
Abstract
Explanted placental fragments may provide a more physiological in vitro model of component cell function than single cell isolates. We have characterized these fragments for cell turnover and have monitored responses from 14 normal placentas under conditions of exogenous TNFα and atypical oxygen concentrations (3% and 17%), conditions associated with abnormal pregnancy and an aberrant in utero environment. Explants were assessed for apoptotic morphology, immunolocalization of Mib-1 (a proliferation marker), caspase 3 activity (an apoptosis promoter), lactate dehydrogenase (a necrosis marker), and human chorionic gonadotrophin [hCG, a marker of cytotrophoblast (CT) differentiation]. Consistent with a reduction in hCG, explants under 17% O2 (with and without TNFα) showed a progressive degeneration of syncytiotrophoblast (ST) (days 0–2) followed by a restoration of hCG (days 4–8) localized to newly differentiated but not syncytialized CTs. In 3% O2, hCG showed the same initial decline but failed to recover thereafter. Proliferation dropped significantly in 17% O2 but was restored and exaggerated sixfold in 3% O2. All reductions in hCG were associated with cell death and caspase-3. Early apoptosis was linked with syncytial loss; later apoptosis (days 8–11) was localized to the non-ST. Prolonged exposure to TNFα (days 4–11) increased ST apoptosis and necrosis but 3% O2 had no significant effect. These findings show that placental explants can accommodate many aspects of CT proliferation, differentiation, and ST apoptosis in culture. TNFα enhanced ST decline but 3% oxygen (compared with 17%) was associated with reduced CT differentiation and a strong shift towards proliferation. These outcomes may reflect previous morphological changes in compromised pregnancies and confirm a possible role for oxygen and TNFα in aberrant trophoblast turnover.
T
To investigate this unusual form of cell turnover, previous studies of CTs have used cultures of isolated primary cells in vitro both before and after differentiation into ST layers and “giant cells” (Morrish et al. 2002). With this technique, it has been suggested that both CTs and STs are sensitive to chronic conditions that may challenge the placenta, such as those of hypoxia, reperfusion, and inflammatory cytokines (Crocker et al. 2001; Hung et al. 2002), conditions identified in preeclampsia (PE) and intrauterine growth restriction (IUGR) (Roberts and Hubel 1999; Serin et al. 2002). Although undoubtedly a useful model, study of cells in isolation produces results of only limited value, as this fails to accommodate both cell-cell and cell-matrix interactions. Recent evidence suggests that fragments of villous tissue in culture may provide a more physiological representation of the cellular kinetics of the placenta (Siman et al. 2001). Although debate surrounds the true agreement between morphological and functional characteristics in this system (Watson et al. 1995), the degeneration and regeneration of the placental syncytium may recreate many fundamental aspects of CT turnover in vivo.
For this study, we proposed that placental villous fragments more closely reflect cellular events within the placental villus than isolated cell preparations in vitro. Moreover, we hypothesized that aberrant placental conditions can disrupt cell responses, affecting both lifespan and turnover of the villous components, with potential implications for inappropriate placental function and development. To test this hypothesis and to characterize this model for future investigations, we have monitored both morphological and biochemical changes of CTs and other cells in term placental villous explants under various oxygen (O2) conditions (3% and 17%) in the presence or absence of exogenous tumor necrosis factor-alpha (TNFα). Particular attention has been paid to the location and loss of cell integrity, either through necrotic and apoptotic cell death, the differentiation of CT and syncytial formation, and the proliferative effects of pathological conditions on both individual trophoblast compartments, stromal cells, and fetal capillary endothelia.
Materials and Methods
Participants
Fourteen placentas were obtained from women at term with uncomplicated pregnancies after elective Cesarean section at St Mary's Hospital (Manchester, UK). All participants gave informed consent before inclusion and the study was approved by the Local Ethics Committee. Women with essential hypertension, preeclampsia (PE), and evidence of aberrant fetal growth or maternal medical complications, such as diabetes or renal disease, were excluded from the study.
Explant Cultures
The explant culture system was developed from the procedures of Siman et al. (2001). Three areas of each placenta were sampled randomly using a transparent sheet bearing a systematic array of sampling windows. Within 30 min of delivery, chorionic non-anchoring villi were dissected from these areas and carefully rinsed in sterile PBS at 37C. The placental tissue was then cut into pieces weighing ∼2–4 mg wet weight. For each individual measurement, four pieces of tissue were cultured on a single Costar Netwell support (15-mm diameter, 74-μm mesh; Corning, Corning, NY) in 1.5 ml of culture medium (CMRL-1066; 5% heat-inactivated fetal bovine serum, 100 IU/ml penicillin, 100 μg/ml streptomycin, 1 μg/ml insulin, 0.1 μg/ml hydrocortisone, and 0.1 μg/ml retinyl acetate; Sigma Chemical, Poole, UK). The tissue was supported on the mesh in the liquid-gas interface. Cultures were maintained at 37C in a humidified gas mixture of 5% CO2/95% air, or 3% O2 with 5% CO2 and 92% N2. Medium was changed every 24 hr. Supernatants were collected and stored at −80C for the analysis of lactate dehydrogenase (LDH) and human chorionic gonadotrophin (hCG). The cultures were maintained for up to 11 days for functional and morphological evaluation. Cultures showing bacterial contamination were interrupted and excluded.
Experimental Conditions
Conditions of reduced oxygen were generated using an atmosphere control chamber with purge airlock and forced air incubator (Coy Laboratory Products; Detroit, MI). All dissections were conducted within the relevant oxygen environments of 3% or 17% oxygen. Pre-equilibration of media was conducted for 24 hr before addition. The percentage of dissolved oxygen in the culture media was analyzed on three separate occasions using an ISO2 dissolved oxygen meter and oxygen electrode. Oxygen in the media exposed to 3% O2 with 5% CO2 and 92% N2 was 3.3 ± 0.4% and to 5% CO2 95% air was 17.4 ± 0.6%.
TNFα was used at a final concentration of 10 ng/ml (Peprotech; London, UK) with interferon gamma (IFN-γ) at 100 U/ml (Peprotech). This combination of cytokines has previously been shown to be bioactive in studies of isolated trophoblast preparations (Crocker et al. 2001; Smith et al. 2002). Both cytokines are expressed in human placental villi during normal pregnancy and their aberrant expression has been implicated in intravillous infections and placental complications (Yui et al. 1994). IFN-γ is believed to enhance responsiveness to TNFα in these systems (Garcia-Lloret et al. 1996).
Analysis of Collected Culture Medium
The concentration of hCG in the culture medium was assessed by a quantitative immunoradiometric assay using a commercially available kit (ICN Pharmaceuticals; Basingstoke, UK). This assay uses a “sandwich technique” in which the solid phase binds the α-subunit of hCG, while a radiolabeled antibody in the liquid phase binds to the corresponding β-subunit of the protein.
LDH in the culture medium was measured using an enzymatic activity assay for the catalyzed reduction of 1.6 mM pyruvate in Tris-buffer, pH 7.5. Enzyme activity was determined from the conversion rate of 0.2 mM NADH to NAD+ at 37C and was measured by the change in absorbance at 340 nm over a 3-min period.
Both LDH and hCG measurements were related to explant protein concentrations at the end of the 11-day cultures. Protein concentrations were measured using a Lowry-type assay. Explants were initially dissolved in 4 ml 0.3 M NaOH. Total protein was recorded using appropriate controls and Bio-Rad DC Protein Assay (Bio-Rad Laboratories; Hemel Hempstead, UK). Protein concentrations were taken from a standard curve using an optical density of 750 nm.
Apoptosis
Using the same random sampling technique, four explants of villous tissue were dissected, fixed in 10% buffered formalin, and embedded haphazardly in a single paraffin block. Sections of 5 μm were cut and mounted on microscope slides, stained with hematoxylin and eosin (H and E), and examined by light microscopy. Each sample was inspected at a magnification of ×400 oil immersion using a Leitz Dialux 22 microscope, with additional Image-Pro Plus 4.5 software (Media Cybernetics; Rockville, MD). In accordance with the protocol of Smith et al. (1998), 51 high-power fields of view (17 per section) were analyzed for each placenta. Each field was standardized using an E11–18mm diameter eyepiece graticule with a 1.0-mm pitch (Datasights; Middlesex, UK). All placental nuclei in each field of view, including those touching the grid parameter, were included. Cells inside the fetal capillary space were excluded. The morphological assessment of apoptosis was made in accordance with previous studies (Smith et al. 1997b). Apoptosis was recognized by the condensation of the heterochromatin within the nucleus. Unlike non-apoptotic nuclei, these condensed bodies show a constant density as the microscope is “racked” up and down (Smith et al. 1997b). The number of apoptotic nuclei was expressed as a percentage of total nuclei in the ST and non-ST compartments. Total nuclei were identified and counted by thresholding of the computerized monochrome image. Cluster analysis of ST nuclei was necessary to distinguish single events. Before total nuclei were counted, cluster measurements of five syncytial regions were verified by eye and thesholding adjusted accordingly.
Caspase Activity Assay
After removing the media and washing with PBS, four explants for each experimental time and condition were homogenized with 200 μl of homogenization buffer (10 mM HEPES, 2 mM EDTA, 0.1% CHAPS, 5 mM DTT, 10 μg/ml pepstatin A, 20 μg/ml leupeptin, 10 μg/ml aprotinin, 10% glycerol, pH 7.0) on ice using a glass tissue grinder (Fischer Scientific; Loughborough, UK). After centrifugation, total protein was recorded in the supernatant by Lowry assay and 40 μl of homogenate was added to 180 μl of reaction buffer (100 mM HEPES, 0.5 mM EDTA, 10% glycerol, 5 mM DTT, 50 μM cytochrome c, 10 mM dATP, pH 7.0) containing either caspase substrate (50 μM Ac-DEVD-amc (Calbiochem-Novabiochem UK; Nottingham, UK) and/or caspase inhibitor (25 μM Z-Vad-FMK; Calbiochem-Novabiochem UK). The samples were transferred to the wells of a 96 white-walled flat-bottomed plate and incubated in the dark for 30 min at 37C. The reaction was stopped by addition of 100 μl stop solution (1% sodium acetate, 175 mM acetic acid) and the fluorescence measured at 380/460 nm with a FL500 fluorimeter (Bio-Tek Instruments; Winooski, VT). Enzyme activity was expressed as fluorescence units per mg protein.
Morphology and Immunohistochemistry
Explants were fixed for 24 hr in fresh neutral buffered 10% formalin, embedded in either paraffin blocks or LKB Historesin, and serial sections of 5 and 1 μm cut, respectively. Resin-embedded tissue was stained with 0.5% toluidine blue and paraffin sections were rehydrated for peroxidase IHC. Microwave pretreatment with 0.01 M citrate buffer, pH 6.0, was performed to encourage epitope exposure, and endogenous peroxidase was blocked by 3% hydrogen peroxide. Mouse anti-human βhCG primary antibody (1:5000, Sigma Chemical) was used in combination with biotinylated secondary anti-mouse antibody (1:200; DakoCytomation, Glostrup, Denmark). Antibody binding was visualized with diaminobenzidine solution (Sigma Chemical). Tissue sections were viewed and photographed using the microscope and imaging software as described.
For electron microscopy, placental explants were harvested and fixed for 3 hr in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer, pH 7.3. Fixed tissue was then washed in buffer containing 3 mM calcium chloride and stored at 4C. For processing, the specimens were dissected and postfixed for 1 hr at 4C in 1% osmium tetroxide in 0.05 M sodium cacodylate buffer. After rinsing in the buffer, the tissue pieces were dehydrated in an ascending series of alcohols, incubated in two 15-min changes of propylene oxide, then infiltrated with Taab embedding resin (Taab Laboratories; Reading, UK) and polymerized in gelatin capsules at 60C for 72 hr. Sections 0.5 μm thick were cut, mounted on copper grids, stained with uranyl acetate and lead citrate, and examined by a Philips EM 301 electron microscope at an accelerating voltage of 60 kV.
Proliferation
The quantification of proliferation was performed as previously described for the Mib-1 staining of paraffin-embedded sections (Smith et al. 1998). Actively proliferating cells were identified as Mib-1-positive nuclei within the stromal, endothelial, or CT compartments. Proliferating nuclei were related to the total non-ST nuclei by image counting as described above.
Statistical Analysis
Unless otherwise stated, statistical significance of difference for normally distributed data was determined using either repeated measures for the ANOVA or Student's t-test for independent samples (with or without a Bonferroni correction for multiple comparisons). Normal distribution was assessed by the Shapiro-Wilk significance level normality test and results are presented as means ± SEM, with the data considered significant at p<0.05.
Results
Morphology
Examination of the explanted sections under all experimental conditions revealed a progressive degeneration of the original ST layers within the first 2 days of culture. Even after day 1 the ST layer appeared vacuolated and was lifting away to expose the underlying CT and basement membranes of the villus (Figure 1A). Although nuclei within this discarded syncytium showed signs of apoptotic morphology, the remaining placental villus and exposed CTs appeared relatively unaffected. The original syncytium, discarded in these early explant cultures, was gradually lost to the medium throughout the remaining 9 days of culture. By day 4, differentiation of exposed CTs could be easily defined by densely stained cytoplasmic regions (Figure 1B) and the development of new microvilli (Figure 2). However, unlike previous descriptions the extensive regeneration of STs from these CTs was not readily observed after exposure to 17% O2 (Watson et al. 1995), although areas of dense CTs did show signs of limited fusion (Figure 1B). By day 11 the cells of the mesenchyme showed signs of vacuolation and general disintegration. Before this time the overall preservation of the villus was good.
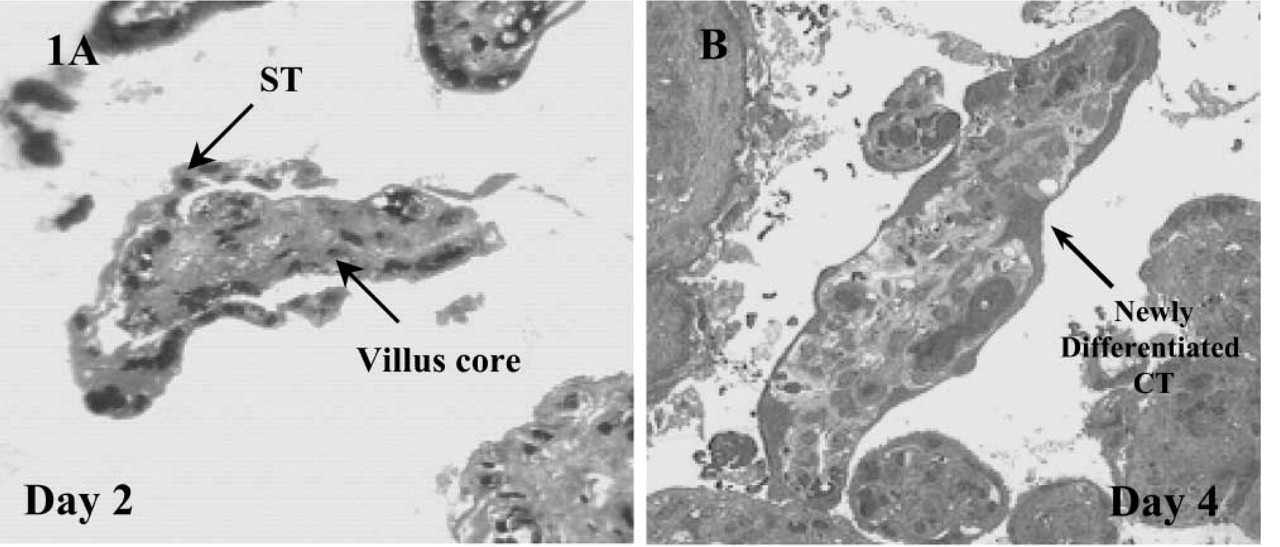
(
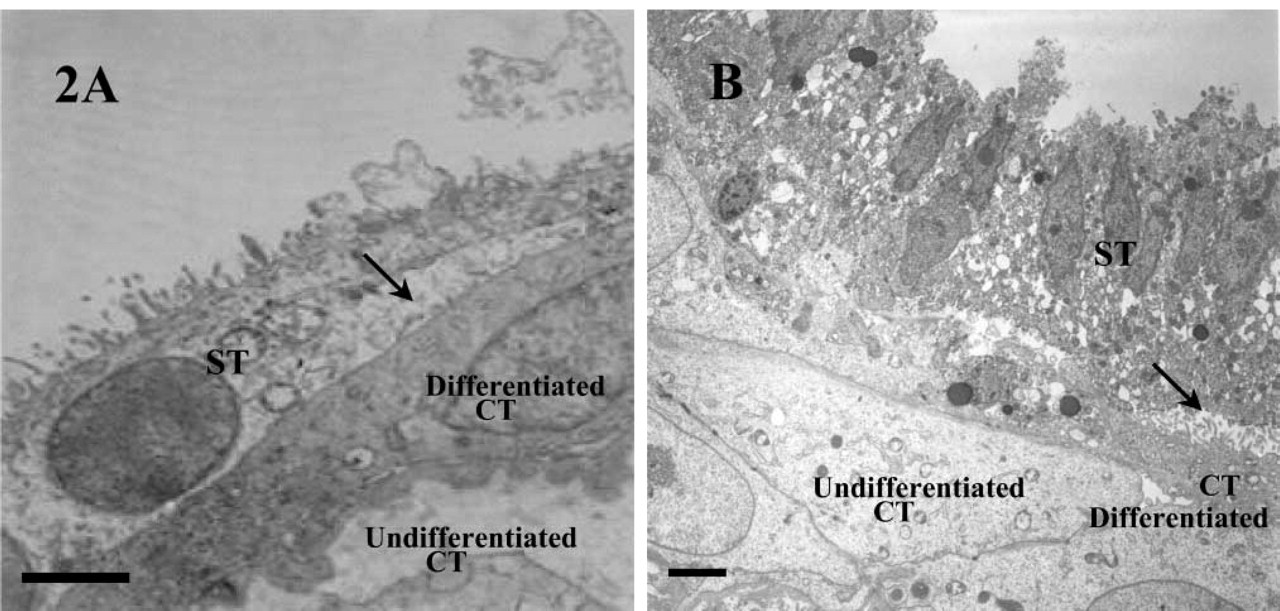
Electron micrographs of placental villous explants harvested on day 4 (
hCG Release from Cultured Placental Explants
In accordance with our morphological observations, the first 2 days of culture elicited a marked fall in ST-derived hCG under all experimental conditions tested (Figure 3). For explants maintained at 17% O2, both in the presence and the absence of TNFα, this loss in hCG quickly recovered from day 2 and increased exponentially thereafter. Given that the detached ST was devoid of antibody staining (Figure 4), this resurgence of hCG could be explained by the differentiation of CTs either from those formed in vitro or those exposed after ST breakdown. The production of hCG throughout the culture period was significantly lower in the TNFα-treated explants than in those without TNFα (p<0.01), suggesting a partial suppression in biochemical transition. Depleting oxygen to 3% abolished hCG production from day 2 in culture, suggesting a complete cessation in CT differentiation. Restoring explants to 17% O2 from day 2 onwards, after an initial depletion of oxygen for 48 hr, partially restored hCG production (Figure 5), indicating that oxygen sensitivity was reversible.
Lactate Dehydrogenase Release
LDH released into the media of the 17% O2 cultures was inversely related to hCG production in the absence of TNFα (Figure 6). The early reduction in hCG was associated with excessive cell death (as indicated by elevated LDH levels), while the subsequent period of differentiation, identified between days 2 and 7, was generally devoid of exaggerated tissue necrosis. By day 11, LDH levels in the 17% O2 cultures began to rise, again consistent with a secondary loss of hCG production and tissue integrity. TNFα initially inhibited the necrotic events of the early cultures. However, prolonged exposure amplified the secondary wave of decline between days 7 and 11. Of all the culture conditions, 3% O2 had the most marked effect on cell viability. Most notably, a reduction in oxygen maintained tissue viability throughout the entire culture period, even during the first few days in culture, when hCG production was rapidly declining.
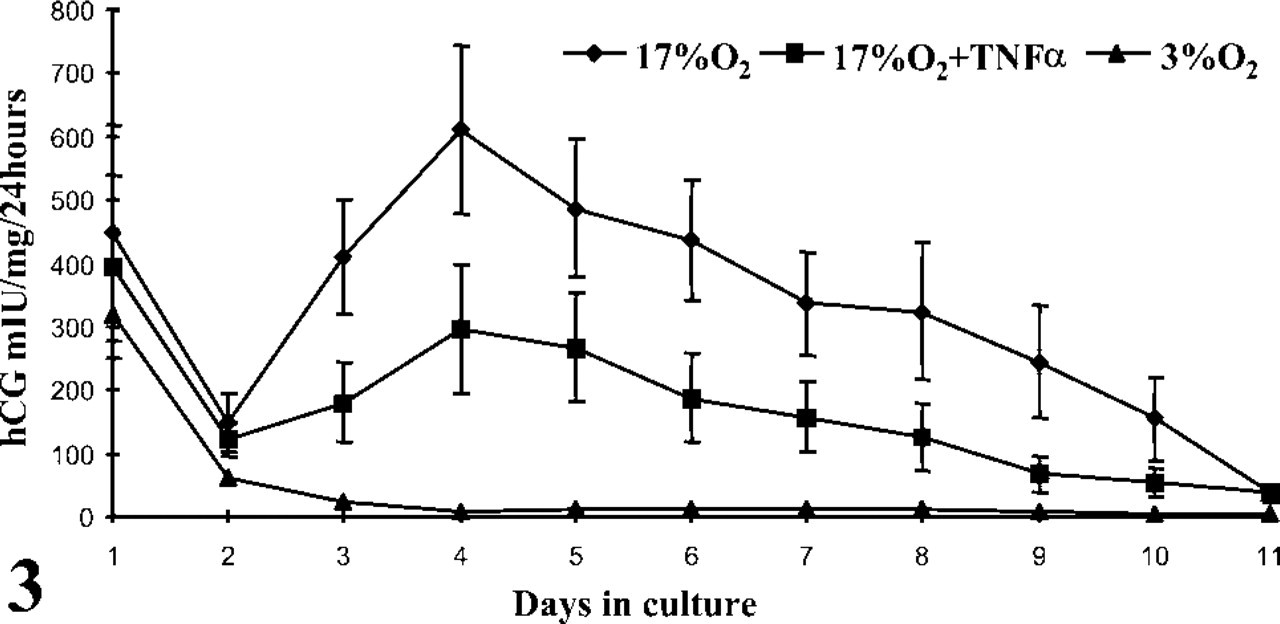
The profiles of hCG released from placental villous explants per 24 hr over 11 days in culture after incubations in 3% O2 and 17% O2 with or without TNFα. Data are means ± SEM for 14 placentas and analyzed by repeated measures ANOVA: 3% O2 vs 17% O2, p<0.001; 17% O2 vs 17% O2 + TNFα, p<0.01.
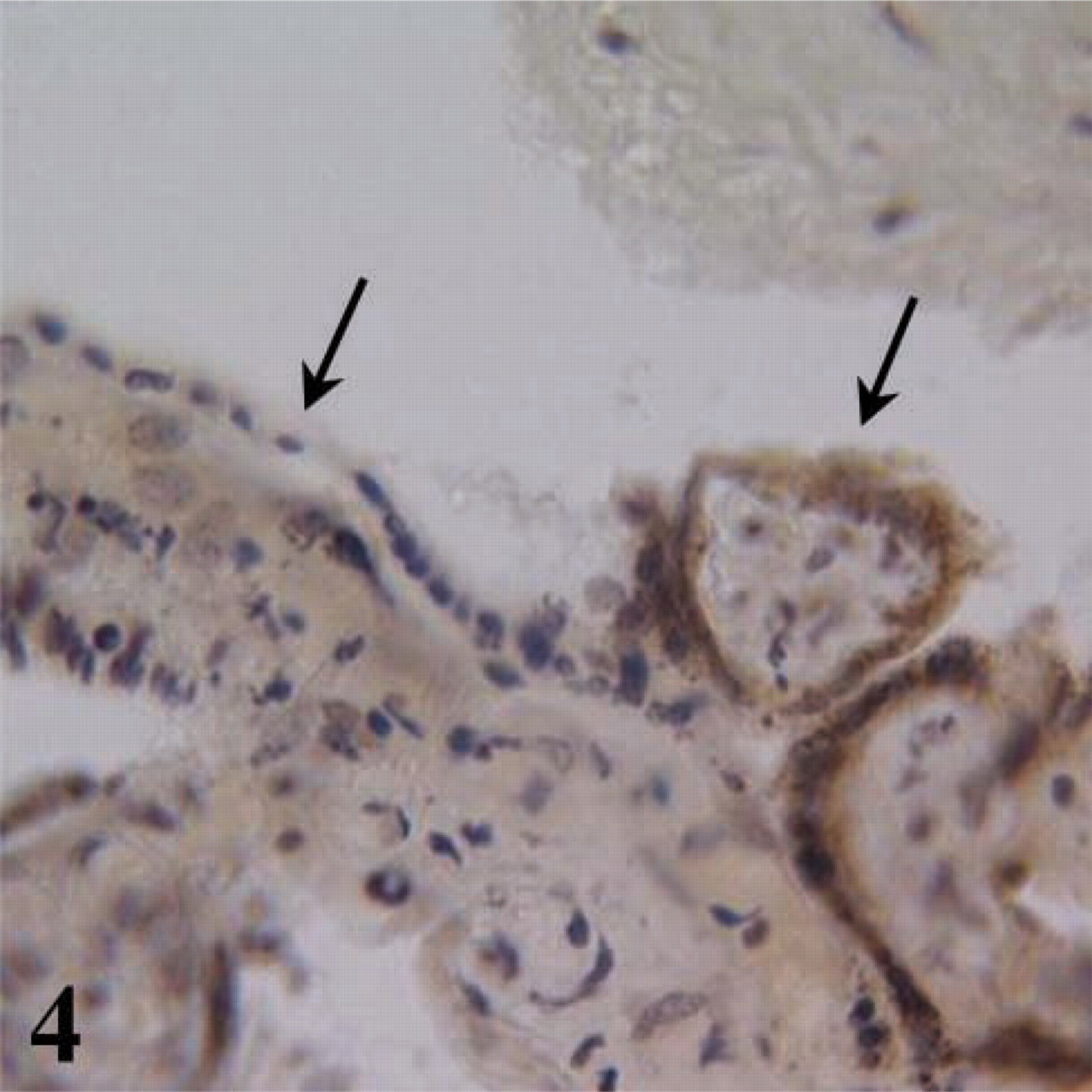
A tissue section of villous explant in 17% O2 at day 4 in culture showing hCG localized to the intact and attached syncytium. Left arrow highlights detached ST without hCG; right arrow indicates positively labeled (brown stained) attached regenerated ST.
Apoptosis
Before culturing, the incidence of apoptosis was greater in the non-ST regions of the explanted villi (Table 1), predominating in the stroma but not in the endothelia or CTs. For the explants cultured in 17% O2, early syncytial degradation was characterized by apoptosis, as determined by caspase activity (Figure 7) and apoptotic morphology (Table 1). Over the initial 2 days of culture, the presence of TNFα was associated with unaltered levels of apoptosis, either in the syncytium or the villous mesenchyme. However, prolonged exposure to TNFα eventually enhanced the apoptotic index in both the attached and detached syncytial compartments (Table 2). A reduction to 3% O2 oxygen had little effect on apoptosis because there were no significant differences between 3% and 17% O2.
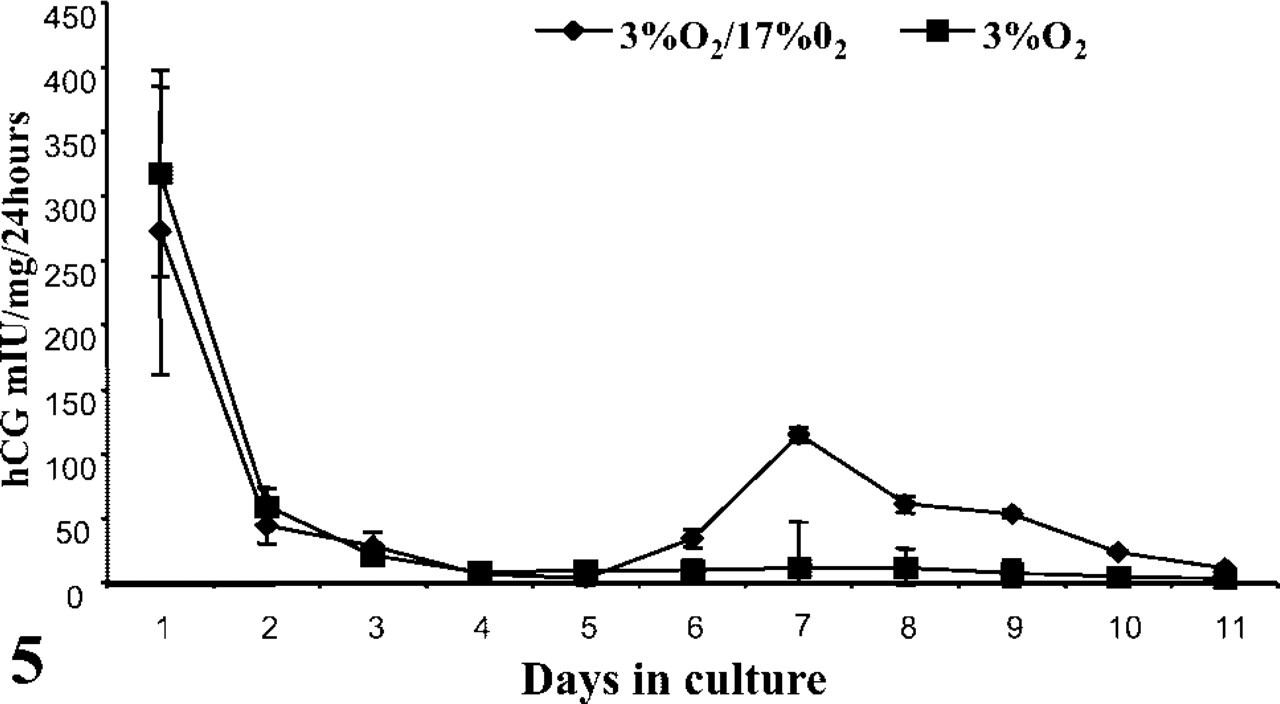
hCG released from placental villous explants in 3% O2 or 17% O2 after an initial incubation in 3% O2 for 48 hr. Data are means ± SEM for explant cultures of four individual placentas.
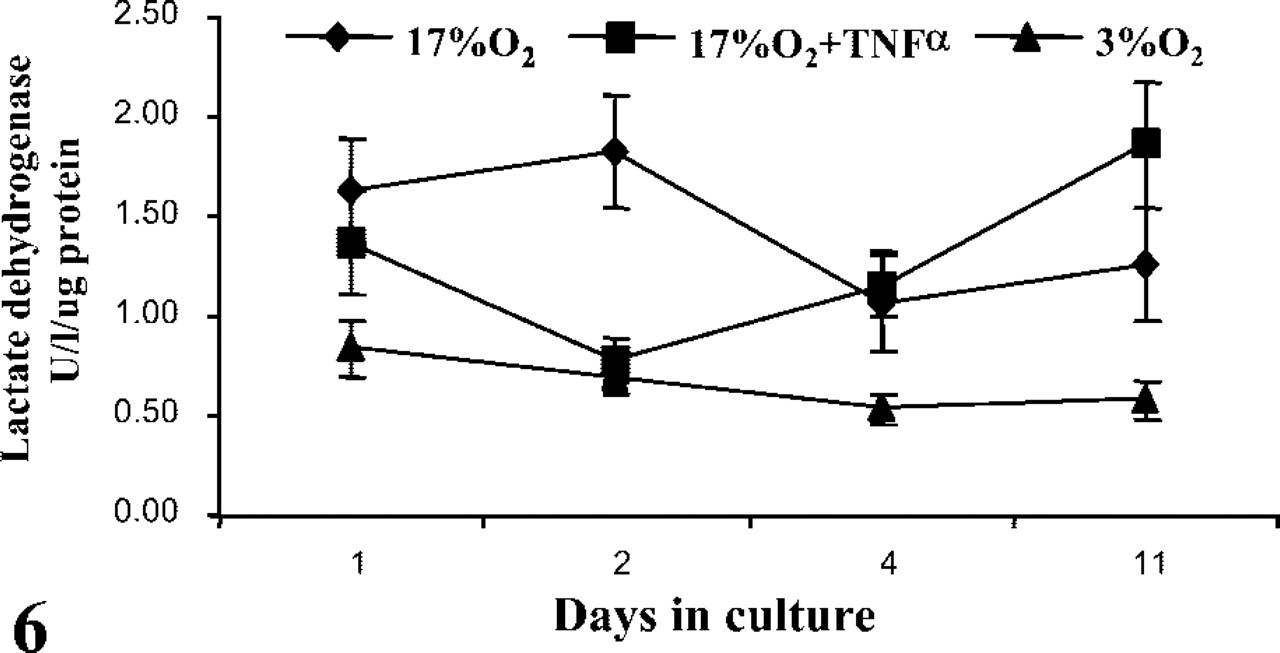
LDH released from placental villous explants at days 1, 2, 4, and 11 in culture, either in 3% O2 or 17% O2 with or without TNFα. Medium was changed daily. Data are means ± SEM for 14 placentas and analyzed by repeated measures ANOVA: 3% O2 vs 17% O2, p<0.0001; 17% O2 vs 17% O2 + TNFα, p<0.05.
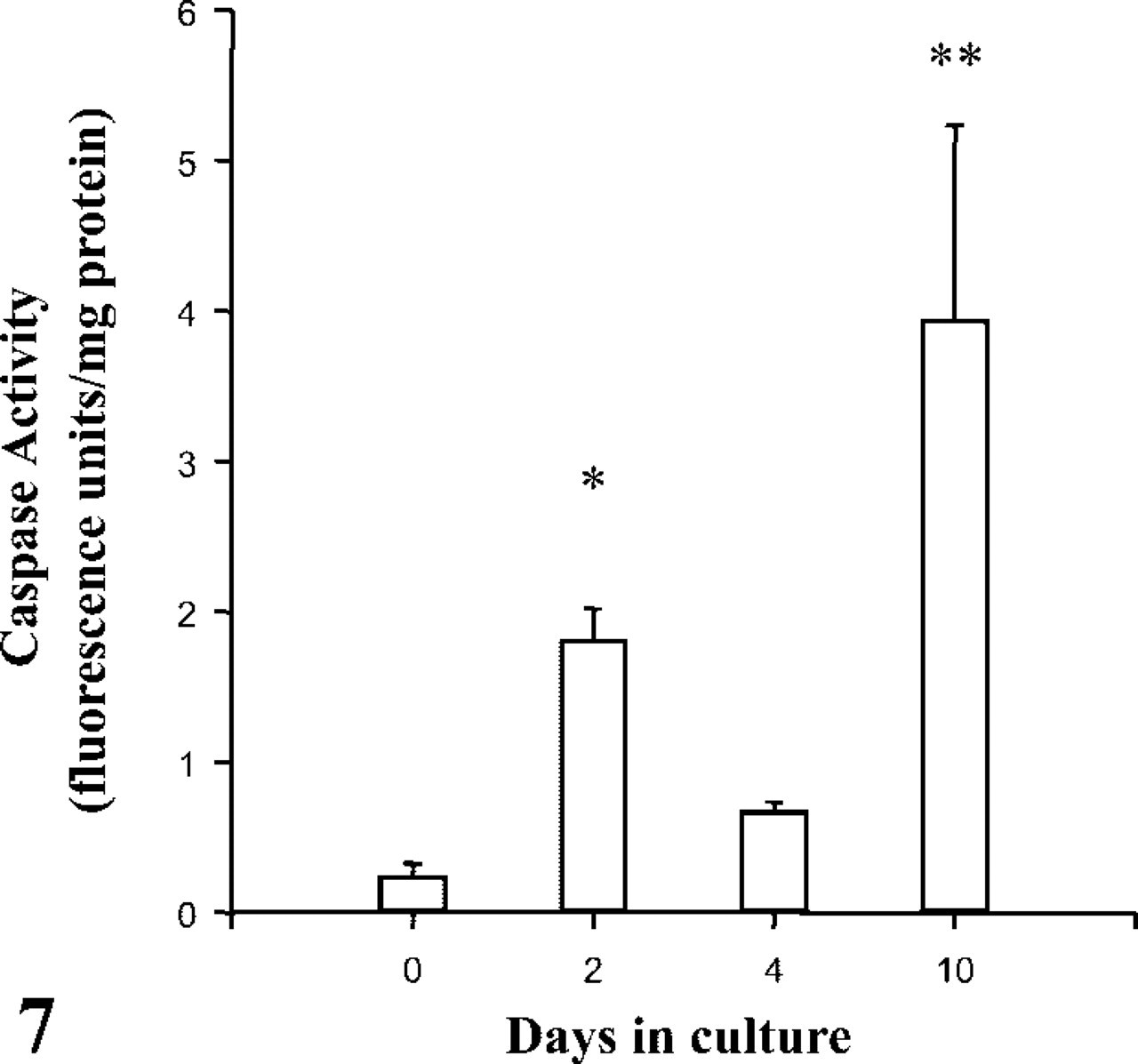
Caspase-3 activity in placental villous explants from normal pregnancies cultured in 17% O2. Data are means ± SEM for six placentas and analyzed by Student's t-test: ∗p<0.01, ∗∗p<0.001.
Proliferation
Before culturing, the proliferative index (i.e., Mib-1-positive events expressed as a percentage of total non-ST nuclei) was almost sixfold greater than that for apoptosis in the equivalent non-ST compartment (Tables 2 and 3). Throughout the culture period proliferation declined under 17% O2 both in the presence and absence of TNFα. From day 4 this reduction was significantly restored in 3% O2. Depleting oxygen to 3% enhanced the rate of proliferation by over threefold in the case of CT and almost ninefold in the stromal compartment (Table 3). Notably, CT proliferation was consistently greater than that of the stroma and proliferative staining of placental endothelial cells was not observed.
Discussion
This report supports previous findings with regard to the culture of term placental explants but extends these observations by monitoring cell kinetics and the influence of oxygen and TNFα. A major aim of this study was to investigate these explants as a model system for the assessment and monitoring of placental villous trophoblast cell turnover. The established life cycle of CT in vivo is one of continuous renewal, involving proliferation, differentiation, the extrusion of cells or cell fragments, and their removal by mechanisms that may or may not involve apoptosis. In normal pregnancy, CT turnover from fusion to syncytial knot shedding may take 2–4 weeks (Huppertz et al. 1998). From the evidence presented, we conclude that explant cultures do not fully replicate this process. However, many aspects, including those of proliferation, differentiation, and ST degradation (by necrosis and apoptosis), are evident and would therefore justify their use in the study of abnormal placental responses and conditions.
In agreement with earlier investigations (Siman et al. 2001; Hung et al. 2002), we have shown that placental villus fragments demonstrate ST degradation within the first 48 hr of culture (under all experimental conditions tested) and that this form of degeneration is characterized by necrosis and apoptosis within the rejected syncytia. Once the original ST has become detached, CT cells are then exposed. These cells rapidly undergo biochemical differentiation into an ST-like phenotype. In contrast to previous reports (Siman et al. 2001), we have been unable to identify extensive regeneration of new ST layers but have shown limited syncytialization in areas where the original CTs were concentrated. To a degree, these results may be explained by the scarcity of CT within the villous placenta at term.
The use of hCG secretion as a determinate of CT differentiation has been questioned in a previous study (Watson et al. 1995), which suggested that hCG generation was derived by membrane-damaging events and not by de novo synthesis. In these investigations we have presented evidence to refute the idea of passive hormone release, particularly with regard to the sustained elevation of hCG observed from day 2 in culture. Our data, in accordance with those of Siman et al. (2001), clearly show that LDH, a non-secreted cytosolic enzyme, does not emulate hCG release but in fact directly opposes it, particularly in cultures maintained at 17% O2. In addition, we have immunolocalized hCG to the attached and functional STs and have found no discernible evidence of hormone production within the lost and degenerate syncytia. These findings, in combination with data showing that hCG reductions can be reversed, uphold the use of hCG measurement and emphasize its usefulness as a tool in the monitoring of ST viability and CT transformations.
Apoptotic index of syncytial and nonsyncytial compartments of placental villous explants cultured in 3% O2 or 17% O2 in the presence and absence of TNFα a
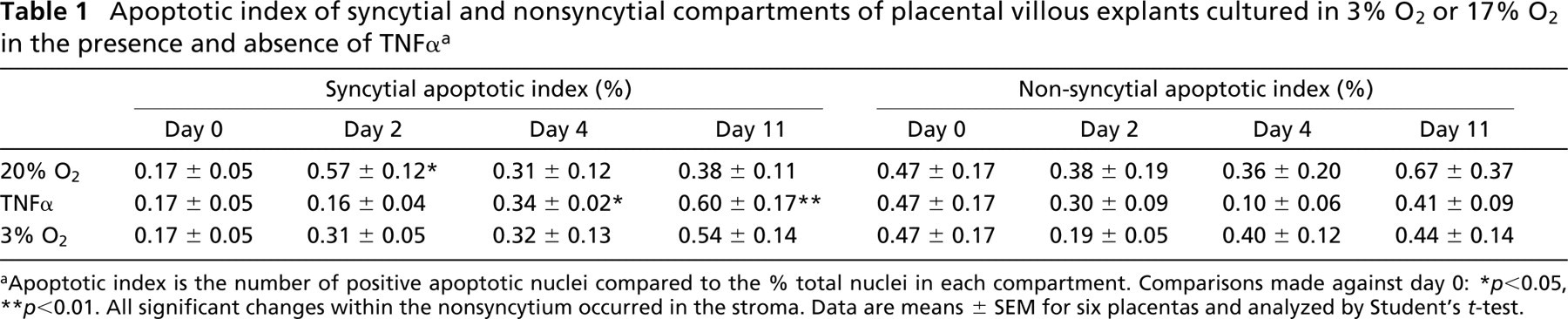
Apoptotic index is the number of positive apoptotic nuclei compared to the % total nuclei in each compartment. Comparisons made against day 0:
p<0.05
p<0.01. All significant changes within the nonsyncytium occurred in the stroma. Data are means ± SEM for six placentas and analyzed by Student's t-test.
Like the placenta, explanted villous fragments can support CT mitosis, as evident with Mib-1 immunostaining, but these replications were limited, particularly at higher concentrations of oxygen. Instead of proliferation, the majority of CTs underwent rapid differentiation at 17% O2 after exposure through ST degeneration. As a rule, the fate of these newly transformed cells did not replicate those of postmitotic CTs in vivo, because syncytial knot formation was never truly defined. However, syncytial and non-syncytial nuclei did show clear and measurable signs of biochemical and morphological apoptosis, a finding that supports a role for explant cultures in the assessment of CT and ST cell death. Unfortunately the depletion of syncytium was never accompanied by fibrinoid deposition, a common feature of the disrupted and denuded villus in vivo (Mayhew and Barker 2001). Nevertheless, these micrographs do show signs of re-epithelization, in which the re-formation of syncytium occurs over the underlying basement membrane or CT in a process akin to that of epithelial repair, as observed extensively ex vivo (Nelson et al. 1990).
Total apoptosis and total proliferation of placental villous explants cultured in 3% O2 or 17% O2 in the presence and absence of TNFα a
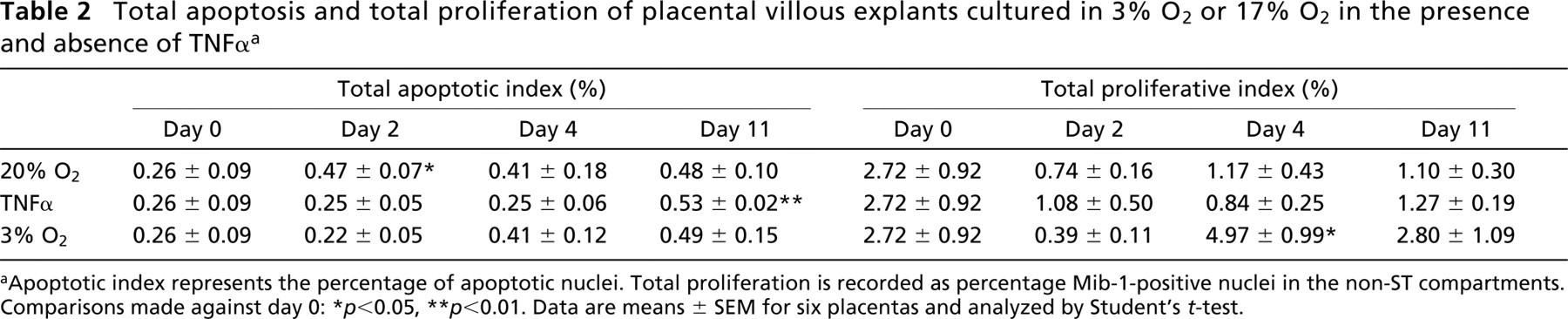
Apoptotic index represents the percentage of apoptotic nuclei. Total proliferation is recorded as percentage Mib-1-positive nuclei in the non-ST compartments. Comparisons made against day 0:
p<0.05
p<0.01. Data are means ± SEM for six placentas and analyzed by Student's t-test.
Compared with normal pregnancy, it has been suggested that the steady state between the numerical ratios of CTs and STs is re-set in PE and IUGR (Mayhew et al. 2003). Because these conditions are characterized by reduced uteroplacental blood flow in mid-pregnancy, its is not surprising that histological features support the idea of placental hypoxia as a possible initiating force (Chappell and Bewley 1998). In our explant experiments, we have exposed placental fragments to two differing oxygen tensions, one of relatively high oxygen, which would be unexpected in the uteroplacental environment, and one of low oxygen, which may be typical of or lower than that usually associated with the placenta (Fujikura and Yoshida 1996). In accordance with recent observations (Huppertz et al. 2003), we have shown that low oxygen, i.e., conditions that may restrict ATP synthesis, have confined CTs to processes of cell division rather than those of differentiation or cell death. These observations correlate with an increase in trophoblastic epithelium, as previously described in IUGR (Mayhew et al. 2003).
Inflammatory cytokines, particularly TNFα, may represent a significant risk factor in PE (Serin et al. 2002); a maternal situation of recognized systemic inflammation (Redman et al. 1999). In this condition, increased apoptosis and shedding of placental material into the maternal circulation is believed to occur (Leung et al. 2001). This, in turn, triggers a general inflammatory cascade (Redman and Sargent 2001). The exact form of this discharged material is as yet unknown, and the mechanism by which it is derived, i.e., apoptosis or necrosis, is currently undetermined. However, TNFα is elevated in the placenta in PE (Conrad and Benyo 1997). Moreover, in isolated CTs and STs it has been shown to elicit a significant apoptotic response (Crocker et al. 2001). In our explant cultures, bioactive levels of TNFα (in combination with IFN-γ) encouraged elevated apoptosis and exaggerated cell death in the ST compartments. Obviously, apoptosis within the natural syncytium is more beneficial than necrosis, but it may still be the case that the overall burden of placental debris lost to the maternal circulation is the trigger for systemic toxemia, as suggested by Redman and Sargent (2000) in PE.
The separate proliferative indices of CT and stromal compartments of the placental explant villi cultured in 3% O2 or 17% O2 in the presence and absence of TNFα a
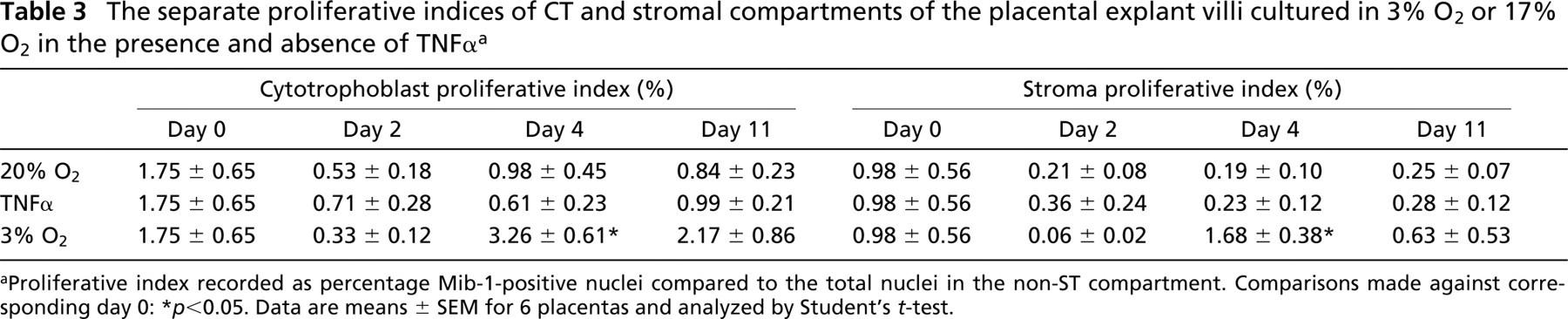
Proliferative index recorded as percentage Mib-1-positive nuclei compared to the total nuclei in the non-ST compartment. Comparisons made against corresponding day 0:
p<0.05. Data are means ± SEM for 6 placentas and analyzed by Student's t-test.
Undoubtedly, IUGR and PE are associated with substantial alterations in placental morphology and are likely to reduce placental efficiency and to contribute to both fetal distress and maternal immune activity (Smith et al. 1997a; Mayhew et al. 2003). The use of explants to identify inappropriate cell kinetics has its limitations, but certain aspects of placental cell turnover are replicated and can be used to reproduce placental conditions and investigate their influence on placental responses in vitro. The data generated should be interpreted in two ways. First, explants are exquisitely sensitive to culture conditions, including oxygen and TNFα. Second, these conditions can be manipulated to investigate different placental issues of interest. For example, by altering the oxygen, the processes of degeneration and repair in the villous placenta can be studied, i.e., at 17% O2. In contrast, by changing to 3% O2, the proliferation of constituent cells can be exaggerated, again for further investigation. This one simple model therefore holds considerable benefits over isolated cells of the villus because these cells are essentially nonproliferative and have produced highly conflicting results, at least with respect to CT turnover in vitro (Levy et al. 2000; Crocker et al. 2001; Kilani et al. 2003).
Overall, these investigations advocate the use of hCG as a marker of syncytial events within the explant model. It is envisaged that further studies will generate important information regarding the pathways of cell turnover within the placental villus and the adaptive or adverse influence of external factors in the etiology and propagation of placenta-associated conditions such as PE and IUGR.
Footnotes
Acknowledgements
Supported by Tommy's-The Baby Charity and by the Castang Trust.