
Abstract
The present study was designed to examine the postnatal developmental changes of atypically shaped cardiomyocytes (ACMs) prepared from the heart of newborn [postnatal day 1 (day-1)] through aged (12-month-old) mice. ACMs were identified as a novel type of self-beating cardiomyocyte with a peculiar morphology in mouse cardiac ventricles. The cell length of ACMs significantly increased during the first three postnatal months and further increased over the following 9 months. In contrast, the population of ACMs was significantly decreased within the first 5 weeks and reached a plateau in the adult stage. ACMs obtained from newborn and adult mice exhibited similar spontaneous action potentials. The expression of the fetal cardiac gene products atrial natriuretic peptide and voltage-gated T-type Ca2+ channel Cav3.2 was confirmed by immunostaining in ACMs obtained from both newborn and aged mice. These observations provide evidence that ACMs that exhibit spontaneous beating survive the long-term postnatal development of cardiac ventricles while preserving the expression of fetal cardiac genes. This manuscript contains online supplemental material at http://www.jhc.org. Please visit this article online to view these materials.
Keywords
T
There is a novel type of resident heart cell in adult mouse cardiac ventricles that spontaneously develops into self-beating cardiomyocytes with a peculiar morphology, defined as atypically shaped cardiomyocytes (ACMs) (Omatsu-Kanbe and Matsuura 2009). ACMs that emerge from the culture of cardiac myocyte–depleted fraction (CMDF) cells appear to be distinct from cardiomyocytes, side population cells (Martin et al. 2004; Oyama et al. 2007), or cardiac progenitor cells expressing stem cell marker proteins, such as stem cell antigen-1 (Sca-1) (Oh et al. 2003; Matsuura et al. 2004), stem cell factor receptor (c-kit) (Beltrami et al. 2003; Pouly et al. 2008), and LIM-homeobox transcription factor islet 1 (islet-1) (Laugwitz et al. 2005; Moretti et al. 2006).
The aim of the present study was to examine the postnatal development of ACMs in the mouse heart. The results indicate that self-beating ACMs appear to increase in size but decrease in number with age, and also survive a long-term postnatal development while preserving the expression of the fetal cardiac gene products atrial natriuretic peptide (ANP) and α1
Materials and Methods
Animals
C57BL/6J mice (Charles River Japan, Inc.; Yokohama, Japan) were used for these experiments. All animals were fed ad libitum in 12:12–hr light-dark cycles, and suckling infants were housed with their mothers. All animal experiments were performed in accordance with the guidelines of the institution's Animal Care and Use Committee.
Solutions
Tyrode solution was used as a standard physiological solution, containing (in mM) 140 NaCl, 5.4 KCl, 1.8 CaCl2, 0.5 MgCl2, 0.33 NaH2PO4, 5.5 glucose, and 5.0 HEPES (pH adjusted to 7.4 with NaOH). Cell isolation buffer (CIB) contained (in mM) 130 NaCl, 5.4 KCl, 0.5 MgCl2, 0.33 NaH2PO4, 22 glucose, 50 μ/ml bovine insulin, and 25 HEPES (pH adjusted to 7.4 with NaOH).
Preparation of CMDF Cells and Ventricular Myocytes
The animals were deeply anesthetized by intraperitoneal injection of a mixture of sodium pentobarbital (100 mg/kg) and heparin (8000 U/kg). CMDF cells were prepared from mice of at least postnatal day 8 (day-8) according to the method described previously (Omatsu-Kanbe and Matsuura 2009). Briefly, the heart was retrogradely perfused at 37C for 10 min with enzyme solution containing 0.1% collagenase, 0.006% trypsin, and 0.006% protease in CIB supplemented with 0.3 mM CaCl2, and then both ventricles were cut out at the level of 2/3 from the ventricular apex. CMDF cells were prepared from these atrial tissue–free ventricles, resuspended by vortexing in a semisolid culture medium containing methylcellulose-based medium MethoCult M3231 (StemCell Technologies; Vancouver, Canada) and Iscove's-modified Dulbecco's medium (StemCell Technologies) at a ratio of 80:20 in the absence of antibiotics, and plated in 35-mm plastic culture dishes or high-quality plastic dishes (μ-Dishes; ibidi GmbH, Martinsried, Germany).
Both ventricles were carefully cut out of newborn (day-1) mice at the level of 2/3 from the ventricular apex under a microscope. These atrial tissue–free day-1 ventricles were minced and digested in the enzyme solution at 37C for 50 min with pipetting. The reaction mixture was then washed with centrifugation, and CMDF cells were prepared with the same procedure used for day-8 or older mice.
Single ventricular myocytes were enzymatically dissociated from the hearts of day-2 and adult (13-week-old) mice using a retrograde Langendorff perfusion method (Shioya 2007).
Microscopy
Phase contrast images of ACMs were acquired using an inverted DIAPHOT 300 microscope (Nikon; Tokyo, Japan) and a charge-coupled device RETIGA 2000R camera (QImaging; British Columbia, Canada), and analyzed using Image Pro Plus software (Media Cybernetics, Inc.; Bethesda, MD). Sample size of each acquired image was 1600 × 1200 pixels (588 × 442 μm with a x20 objective).
Cell Length and Population
Phase contrast images of ACMs were analyzed in the computer using the Image Pro Plus software package to measure cell length. Beating ACMs were counted under a Hoffman-modulated inverted microscope to determine the number of cells.
Immunostaining
The cells cultured on μ-Dishes were fixed and immunostained as described previously (Omatsu-Kanbe and Matsuura 2009). The primary antibodies were mouse monoclonal anti–α-actinin (A7811, clone EA-53, 1:400; Sigma, St. Louis, MO) and anti–connexin-43 (C8093, clone CXN-6, 1:4000; Sigma), rabbit polyclonal anti-ANP (AB5490,1:200; Chemicon-Millipore, Billerica, MA), and anti–α1
Membrane Potential Recordings
The membrane potential of self-beating ACMs adhering to the bottom of 35-mm plastic culture dishes was measured in the current-clamp mode of the EPC-8 patch-clamp amplifier (HEKA; Lambrecht, Germany) immediately after establishing whole-cell access. The patch electrode had a resistance of 2.5–3.5 MΩ when filled with the pipette solution containing (in mM) 70 potassium aspartate, 40 KCl, 10 KH2PO4, 1 MgSO4, 3 Na2-ATP, 0.1 Li2-GTP, and 5 HEPES (pH adjusted to 7.2 with KOH), and the cells were continuously super-fused at a flow rate of 2.5–3 ml/min with Tyrode solution at 36–37C.
Statistics
All of the averaged data are expressed as the means ± SEM. Statistical comparisons were made using oneway ANOVA followed by Tukey's test, and differences were considered to be significant at p<0.001.
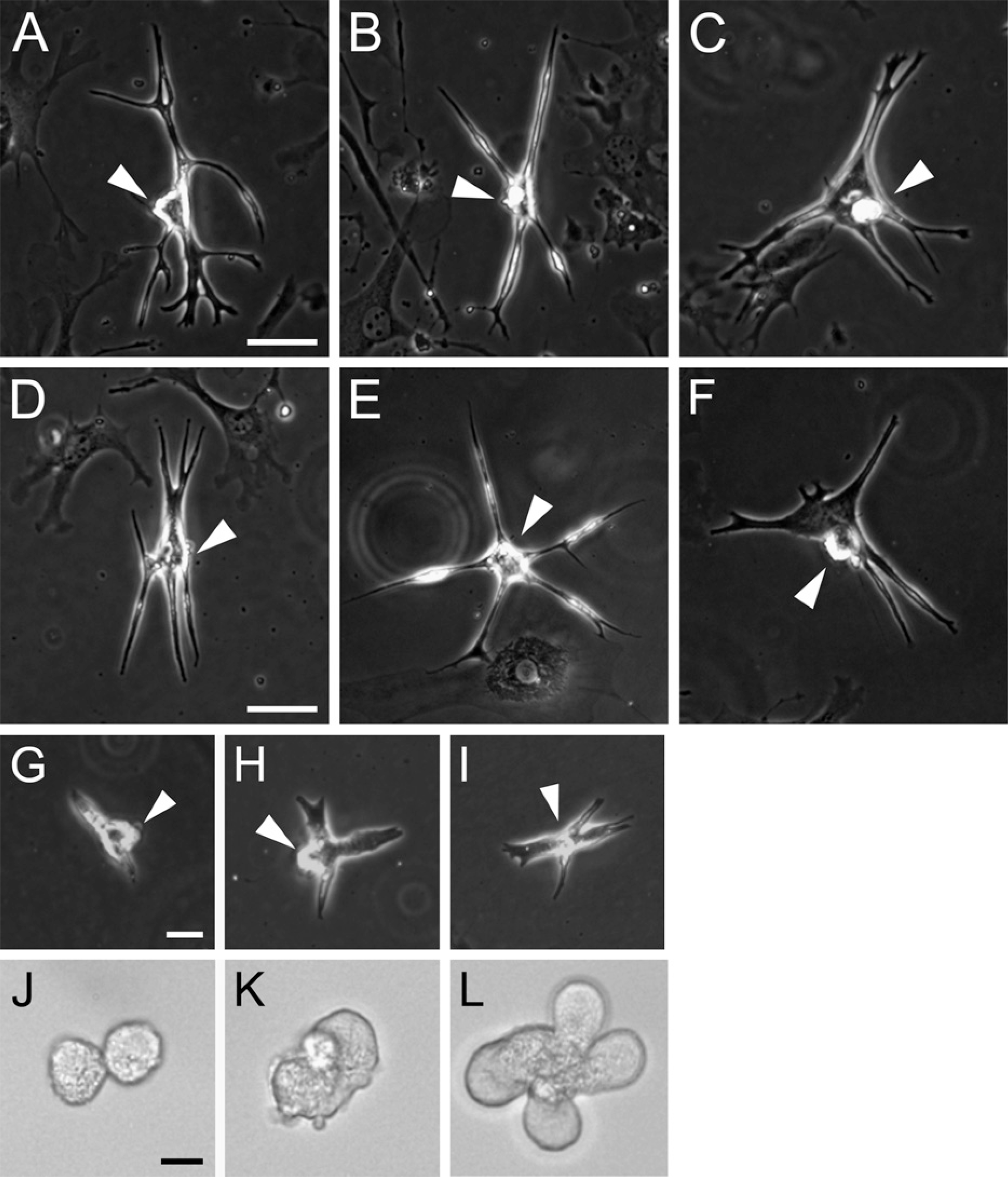
Atypically shaped cardiomyocytes (ACMs) prepared from adult and newborn mice. Phase contrast microscopy of self-beating ACMs from 15-week-old C57BL/6J (
Results
Morphological Characteristics of Self-beating ACMs in Day-1 Heart
Figures 1A–1C demonstrate that the ACMs obtained from adult C57BL/6J mice display a peculiar morphology with branching, projections, and/or bulges, as previously described (Omatsu-Kanbe and Matsuura 2009). To examine whether those ACMs were also resident in the hearts of other mouse strains, CMDF cells were obtained from adult ICR mice (Charles River Japan, Inc.) and cultured in a semisolid methylcellulose-based culture medium. As demonstrated in Figures 1D–1F, self-beating ACMs emerged from the CMDF cell culture of ICR mice, indicating that ACMs were not unique to the C57BL/6J mouse strain. The present study investigated the postnatal developmental changes in ACMs using C57BL/6J newborn to 12-month-old mice.
Some CMDF cells prepared from day-1 mice spontaneously developed into self-beating ACMs with a peculiar morphology, displaying branched and elongated shapes with projections, and often possessing small bulges on the cell surface (Figures 1G–1I), similar to the morphologies observed in ACMs obtained from adults (Figures 1A–1C). Although the bulges on ACMs from day-1 hearts and those from adult hearts were similar in size (diameter 5–10 μm), the ranges of elongation and the numbers of branching and projections observed in day-1 ACMs were somewhat less compared with those in adult cell preparations, giving newborn ACMs an “infantile” appearance. Although most of the day-1 ACMs were beating independently, in some cases, two or more ACMs that were connected at the edge of the cell membrane were beating synchronously (Movie SM1).
Interestingly, some of the self-beating ACMs obtained from day-1 heart displayed rounded shapes (diameter, ×10 μm) without any projections (Figures 1J–1L and Movie SM2). Such rounded beating cells were not observed in cell preparations obtained from the adult heart (Omatsu-Kanbe and Matsuura 2009). These “morphologically immature” types of ACMs were mostly found to have a fused appearance consisting of various numbers of rounded cells [e.g., two (Figure 1J), three (Figure 1K and Movie SM2), and five (Figure 1L) cells], and all of the fused cells were beating synchronously.
Repetitive Action Potentials Recorded in Day-1 ACMs
The electrophysiological basis for spontaneous beating of ACMs from day-1 mice was examined by recording the membrane potentials using whole-cell current-clamp techniques. As demonstrated in Figure 2, action potentials were spontaneously evoked by the preceding slow diastolic depolarization (pacemaker depolarization). The parameters of the spontaneous action potentials are summarized in Table 1 and are found to be somewhat similar to those of mammalian sinoatrial (SA) node cells (Mangoni and Nargeot 2001). It is thus likely that repetitive firing of action potentials underlies the spontaneous beating of ACMs from day-1 mice, similar to those observed in ACMs from adults (Omatsu-Kanbe and Matsuura 2009).
Postnatal Increase in Cell Dimension of ACMs
It is generally thought that cardiomyocytes increase in cell size, length, width, and volume during normal development (Katzberg et al. 1977; Korecky and Rakusan 1978; Anversa et al. 1986). We therefore examined the cell length of ACMs obtained from newborn to aged mice. As demonstrated in Figure 3A, the cell length of ACMs significantly increased during the postnatal ×3 months from 68.1 ± 3.1 μm (day-1) to 200.4 ± 5.74 μm (8–12-week-old), and further increased in the following ×9 months to 239.3 ± 9.5 μm (12–month-old).
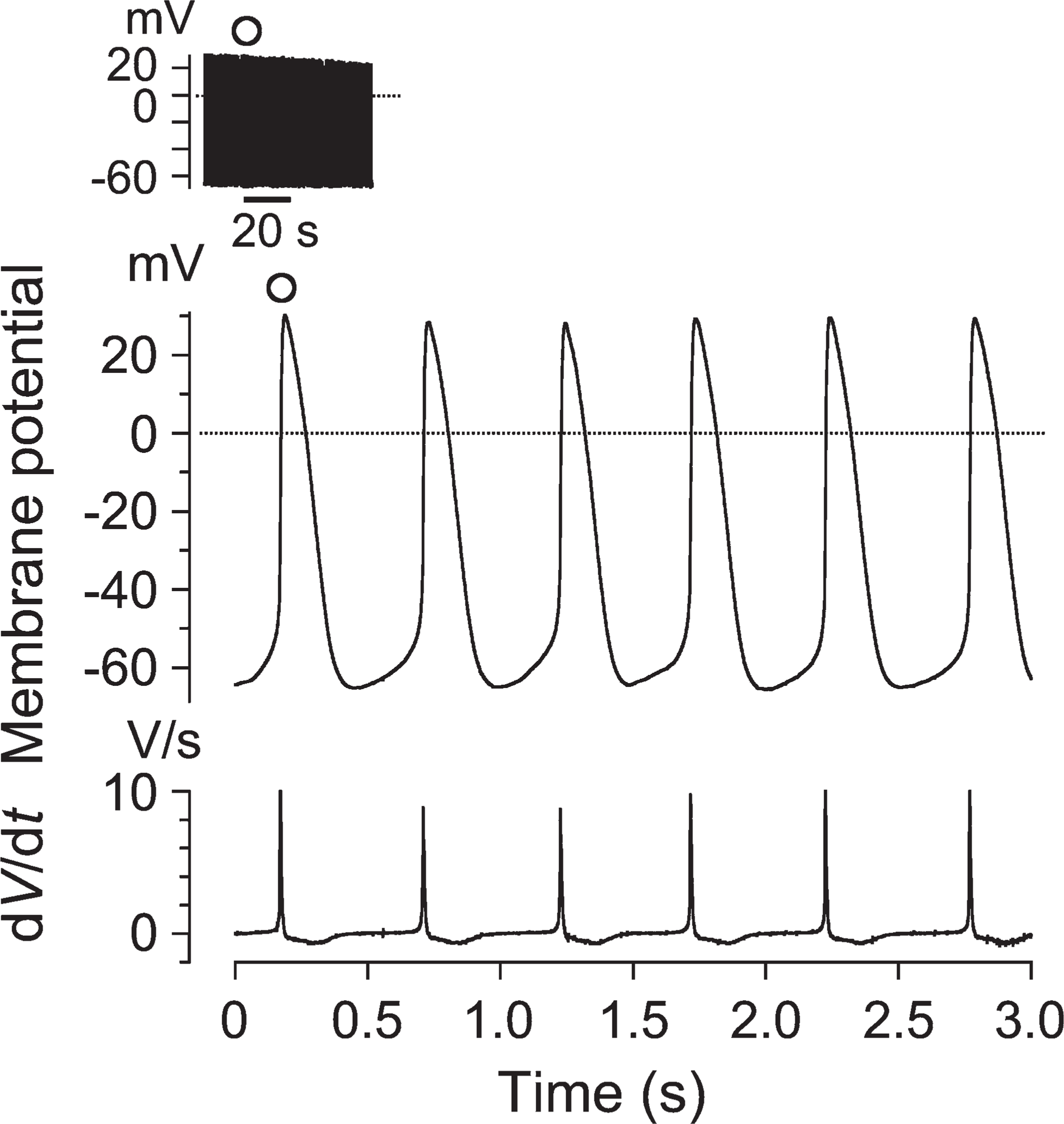
Action potentials recorded from day-1 ACMs. Chart record of the action potentials recorded from an ACM (left shoulder) and expanded traces (lower) at the time denoted by the open circle on a faster time scale. Lower top indicates the repetitive action potentials and lower bottom the first derivative of membrane potentials (dV/dt). Dotted line in the membrane potential records indicates the zero-voltage level.
Postnatal Decrease in Cell Population of ACMs
We next investigated the changes in the cell population of self-beating ACMs obtained from the hearts of mice of different ages. As demonstrated in Figure 3B, the number of ACMs was apparently highest at day-1, 4190 ± 163. However, it is possible that the cell numbers of day-1 ACMs were overestimated, because neonatal cardiomyocytes prepared by the tissue-mincing method have been reported to spontaneously beat in culture (Simpson and Savion 1982; Nuss and Marban 1994). In contrast, morphologically distinguishing ACMs from ventricular myocytes was much easier in cells of day-8 or older mice prepared by the retrograde perfusion method. Therefore the changes in the numbers of ACMs obtained from day-8 mice, 3219 ± 166, were compared with those obtained from the older mice. The population of ACMs significantly decreased to 823 ± 79 at 5-week-old (26% that of day-8 mice), whereas there were no significant differences in the numbers of ACMs in the following period, e.g., 558 ± 50 at 10-week-old (17% that of day-8 mice) and 613 ± 76 at 12-month-old (19% that of day-8 mice).
Action potential parameters of ACMs from day-1 mice

ACMs, atypically shaped cardiomyocytes; MDP, maximal diastolic potential; DDR, diastolic depolarization rate; SCL, spontaneous cycle length; AP, action potential.
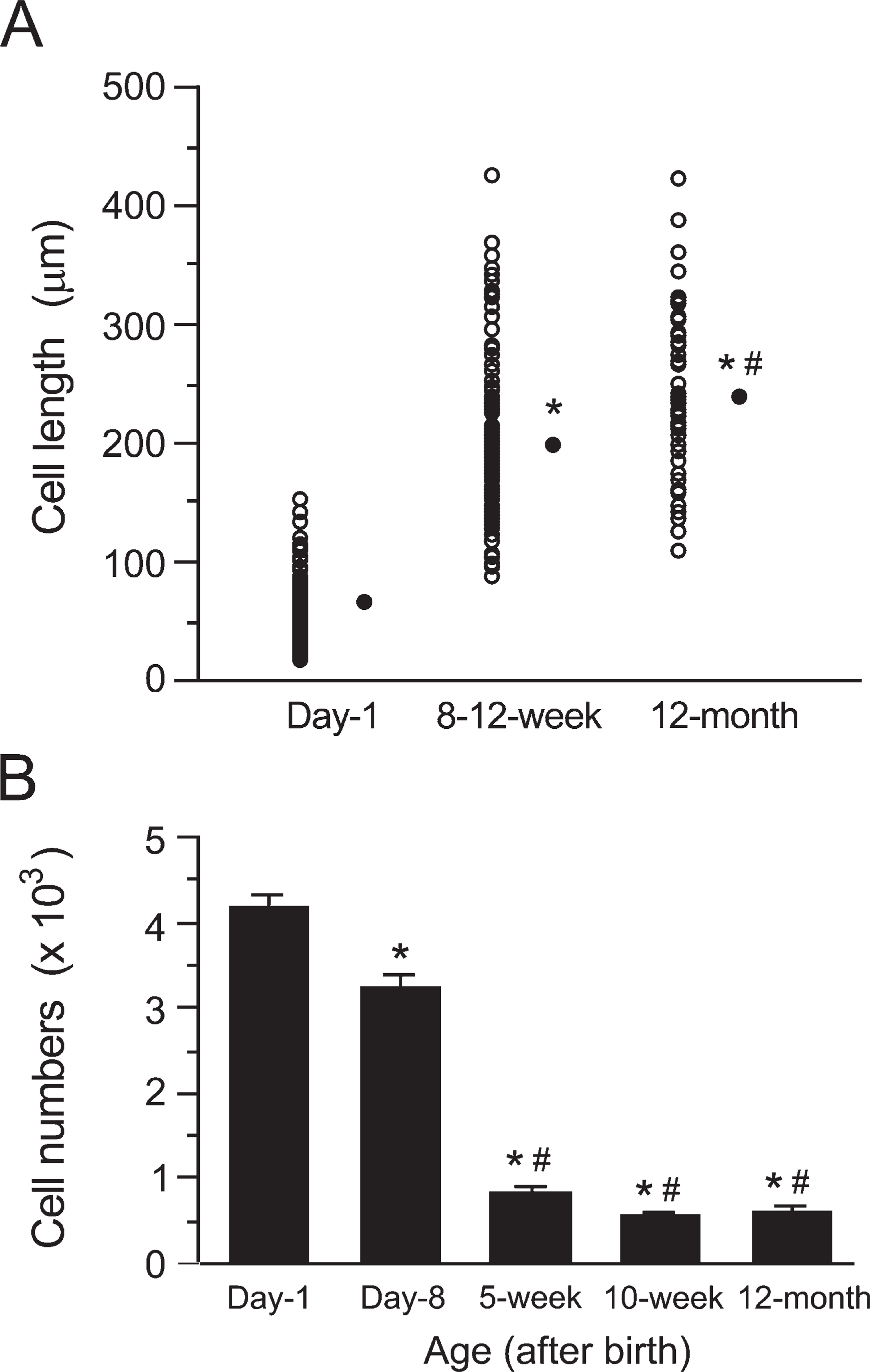
Postnatal changes in cell length and number of ACMs. (
Expression of Fetal Cardiac Gene Products ANP and Cav 3.2 in ACMs
One of the most interesting observations about ACMs obtained from adult hearts was that those cells exhibited characteristics of both SA node pacemaker cells and ventricular myocytes (Omatsu-Kanbe and Matsuura 2009), namely, self-beating activities and repetitive action potentials specific for the SA node, and the expression of connexin-43, the major isoform of gap junction proteins in ventricular myocytes (Verheijck et al. 2001; Musa et al. 2002). Immunocytochemical analyses confirmed the abundant expression of connexin-43 in ACMs from both day-1 (Figures 4A–4C) and aged [12-month-old (Figures 4D–4F)] hearts, displaying punctate staining throughout the whole-cell images. On the other hand, the expression of connexin-43 in isolated ventricular myocytes was mainly observed at the cell periphery, as expected (Figures 4G–4I).
We next examined whether ACMs express fetal cardiac gene products. It is well known that ANP is present in fetal ventricles but not in adult ventricles (Bloch et al. 1986; Cameron and Ellmers 2003). Figure 5 demonstrates that the expression of ANP was confirmed by immunostaining in ACMs from both day-1 (Figures 5A–5C) and aged (Figures 5D–5F) hearts, but was negligible in adult ventricular myocytes (Figures 5G–5I). Immunocytochemical analyses of α-actinin confirmed that day-1 ACMs, as well as aged ACMs, already exhibited well-organized sarcomeric structures (Figures 5 and 6), similar to the sarcomeric appearances observed in isolated ventricular myocytes from day-2 mice (Figures 5J and 5K).
Cav3.2 is also preferentially expressed in fetal ventricles, but less so in adult ventricles, and therefore the T-type Ca2+ current is very small or even undetectable in adult ventricular myocytes (Perez-Reyes 2003; Chiang et al. 2009). ACMs from both day-1 (Figures 6A–6C) and aged (Figures 6D–6F) hearts also expressed Cav3.2 predominantly in the intracellular space rather than the cell periphery, whereas very weak signals were detected near the plasma membrane in adult cardiac ventricular myocytes (Figures 6G–6I). These results show those fetal cardiac genes to be well preserved in ACMs during postnatal development.
Discussion
The present study provides evidence that self-beating ACMs obtained from mouse hearts increase in size but decrease in number during postnatal development, while preserving the expression of the fetal cardiac gene products ANP and Cav3.2 until the aged stage. We have previously demonstrated that self-beating ACMs develop spontaneously from rounded CMDF cells with a diameter of ×10 μm obtained from adult mouse ventricles (Omatsu-Kanbe and Matsuura 2009). ACMs are distinct from SA or atrioventricular nodal cells, because CMDF cells are prepared from cardiac ventricles containing no atrial tissues. Furthermore, ACMs are thought to be different from Purkinje cells: Purkinje cells exhibit a ventricular myocyte–like appearance (Han et al. 2002; Di Maio et al. 2007), whereas ACMs display a peculiar morphology with highly branched and elongated shapes with many projections. Taken together, ACMs can be identified as a novel type of heart cell that is present in adult cardiac ventricles.
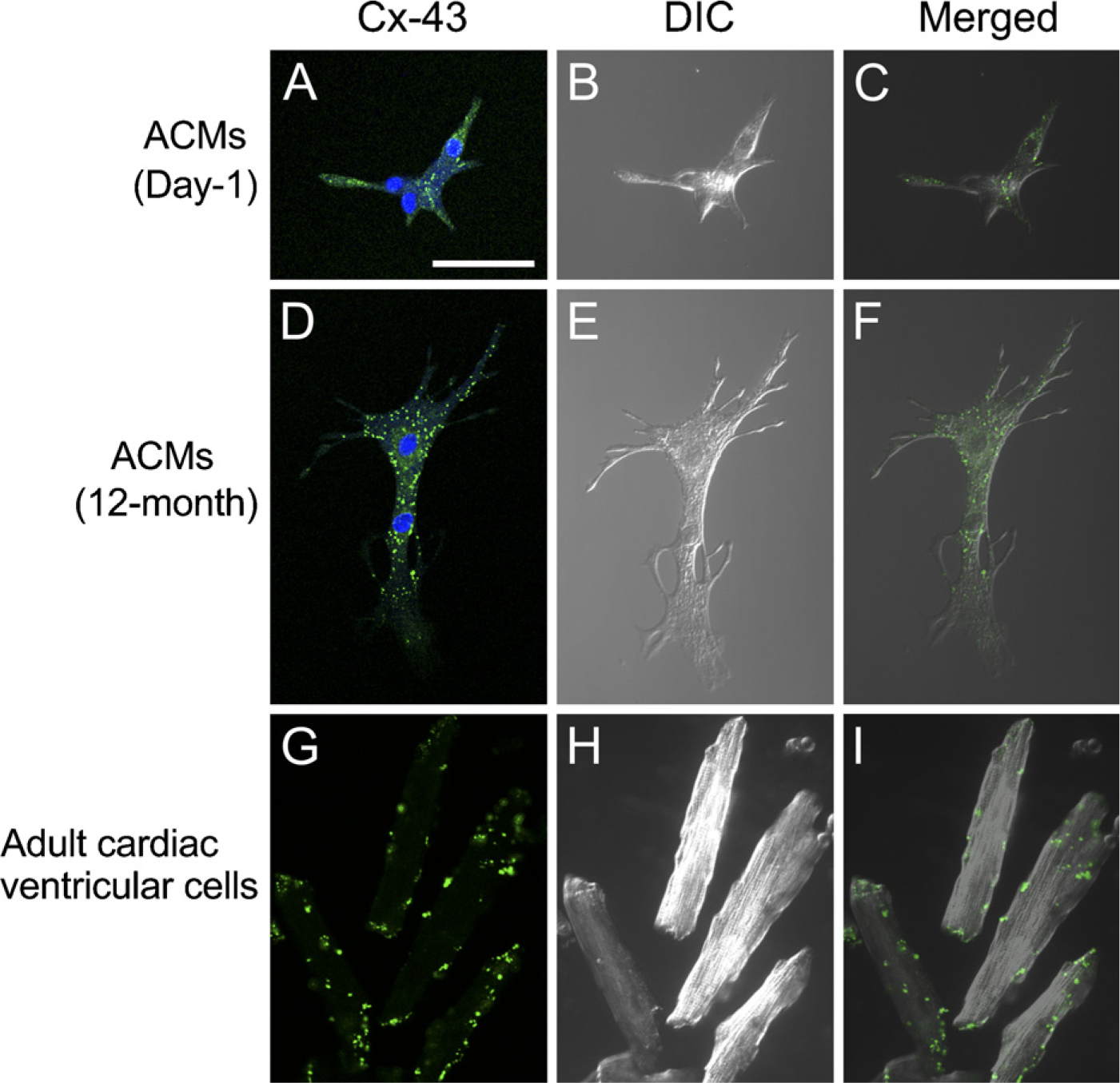
Immunostaining of connexin-43. Confocal laser scanning microscopy of immunostaining of connexin-43 (Cx-43, green), 4'-6-diamino-2-phenylindole (DAPI) staining (blue), and differential interference contrast (DIC) images. ACMs from day-1 (
The present study characterizes the developmental changes in ACMs in the postnatal period. Although ACMs from the adult heart do not proliferate appreciably (Omatsu-Kanbe and Matsuura 2009), it still remains unknown whether the ACMs from the newborn heart could increase their population. We did not detect any appreciable proliferation in day-1 ACMs in either a semisolid culture medium or in the standard liquid culture medium. It is possible that the cell clusters of morphologically immature types of ACMs might represent the results of cell division (Figures 1J–1L). However, as judged from the observation that these cell clusters often contain an odd number of cells (Figures 1K and 1L, and Movie SM2), it is likely that ACMs fuse and thereby form connected structures rather than undergoing cell division. Prolonged culture of day-1 CMDF cells in a dense cell population often yields a large cell assembly comprising morphologically mature ACMs (Figure 7), and the entire area of these assemblies contracts at a single rate (Movie SM3 for Figure 7B). The fact that those large cell assemblies are rarely observed when CMDF cells are well dispersed and cultured at a low cell density (Figures 1 and 4–6) further supports the view that day-1 ACMs tend to fuse to each other but do not proliferate, or even if they do proliferate, few cells could divide under the present culture conditions.
The cell lengths of ACMs from the adult heart are significantly larger than those from day-1 hearts (Figure 3A), and furthermore, even in the adult stage, ACMs from aged mice (12-month-old) display larger cell size, compared with those from younger adults (8–12-week-old). This age-dependent enlargement of ACMs seems to be one of the cardiomyocyte-like characteristics, although it is likely that ACMs remain quiescent throughout their lifetime under in vivo physiological conditions.
In contrast to the age-dependent growth in size (Figure 3A), the cell population of ACMs markedly decreases within the first few weeks of life and then reaches a plateau in the adult stage (Figure 3B). These observations suggest that ACMs undergo a process of cell death in the early postnatal period, similar to the observations reported in cardiomyocytes (Limana et al. 2002). Although it is presently unclear whether ACMs regenerate in vivo, the rate of cell death is thought to be much higher than that of cell renewal in the postnatal heart.
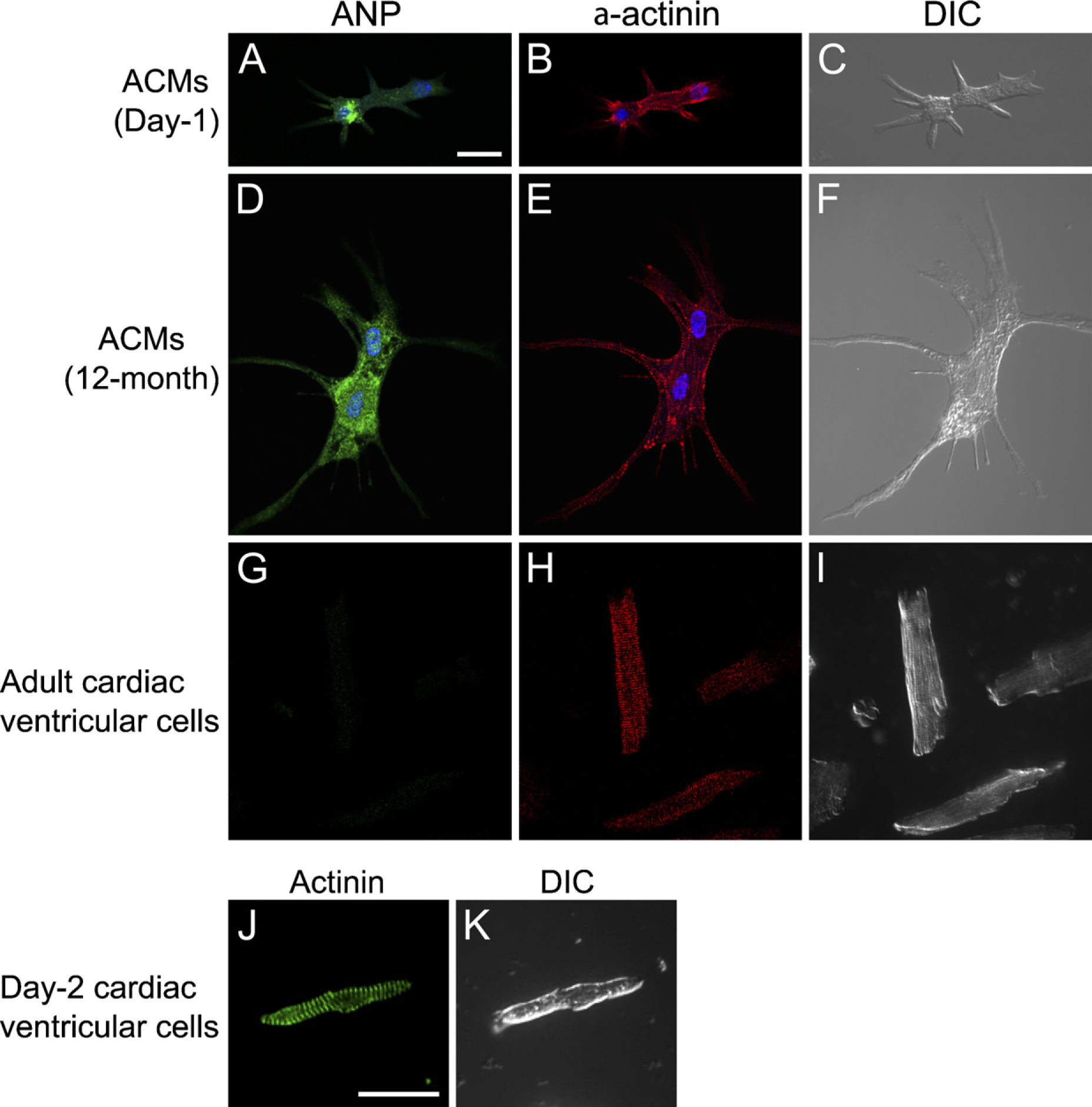
Immunostaining of atrial natriuretic peptide (ANP) and α-actinin. (
The present study detected ×560–820 ACMs in roughly two thirds of the adult mouse cardiac ventricles (Figure 3B), which is smaller in number than that of SA node cells in the mouse (×1700; Mangoni and Nargeot 2001), guinea-pig (<1000; Opthof et al. 1985), and rabbit (×5000; Opthof et al. 1985). Future studies are required to investigate the mechanism of the cell death, as well as the function, of ACMs in the heart.
Newborn ACMs are reasonably expected to express some fetal cardiac gene products, such as ANP (Figures 5A–5C) and Cav3.2 (Figures 6A–6C). It is notable that the ACMs obtained from an aged heart express both ANP (Figures 5D–5F) and Cav3.2 (Figures 6D–6F), which contrasts with the absence of these proteins in adult ventricular myocytes (Figures 5G–5I and 6G–6I). ACMs are thus found to preserve some fetal characteristics. Cardiac hypertrophy reactivates the fetal myocardial gene expression pattern, and the ventricular expression of ANP is considered to be one of the best biological markers associated with ventricular overload (Swynghedauw 1999). The physiological and/or pathophysiological roles of ACMs have yet to be elucidated. However, the origin of ACMs is thought to be functionally masked under in vivo physiological conditions, but might become unmasked in an injured myocardium, thus resulting in the development of self-beating ACMs (Omatsu-Kanbe and Matsuura 2009). Further experiments are needed to examine the quantitative changes in the expression of these fetal genes or gene products in ACMs during postnatal development.
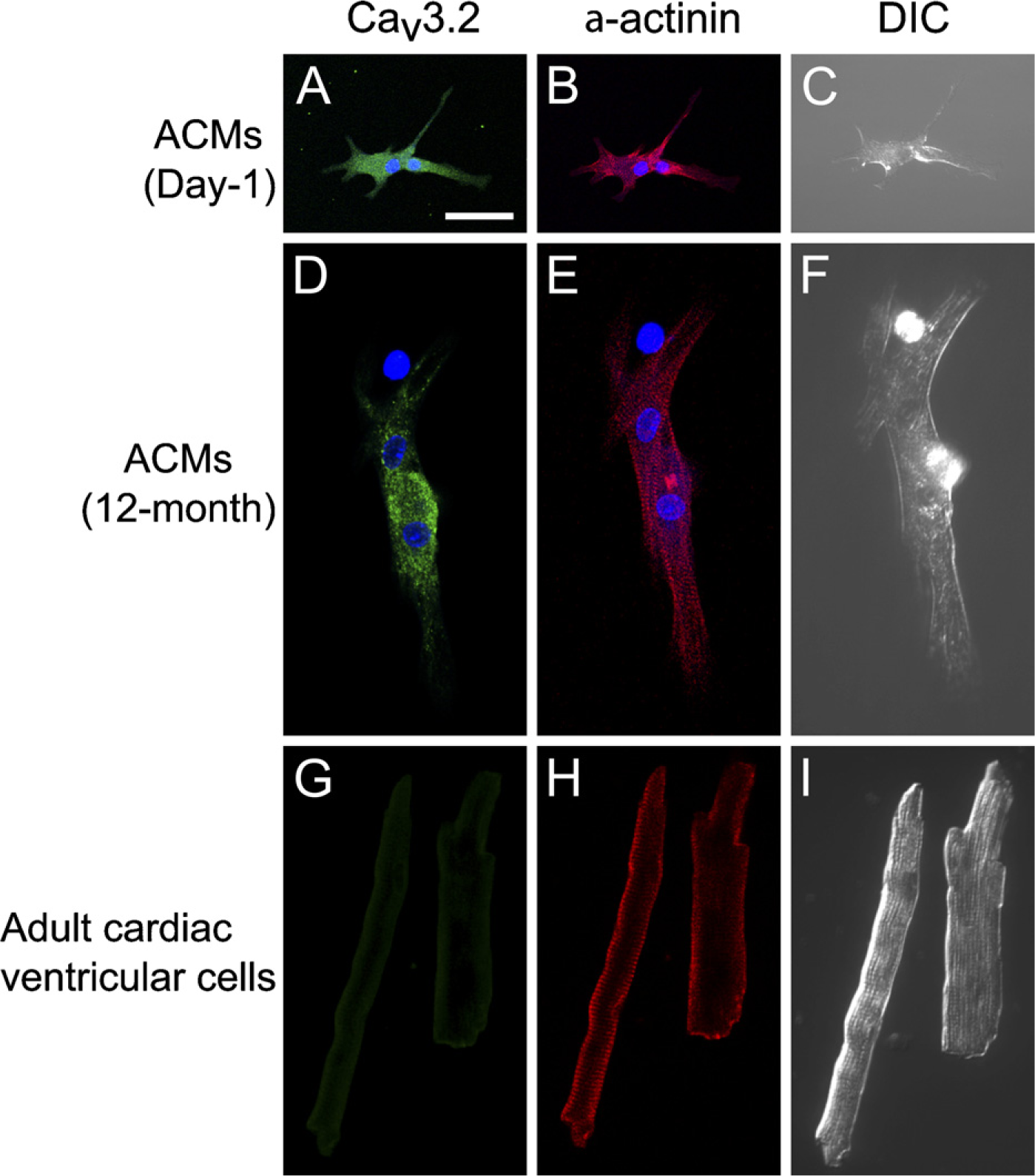
Immunostaining of Cav3.2 and α-actinin. (
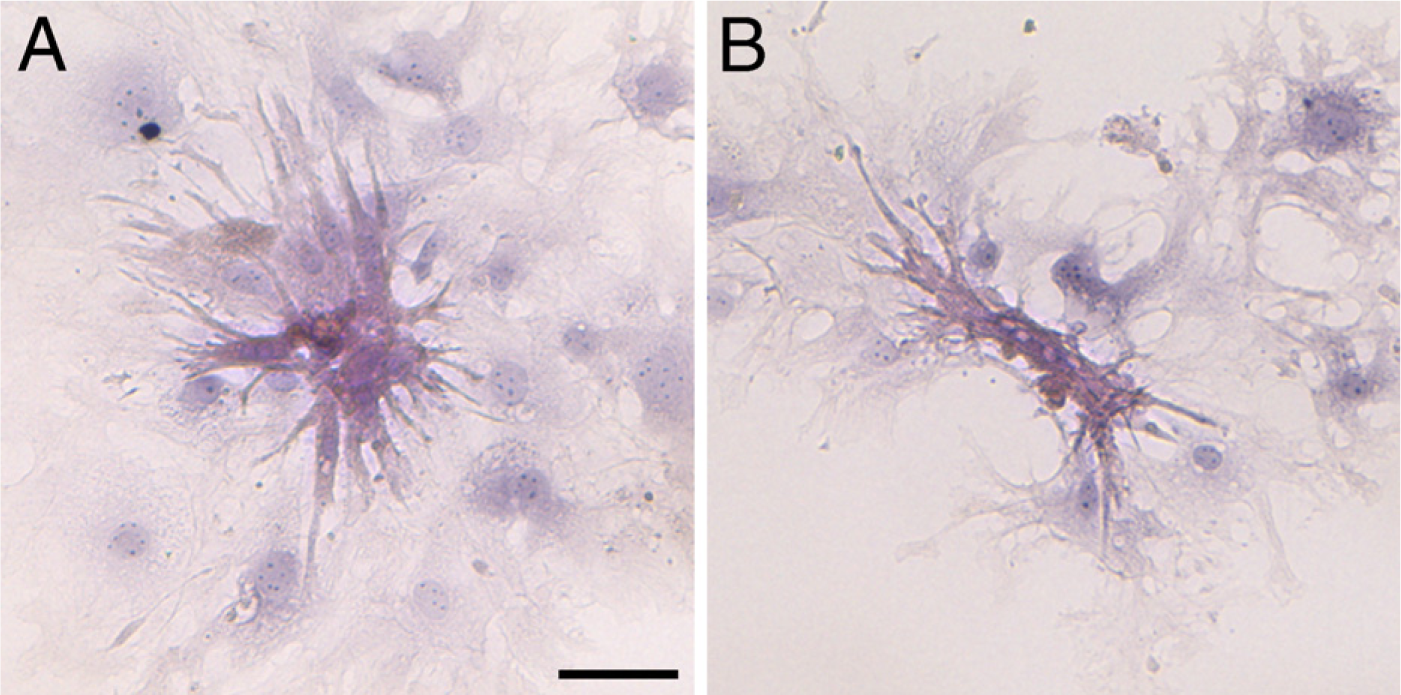
Hematoxylin–eosin (HE) staining in assemblies of day-1 ACMs. ACMs from day-1 mice cultured for 12 days were fixed with paraformaldehyde and stained with HE. Before fixation, the entire area of the ACM assembly in both
In summary, this study confirmed the presence of ACMs in mouse ventricles with the persistent expression of the fetal cardiac genes ANP and Cav3.2 throughout the lifetime of the mouse.
Footnotes
Acknowledgements
This work was supported by a grant-in-aid for scientific research from the Japan Society for the Promotion of Science (no. 19590207 to MO-K).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.