
Abstract
High levels of expression of galectin-1 and galectin-3, the β-galactoside-binding proteins, have been recently described in malignant thyroid tumors but not in adenomas nor in normal thyroid tissue. However, there are no data about the expression of these galectins during fetal thyroid development. In this study we analyzed immunohistochemically the presence of galectin-1 and galectin-3 in human fetal thyroid glands (16–37 weeks of gestation). Weak to moderate cytoplasmic staining for galectin-1 was observed in follicular cells of all fetal thyroids. Galectin-3 could not be detected in thyroid follicular cells of any fetal thyroid investigated. Both galectins were detected in stromal tissue, but staining for galectin-1 was more intense. The absence of galectin-3 in thyroid cells during fetal development suggests that galectin-3 is expressed de novo during malignant transformation of thyroid epithelium, and that galectin-1 could be considered an oncofetal antigen. The results obtained indicated potential roles for galectin-1 and galectin-3 during the investigated period of human fetal thyroid gland development. Both galectins might participate in developmental processes regarding stromal fetal thyroid tissue organization, whereas galectin-1 might have a function in thyroid epithelium maturation.
G
One of the striking properties of galectins is developmental regulation of their expression, localization, and activity (Hirabayashi and Kasai 1993; Leffler 1997, 2001). For example, differences in expression of galectin-1 and/or galectin-3 were found during human embryogenesis (Van den Brule et al. 1997; Winyard et al. 1997), fetal development of bovine respiratory and digestive tracts (Kaltner et al. 2002), rat lung (Clerch et al. 1987), and during mouse organogenesis (Colnot et al. 1997). Depending on the cell or tissue type, they can be up- or downregulated or expressed transiently, with peaks at critical points of development and morphogenesis. Galectin-1 and galectin-3 can be detected in the cytoplasm, nucleus, at the cell surface, or in the extracellular matrix, and their localization is also characteristically changed in different developmental stages and physiological conditions. The dynamic pattern of expression therefore suggests a role in the modulation of cell adhesion and signaling during growth and differentiation (Zalik 1991; Zhou and Cummings 1992; Van den Brule et al. 1997; Leffler 2001). With regard to development of the human thyroid, there are no data about galectin expression in this gland during fetal life. Thus far, thyroid galectins have been examined only in adult tissue (Xu et al. 1995; Fernandez et al. 1997; Cvejić et al. 1998,2000; Orlandi et al. 1998; Inohara et al. 1999; Kawachi et al. 2000; Saggiorato et al. 2001; Coli et al. 2002). The results obtained indicated that galectin-1 and galectin-3 are absent from normal tissue and benign tumor tissue. Conversely, thyroid carcinomas express both galectin-1 and galectin-3, the latter being suggested as a reliable presurgical tumor marker (Orlandi et al. 1998; Inohara et al. 1999).
In this study we examined immunohistochemically the expression of galectin-1 and galectin-3 in human fetal thyroid, in an attempt to determine whether or not galectin expression in thyroid carcinoma represents a de novo expression. The results obtained indicated a characteristic pattern of distribution in the cells and extracellular compartment, suggesting involvement of galectins in particular developmental processes in the human thyroid gland.
Materials and Methods
Tissue Samples
A total of 23 thyroid glands were obtained from human fetuses (gestational age range of 16 to 37 weeks) with local ethical committee approval. Fetuses were obtained from spontaneous or therapeutic abortions for social-legal causes and they had no visible developmental abnormalities. The gestational ages of fetuses were estimated from the maternal menstrual history and measurement of fetal size. The thyroid tissue was fixed in Bouin's solution and processed after paraffin embedding.
Immunohistochemistry
The following antibodies were used: (a) a rabbit anti-human placental galectin-1 antiserum and (b) a rat monoclonal antibody (MAb) M3/38 against galectin-3. A polyclonal antibody to galectin-1 was produced and characterized in our laboratories as described earlier (Vićovac et al. 1998). In brief, galectin-1 was isolated from human term placenta by affinity chromatography on an asialofetuin-Sepharose 4B column. The affinity-purified lectin was used for production of antiserum by repeated immunization of rabbits. The antiserum obtained gave one sharp precipitation line in double diffusion against total extract of placental tissue and was checked for specificity in a blotting assay. No crossreactivity with galectin-3 was found.
A rat MAb M3/38 (IgG2a subclass) is a hybridoma supernatant produced by TIB 166 (American Type Culture Collection; Rockville, MD) against murine macrophage cell surface antigen Mac-2 (Ho and Springer 1982), which is identical to L-29 (Huflejt et al. 1993), i.e., galectin-3. This antibody was characterized previously and found to crossreact with human galectin-3 (Cherayil et al. 1990). Antibody was kindly supplied by Dr M. E. Huflejt (La Jolla Institute for Allergy and Immunology; San Diego, CA).
Immunostaining was performed on 4–6-μm-thick sections using the avidin-biotin–peroxidase complex (ABC) technique (Hsu et al. 1981) with Vectastain Elite ABC Kit (Vector Laboratories; Burlingame, CA).
After deparaffination and rehydration, endogenous peroxidase activity was blocked with 0.3% H2O2/methanol followed by nonimmune serum for 20 min to block nonspecific binding. The sections were then incubated with primary antibody to galectin-1 or to galectin-3 at 4C overnight at a dilution of 1:600 and 1:200, respectively. This was followed by incubation with biotinylated goat anti-rabbit IgG for galectin-1 detection or biotinylated horse anti-mouse IgG for galectin-3 detection. Both second antibodies were from Vector Laboratories. Next, the sections were incubated with avidin-biotinylated peroxidase complex for 30 min. Between each step, sections were washed three times in PBS. The reaction was visualized using 3,3′-diaminobenzidine tetrahydrochloride (DAB Peroxidase Substrate Kit; Vector Laboratories). After counterstaining with hematoxylin, slides were dehydrated, coverslipped, and examined with a Reichert-Jung microscope supplied with a Photostar automatic camera system (Vienna, Austria). Negative controls included omission of the primary antibody and the use of nonimmune serum, both resulting in complete absence of staining. Staining of thyroid epithelium or stromal compartment, after evaluation by two observers independently was scored as follows: −, no staining; +, weak staining; ++, moderate staining; and +++, strong staining.
Results
The results of the immunohistochemical analysis of galectin-1 and galectin-3 in human fetal thyroid tissue are presented in Table 1. Some representative photomicrographs are shown in Figures 1A-1F.
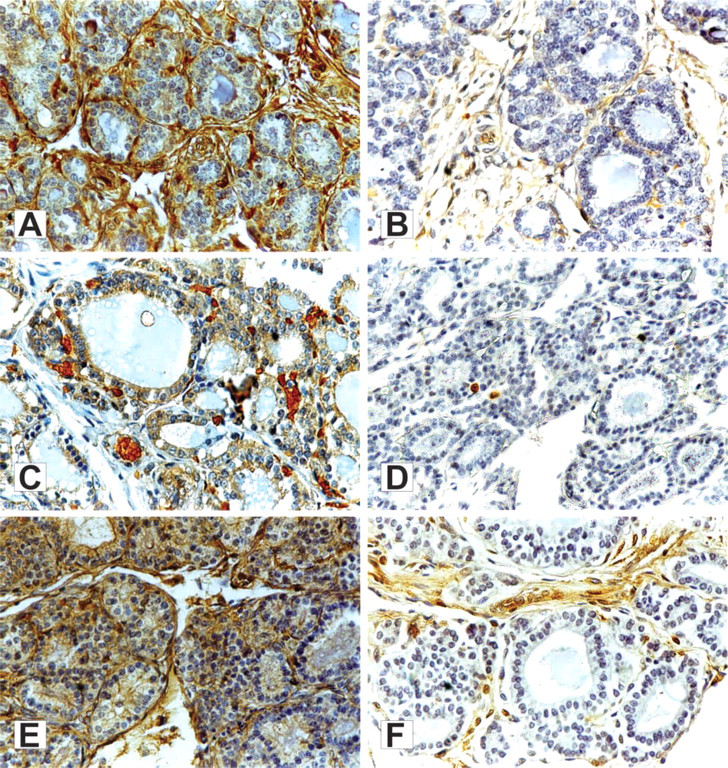
Immunohistochemical analysis of galectin-1 (
Expression of galectin-1 and galectin-3 in the thyroid tissue of human fetuses a
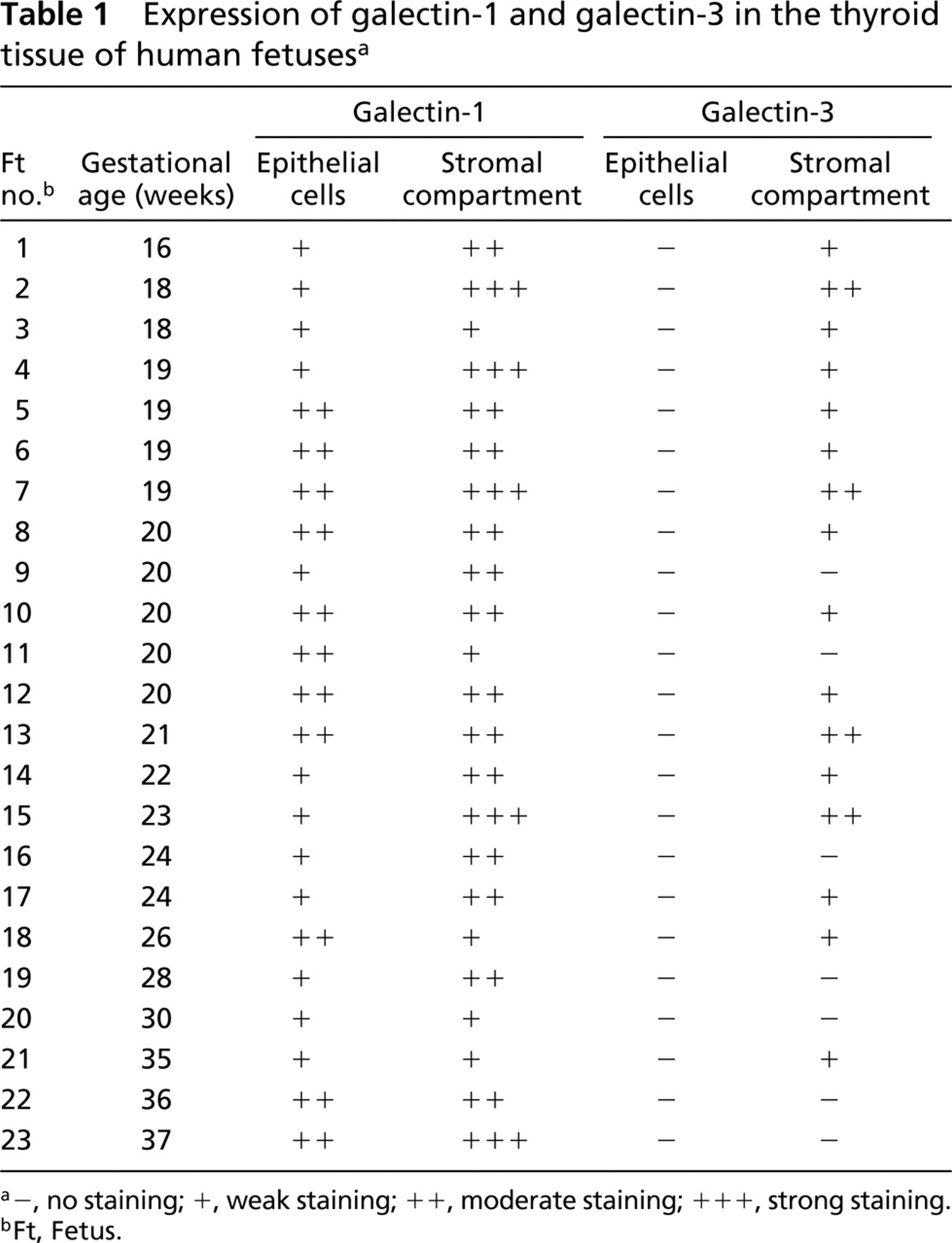
−, no staining; +, weak staining; ++, moderate staining; +++, strong staining.
Ft, Fetus.
Light microscopic examination of fetal thyroid tissue revealed a more or less organized follicular structure, which was generally in accordance with the gestational ages of the fetuses.
Immunohistochemical staining using a polyclonal antibody against galectin-1 was observed in all cases examined (Table 1; Figures 1A, 1C, and 1E). Galectin-1 was localized in the cytoplasm of thyroid epithelial cells. Cytoplasmic staining was diffuse, ranging from weak (12 cases) to moderate (11 cases). Nuclear staining was not found. The apical part of the cell membrane, which faces the follicle lumen, was occasionally stained. Immunoreactivity was also found in stromal cells, extracellular matrix, and host-derived nonthyroid cells. The intensity of staining in the stromal compartment varied from weak to moderate or strong, as shown in Table 1.
Immunostaining using an MAb against galectin-3 showed positivity only in the extracellular compartment (Figures 1B and 1F). Thyroid epithelium was negative in all fetal thyroid tissues examined. Staining for galectin-3 in the stromal compartment varied from weak (12 cases) to moderate (four cases) and in general was less intense than staining for galectin-1. In seven cases no immunoreactivity was found either in follicular cells or in the stromal compartment (Figure 1D).
Discussion
This immunohistochemical study has for the first time analyzed galectin-1 and galectin-3 expression in human fetal thyroid glands. The results obtained demonstrate that they are differentially expressed during the investigated period of fetal life.
During thyroid gland development, the histological differentiation of the follicular epithelium can be considered to pass through three stages (Shepard 1967), depending on the general level of colloid development: the precolloid stage (7–13 weeks), the initial colloid stage (13–14 weeks), and the follicular stage (>14 weeks of gestation). Because new cells are constantly being added, however, all older fetal ages also include follicles at less advanced histogenetic stages (Pinter 1996). We have shown here that galectin-1 is expressed in thyroid cells during the follicular phase of histogenesis of the thyroid follicle. Whereas galectin-1 was consistently found in the cytoplasm of thyroid follicular cells, galectin-3 could not be detected in thyroid cells of any fetal thyroid gland investigated. On the other hand, at the studied developmental stage, both galectin-1 and galectin-3 were detected in stromal cells and extracellular matrix.
The follicular phase is a period during which maturation of the thyroid epithelium progresses in terms of multiplication of follicles and enlargement of the follicular lumen (Sinadinović et al. 1986; Pinter 1996). In addition, the volume of thyroid stroma increases continuously (Bocian-Sobkowska et al. 1992,1997). At the same time the ratio among different histological compartments (epithelium, colloid, and stroma) changes considerably (Bocian-Sobkowska et al. 1992,1997). In general, galectins have a number of properties that contribute to tissue remodeling, including the regulation of cell adhesiveness and motility and promotion of cell growth (Zhou and Cummings 1992; Barondes et al. 1994; Leffler 2001; Hughes 2001). According to the possible role of galectins in cell-cell and cell-matrix interactions, thyroid galectins could be involved in the important morphological and cellular changes that occur at this developmental stage. In this context, it is possible that galectin expression in stromal fibroblasts is related to their proliferative status during extracellular matrix organization in fetal thyroid tissue. On the other hand, cytoplasmic localization of galectin-1 in thyroid follicular cells might be important for intracellular processes.
The results of our work and available data about galectin expression in normal and transformed adult thyroid tissue indicate that galectin-1 could be considered as an oncofetal antigen with regard to thyroid epithelial cells. As for galectin-3, it appears to be neoexpressed during malignant transformation of thyroid epithelium. However, we cannot rule out the possibility of galectin-3 expression in thyroid cells in the early stages of embryonic/fetal life (before 16 weeks).
Although galectin-1 and galectin-3 share structural and biochemical similarities, their different expression patterns and cellular localization suggest different biological roles in the fetal thyroid. Identification of their endogenous ligands at a specific developmental stage or in pathological conditions, and in relation to thyroid cell cytosol and extracellular matrix, will be informative regarding their possible roles in fetal thyroid gland development.
Footnotes
Acknowledgements
Supported by the Ministry of Science, Technologies and Development of the Republic of Serbia, project 1504 “Glycobiological aspects of physiological and pathophysiological processes.”
We are grateful to Dr M. E. Huflejt (La Jolla Institute for Allergy and Immunology; San Diego, CA) for providing a monoclonal antibody against galectin-3. We also thank Dr Miloš Cvetković (Department of Gynecology and Obstetrics) and Dr Radoslav Radosavljević (Department of Histopathology, Clinical Center of Serbia) for their help in obtaining human fetal thyroid glands and providing necessary data about the fetuses.