
Abstract
Macrophages have multiple roles in the heart including immune surveillance and extracellular matrix remodeling. Aging increases both collagen deposition and macrophage number in the heart; however, rodent models used to study cardiac macrophages have age-related comorbidities such as atherosclerosis and hypertension. The Fischer 344 rat does not develop these conditions with aging; therefore, the purpose of this study was to evaluate macrophage number and polarization in the hearts of aged (24-month) and young (6-month) Fischer 344 rats. Paraffin-embedded hearts were assessed for collagen deposition and immunolabeled for CD68, CD163, CD206, and galectin-3. Compared with young rats, significantly greater collagen deposition was observed in the old rats. There were no significant differences in CD68+ or CD163+ cells between age groups, but both CD206+ and galectin-3+ cells were more numerous in the aged animals. Double-immunofluorescence studies demonstrated that galectin-3 colocalized with both CD68 and CD163, suggesting that galectin-3 is found in cardiac macrophages. Further colocalization studies demonstrated similar proportions of CD68+/CD163−, CD68+/CD163+, and CD68−/CD163+ cells between age groups, suggesting that aging does not affect macrophage polarization. As CD206+ and galectin-3+ cells promote fibrosis, these results warrant future studies that delineate the specific roles of these cells in the aged heart:
Keywords
Introduction
Changes in tissue macrophages with aging have become an area of growing interest. Several investigations have demonstrated that macrophages increase in number with age in most organs studied. 1 Macrophages populate the heart where they facilitate monocyte recruitment after ischemia, 2 promote fibrosis after myocardial infarction, 3 and aid in electrical conduction. 4 Macrophages can alter their phenotype (or polarization) between pro-inflammatory (M1) and anti-inflammatory/matrix-building (M2) subtypes. It should be noted that these are the extremes of the spectrum that are typically induced in isolated cells by lipopolysaccharide/interferon-gamma (M1 macrophages) 5 or Il-4/Il-10 (M2 macrophages). 3 In vivo, macrophage phenotype is complex; several intermediate phenotypes have been described, 6 and macrophages can express markers of both phenotypes.7,8
Macrophages in the heart become more numerous in several models of aging. For example, 24-month-old male Sprague-Dawley rats have more CD68+ (a pan-macrophage marker) and CD206+ cells (M2 marker) than 3-month-old animals. 9 Also, F4/80+ cells (a mouse pan-macrophage marker) are greater in 20-month-old compared with 3-month-old female C57BL/6 mice. 10 Furthermore, in a combined sample of male and female C57BL/6 mice, Mac-3+ cells (Lamp2/CD107B) were more numerous in senescent (26- to 34-month-old) compared with young (6- to 9-month-old) mice. 11 Using flow cytometry, the same laboratory demonstrated that M1 macrophages (defined by the authors as F4/80+/CD206− cells) increased with aging, while M2 macrophages (F4/80+/CD206+) declined. 11
While these investigations show consistent effects of aging on cardiac macrophages, the rodent models utilized develop cardiovascular comorbidities that could affect the age-related differences observed. For example, Sprague-Dawley rats develop hypertension with aging 12 and hypertension increases macrophage number in the heart.13,14 Also, the C57BL/6 mouse strain is highly susceptible to atherosclerosis, cardiomyopathy, and systemic inflammation with aging,15,16 factors that increase cardiac macrophage number. 17 Thus, to better model healthy aging, there is a need to study heart macrophage abundance and polarization in a rodent model with fewer comorbidities. The Fischer 344 rat represents such a model, because it does not develop age-related atherosclerosis or hypertension.18,19 However, these rats do develop systemic inflammation and myocardial fibrosis with aging;20,21 thus, it is reasonable to surmise that these conditions would affect cardiac macrophages. Therefore, the purpose of this study was to quantify macrophages in the hearts of young (6-month-old) and aged (24-month-old) Fischer 344 rats and to further investigate their polarization.
Materials and Methods
Animals
Heart samples are from animals utilized in a previous study 22 conducted at the University of Iowa. The animal protocol was approved by the University of Iowa Animal Care and Use Committee (#0606117). Young (6-month) and old (24-month) male Fischer 344 rats (n=7 per age group) were housed at the University of Iowa animal care facility, fed ad libitum, and kept in individual cages. They were euthanized with an overdose of pentobarbital sodium. Hearts were removed from the animals and sections of the left ventricles were processed for histology as described. 22
Trichrome Staining
Paraffin sections (5 μm) were prepared and then sent to the histology laboratory of IHC World (Ellicott City, MD) where they were stained using the Masson’s trichrome procedure and sent back to the author. The degree of trichrome staining was then assessed in blind fashion by viewing 10 non-overlapping fields per animal with the 20× objective and giving each field a score based on the amount of blue staining. The scale used was as follows: 0—no blue staining or collagen confined to the stroma of blood vessels; 1—infrequent interstitial collagen, light staining; 2—regular and dense interstitial collagen; 3—widespread interstitial collagen, with dark blue staining.
Immunohistochemistry and Cell Counting
Additional 5-μm slides were prepared for immunohistochemistry for macrophage markers using Vector’s ABC reagents and the brown reaction product of diaminobenzidine (DAB) as described previously. 22 Primary antibody dilutions and sources are given in Table 1. These antibodies have been validated in liver where they stain nonparenchymal cells 22 and match staining for each antigen shown in the Human Protein Atlas. 23 Optimization experiments utilizing different concentrations of primary antibodies were performed before staining of all samples. Negative control slides were carried out using isotype-specific sera and did not display a signal or background staining. Two antigens (HO-1 and iNOS) that readily label macrophages in the liver were undetectable in heart samples (Appendix Fig. A1). The lack of iNOS expression in the heart is consistent with previous studies.24,25 Secondary antibody (biotinylated mouse or rabbit; Vector BA-2001 and BA-1000, respectively) concentrations were 1:200. Sections were lightly counterstained with Gill’s hematoxylin, dehydrated, cleared, and coverslipped.
Antibodies and Concentrations, Degree of Staining in the Heart.
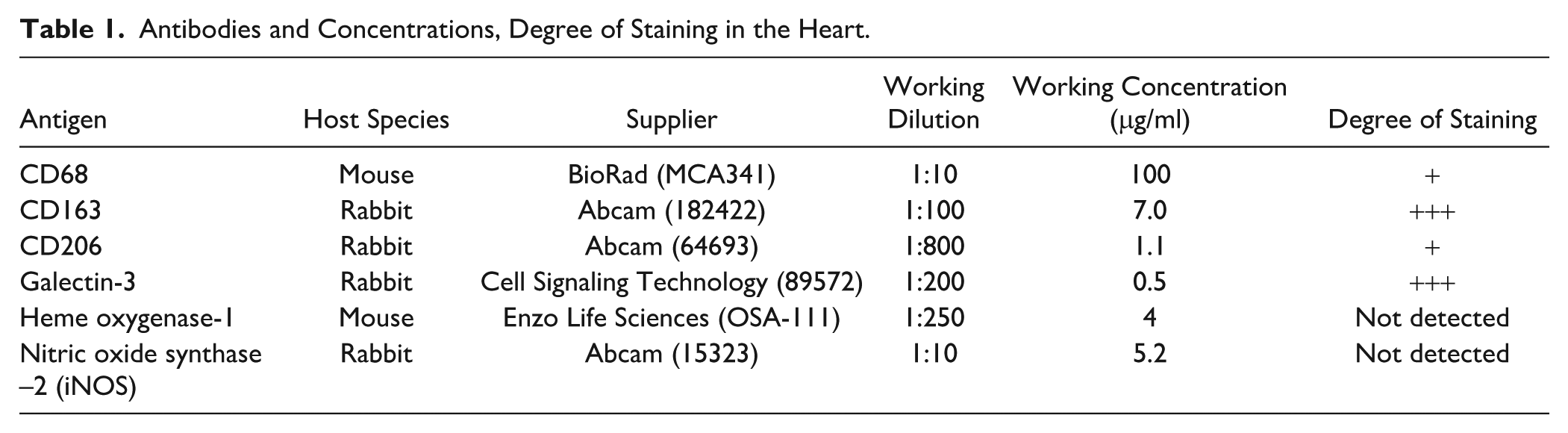
Imaging and micrographs were taken with a Nikon Eclipse E800 microscope and Nikon DXM1200F digital camera. Positive cells (CD163+ and galectin-3+) were counted in 10 non-overlapping images from the 40× objective with the counter blinded to the animal identity. Results are presented as the average number of cells per 40× field. Because CD68 and CD206 cells were less frequent, positive cells were counted in all low-magnification (10× objective), non-overlapping fields on the tissue slice, and then the area in square millimeters of the entire heart slice was determined using Image J. Results are presented as the number of cells per square millimeter of tissue area.
Double immunofluorescence was performed as described 22 with primary antibodies from different hosts. Tissue sections were incubated overnight with the following combinations of primary antibodies: (1) rabbit anti-galectin-3 (1:200) and mouse anti-CD68 (1:10); (2) rabbit anti-galectin-3 (1:200) and mouse anti-CD163 (1:10; ED2 clone, Serotec, MCA342GA); and (3) mouse anti-CD68 (1:10) and rabbit anti-CD163 (1:100, Abcam). Sections were subsequently incubated for 45 min in anti-mouse Alexa 488 (1:100; Vector) and anti-rabbit Cy3 (1:100 Life Technologies, A10520). Attempts to quantify the percentage of galectin 3+ cells that were also CD163+ were unsuccessful due to reductions in positive staining that occurred when using different antibodies. However, using the Serotec CD163 antibody, half of all the animals (4 young, 3 old) demonstrated definitive CD163 staining and colocalization of Gal-3 and CD163 (Fig. 2).
Statistics
Based on standard deviations in cell counts from a previous investigation, 22 power tests indicated that the sample sizes herein were sufficient for a 90% chance of detecting meaningful differences in cell counts at an alpha level of 0.05. All data sets were tested for normality before statistical tests of differences in the means. The CD68 and CD206 cell counts were not normally distributed; therefore, the means of 6- and 24-month-old animals were compared with a Kruskal–Wallis test. The other data sets were normally distributed and were compared via Student’s t-test for independent samples. A p-value of less than 0.05 was considered statistically significant.
Results
Consistent with previous findings on the aged heart,9,21 24-month-old animals displayed significantly greater fibrosis than the 6-month-old animals (p<0.001, Fig. 1).
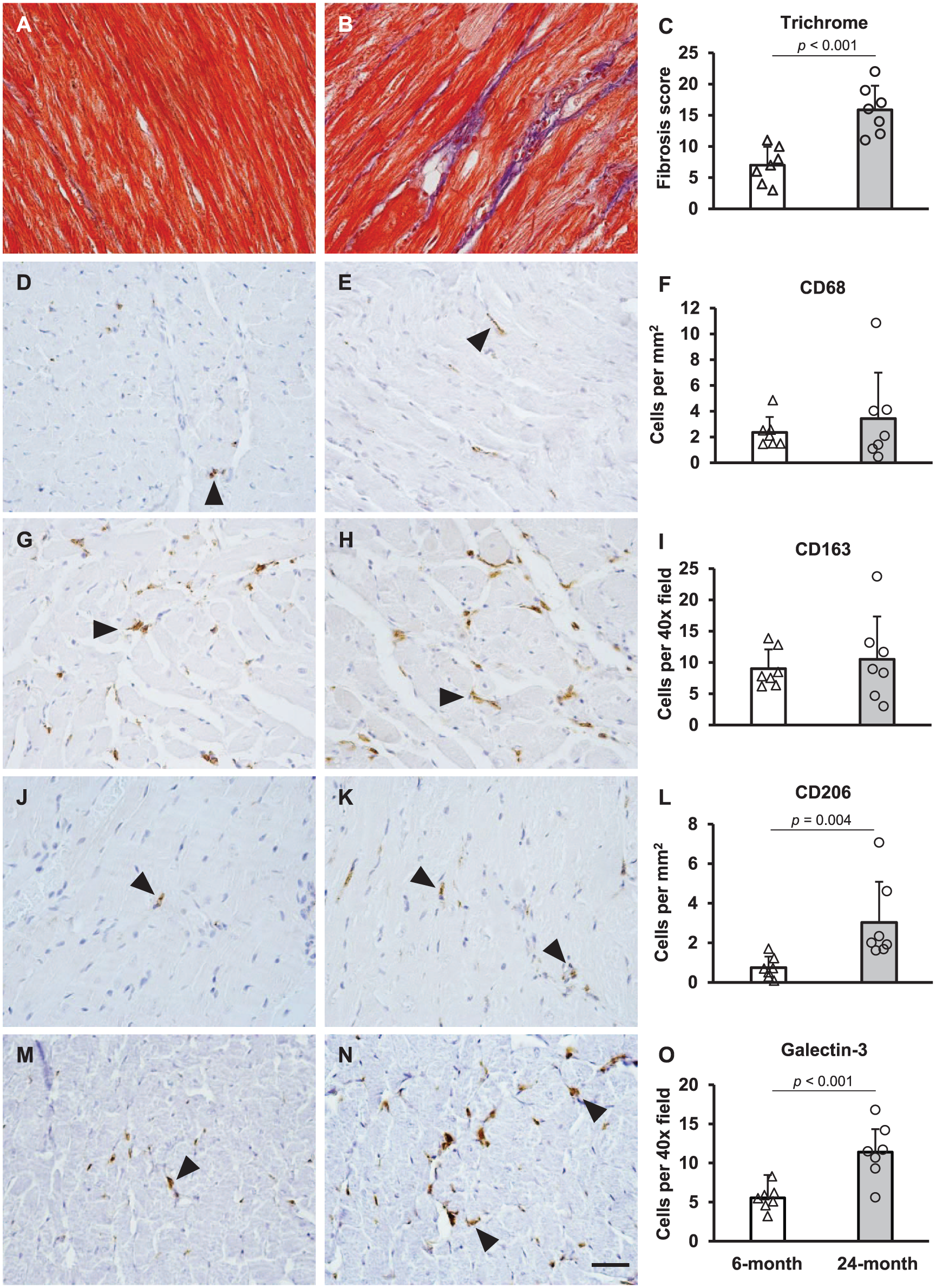
Age-related changes in fibrosis and macrophage markers in Fischer 344 rats. Heart sections from 6-month (A) and 24-month-old rats (B) were stained with trichrome for collagen. Sections were assessed for the degree of fibrosis and scores are quantified in Panel C. The remaining panels show CD68 (D-F), CD163 (G-I), CD206 (J-L), and galectin-3 (M-O) staining in 6-month-old (D, G, J, M) and 24-month-old (E, H, K, N) animals. Cell counts are quantified in panels F, I, L, and O with open bars representing 6-month-old animals and gray bars representing 24-month-old animals. Results are displayed as mean ± SD with individual values shown (n=7 for each age group). p-values are indicated where 6- and 24-month-old animals were significantly different from each other (trichome, CD206, and galectin-3). Positive cells (brown reaction product) are indicated by arrowheads, and the scale bar (Panel N) is 50 μm.
Positive staining for macrophage markers appeared in rounded or spindle-shaped cells interspersed among cardiomyocytes. Differences in the overall abundance of the macrophage markers within cardiac tissue were observed: CD68+ and CD206+ cells were less frequent than CD163+ and galectin-3+ cells (Table 1). The difference in frequency between CD68+ and CD163+ is consistent with a previous investigation. 7 Aging resulted in differential counts of macrophage markers; 24-month-old animals had significantly greater CD206+ (p=0.004) and galectin-3+ cells (p<0.001) than 6-month-old animals (Fig. 1). However, there were no significant differences in the numbers of CD68+ or CD163+ cells between the age groups (Fig. 1).
Macrophages express galectin-3, 26 but this protein is found in other cell types as well. To further characterize the galectin-3+ cells, I performed double-staining immunofluorescence experiments with two macrophage markers: CD68 (a pan-macrophage marker 27 ) and CD163 (an M2 macrophage marker 5 ). The percentage of total galectin-3-positive cells (Gal3+/CD68+ plus Gal3+/CD68−) that were also CD68+ was 31% in young animals and 23% in old animals (not significant; Fig. 2K). Thus, the majority of cells in both young and old animals were positive only for galectin-3 and were likely not macrophages. Half of the animals displayed definitive galectin-3/CD163 staining, demonstrating that galectin-3 also colocalizes with CD163. Overall, these results demonstrate that galectin-3 is found within a fraction of cardiac macrophages.
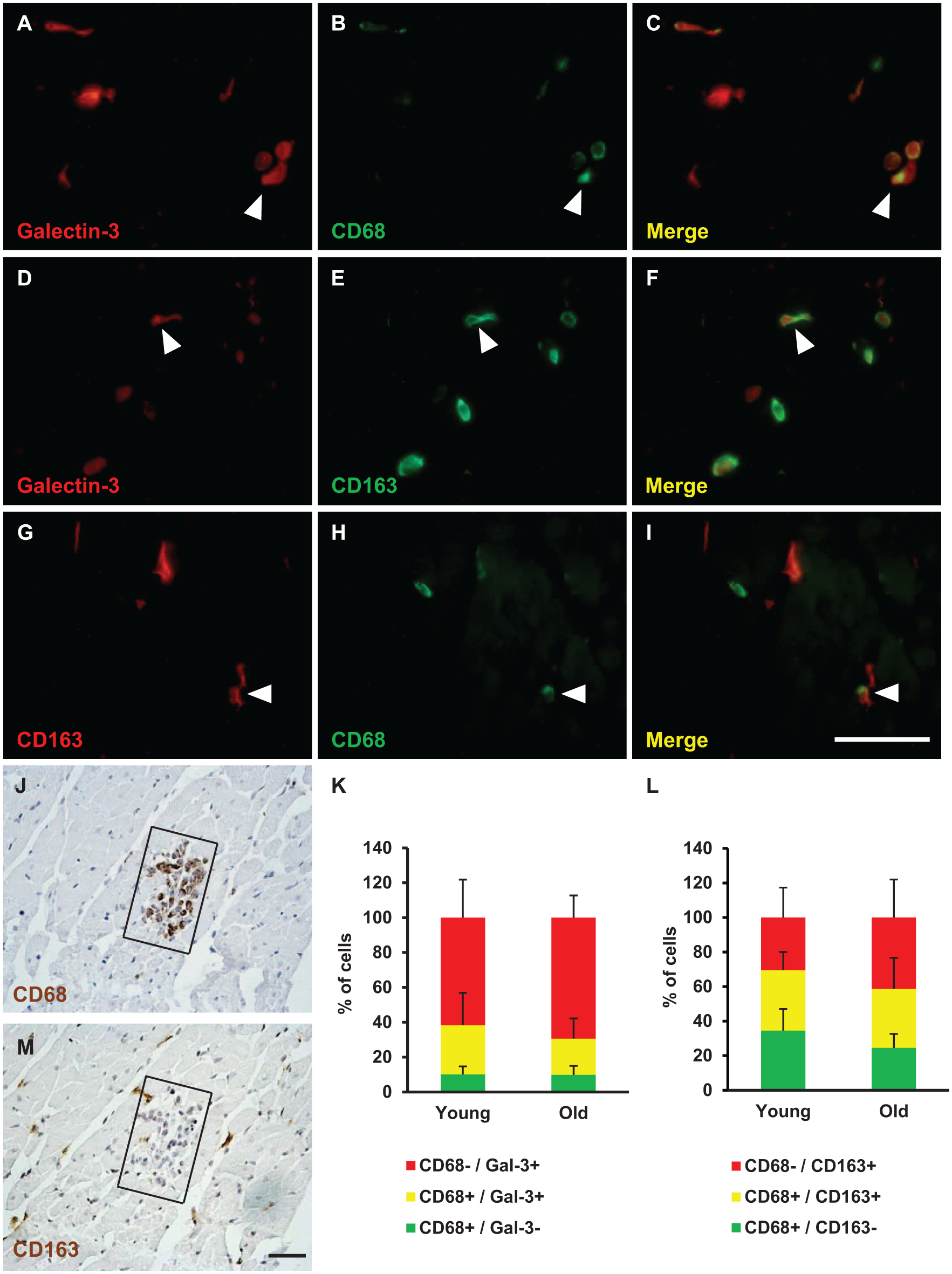
Colocalization of galectin-3 with macrophage markers. Representative images of hearts double-stained with galectin-3 and CD68 (Panels A–C), galectin-3 and CD163 (Panels D–F), and CD163 and CD68 (Panels G-I). The proportions of single and double stained cells are quantified in Panel K (CD68 and galectin-3) and Panel L (CD163 and CD68). Double-stained cells are indicated by arrowheads. Images are representative of young and old animals, but the images here are from young animals. There were no significant differences in the proportions of cells between young and old animals. Results in K and L are expressed as mean percentages of cells plus standard deviation. Panels J and M are serial 5-μm sections from one animal showing an inflammatory infiltrate stained for CD68 (Panel J) and CD163 (Panel M). The scale bars (Panels I and M) are 50 μm.
To distinguish the polarization of cardiac macrophages, I double-stained for CD68 and CD163. 5 Macrophages that are CD68+/CD163− are more responsive to interferon-gamma (IFNγ) than macrophages that are CD68−/CD163+. 28 As IFNγ causes macrophages to adopt the M1 phenotype, CD68+/CD163− cells are likely M1 macrophages. Also, the ratio of CD163 to CD68 macrophages has been used to determine the proportion of M2 macrophages in the heart. 29 Therefore, double-staining for these antigens can assess macrophage polarization. One animal possessing an inflammatory infiltrate provided an opportunity to compare these markers. The infiltrate stained positive for CD68 and negative for CD163 (Fig. 2J and M). This confirms the specificity of antibodies and suggests that CD68+ and CD163+ cells are phenotypically distinct (at least in infiltrates). In young animals, 34.6% of cells were CD68+/CD163− (M1), 34.9% of cells were CD68+/CD163+ (mixed phenotype), and 30.5% were CD68−/CD163+ (M2). In old animals, 24.6% of cells were CD68+/CD163− (M1), 34.1% of cells were CD68+/CD163+ (mixed phenotype), and 41.3% were CD68−/CD163+ (M2; Fig. 2L). There were no significant differences between age groups in these cell populations.
Discussion
In a rodent model of aging in which animals do not develop atherosclerosis or hypertension, this investigation has demonstrated that cardiac CD206+ and galectin-3+ cells are more numerous in aged, compared with young animals. This suggests that aging, in the absence of cardiovascular comorbidities, results in macrophage accrual in the heart. CD206 is a mannose receptor on macrophages, commonly utilized to identify the M2 macrophage phenotype. 5 Galectin-3 is a multifunctional protein that promotes inflammation and fibrosis. It is expressed by both M1 and M2 macrophages in vitro, 26 and the current investigation demonstrates that galectin-3 is expressed by macrophages in cardiac tissue. Two investigations have also shown an age-related increase in M2 macrophages in the heart: CD163+ cells in Rhesus Macaques 30 and CD206+ cells in Sprague-Dawley rats. 9 The greater numbers of galectin-3+ cells in the old rats compare favorably to the work of Kasacka and associates, showing an increase in galectin-3 staining intensity in hearts from 57-year-old men compared with 38-year-old men. 31
While the present investigation has not established causality, more numerous CD206+ and galectin-3+ cells could contribute to the greater degree of collagen deposition in aged hearts as M2 macrophages have a role in extracellular matrix production. After myocardial infarction, CD206+ macrophages accumulate in the infarct region, and their depletion impairs the fibrotic response. 32 Similarly, galectin-3 promotes cardiac fibrosis. Infusion of galectin-3 into rats increases left ventricular collagen content 33 and overexpression of cardiac galectin-3 increases trichrome staining in the heart. 34 Future cell-specific ablation studies in aged organisms are needed to confirm the roles of CD206 and galectin-3 in age-associated fibrosis of the heart.
Further evaluation of macrophage polarization in this investigation has suggested that cardiac macrophages are not strongly polarized, which is consistent with Mylonas and associates. 35 While iNOS, a canonical M1 marker, could not be utilized, studies have shown that the lack of an M2 marker on a cell labeled with a pan-macrophage marker identifies an M1 macrophage.11,28,29 Therefore, the CD68+/CD163− cell population in this study are likely M1 macrophages. There were no significant differences in the percentages of CD68+ and CD163+ cells between young and old animals, suggesting that aging does not result in a shift in macrophage polarization in the heart. This is consistent with results in the livers of Fischer 344 rats showing that while macrophage numbers increased with aging, the proportions of M1 and M2 macrophages were similar. 8 However, this result conflicts with the results of Ma et al, 11 who showed a shift toward M1 cardiac macrophages in aged mice. As described earlier, this discrepancy could be explained by the C57 mouse being more prone to inflammation and atherosclerosis, comorbidities that could influence macrophage polarization.
There are some limitations to this report which are discussed here. The inclusion of a canonical marker for M1 macrophages would provide a more comprehensive assessment of macrophages in the aged heart. Unfortunately, iNOS protein is not detectable in rat myocardium (Appendix Fig. A1).24,25 Other specific M1 markers such as TNFα and IL-1β are cytokines that are either not detectable via immunohistochemistry or do not label cardiac macrophages.23,36,37 One investigation used CD68 as an M1 marker and showed that CD68+ cells increased in the heart between 1 and 7 days after isoproterenol injection. 7 At 3 days, there was very little overlap of CD68+ and CD163+ macrophages, suggesting that they labeled different macrophage phenotypes. 7 However, assessment of CD68+ cell polarization in the quiescent heart was not performed. Therefore, definitive immunohistochemical labeling of M1 macrophages in the rat heart remains a challenge. Methods such as flow cytometry on isolated cardiac macrophages would be helpful on this point, to better quantify cell numbers as well as the levels of expression of macrophage polarization markers in heart tissue. However, as it is not always feasible to isolate fresh hearts from aged animals, the present study is useful because it utilizes a standard technique on preserved samples to quantify cardiac macrophages. Future studies should also include multiple age groups to better assess the trajectory of changes in macrophages during aging. Finally, this study is limited to male rats, and it would be insightful to perform similar studies on female rats.
Overall, these results confirm that cardiac macrophages accrue with aging in the Fischer 344 rat, which develops fewer comorbidities than other rodent models. The results of this investigation warrant future studies on why galectin-3+ and CD206+ cells accumulate with aging and the role of these proteins in age-related cardiac fibrosis.
Footnotes
Appendix
Acknowledgements
I thank Kathleen Bloomer, Megan Sullivan, and Anastasiya Volyanyuk for assistance with histological techniques and cell counting.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
SAB designed the study, performed the immunohistochemistry, counted cells, analyzed the data, and wrote the manuscript.
Data Availability
Data are available from the author upon reasonable request.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The author was funded by a Faculty Development Grant from Penn State Abington.