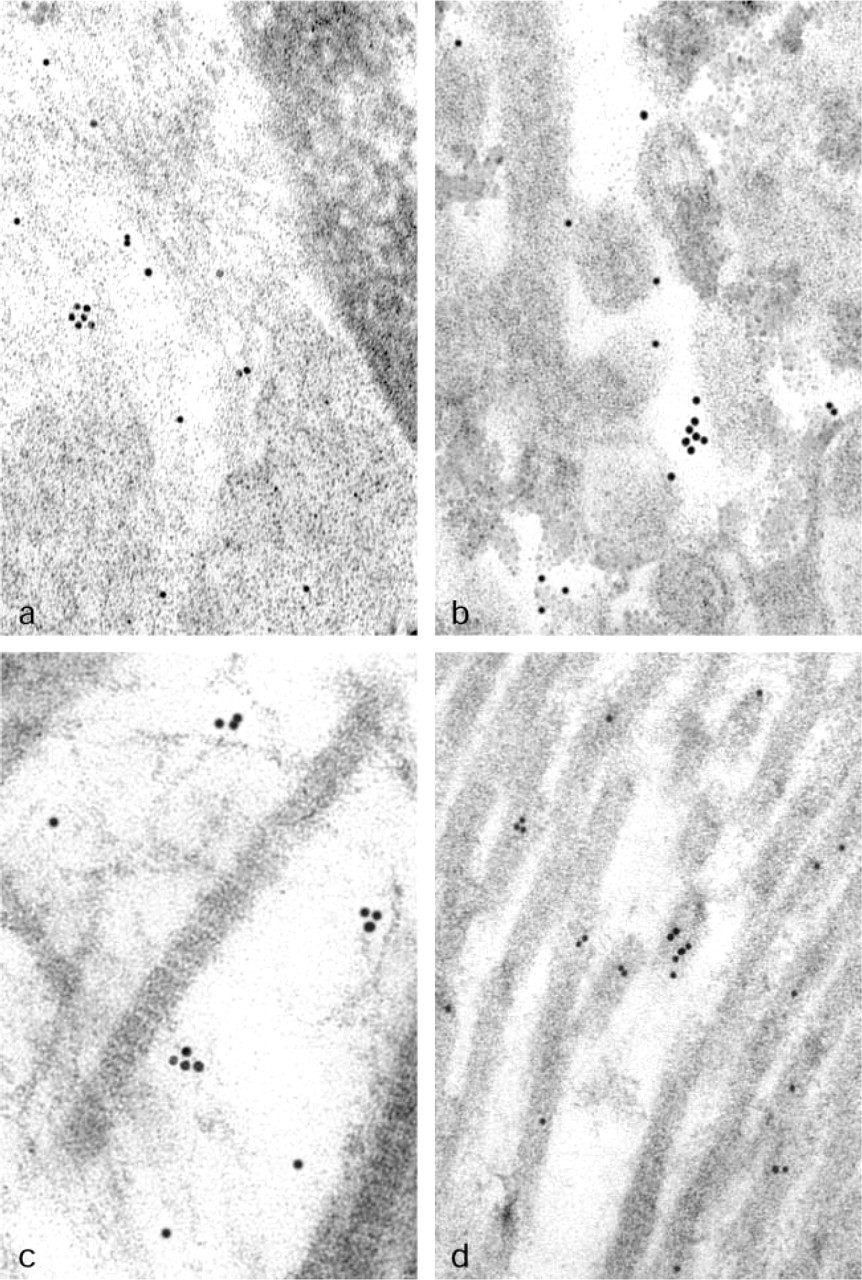
Subcellular localization of HtrA1 protein in human placenta using electron microscopy. (
The presence of an IGF-binding domain is also important. The IGF family plays an important role in implantation and placental physiology (Giudice et al. 2002). IGF-II is abundantly expressed by placental trophoblasts and has effects on trophoblast function modulators of invasion (Irwin et al. 2001), and regulation of this growth factor in the placental bed is important for trophoblast homeostasis and invasion (Giudice et al. 2002). Ablation of either the IGF-I or the IGF-II gene reduced fetal size to 60% of that in normal littermates (Warburton and Powell-Braxton 1995) and when both genes were deleted, pup size was reduced to 30% (Liu et al. 1993; Warburton and Powell-Braxton 1995; Nie et al. 2003b). It has been demonstrated that IGFBPs regulate IGF bioavailability. Among IGFBP proteins, IGFBP-4, a potent inhibitor of IGF actions, is abundantly expressed by the maternal decidua (Han et al. 1996). Human trophoblasts secrete an IGF-dependent IGFBP-4 protease, PAPP-A (Giudice et al. 2002) that is most similar to HtrA1 (Zumbrunn and Trueb 1996). PAPP-A levels increase in the maternal circulation as pregnancy progresses (Brambati et al. 1993). Because HtrA1 possesses an IGF-binding domain and becomes highly expressed as pregnancy proceeds, it is reasonable that HtrA1 might function in the regulation of the IGF–IGFBP system to regulate placental function and development.
In conclusion, the expression pattern of HtrA1 in human placenta strongly suggests a role for this protein in pregnancy. Moreover, its subcellular distribution may imply that HtrA1 acts on different targets, such as intracellular growth factors or extracellular matrix proteins, to favor the correct formation/function of the placenta. Further studies are needed to better define the involvement of this protein in the complex mechanism of trophoblast maturation and to demonstrate the possible therapeutic utility of targeting this pathway.
Footnotes
Acknowledgements
Supported in part by Second University grants to ADL and AB, by Futura-ONLUS to AB, and by AIRC and Ministero della Salute grants to MGP. We thank ISSCO for its continuing support. Dr I.L. Tuduce is on leave of absence from the Dept. of Biology, Faculty of Chemistry, Biology and Geography, West University of Timisoara, Romania and is recipient of a fellowship “Marie Curie Training Site,” contract no. HPMY-CT-2000–00210.
We also thank Mr. Giuseppe Falcone for his contribution to the image elaboration.