
Abstract
The six mammalian glycosaminoglycans (GAGs), chondroitin sulfate, dermatan sulfate, heparin, heparan sulfate, hyaluronan, and keratan sulfate, are linear polysaccharides. Except for hyaluronan, they are sulfated to various extent, and covalently attached to proteins to form proteoglycans. GAGs interact with growth factors, morphogens, chemokines, extracellular matrix proteins and their bioactive fragments, receptors, lipoproteins, and pathogens. These interactions mediate their functions, from embryonic development to extracellular matrix assembly and regulation of cell signaling in various physiological and pathological contexts such as angiogenesis, cancer, neurodegenerative diseases, and infections. We give an overview of GAG–protein interactions (i.e., specificity and chemical features of GAG- and protein-binding sequences), and review the available GAG–protein interaction networks. We also provide the first comprehensive draft of the GAG interactome composed of 832 biomolecules (827 proteins and five GAGs) and 932 protein–GAG interactions. This network is a scaffold, which in the future should integrate structures of GAG–protein complexes, quantitative data of the abundance of GAGs in tissues to build tissue-specific interactomes, and GAG interactions with metal ions such as calcium, which plays a major role in the assembly of the extracellular matrix and its interactions with cells. This contextualized interactome will be useful to identify druggable GAG–protein interactions for therapeutic purpose:
Keywords
Introduction
The six mammalian glycosaminoglycans (GAGs), namely, chondroitin sulfate (CS), dermatan sulfate (DS), heparin (HP), heparan sulfate (HS),1,2 hyaluronan (HA),
3
and keratan sulfate (KS),4,5 are linear polysaccharides made of disaccharide repeats. These disaccharides are composed of an uronic acid and an hexosamine,
6
except for KS, which lacks the uronic acid and consists of a sulfated poly-N-acetyl lactosamine chain made of
The GAG chains, ranging from a few kDa to over a 100 kDa, contribute to the physicochemical properties and biological functions of proteoglycans.6,10,11 HS, for example, affects the conformation, stability, and diffusion of proteins. 1 Other unique sulfated GAGs with biological functions such as anticoagulant activity and neurite outgrowth–promoting activity exist in primitive chordates, but they are not discussed in this review.12,13
GAGs play numerous biological roles, ranging from embryonic development,14,15 extracellular matrix (ECM) assembly to the regulation of cell signaling1,6 in various physiological and pathological contexts such as angiogenesis,16,17 cancer,18,19 inflammation,18,20 neural development, 21 neurodegenerative diseases,22,23 and host–pathogen interactions. 24 These roles are mediated by the interactions of GAGs with a variety of proteins, including soluble proteins (growth factors, morphogens, and chemokines25,26), ECM proteins and bioactive fragments, membrane receptors such as integrins,27,28 and lipoproteins. 29 GAGs also bind to numerous pathogens, including viruses, 30 bacteria, 31 parasites,32,33 and fungi.24,34 The interaction of HS with a number of viruses contributes to their attachment to the host cells or to their increased concentration at the cell surface, whereas HS proteoglycans may serve as entry receptors for some viruses. 35 It has been recently reported that HP binds to the spike glycoprotein of severe acute respiratory syndrome coronavirus (SARS-CoV), Middle-East respiratory syndrome coronavirus (MERS-CoV), and to both monomeric and trimeric spike of SARS-CoV-2, responsible for the COVID-19 pandemic. 36 The interaction between HP and the SARS-CoV-2 Spike S1 protein is mediated by the receptor-binding domain of the protein. 37 GAG–protein interactions may be targeted for therapeutic purpose as shown for HP– and HS–protein interactions38,39 and microbial GAG–protein interactions. 40
We give here an overview of the molecular characterization of GAG–protein interactions (i.e., specificity and chemical features of GAG and protein-binding sequences), and review the available GAG–protein interaction networks. We also provide the first comprehensive draft of the GAG interactome composed of 827 proteins and 932 protein–GAG interactions. This network is a scaffold, which in the future should integrate structures of GAG–protein complexes, quantitative data on the abundance of GAGs in tissues to build tissue-specific interactomes, and GAG interactions with metal ions such as calcium, which plays a major role in the ECM assembly and in ECM–cell interactions. This contextualized interactome will be useful to identify druggable GAG–protein interactions for therapeutic purpose. The major perspectives and challenges to overcome to reach this goal are presented in the conclusion.
Glycosaminoglycan–Protein Interactions
The methods to identify and characterize GAG interactions were recently reviewed. 6 They include label-free techniques monitoring interactions in real time such as surface plasmon resonance and bio-layer interferometry, 41 solid-phase binding assays, protein arrays probed with GAGs, 42 and GAG arrays probed with proteins.41,43-45 Very large interaction data sets are not available for GAG interactions because high-throughput techniques used to collect interaction data are gene-centric (two hybrid approaches) or protein-centric (affinity-purification-mass spectrometry). Libraries of GAG-like oligosaccharides have been synthesized either chemically and/or enzymatically, and may be used to design GAG microarrays for studying GAG–protein interactions. 43 A cell-based library of displayed GAGs, the GAGOme, has been developed for cell-binding assays and the assembly of GAG microarrays, and the identification of further GAG-binding partners. 46 A number of GAG–protein interactions experimentally supported has been manually curated according to the curation rules of the Inter-national Molecular Exchange (IMEx) consortium 47 and stored in the publicly and freely available interaction database MatrixDB (http://matrixdb.univ-lyon1.fr/) that we have developed.48-50 MatrixDB, the ECM interaction database,48,49 contains GAG–protein and protein–protein interaction data manually curated from literature, and has been recently updated with a focus on GAGs. 50
GAG–protein interactions are driven by the features of GAGs (chemical groups, length, and conformation) and of their protein partners (amino acid residues in linear sequences or three-dimensional [3D] structures). The characterization of binding interfaces is required to decipher the molecular mechanisms underlying GAG–protein interactions, their specificity, and their biological functions. The various modes of interaction between HS and its protein partners have been reviewed.6,29,51 The basic consensus sequences, X-B-B-X-B-X and X-B-B-B-X-X-B-X, where B is an arginine or lysine residue, and X is a hydrophobic amino acid, identified by Cardin and Weintraub 52 and called CW motifs, are found in HP- or HS-binding proteins. However, the network analysis of the amino acid sequences of 437 HP-binding proteins has shown that HP-binding sequences may be composed of numerous, short, spaced, basic amino acid sequences forming in a concerted fashion a 3D arrangement on the protein surface rather than linear basic sequences. 53 The fact that HP-binding sites are only made of stretches of contiguous arginine and lysine residues has also been challenged by the finding that numerous ionic and non-ionic amino acid residues contribute to the specificity of GAG-binding sites. The uncharged, polar, residues, asparagine or glutamine, are involved in the formation and specificity of GAG-binding sites. Highly specific and nonspecific GAG–protein interactions can be discriminated by identifying “hot spots” in GAG-binding sites. (i.e., neutral loci among arginine/lysine residues that present high electrostatic potential). 38 This will have direct applications in the identification of druggable GAG–protein interactions, the design of GAGs and GAG mimetics as potential drugs, and in the identification of the GAG interactome by computational analysis. 38 HP-binding sites may be partially disordered, and we have shown that the association rate of proteins with HP is positively correlated with the percentage of disordered residues in HP-binding sites. 54
More than 75 GAG–protein complexes are available in the Protein Data Bank, 7 and the structural biology of the HP interactome based on GAG–protein complexes has been reviewed. 55 When no structural experimental data are available, and when the sequence of GAG oligosaccharides interacting with proteins is known, it is possible to build their 3D models, and to use them for docking onto the 3D structure of their protein partners to determine the chemical features of GAGs and amino acid residues (or domains) of proteins involved in the interactions. Coarse-grained models of GAGs have been generated,56,57 and quantum mechanics, molecular mechanics and dynamics, coarse graining and docking methods have been used for the multiscale modeling of GAG structure, from monosaccharides to GAG chains. 58
We have designed a pipeline to build 3D models of GAG oligosaccharides from their GlycoCT codes, a machine-readable encoding scheme for complex carbohydrates, 59 with the GAG builder tool we developed. This tool is based on a repertoire of conformations experimentally validated by data extracted from GAG–protein crystals. 7 A converter (the CT23D converter) automatically translates the GlycoCT code of a GAG sequence into the input file required for GAG builder, 7 which is freely available on the web site of MatrixDB database. 50 We have shown that the Φ and Ψ torsion angles of the HP/HS and HA oligosaccharides co-crystallized with proteins are clustered into two major areas, emphasizing the role of GAG conformation in protein binding. 7
The sulfation patterns of GAGs, referred to as the sulfation codes, are key features for the molecular recognition of GAG partners. Alterations of the degree of GAG sulfation are often associated with disorders such as degenerative diseases and malignant neoplasms. 60 The chemical modifications of HS chains govern their specific interactions with a number of proteins, but their organization into sulfated and N-acetylated domains also contributes to the specificity of protein interactions. 29 The role of HS organization has been shown for its interaction with endostatin, the C-terminal fragment of collagen XVIII, which binds to a 10/12-mer containing two N-sulfated regions separated by at least one N-acetylated glucosamine unit. 61
Another parameter contributing to the specificity of GAG–protein interactions is the internal mobility of GAG chains. The internal mobility of sulfated domains of HS chains is mostly due to the conformational flexibility of iduronic acid residues, but the sulfated domains are less flexible than the non-sulfated ones. 62 The organization of HS chains in domains of variable flexibility allows multiple interactions to occur along the same chain.
Docking combined with molecular dynamics simulations has been used to investigate GAG–protein complexes 63 such as those formed with growth factors (e.g., vascular endothelial growth factor-A-HP), 64 with ECM proteins such as those formed by procollagen C-proteinase-enhancer-1 with several GAGs 65 or by ECM bioactive fragments such as the propeptide of lysyl oxidase with a HP hexasaccharide. 66 A benchmarking study, including eight docking softwares for 28 GAG–protein complexes, has concluded that the combination of several molecular docking programs and other modeling approaches such as molecular dynamic improves the prediction and evaluation of GAG-binding poses. 67 Computational approaches are valuable tools to address the following points: “does a protein bind to GAG,” “where does the GAG bind,” “is the GAG–protein complex stable,” and “what is the most optimal GAG binding sequence” as summarized in the flowchart designed by Sankaranarayanan et al. 68 The affinity and kinetics of GAG interactions also provide information. Matrisome-associated proteins (e.g., growth factors) form transient complexes with HP but bind to this GAG with a higher affinity than core matrisome proteins. 54 Kinetics and affinity can be used to rank interactions in networks. We have integrated them in the interaction network of an endogenous angiogenesis inhibitor, endostatin, which is a HP-binding protein interacting with other HP-binding proteins at the surface of endothelial cells. 54
The First Draft of the Glycosaminoglycan Interactome
Several networks focused on a specific GAG, a subset of GAG-binding proteins, or a biological process (e.g., angiogenesis as described above) 54 have been built. HP and HS interactions with angiogenesis regulators have been collected to create the “angiogenesis glycomic interactome” comprising 56 proteins, including growth factors, proangiogenic receptors, angiogenesis inhibitors, and effectors. 16 An interaction network of HP/HS was built in 2018 using MatrixDB data. It contained 262 interactions of HP/HS with ECM proteins, ECM bioactive fragments, cytokines, growth factors, proteins from the complement system, membrane proteins such as integrins, and intracellular proteins. 6 The most comprehensive HP/HS available so far was the HP/HS interactome containing 435 human proteins binding to HP or HS, which was generated by combining data from the literature and affinity proteomics experiments performed on plasma membrane–enriched fractions isolated from rat liver. 69
MatrixDB data have also been used to build an interaction network comprising five of the six mammalian GAGs, 135 proteins, and 211 GAG–protein interactions. 41 In this network, 60.5% of HP partners, 38.5% of HA partners, and 20% of CS partners bound only to HP, HA, and CS respectively, whereas none of the DS partners interacted only with DS. 41 HP was thus the GAG with the highest number of specific protein partners, and DS had no specific protein partners. In contrast, less than 2% of HP partners interacted with five GAGs, whereas 13% to 15% of CS, DS, and HA partners bound to five GAGs. 41
GAGs have been reported to bind to more than 500 proteins. 38 To generate the first comprehensive draft of the mammalian GAG interactome, we used the roadmap we designed to build extracellular and pericellular protein–GAG interaction networks. 70 Several sources of interaction data were combined to build the protein network of the six mammalian GAGs (i.e., CS, DS, HS/HP, HA and KS). HP and HS interactions were clustered because these GAGs share the same disaccharide unit, and HP is often used as a model for HS. Partners of KS (217 proteins), CS A (24 proteins), and HA (six proteins) identified using protein arrays and biotinylated GAGs, 42 and of HP/HS identified by affinity chromatography-mass spectrometry 69 were included in the GAG interaction data set together with the CS-binding proteins identified in the central nervous system. 71 Further manual curation of the literature was performed with a focus on ECM proteins. UniProtKB 72 and Gene Ontology (GO) database 73 were queried using keywords and GO terms related to GAG binding (e.g., KW-0358 heparin binding, and GO:1904399 heparan sulfate binding).
The resulting network is the first comprehensive interactome of the six mammalian GAGs (HP and HS being clustered). It comprises 832 biomolecules (827 proteins and five GAGs) and 932 protein–GAG interactions (Fig. 1). The experimental settings used to identify interactions are detailed in the articles listed by their PubMed reference number (PMID) in Supplementary Table S1. When an interaction has been reported in several studies, all the publications are listed. HP/HS, which are the most extensively studied GAGs; have 580 partners, KS and CS bind to 218 and 72 proteins, respectively; and HA and DS have the lowest number of partners (43 and 19, respectively). About 89% of the HP-/HS-binding proteins interact only with HP/HS, 85% of KS-binding proteins interact only with KS, 63% of HA partners bind only to HA, and 26% of CS partners bind specifically to CS (Fig. 2). In contrast, none of the partners of DS binds specifically to it (Fig. 2) as previously reported for a smaller GAG network. 41 HP, HS, and KS have thus the highest number of specific protein partners, whereas DS has no specific protein partners but has the highest percentage of partners able to bind to four (26%) or five (5%) GAGs. However, it is difficult to draw firm conclusions on the specificity of the GAG partners given that binding assays have been performed with different sets of proteins for each GAG, and that negative results (i.e., lack of binding to a protein) are not recorded in most databases. This emphasizes the need to design proteome-wide binding assays to increase GAG interactome coverage.
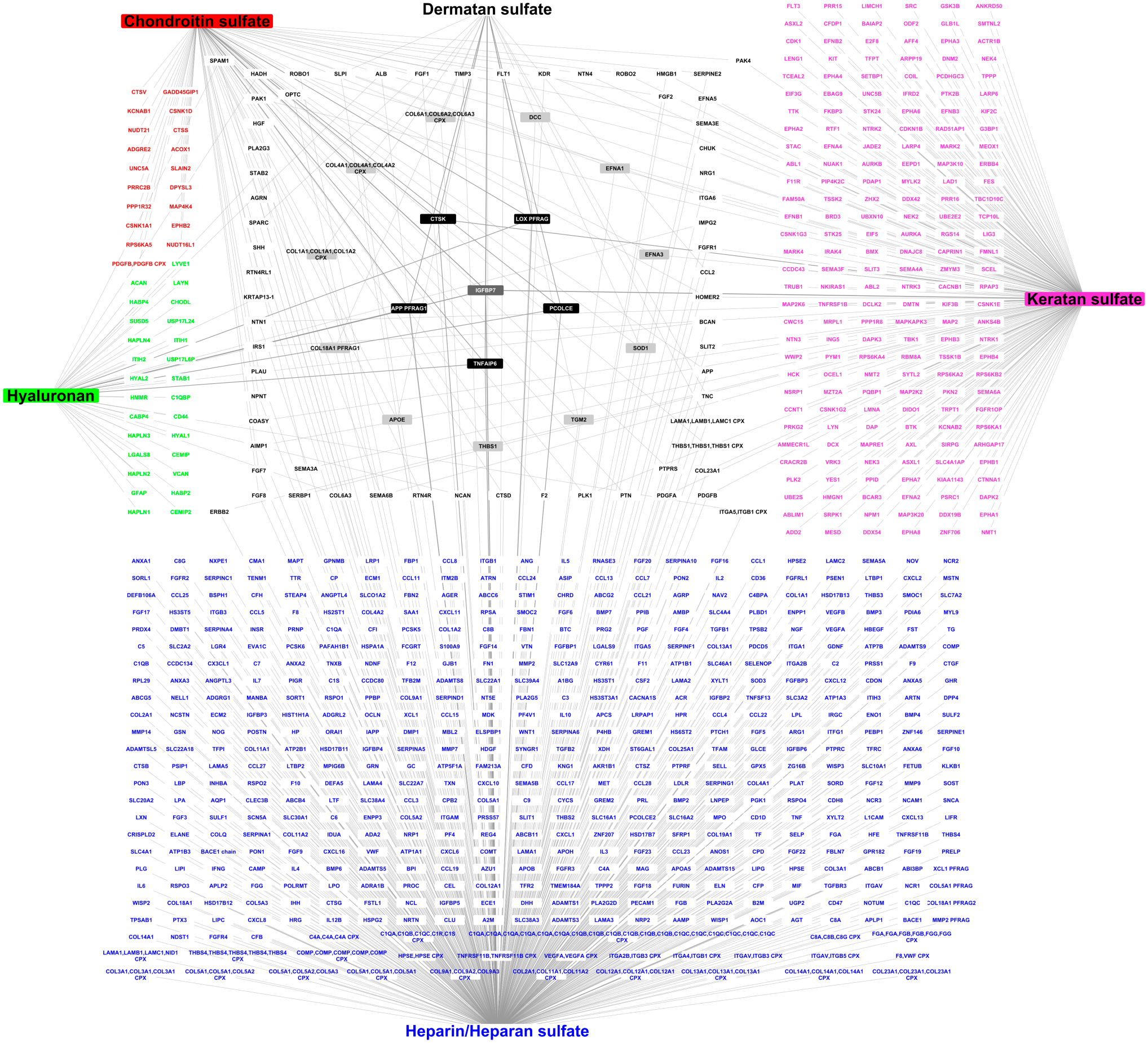
The interactome of the six mammalian glycosaminoglycans (GAGs) comprising 827 proteins and 932 GAG–protein interactions (72 for chondroitin sulfate, 19 for dermatan sulfate, 580 for heparin/heparan sulfate, 43 for hyaluronan, and 218 for keratan sulfate). The partners of GAGs are displayed according to the number of GAGs they interact with. The colored nodes, located at the periphery of the network, bind only to the GAG with the same color, the square correspond to proteins interacting with two GAGs, and the two inner circles correspond to proteins interacting with three (light gray) and four (dark) GAGs. The protein, which binds to all GAGs, is at the center of the network (CPX: complex, i.e., multimeric proteins, PFRAG: bioactive protein fragments, e.g., COL18A1 PFRAG1: endostatin).
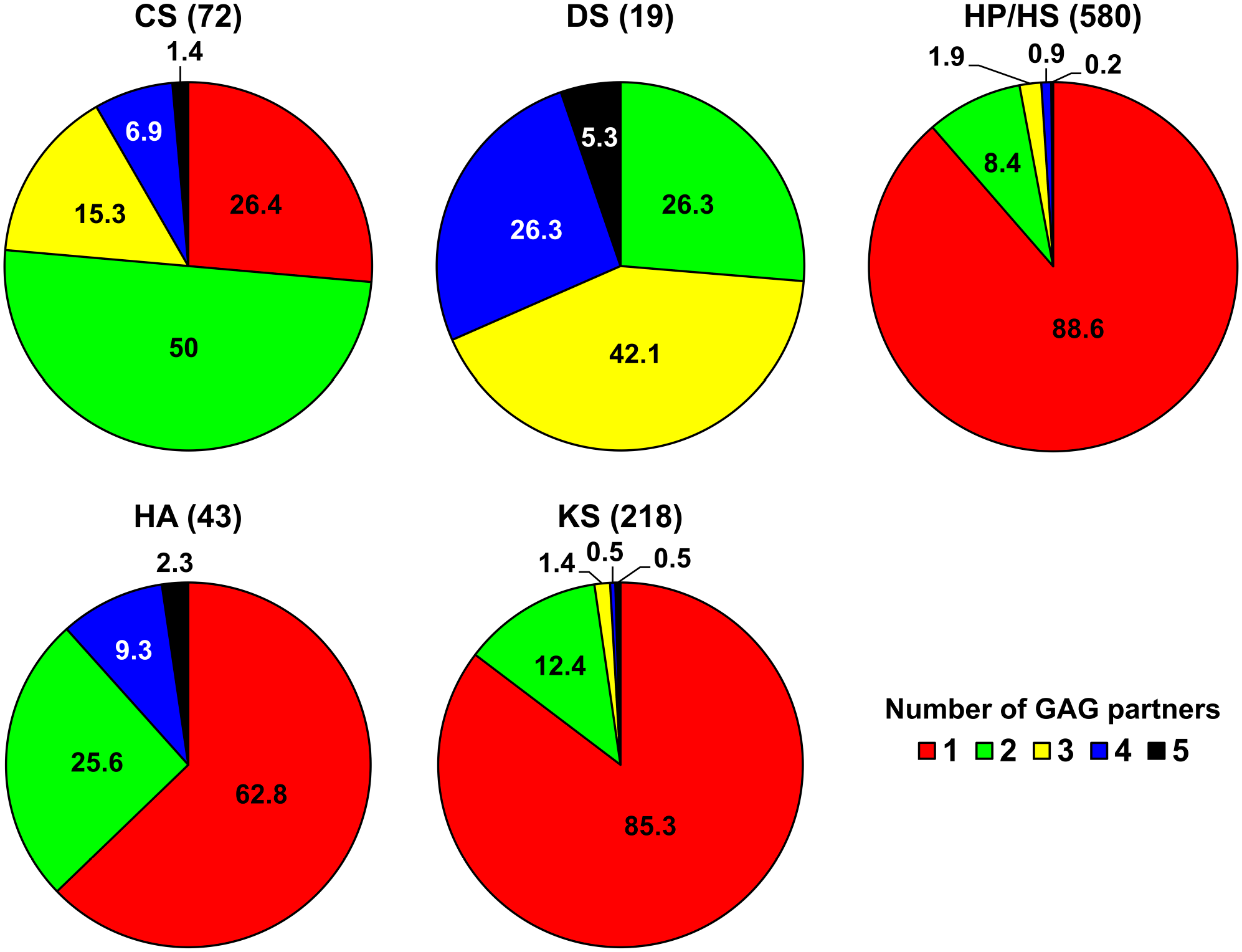
Specific and common protein partners of the six GAGs. The percentages of partners, which bind to one (red), two (green), three (yellow), four (blue) or five (black) GAGs, are indicated for each GAG. The total number of partners of each GAG is indicated in brackets. Abbreviations: GAGs, glycosaminoglycans; CS, chondroitin sulfate; DS, dermatan sulfate; HA, hyaluronan; HP, heparin; HS, heparan sulfate; KS, keratan sulfate.
The number of partners common to GAG pairs is displayed in Fig. 3. Although HA is not sulfated, it has eight partners in common with HP/HS, which are highly sulfated. The proteins binding to three, four, of five GAGs are displayed in Fig. 1 and Supplementary Table S2. Eleven proteins bind to three GAGs, including ECM proteins (collagens I, IV, and VI, and thrombospondin-1), a matricryptin of collagen XVIII (endostatin), and proteins involved in nervous system development (netrin receptor DCC, and ephrins A1 and A3). Five proteins interact with four GAGs, an ECM protein, procollagen C-proteinase enhancer-1, cathepsin K, tumor necrosis factor–inducible gene 6 protein, a multifunctional protein with anti-inflammatory and tissue-protective properties, 74 a β-amyloid peptide, and the propeptide of lysyl oxidase (Fig. 1 and Supplementary Table S2). A single protein, the insulin-like growth factor–binding protein 7, binds to all GAGs (Fig. 1 and Supplementary Table S2). Because the partners of GAGs may interact with each other in vivo, we have added them to build a comprehensive GAG network, including not only GAG–protein interactions but also protein interactions between GAG partners. Protein–protein interactions were collected by manual curation of the literature, and by querying MatrixDB, the Adhesome network (http://adhesome.org) 75 and the Bioplex 2.0 data set. 76 The resulting network comprises 827 proteins and 2240 interactions (Supplementary Fig. S1 and Supplementary Table S1).
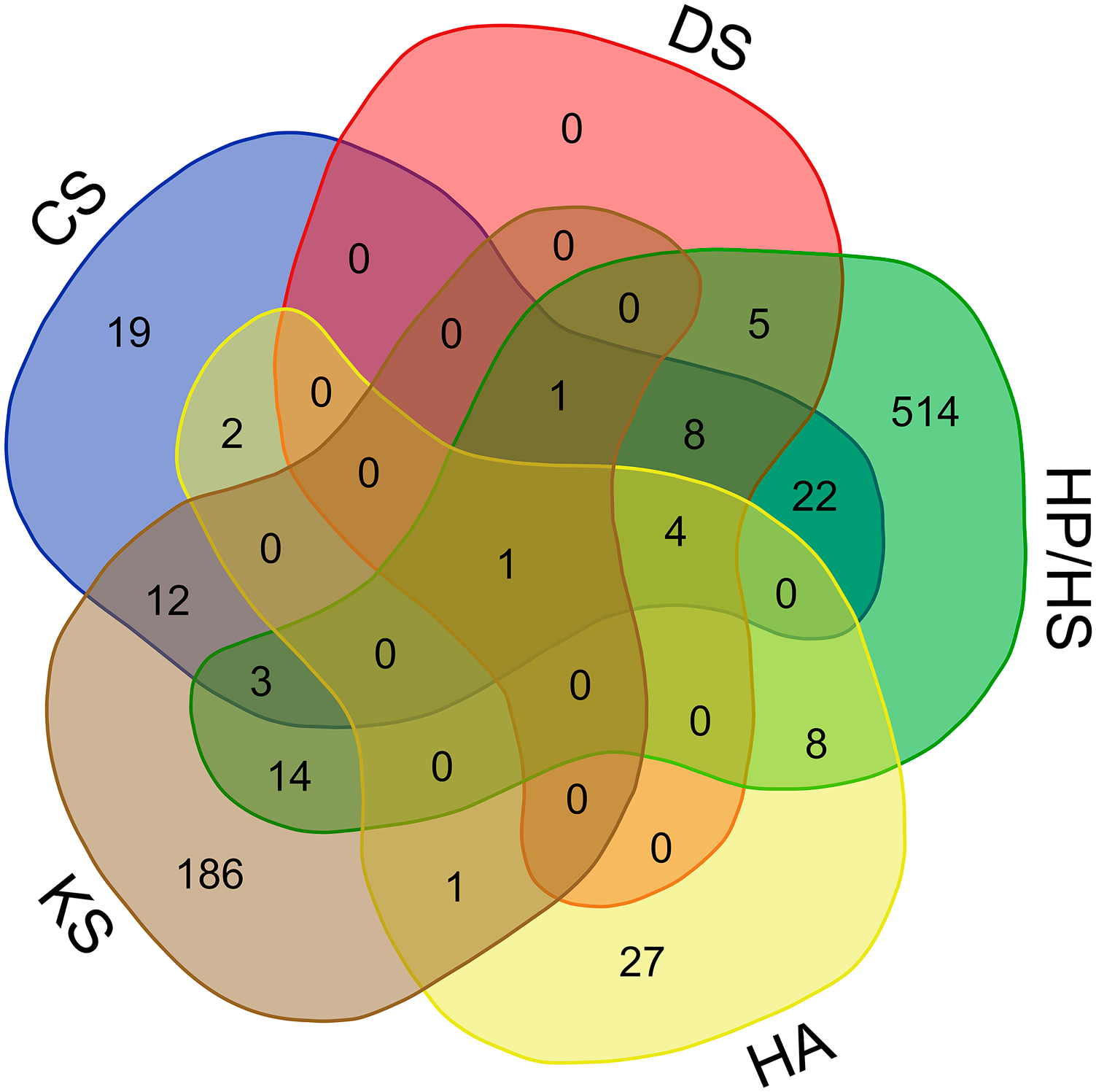
The Venn diagram showing the number of specific partners of individual GAGs, and the number of protein partners shared by two, three, four, or five GAGs. Partners interacting with three, four, or five GAGs are listed in Supplementary Table S2. The diagram was built with a web-service (http://bioinformatics.psb.ugent.be/webtools/Venn/). Abbreviations: GAGs, glycosaminoglycans; CS, chondroitin sulfate; DS, dermatan sulfate; HA, hyaluronan; HP, heparin; HS, heparan sulfate; KS, keratan sulfate.
Enrichment analyses of the GAG interactome were carried out using the functional enrichment analysis tool FunRich, and FunRich database as background. 77 The full report of these analyses is included in Supplementary Table S3. They show that 55% of GAG partners are extracellular, 30% are located in plasma membranes, 27% in exosomes, and 20% in lysosomes. These cellular component annotations are significantly enriched in the GAG interactome (p<0.001). Several molecular functions are also significantly enriched in GAG partners (p<0.001), but they concern less than 10% of them (ECM structural constituent: 8%, serine/threonine kinase activity: 5%) or less than 5% of them. No protein domain was significantly overrepresented in numerous GAG partners, except for the presence of a signal peptide in 62% of GAG partners (p<0.001), indicating that most GAG partners are secreted proteins. The epidermal growth factor domain is the next most abundant domain in GAG partners, but it is present in only 7% of them (p<0.001), followed by the tyrosine kinase, catalytic domain (TyrKc; 5%, p<0.001).
The biological processes significantly enriched (p<0.001) in the GAG interactome are signal transduction (36% of the GAG partners), cell communication (34%), cell growth and/or maintenance (12%), and immune response (10%). Pathway analysis was then performed using Reactome.78,79 The major significant pathways identified by Reactome are displayed in Fig. 4, and the 25 most relevant pathways sorted by p value (10−16<p<10−13) are listed in Supplementary Table S4. They include pathways related to ECM organization (assembly of collagen fibrils and other multimeric structures, collagen and ECM degradation, collagen formation and chain trimerization, ECM proteoglycans, non-integrin membrane–ECM interactions, and integrin cell surface interactions), signal transduction (phospholipase C-mediated cascade, FGFR2 [fibroblast growth factor receptor 2], FGFR2 ligand binding and activation, PI3K/AKT signaling, PI3K/AKT signaling in cancer, chemokine receptors and signaling by receptor tyrosine kinases), hemostasis (platelet degranulation, and response to elevated platelet cytosolic Ca2+), developmental biology (nervous system development, axon guidance, EPH–ephrin-mediated repulsion of cells), and metabolism of proteins (posttranslational protein phosphorylation, and regulation of insulin-like growth factor [IGF] transport and uptake by IGF-binding proteins).
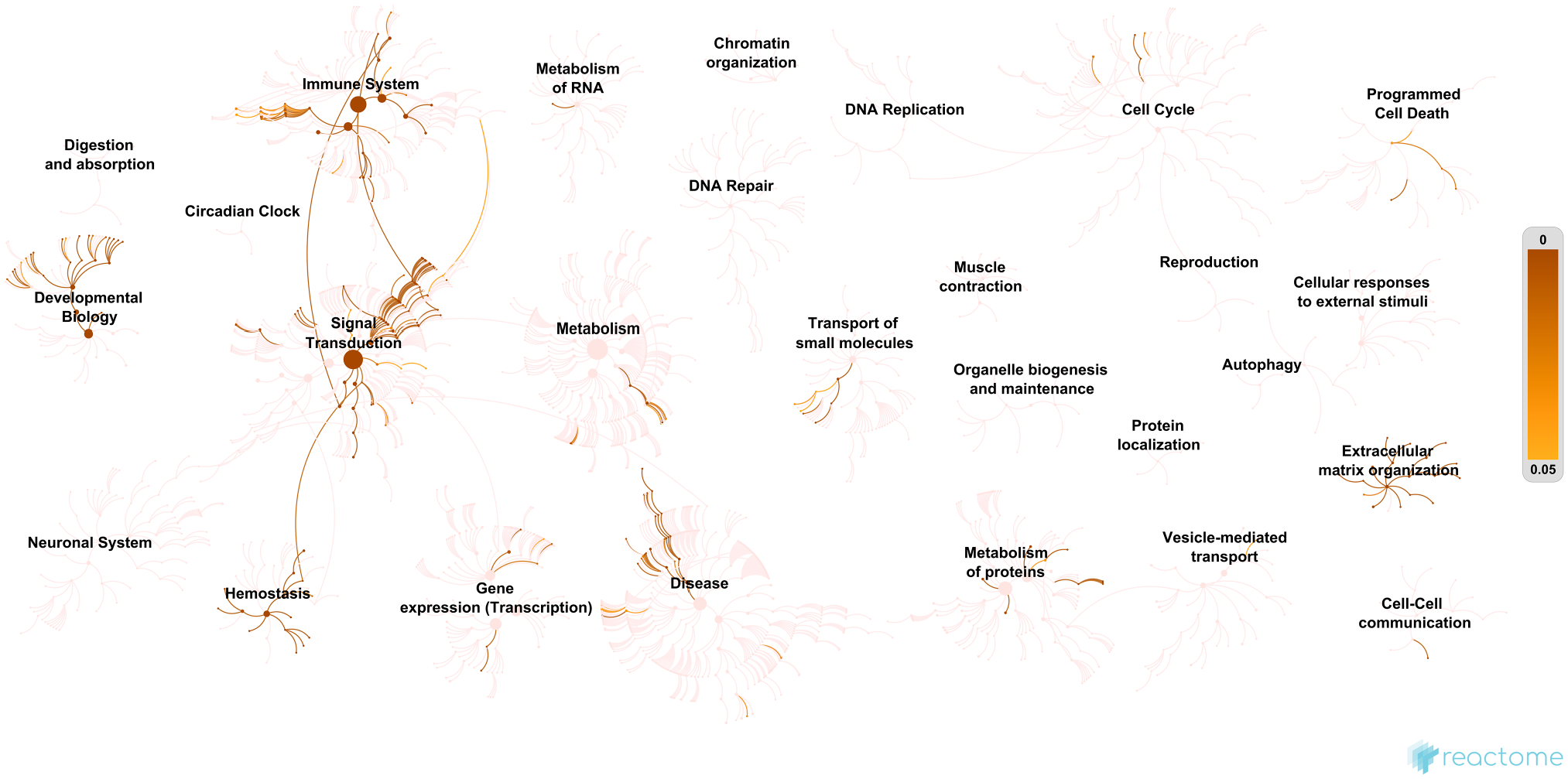
A human genome-wide overview of the results of the pathway analysis of the glycosaminoglycan (GAG) interactome using Reactome.78,79 “Reactome pathways are arranged in a hierarchy. The center of each of the circular ‘bursts’ is the root of one top-level pathway. Each step away from the center represents the next level lower in the pathway hierarchy.” 78 The size of the nodes is proportional to the number of physical entities (proteins, complexes, chemicals) it contains. The color code denotes overrepresentation of pathways. Light color indicates that the corresponding pathways are not significantly overrepresented in the GAG interactome.
Several transcription factors regulating the expression of GAG partners are significantly enriched (p<0.001) in the GAG interactome, including hepatocyte nuclear factor 4-alpha (27%), nuclear factor 1 C-type (21%), and transcription factor E2-alpha involved in the initiation of neuronal differentiation (TCF3, 19%) (Supplementary Table S3). The analysis of GAG partners using FunRich and the Catalogue of Somatic Mutations in Cancer, a comprehensive resource for exploring the effect of somatic mutations in human cancer, 80 shows a significant enrichment (p<0.001) in somatic mutations associated with the central nervous system (74%), skin (71%), prostate (60%), and hematopoietic and lymphatic tissues (56%).
In conclusion, the first draft of the GAG interactome is a scaffold which should integrate further data in the future. The integration of quantitative data on the presence/abundance of GAGs in healthy and diseased tissues or biological fluids (i.e., glycosaminoglycomic data) 81 would allow us to generate tissue-specific and disease-specific GAG networks. Methods and standards to analyze GAGs are under development, and will speed up the collection and use of glycosaminoglycomic data in the coming years.82-84 Structural analyses have been done for individual complexes 55 but not at the GAG interactome scale. Experimental data are available in the Protein Data Bank for about 75 GAG–protein complexes, 7 but additional data could be generated using docking experiments when the 3D structure of GAG-binding proteins is known. Three-dimensional models of GAG sequences binding to proteins could also be built using several approaches as described above. This would be useful to decipher the glycocode resulting from the combinatorial of chemical groups, mostly sulfate groups, along GAG chains.
It would be interesting to include the interactions of GAG–metal ion interactions in the next version of the GAG interactome. Indeed, GAGs can bind to metal ions associated with proteins or to metal ions in solution, and metal ions can modulate the specificity and affinity of protein–GAG interactions. 85 The binding of prion protein to GAGs is stabilized by Cu2+or Zn2+ ions, and the interaction of endostatin with HS is increased in the presence of Zn2+ ions. 86 In addition, HP binds to calcium, 85 which plays a major role in ECM assembly and functions, 87 and in ECM–cell interactions 88 as reported for syndecans, which interact with transient receptor potential channels involved in Ca2+ entry mechanisms and can regulate them. 89 Here again, modeling approaches could provide new insights on the effects of ions on protein–GAG interactions as shown for the complex formed by the ECM protein procollagen C-proteinase enhancer-1 and GAG oligosaccharides. 65 It would be also crucial to determine the role of the proteoglycan protein cores in GAG–protein interactions to better understand the mechanisms of GAG–protein interactions in vivo. Last, partners of the unique sulfated GAGs found in primitive chordates, which have important biological functions such as anticoagulant activity and neurite outgrowth promoting activity,12,13 should be added to the mammalian GAG interactome.
Supplemental Material
Supplemental Tables S1
List of glycosaminoglycan-protein interactions and protein-protein interactions between the protein partners of glycosaminoglycans included in the comprehensive GAG interactome.
Supplemental Material
Supplemental Tables S2
List of proteins binding to three, four or five glycosaminoglycans.
Supplemental Material
Supplemental Tables S3
Enrichment analysis report of the GAG interactome using FunRich.
Supplemental Material
Supplemental Tables S4
List of the 25 most relevant pathways sorted by p-value identified by pathway analysis of the GAG interactome using Reactome.
Supplemental Material
Supplemental Figure S1
The interactome of the six mammalian glycosaminoglycans (GAGs) including GAG-protein interactions and interactions between GAG protein partners.
Footnotes
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
SDV performed literature curation and database searches, and designed the figures; OC retrieved glycosaminoglycan (GAG)–protein interactions from Gene Ontology and UniProtKB, assembled the interaction data set, and implemented new GAG features in MatrixDB database; and SR-B manually checked retrieved GAG interaction data, performed enrichment and pathway analyses of the GAG interactome, and wrote the manuscript. All authors have read and approved the final manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work performed by the authors was supported by the “Fondation pour la Recherche Médicale,” France (Grant No. DBI20141231336 to S.R.-B.), the Institut Français de Bioinformatique (ANR-11-INBS-0013), Glycomatrix project, call 2015 to S.R.-B. and O.C., and the Groupement de Recherche (GDR) GagoSciences (CNRS, GDR 3739, Structure, Fonction et Régulation des Glycosaminoglycanes to S.R.-B., S.D.V., and O.C.).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.