
Abstract
Epoxyeicosatrienoic acids (EETs) are cytochrome P450 metabolites of arachidonic acid, which function in the brain to regulate cerebral blood flow and protect against ischemic brain injury. EETs are converted by soluble epoxide hydrolase (sEH) to the corresponding inactive diol metabolites. Previous animal studies have indicated that sEH gene deletion or treatment with sEH inhibitors results in increased levels of EETs and protection against stroke-induced brain damage. To begin elucidating the underlying mechanism for these effects, we sought to determine the distribution, expression, and activity of sEH in human brain samples obtained from patients with no neurological changes/pathologies. Immunohistochemical analyses showed the distribution of sEH mainly in the neuronal cell bodies, oligodendrocytes, and scattered astrocytes. Surprisingly, in the choroid plexus, sEH was found to be highly expressed in ependymal cells. Vascular localization of sEH was evident in several regions, where it was highly expressed in the smooth muscles of the arterioles. Western blot analysis and enzyme assays confirmed the presence of sEH in the normal brain. Our results indicate differential localization of sEH in the human brain, thus suggestive of an essential role for this enzyme in the central nervous system. This manuscript contains online supplemental material at http://www.jhc.org. Please visit this article online to view these materials.
Keywords
S
In addition to its C-terminal EH domain, sEH also contains an N-terminal phosphatase domain (Cronin et al. 2003; Newman et al. 2003). These two domains have functionally distinct catalytic sites (Newman et al. 2003). Likely, endogenous substrates of the C-terminus include arachidonic acid epoxides [epoxyeisocsatrienoic acids (EETs)] and linoleic acid epoxides, which have been shown to regulate cardiovascular, renal, and inflammatory functions (Node et al. 1999; Capdevila and Falck 2001; Roman 2002; Sun et al. 2002). Endogenous substrates for the N-terminal phosphatase are thought to be phosphorylated lipid metabolites, some of which act as precursors in the cholesterol biosynthesis pathway and in isoprenylation (Newman et al. 2003; Tran et al. 2005; EnayetAllah and Grant 2006).
EETs are potent vasodilators in the brain (Ellis et al. 1990) and are released from astrocytes after glutamate receptor activation (Alkayed et al. 1996, 1997). Inhibitors of EET synthesis block functional hyperemia in brain (Alkayed et al. 1997; Peng et al. 2002, 2004), suggesting that EETs may play a role in neurovascular coupling (Harder et al. 1998). Besides their role as vasodilators, EETs are considered neuroprotective agents because of their anti-inflammatory, antipyretic, antithrombotic, and proangiogenic effects (Larsen et al. 2006). EETs are formed from arachidonic acid through the enzymatic action of CYP-450 epoxygenases (Zeldin 2001), and all four regioisomers of EETs are found in the rodent brain: 5, 6-EET, 8, 9-EET, 11, 12-EET, and 14, 15-EET (Amruthesh et al. 1992; Roman 2002). Studies using CYP-450 epoxygenase inhibitors in the brain showed a 30% reduction in blood flow (Alkayed et al. 1997), thus suggesting a vital role of EETs for regulating cerebral blood flow. Studies in mice showed that sEH inhibition was protective against ischemic brain damage (Zhang et al. 2007). DHETs, the metabolites formed by the hydrolysis of EETs, are generally less potent in their activity compared with EETs; thus, sEH is thought to attenuate EET action (Spector and Norris 2007). The CYP epoxygenase isoforms in the rat brain include CYP 2C11, 2C12, 2D8, 2D9, 2C29, 2C38, and 2C39 (Roman 2002). Immunocytochemical studies suggest that epoxygenase expression in rat brain is localized mainly in astrocytes (Alkayed et al. 1996, 1997).
Several polymorphisms of human sEH affecting the C-terminus have been previously reported (Sandberg et al. 2000; Przybyla-Zawislak et al. 2003), and recently, sEH polymorphisms have been linked to neuronal survival after ischemic injury (Koerner et al. 2007). Sato et al. (2004) showed an association between the Arg287Gln variant and high plasma cholesterol levels in familial hypercholesterolemia patients, thus suggestive of a role for sEH in the isoprenoid/cholesterol biosynthesis pathway (Sato et al. 2004).
In human brain, the mean concentration of unesterified cholesterol is higher than in any other body tissue (23 mg/g), and the majority of this pool is in the myelin sheath of axons (Dietschy and Turley 2004). Cholesterol is produced mainly by oligodendrocytes, a type of glia in the central nervous system (CNS). Currently there is no evidence of net transfer of sterols from blood into the brain or spinal cord, implying that the brain relies solely on de novo synthesis to meet its cholesterol requirements (Bjorkhem and Meaney 2004). Thus, sEH may play a role in regulation of intermediates in the cholesterol/isoprenoid pathway in the CNS.
Relatively little is known about cytochrome P-450 catalyzed arachidonic acid metabolism and the biological effects of the resulting eicosanoids in humans. sEH expression is evident from activity measurements and localization in human liver, kidney, and placental sub-cellular fractions (Pacifici et al. 1988; EnayetAllah et al. 2004). A previous study in rodents (Zhang et al. 2007) showed that sEH was localized in neuronal cell bodies and processes. Currently, rodent models are extensively used to study the potential role of sEH in hypertension and stroke; however, there is considerable variation between the activity and expression of sEH between rodentsandhumans (Oesch et al. 1986; Jung et al. 2005). This study focuses on the distribution, expression, and activity of sEH in the human CNS as a means for understanding the possible neural function of sEH.
Materials and Methods
Tissues and Antibodies
Postmortem normal brain tissue samples (frozen and formalin-fixed paraffin-embedded) and CNS tissue microarrays were obtained from the Cooperative Human Tissue Network–Mid-Atlantic Division (Charlottesville, VA). In most cases, samples were provided with background information (e.g., age, sex), which are listed in Table 1. Frozen tissues were kept at −80C until used.
For IHC procedures, 4-μm sections of paraffin-embedded tissues were used. Multiple sections were histologically evaluated with no pathological findings. Sections of the brain showing signs of freeze-thaw alterations were not used.
Liver sections were used as a positive control for the anti-human sEH (polyclonal rabbit anti-sEH, a gift from Dr. B. Hammock, University of California, Davis) antibody, and cerebellum was used as a positive control for the mouse anti-human glial fibrillar acidic protein (GFAP) antibody as recommended by the manufacturer (Sigma-Aldrich; St. Louis, MO).
The specificity of the anti-human sEH (hsEH) antibody was confirmed by preabsorption and immunoblot analysis. For preabsorption, we incubated the anti-hsEH antibody with excess purified hsEH protein and found no observable staining in brain tissues. Similarly, immunoblot analysis of brain tissues showed a single band at ∼62 kDa, corresponding to hsEH. These findings are consistent with previous studies in our laboratory using immunoblot analysis and preabsorption in a wide array of human tissues (EnayetAllah et al. 2004). The specificity of the GFAP antibody has been confirmed by the manufacturer (Sigma-Aldrich).
Single IHC
Sections on slides were deparaffinized and hydrated by passage through a series of xylene, ethanol, and distilled water washes. For staining with anti-sEH antibody, sections were heated to 95C in Retrieve-All (Signet; Dedham, MA) to unmask antigenic sites as recommended. The sections were washed in Tris buffer (0.096 M Tris-HCl, 0.029 M Tris base, 0.347 M NaCl with 0.025% Triton X-100, pH 7.6) for 10 min. Sections were treated with 3% hydrogen peroxide to quench endogenous peroxidase activity. Nonspecific binding was blocked with 10% normal goat serum and incubated with primary antibody against sEH (polyclonal rabbit anti-sEH, 1:400) for 1 hr. The sections were incubated with goat anti-rabbit horseradish peroxidase (HRP)-conjugated secondary antibody (1:350; Zymed Laboratories, South San Francisco, CA) for 30 min and visualized using the DAB kit (Zymed Laboratories). The DAB kit contains H2O2 solution, DAB solution, and substrate buffer and was used as per the manufacturer's recommendations. Tris buffer was used for washing. The sections were counterstained with hematoxylin, washed in distilled water, passed through a series of rapid dips in ethanol and xylene, and coverslipped. Negative control studies were done using preimmune rabbit serum or IgG for the sEH antibody at equivalent concentrations and incubation periods. All experiments were run in replicate (n=2−5 replicates per experiment), and experiments were repeated on different occasions to confirm constancy of staining in each sample. The expression level of enzyme in different tissues was evaluated blindly by three independent observers (PS, RS, and AE). Distinction between cell types, neuron, and glia was made based on cell size and morphology from both hematoxylin and eosin–stained slides and IHC slides.
Background information of donors (age and sex)
Double IHC
Double immunostaining for sEH and GFAP was performed using HRP and alkaline phosphatase (AP) labeling systems, respectively. sEH antibody was polyclonal (rabbit), and the GFAP antibody was monoclonal (mouse). The sections were deparaffinized and pre-treated in the same way as described above. They were incubated first with the primary antibody against sEH (1:400) for 1 hr, followed by the secondary antibody, goat anti-rabbit HRP (Zymed Laboratories) at room temperature for 30 min. After color development with DAB, the sections were washed with Tris buffer, blocked for 30 min, and incubated with mouse anti-human GFAP (1:600) primary antibody for 1 hr. The Vectastain Elite ABC avidin–biotin–alkaline phosphatase kit (Vector Laboratories; Burlingame, CA) was used for the second label with GFAP. The sections were incubated for 30 min with biotinylated goat anti-mouse secondary antibody followed by AP reagent for 30 min. To detect bound antibodies, 5-bromo-4-chloro-3-in-dolyl phosphate/nitro blue tetrazolium was used as substrate. No cross-labeling was detected in the negative controls where the first or second primary antibodies were replaced by preimmune rabbit serum or mouse IgG, confirming the efficiency of the blocking step for double IHC.
Sample Preparation
Subcellular fractions of frozen brain tissues (frontal lobe, parietal lobe, occipital lobe, temporal lobe, and thalamus) were prepared by homogenizing in a glass homogenizer. Approximately 500 mg of frozen tissue (stored at −80C) was transferred to 1 ml lysis buffer (10 mM HEPES, 10 mM NaCl, 1 mM KH2PO4, 5 mM NaHCO3, 5 mM EDTA, 1 mM CaCl2, 0.5 mM MgCl2, and protease inhibitor cocktail tablets) (Guillemin et al. 2005). All homogenates were centrifuged at 13,000 × g for 20 min at 4C. The supernatant obtained was centrifuged at 100,000 × g for 60 min at 4C. The resulting pellet yielded microsomes, and the supernatant yielded cytosol. Total protein was quantified on all samples with BCA reagent from Pierce (Rockford, IL) using BSA as the standard, according to the manufacturer's directions. Aliquots of the samples were frozen at −80C and stored for enzyme assays.
Enzyme Activity
Cytosolic fractions of brain tissue were isolated as described above and used to measure sEH activity using trans-[2-3H] 1, 3-diphenylpropene oxide (t-DPPO) as described previously (Borhan et al. 1995). Briefly, 1 μl of a 5 mM solution of [3H] t-DPPO in dimethylformamide was added to 100 μl of enzyme preparation in sodium phosphate buffer (0.1 M; pH 7.4) containing 0.1 mg/ml of BSA. The enzyme was incubated at 30C for 20 min, and the reaction was quenched by the addition of 60 μl of methanol and 200 μl of isooctane. Enzyme activity was determined based on the amount of radioactive diol formed in the aqueous phase using a scintillation counter. Assays were performed in triplicate.
Electrophoresis and Immunoblotting
Cytosolic fractions of frozen brain tissue (cerebral cortex, frontal lobe, occipital lobe, parietal lobe, and temporal lobe, and thalamus) were prepared as previously described. Twenty μl (15 μg total protein) of the supernatant fraction was added per lane for SDS-PAGE and Western blotting. Protein concentrations were measured by BCA reagent from Pierce as recommended. SDS-PAGE was performed using 12% resolving gels. Proteins were transferred to 0.2-μm polyvinylidene difluoride transfer membrane (Millipore; Bedford, MA) overnight at 30 V in Tris-glycine buffer with 0.037% SDS and 20% methanol. The membrane was blocked for 1 hr in 5% (w/v) non-fat dry milk plus 0.25% Tween-20 in PBS (pH 7.4) and incubated with polyclonal rabbit anti-human sEH (1:3000) for 2 hr. The secondary antibody, goat anti-rabbit–IgG–peroxidase conjugate (1:5000; Sigma) was incubated for 2 hr. Bound secondary antibodies were detected with super-signal chemiluminescent substrate as recommended (Pierce). Bands were visualized with a Kodak image station 440CF, with Kodak 1D software (Carestream Molecular Imaging; New Haven, CT). Human liver and His-tagged purified sEH were used as positive controls.
Semiquantitative assessment of sEH expression in the brain regions evaluated by three independent observers

The expression level of sEH in different regions was described as low (+), moderate (++), or high (+++). sEH, soluble epoxide hydrolase; GFAP, glial fibrillar acidic protein.
Statistical Analysis
Differences in enzyme activity were analyzed by one-way ANOVA. The criterion for statistical significance was set at p≤0.05.
Results
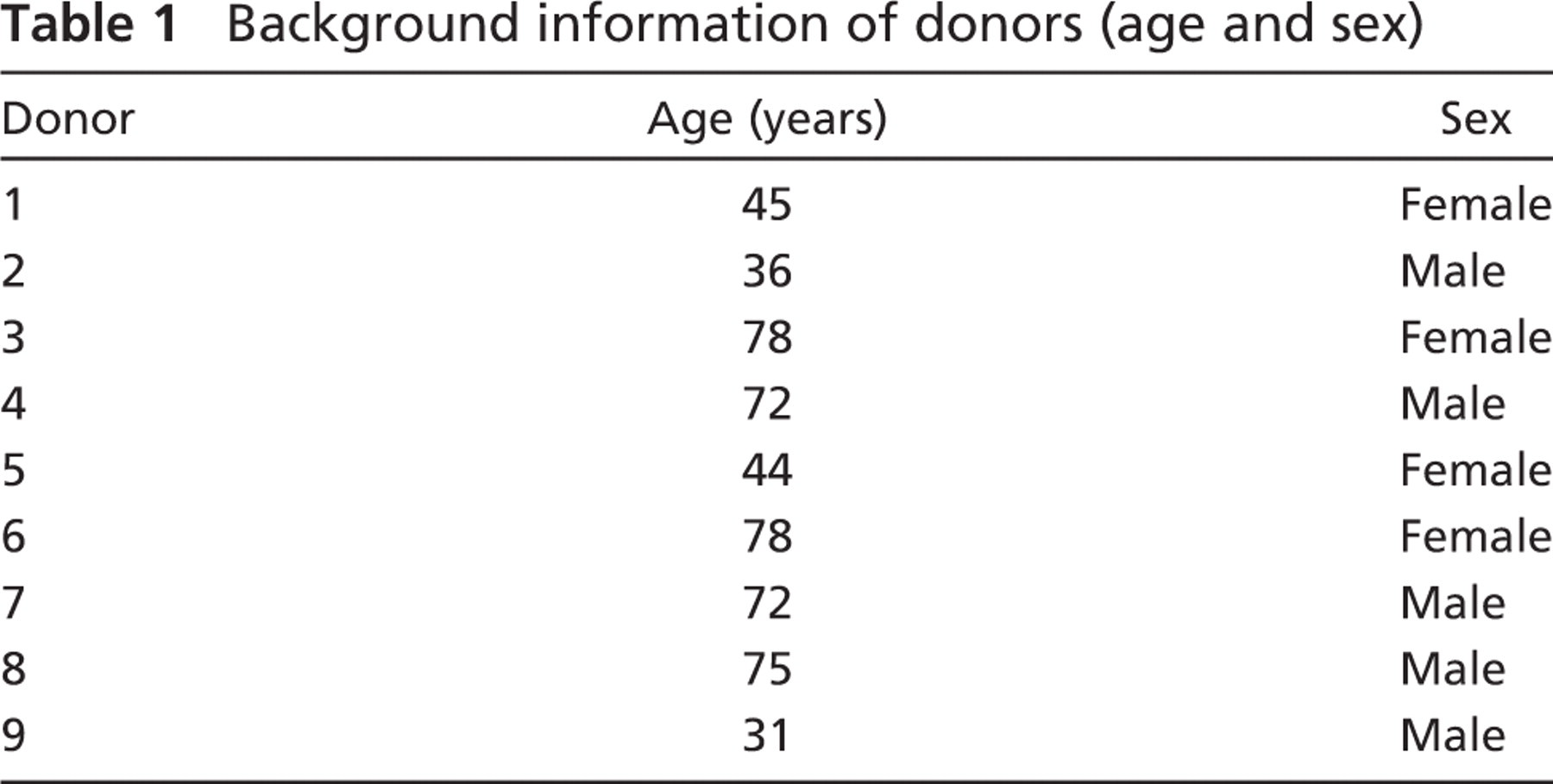
Immunoreactivity for sEH was detected in all brain regions evaluated; however, region- and cell-specific differences were apparent (Table 2). sEH was detected in vascular and non-vascular regions, with predominant expression in the glial subtype (thought to be oligodendrocytes as determined by double labeling), neuronal cell bodies, and neuropil surrounding the neurons. No staining was observed in the negative control slides (Figure 1A). Low-magnification images showing the overall distribution of sEH are provided as a supplementary figure (Supplementary Figure 1). In gray matter (cerebral cortex), sEH immunoreactivity was predominantly in the neurons, oligodendrocytes, and a few scattered astrocytes (Figures 1B–1D). On the other hand, in white matter (Figure 1E), there appeared to be marked immunoreactivity within and surrounding the oligodendrocytes. In regions such as the putamen and globus pallidus, there was punctate staining with distribution predominantly around the oligodendrocytes and astrocytes and occasional cytoplasmic staining of neurons in the latter region. In the substantia nigra (Figure 1F), thalamus, and hippocampus, the distribution was similar to putamen, with the exception of marked immunoreactivity surrounding and within the substantia nigral neurons, thalamic neurons, and hippocampal neurons. Marked immunoreactivity for sEH was detected in the pituitary (Figure 1G). There was diffuse, marked cytoplasmic staining in acidophils and chromophobes; however, no staining was observed in basophils. In the cerebellum, there was scattered staining of the granular cell layer and glial cells of the molecular layer (Figure 1H). In the pons and medulla oblongata (Figure 2A), staining was mainly observed in the neuronal cell bodies.
Soluble epoxide hydrolase (sEH) immunoreactivity (brown) in sections of human brain. (
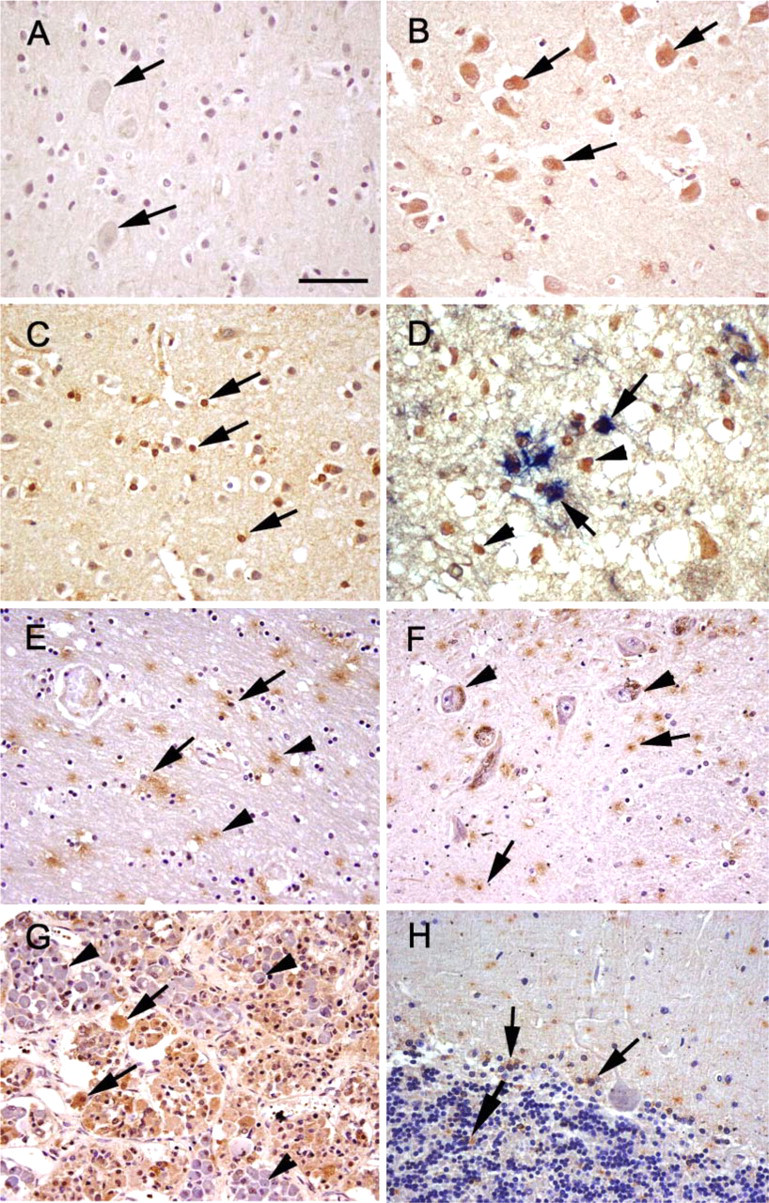
There was robust staining of the neurons of the spinal cord and meningeal blood vessels (Figures 2B and 2C). Marked sEH reactivity was observed in the modified ependymal cells lining the choroid plexus (Figure 2D).
Double labeling of sEH with GFAP was used to differentiate between the two major glial cell populations: astrocytes and oligodendrocytes. sEH distribution was observed mainly in GFAP-negative cells, assumed to be oligodendrocytes based on cell size and morphology. However, occasional colocalization of sEH with GFAP was also observed, especially in regions of the cerebral cortex (Figure 1D).
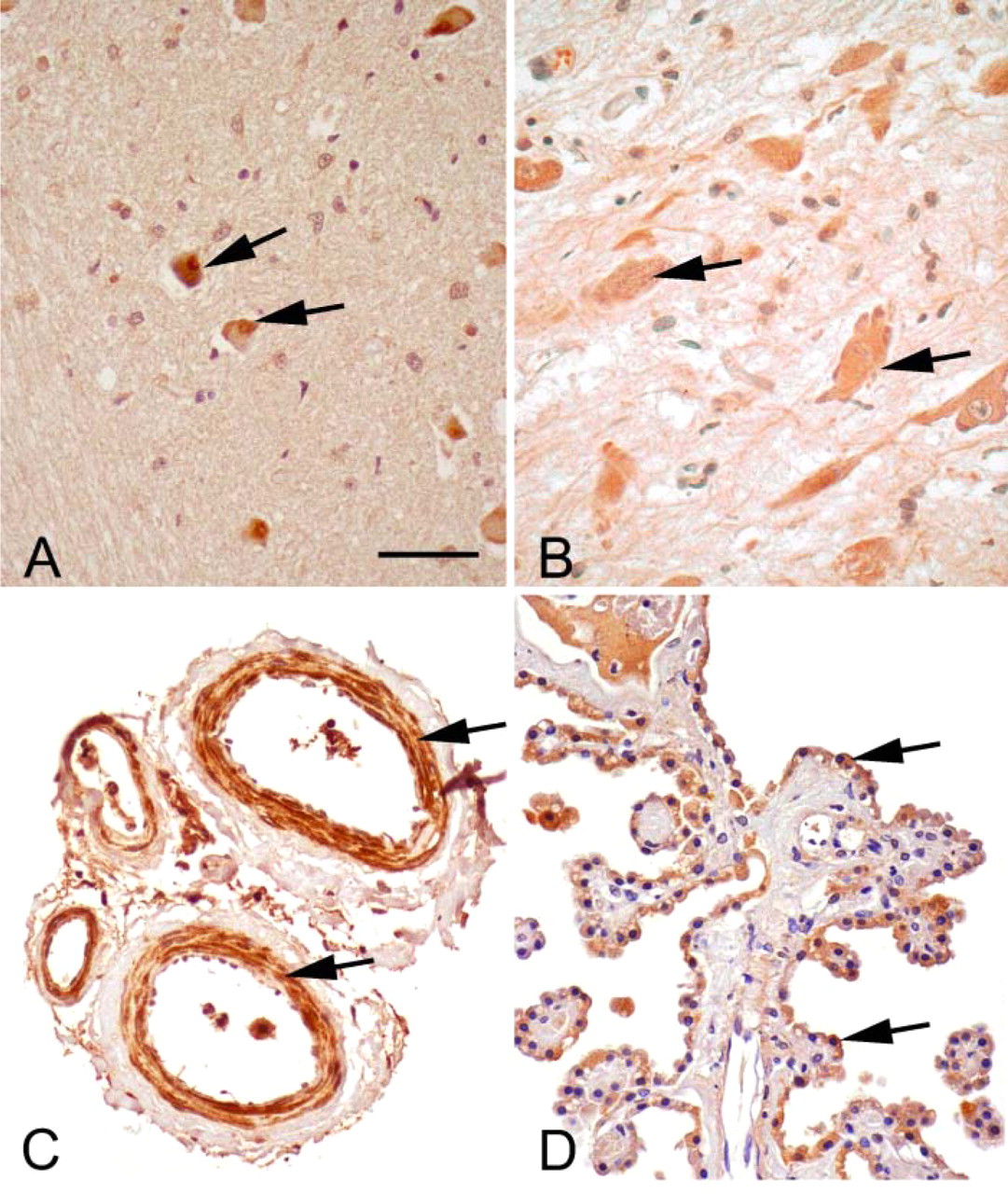
Expression of sEH in the human brain was confirmed by Western blot analysis. sEH immunoreactive protein was detected at the predicted molecular mass of ∼62 kDa (Figure 3). Purified His-tagged sEH was detected at a slightly higher molecular mass than 62 kDa as expected because of the presence of the His-tag. The level of sEH expression in the two brain regions (cerebral cortex and thalamus) was comparable, with little to negligible difference between the different lobes of the cerebral cortex.
sEH immunoreactivity (brown) in medulla, spinal cord, meningeal blood vessels, and choroid plexus. (
Western blot analysis of sEH expression in the human brain. There is sEH expression in the human frontal lobe, occipital lobe, parietal lobe, temporal lobe, and thalamus. His-tagged purified human sEH (hsEH) and human liver are used as positive controls. β-actin was used as a loading control for brain tissue samples.
sEH enzyme activity was measured in all brain tissues using the substrate t-DPPO. Hydration of t-DPPO by the cytosolic fraction of brain homogenates ranged from 1.7 to 2.1 nmol.min-1.mg protein-1 (Table 3). There was no significant difference in enzyme activity between the different brain regions evaluated (ANOVA, p=0.8491). However, the enzyme activity of the liver homogenate obtained from an age-matched individual showed that the liver (16.2 nmol.min-1.mg protein-1) had significantly higher sEH activity (ANOVA, p= 0.002; Table 3).
Discussion
This study provides the first description of the localization, distribution, and activity of sEH in the human brain. Previous reports have documented the presence of sEH in the mouse brain (Shin et al. 2005; Zhang et al. 2007). Since its documentation as a separate enzyme from the microsomal epoxide hydrolase (Ota and Hammock 1980), sEH has been widely studied to understand its expression profile, activity, and potential function. Recently, this enzyme has been implicated in brain pathology, especially in stroke (Zhang et al. 2007). In addition, by regulating cerebral blood flow, substrates of this enzyme (EETs) are thought to be neuro-protective. sEH is now known to contain a phosphatase domain suggested to play a role in sterol biosynthesis and isoprenylation (Tran et al. 2005; EnayetAllah and Grant 2006).
Specific activity of sEH in different brain regions compared with the liver
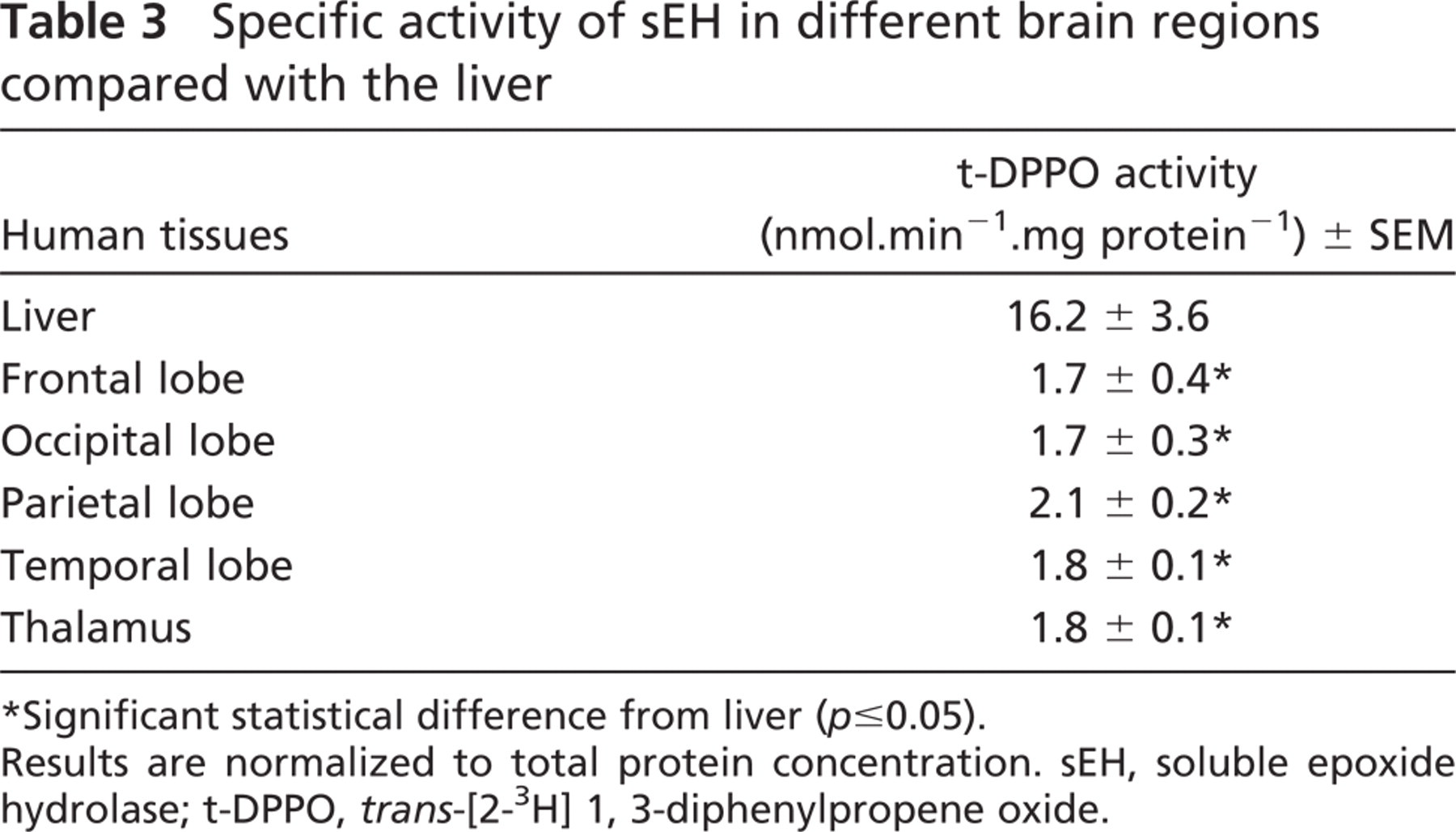
∗ Significant statistical difference from liver (p≤0.05).
Results are normalized to total protein concentration. sEH, soluble epoxide hydrolase; t-DPPO, trans-[2-3H] 1, 3-diphenylpropene oxide.
Although there is evidence indicating that sEH is present in the normal rodent brain (Shin et al. 2005; Zhang et al. 2007), its function in the nervous system is mainly attributed to levels of EETs and its implication to regulation of cerebral blood flow. This study documented the localization of this enzyme in the human nervous system, predominantly in the cytoplasm of neurons, oligodendrocytes, astrocytes, modified ependymal cells, and the smooth muscles of medium-sized arterioles. These results extend and confirm a previous report of localization of sEH in the mouse brain (Zhang et al. 2007) and suggest an important role for sEH in the human brain. The localization of this enzyme in vascular and non-vascular regions implies a potential for different functional roles (e.g., regulation of cerebral blood flow in blood vessels or localized biotransformation reactions in regions such as the choroid plexus).
Oligodendrocytes are responsible for myelin synthesis and maintenance of the integrity of myelinated axons. The localization of sEH in this cell type is suggestive of a role in myelin synthesis, possibly by regulating sterol synthesis. In addition, sEH was detected in certain cell types in the pituitary such as acidophils and chromophobes (Figure 2B). There have been previous reports suggesting the influence of EETs on the release of growth hormone (Snyder et al. 1989) and prolactin (Cashman et al. 1987), which are produced by acidophilic cells of the anterior pituitary. Thus, sEH maybe involved in hormone synthesis and regulation. Also, the pituitary is a region devoid of the blood–brain barrier and thus has higher xenobiotic metabolizing enzyme activities than other parts of the brain (Ghersi-Egea et al. 1992).
The presence of sEH in the smooth muscles on certain arterioles, especially of the meninges and spinal cord, reinforces previous findings of regulation of cerebral blood flow by degrading EETs, which are produced in the brain predominantly by astrocytes and endothelial cells (Alkayed et al. 1996, 1997). EETs have been shown to be anti-inflammatory (Node et al. 1999) and thus important in the progression of stroke. Therefore, sEH may serve as a potential therapeutic target in the treatment of stroke and other neurodegenerative diseases that have inflammatory components.
The high level of sEH expression was striking in the choroid plexus. This distribution was similar to what was observed for CYP4×1 epoxygenase in the mouse brain (Al-Anizy et al. 2006). The choroid plexus is actively involved in the production and circulation of cerebrospinal fluid. Thus, sEH could be performing localized biotranformation reactions in these regions.
In summary, we showed that sEH is present in relatively high abundance in the human brain, with preferential expression in neuronal cell bodies, oligodendrocytes, astrocytes, meningeal blood vessels, and the choroid plexus, suggestive of a neuromodulatory role for sEH. Abundant sEH expression in smooth muscle cells of meningeal blood vessels is consistent with a role for sEH in the regulation of biologically active EET levels. The possibility that interindividual variation in sEH expression and function contributes to the underlying pathogenesis of diseases such as stroke requires further study using a larger sample size. These results are consistent with what is observed in rodents. However, in addition to distribution in neuronal cell bodies and processes, in the human brain, sEH was observed in oligodendrocytes, astrocytes, meningeal blood vessels, and the choroid plexus. This suggests that, in the human brain, sEH may be important in processes other than regulation of EET levels and effects on cerebral blood flow.
Footnotes
Acknowledgements
This work was supported by National Institutes of Health Grant ES-011630.
We thank Dr. B. Hammock (University of California, Davis) for providing the polyclonal anti-human soluble epoxide hydrolase rabbit serum. We also thank Denise Woodward, Ione Jackman, Lynn Howlett (Histology Laboratory, University of Connecticut), and Alex Bothell for advice and assistance during the completion of this work.