
Abstract
MicroRNAs (miRs) regulate endometrial function and their dysregulation could underlie unexplained infertility in women. Ribonucleases including DICER and DROSHA, and the proteins, ARGONAUTE 1 (AGO 1) and 2 (AGO 2) regulate the biogenesis/maturation of miRs. We aimed to elucidate the expression and localization of miR biogenesis machinery components during the human menstrual cycle and compare their levels in endometrium from women with normal fertility and primary unexplained infertility. miR biogenesis components were measured by quantitative-RT-PCR and immunohistochemistry. In the endometrium of women with normal fertility, DROSHA immunolocalized maximally to the epithelium during the early and mid-secretory phases compared with the proliferative and late-secretory phases. Stromal DICER immunostaining intensity was higher in the late-secretory phase compared with all other phases in fertile women. DROSHA mRNA was reduced in the mid-secretory-infertile whole endometrial tissue (has all cells of the tissue), and primary epithelial and stromal cells while no differences were found in DICER, AGO1, and AGO2 mRNA. In the luminal epithelium, DROSHA staining intensity was reduced in early and mid-secretory-infertile while DICER staining was reduced in the early secretory-infertile compared with their respective fertile groups. DICER and DROSHA were dynamically regulated across the menstrual cycle and reduced levels during receptivity phase could underlie implantation failure/infertility.
Introduction
Implantation failure is a major factor underlying infertility and low pregnancy success rates in women conceiving naturally and via assisted reproductive technologies such as in vitro fertilization (IVF). For pregnancy to be achieved, successful blastocyst implantation has to occur via a blastocyst firmly adhering on an adequately receptive endometrial uterine luminal epithelium. The endometrium undergoes extensive remodeling during each menstrual cycle and becomes receptive during the mid-secretory phase, within approximately days 19 to 23 of a 28-day menstrual cycle. Therefore, the uterine microenvironment is important to provide the necessary regulatory molecules and microvesicles during endometrial remodeling and any abnormalities could impair embryo implantation, causing infertility. Some known regulators include leukemia inhibitory factor (LIF), vascular endothelial grown factor (VEGF), and Interleukin 11 (IL-11).1–4
More recently, studies have identified a widespread alteration of small non-coding RNA, microRNAs (miRs) in the endometrium of infertile women5–8 and in embryos.9–13 miRs are small, non-coding 22 to 24 long nucleotides generated by endogenous hairpin-shaped transcripts. 14 miRs control gene expression silencing which leads to inhibition of target genes involved in numerous pathways such as cell differentiation, apoptosis, and cell proliferation. 15 The biogenesis mechanism of miRs is strictly controlled by RNA machinery, in particular, DICER and DROSHA. DROSHA is an RNase III enzyme and together with its partner DiGeorge syndrome critical region 8 (DGCR8) is essential for miR maturation. In the cytoplasm, the second RNase III endonuclease DICER takes another cut and mature miR is loaded into the Argonaute family transporting proteins (i.e., AGO1 and AGO2). Animal studies have demonstrated that miR biogenesis molecules might play a role in unexplained fertility16–20; however, the expression levels of miR biogenesis molecules in relation to human fertility remain elusive. The aim of this study was to determine the spatial and temporal expression of miR biogenesis molecules in endometrium across the human menstrual cycle and to investigate whether their gene and protein expression levels were associated with fertility status in women. We hypothesized that the expression of miR biogenesis molecules is dysregulated in the endometrium from women with unexplained infertility compared with normal fertility (controls).
Methods
Primary Tissue Collection
Endometrial tissue was collected at Monash Medical Centre (Melbourne, Australia) from women across the menstrual cycle (proliferative and early, mid-, and late-secretory tissue). Written informed consent was obtained from each patient, with protocols approved by and in accordance with the Monash Health Human Research Ethics Committee, Melbourne, Australia. Fertile women (n=5–10/phase of the menstrual cycle) had proven parity, while infertile women (n=5–10/group) had primary unexplained infertility. The women had received no steroid hormone treatment for at least 3 months before tissue collection. The collected tissues were examined by an experienced gynecological pathologist to confirm cycle stage and absence of endometrial dysfunction.
Endometrial Epithelial and Stromal Cell Isolation
Primary human endometrial epithelial cells (HEEC) and stromal cells (HESC) were isolated from whole endometrial tissue as previously published (n=5–10/group)21–23 and cultured in DMEM/F12 (Thermo Electron Corporation, Australia) medium with 10% fetal calf serum (Invitrogen, USA) and 1% antibiotic-myotic solution (Gibco, North Shore, New Zealand).
RNA Isolation and Real-time Quantitative PCR
RNA was isolated from HEEC, HESC, or whole endometrial tissue biopsies using TRI Reagent (Sigma-Aldrich, Darmstadt, Germany) following the manufacturer’s instructions. RNA concentrations were determined using NanoDrop spectrophotometry. RNA was then reverse transcribed using the Superscript III First strand synthesis kit (Invitrogen) to yield complementary DNA. Real-time qPCR was then performed using the SYBR Power (Applied Biosystems) in combination with specific oligo primer pairs (Sigma) as listed in Supplementary Table 1. Expression levels were normalized to 18S for target gene analysis. Relative expression levels were calculated using comparative cycle threshold method (ΔΔCt).
Immunohistochemistry
Endometrial tissue was fixed in neutral-buffered formalin (10%) and embedded in paraffin. Sections were dewaxed in xylene and rehydrated in graded ethanol. Antigen retrieval was performed with heated citrate buffer (pH 6.0) in a microwave at 80 watts for 5 min before an endogenous peroxidase block using 3% hydrogen peroxide in methanol. Following Tris buffered saline (TBS) washes, non-immune blocking was completed with 10% goat serum and 2% human serum in TBS for 45 min at room temperature, under agitation. DROSHA antibody (Cell Signaling #3364P) was added at 8.0 µg/ml and DICER (Cell Signaling #5362P) was added 1.39 µg/ml overnight at 4C, with a concentration matched rabbit IgG (Dako) for each as negative control to show specificity of the immunohistochemical staining. Sections were washed with TBS Tween 0.6% and TBS, subsequently sections were incubated in biotinylated goat anti-rabbit secondary antibody (Vector) for 30 min at room temperature. Avidin-biotin complex (Vector) was added for 30 min at room temperature following extensive TBS Tween 0.6% and TBS washes. Diaminobenzidine (DAB; Dako) was added and sections were immersed in distilled water as brown staining appeared under light microscopy. Sections were counterstained with haematoxylin, dehydrated through graded ethanol, and cleared with xylene before coverslips were adhered using DPX. Staining intensity scores were determined by scorers blinded to the patient characteristics as previously published. 24 , 25 A score of 0 denoted no positive brown DROSHA or DICER staining and 3 was intense staining. Scores were averaged and plotted using Graphpad Prism 7. Images were captured using Olympus light microscope and CellSens software. Scale bars were added in Fiji Image software.
Statistical Analysis
All values are expressed as the mean ± SEM (standard error of the mean). All data were analyzed using tools within Graphpad Prism 7. The one-way analysis of variance (ANOVA) and Tukey’s post hoc tests were used to analyze changes in DROSHA and DICER expression across the menstrual cycle. The two-tailed unpaired Student’s t-test was used to compare miR machinery expressions between fertile and infertile groups. Probability level of 5% (p<0.05) was considered significant for all statistical tests.
Results
DROSHA and DICER Protein Expression in the Endometrium Are Altered Across the Menstrual Cycle
To determine endometrial DROSHA and DICER protein expression levels across the menstrual cycle, DROSHA and DICER immunostaining were assessed in fertile and infertile endometrium tissue. In the fertile endometrium, DROSHA staining was significantly higher during the early and mid-secretory phases compared with both the proliferative and late-secretory phases in the luminal or endometrial surface epithelium basal lamina (p<0.05, Fig. 1A). In parallel, DROSHA staining was significantly higher in the glandular epithelium membrane in the mid-secretory phase compared with the late-secretory phase in the fertile endometrium (p<0.05, Fig. 1A). In the infertile endometrium, DROSHA staining was significantly higher in the basal lamina of the luminal epithelium during the mid-secretory phase compared with the late-secretory phase (p<0.05, Fig. 1B). DROSHA staining of the basal laminar of the glandular and endometrial epithelium that opens up into the endometrial lumen was significantly higher in the early and mid-secretory phases compared with the late-secretory phase (p<0.01, Fig. 1B). No significant differences in DROSHA staining were observed in the stromal compartment (comprising of fibroblasts) across the menstrual cycle in both the fertile and infertile endometrium (Fig. 1A and B). In the fertile endometrium, DICER staining intensity was significantly higher in the stromal compartment during the late-secretory phase compared with the proliferative, mid-secretory, and late-secretory phases (p<0.01 and p<0.001, Fig. 1C), in contrast, no significant differences were observed in the infertile endometrium. The luminal and glandular epithelium basal lamina also showed no significant differences in DICER staining across the menstrual cycle in the fertile and infertile endometrium (Fig. 1C and D).
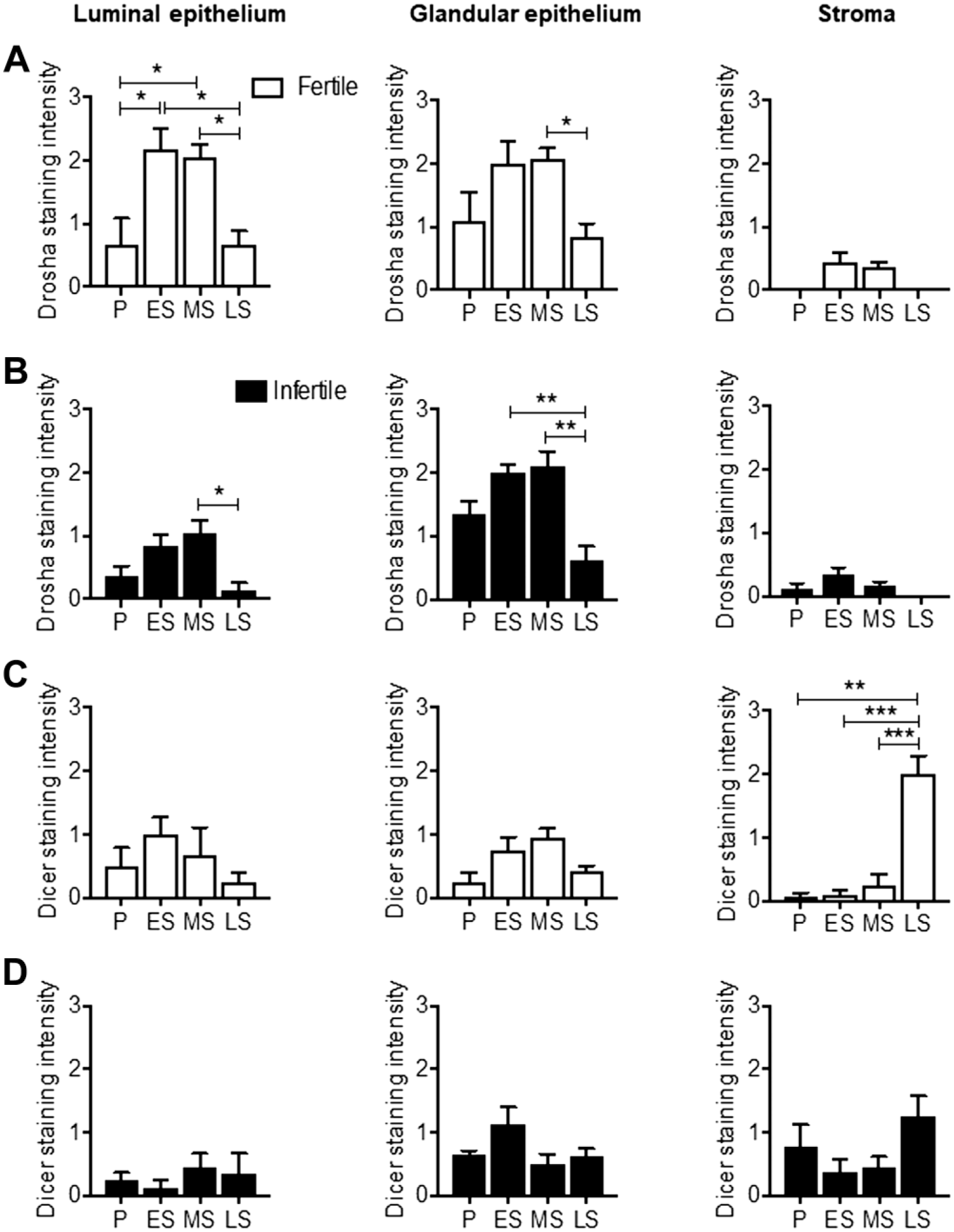
DROSHA and DICER protein expression across the menstrual cycle in the endometrium of fertile and infertile women. Immunoscoring of DROSHA in the (A) fertile and (B) infertile luminal epithelium, glandular epithelium, and stroma during the P, ES, MS, and LS phases. Immunoscoring of Dicer in the (C) fertile and (D) infertile luminal epithelium, glandular epithelium, and stroma during the P, ES, MS, and LS phases. Data are presented as mean ± SEM (n=5/group). Abbreviations: P, proliferative; ES, early secretory; MS, mid-secretory; LS, late-secretory; SEM, standard error of the mean. One-way ANOVA for significance (*p<0.05, **p<0.01, ***p<0.001).
DROSHA mRNA Expression Was Reduced in Infertile Human Endometrium Tissue Compared With Fertile Controls During the Mid-secretory Phase
To determine whether miR machinery genes were altered in infertile endometrium, DROSHA and DICER mRNA expression were compared in the fertile and infertile endometrium during the mid-secretory phase. Relative quantification with qPCR revealed DROSHA mRNA expression levels were significantly reduced in mid-secretory phase infertile whole endometrial tissue compared with mid-secretory fertile endometrium (p<0.05, Fig. 2A). DICER, AGO1, and AGO2 mRNA expressions showed no difference between fertile and infertile endometrium in the mid-secretory phase (Fig. 2B to D). Investigation of DROSHA mRNA expression in isolated HEEC and HESC revealed significantly lower DROSHA expression in the HEEC from the infertile endometrium compared with the fertile control in the mid-secretory phase (Fig. 2A); however, no differences were observed in the stromal compartment. On the contrary, no significant differences were observed for DICER, AGO1, and AGO2 mRNA expression between the fertile and infertile HEEC and HESC isolated from the mid-secretory phase (Fig. 2B to D).
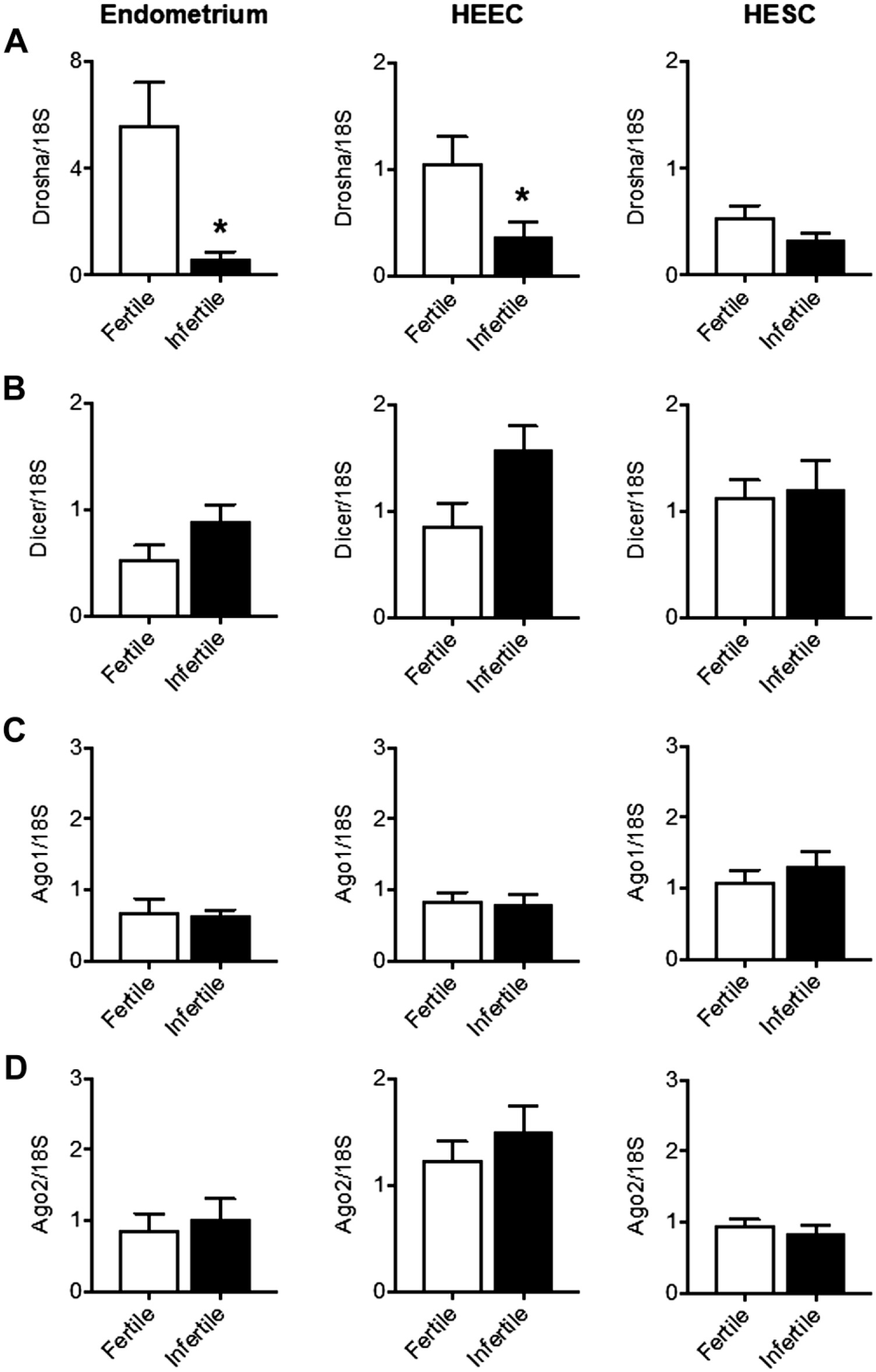
DROSHA, DICER, AGO1, and AGO2 mRNA expression in the endometrium of fertile and infertile women. (A) DROSHA, (B) DICER, (C) AGO1, and (D) AGO2 mRNA expression were measured from whole endometrium tissue, human endometrial epithelial cells (HEEC) and human endometrial stromal cells (HESC). Data are presented as mean ± SEM (n=5/group). Abbreviation: SEM, standard error of the mean. Unpaired t-tests for significance (*p<0.05).
DROSHA and DICER Protein Expressions Are Reduced in the Infertile Endometrium
To determine whether DROSHA and DICER protein expressions are altered in the early and mid-secretory infertile endometrium compared with the fertile controls, DROSHA and DICER immunostaining were assessed. The luminal epithelium basal lamina DROSHA staining was significantly lower in both the early and mid-secretory infertile compared with their respective fertile groups (p<0.01, Fig. 3A) while no significant differences were observed in the glandular epithelium and stromal compartment (Fig. 3A and B). DICER staining was significantly lower in the infertile luminal epithelium basal lamina compared with the fertile group during the early secretory phase whereas no significant differences were observed during the mid-secretory phase (Fig. 4A and B). Furthermore, no differences in DICER staining were observed between the fertile and infertile endometrium in the glandular epithelium and stromal compartments (Fig. 4A and B).
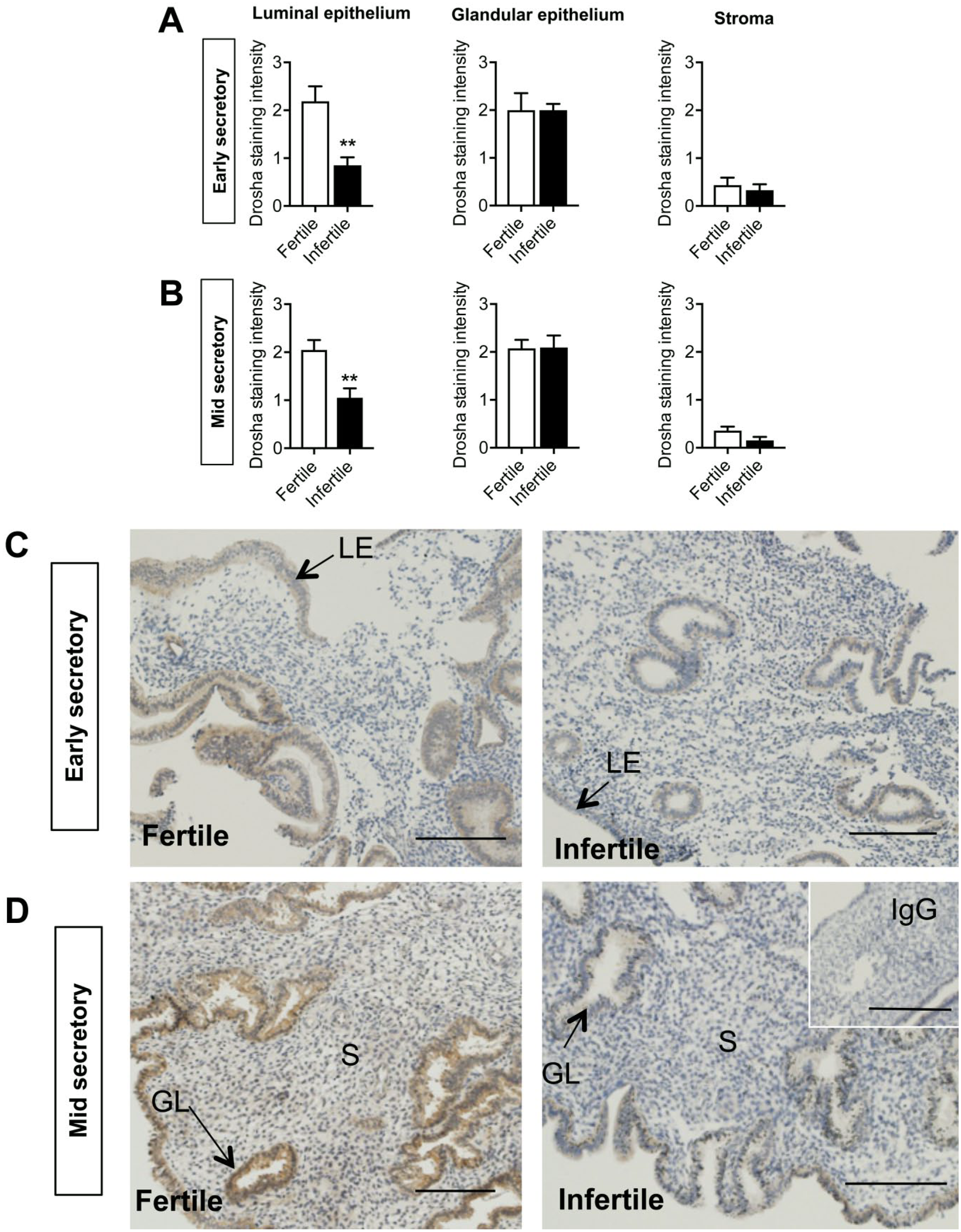
DROSHA immunolocalization in the endometrium during the early and mid-secretory phases in fertile and infertile women. Immunoscoring of DROSHA in the luminal epithelium, glandular epithelium, and stroma during the (A) early and (B) mid-secretory phases in fertile and infertile endometrium. Representative images of DROSHA immunostaining in the (C) early secretory and (D) mid-secretory phases in fertile and infertile endometrium. Negative control in inset. Scale bar represents 100 μm. Data are presented as mean ± SEM (n=5/group). Abbreviation: LE, luminal epithelium; GE, glandular epithelium; SEM, standard error of the mean. Unpaired t-tests for significance (*p<0.05).
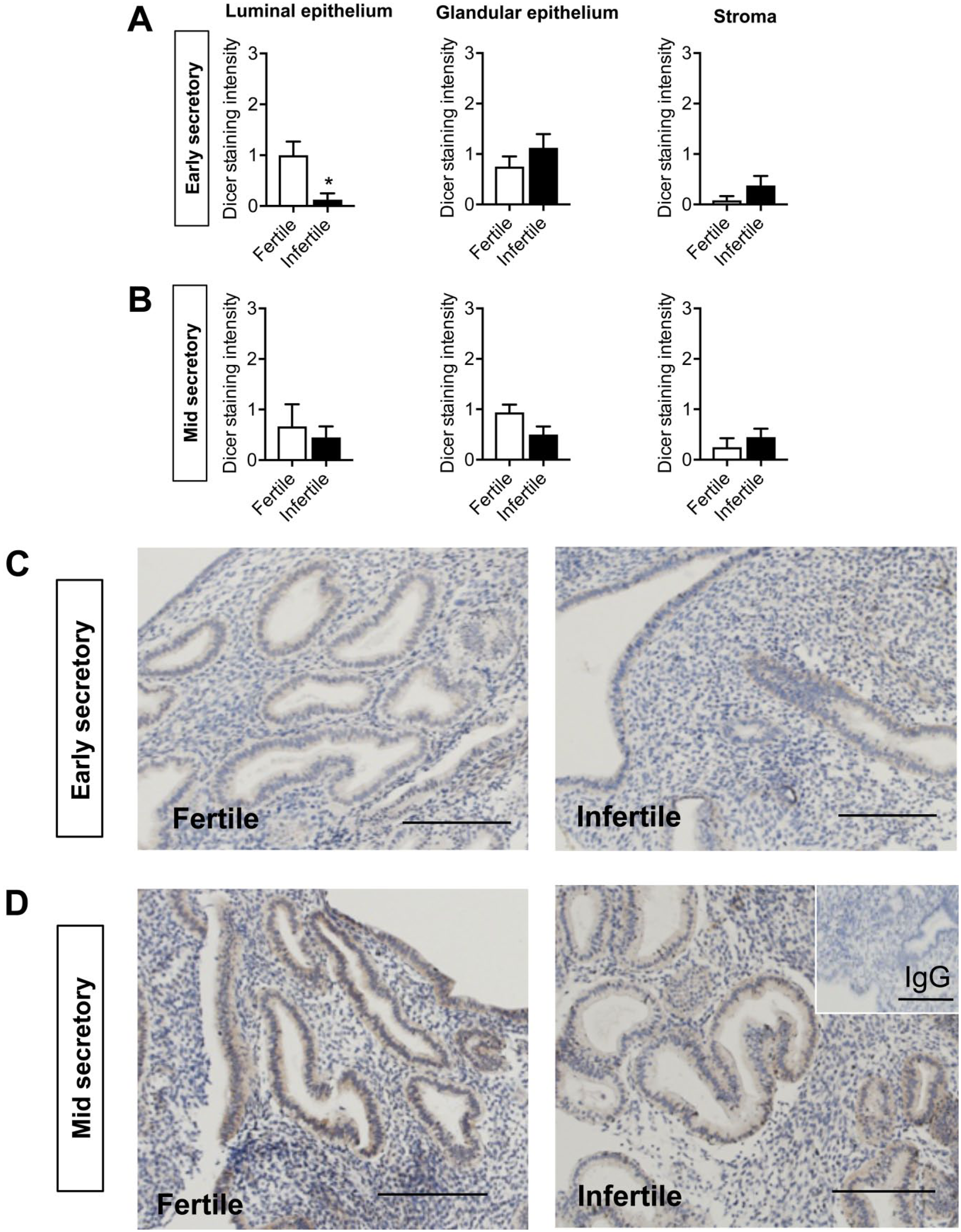
DICER immunolocalization in the endometrium during the early and mid-secretory phases in fertile and infertile women. Immunoscoring of DICER in the luminal epithelium, glandular epithelium, and stroma during the (A) early and (B) mid-secretory phases in fertile and infertile endometrium. Representative images of DICER immunostaining in the (C) early secretory and (D) mid-secretory phases in fertile and infertile endometrium. Negative control in inset. Scale bar represents 100 μm. Data are presented as mean ± SEM (n=5/group). Abbreviation: SEM, standard error of the mean. Unpaired t-tests for significance (*p<0.05).
Discussion
In the endometrium, several miRs are spatiotemporally regulated across the menstrual cycle.26–29 The miR biogenesis machinery components such as DROSHA and DICER play a central role in regulating miR expression; hence aberrant regulation of these components could affect miR expression. Abnormal levels of DROSHA and DICER have been mainly implicated in several types of cancer including ovarian, lung, and breast cancer.30–34 In the present study, we demonstrated for the first time that DROSHA is dynamically regulated across the menstrual cycle in the luminal and glandular epithelium. In parallel, DICER was significantly elevated in the late-secretory stromal compartment compared with other phases suggesting a role in decidualization of endometrial stromal cells. This study also demonstrated that DROSHA mRNA and protein expression is reduced in the early and mid-secretory infertile endometrium compared with fertile endometrium suggesting a role in endometrial receptivity. DROSHA may act to regulate miR expression and therefore protein production, although this remains to be determined. By contrast, DICER protein staining intensity was reduced in the early secretory infertile endometrium compared with fertile endometrium and may also affect miR and protein production in the lead up to receptivity.
Our findings revealed that DROSHA is increased during the early and mid-secretory phases and may therefore affect endometrial receptivity and preparation for implantation. While the upregulation of DROSHA could be associated with the increased miR expression, these findings contradict the downregulation of other miRs during the secretory phases. This could be explained by the alternative mechanisms of miR regulation that include DROSHA and DICER-independent pathways that generate functional miRs. 35 Nonetheless, these findings indicate that DROSHA is important in regulating miRs that are required to facilitate implantation and establishment of pregnancy. DICER was highly expressed while DROSHA staining was absent at the late-secretory stromal compartment. These findings suggest that Dicer could be involved in decidualization, a spontaneous process of HESC differentiating during the mid- to late-secretory phase of the menstrual cycle. In support, DICER mRNA and protein expression in HESC are increased following decidualization treatments in vitro. 36 However, the previous study also suggests that DICER plays a minor role in decidualization, suggesting that other DICER alternative biogenesis pathways are involved in regulating this process. 36
The luminal epithelium is the first point of contact for blastocyst and differentiates to facilitate embryo attachment and subsequently implantation. Considering that DROSHA and DICER expressions were reduced in the infertile compared with the fertile luminal epithelium, both DROSHA and DICER may be important to ensure successful embryo implantation. The increased expression of DROSHA and DICER in endometrium during the early secretory phase suggests that the miR biogenesis machinery components are important in the lead up to implantation. Both components may regulate miR expression and their protein targets which could occur at the beginning of the receptive phase. In addition, the increased expression of DROSHA during the mid-secretory phase, also known as the implantation window, indicates its importance for embryo implantation and subsequently pregnancy. In support, Drosha expression primarily localizes in the mouse uterine epithelium and is gradually elevated during the implantation window, reaching maximal expression on day 6 of the pregnancy. 37 The increase in Drosha expression was not induced by blastocysts in mice, further suggesting that Drosha is essential to prepare the endometrium before embryo implantation. 37 Furthermore, Dgcr8 knockout mice were found to be infertile due to abnormalities in uterine development, 38 indicating that the Drosha-Dgcr8 complex is important for uterine development and female fertility. Hence, low levels of Drosha in the infertile endometrium could be one of the underlying causes for infertility. Further studies investigating whether DGCR8 is also aberrantly regulated in the infertile endometrium would be crucial. While the role of Drosha in fertility of female mice remains elusive, several studies have demonstrated that the loss of Dicer in the reproductive tract of female mice leads to sterility.16–19 Specific miRs (miR-181c, miR-200b, miR-101) were downregulated in the conditional Dicer knockout mice, while pro-apoptotic genes were upregulated, implicating an apoptotic phenomenon. 19 These studies highlight the importance of miR biogenesis machinery components in endometrial development, function, and fertility.
Studies have emerged suggesting that miRs could be involved in regulating the synchronization of the estradiol and progesterone surge during the menstrual cycle. 27 , 39 A genomic profiling study identified 12 specific miRs predicted to suppress the expression of cell cycle regulators were upregulated in the progesterone-driven secretory phase. 27 In support, mice studies revealed that numerous uterine miRs are highly expressed during pregnancy whereby progesterone levels are increased. 40 , 41 Furthermore, uterine Drosha expression in ovariectomized mice was elevated following progesterone treatment whereas estrogen treatment had no effect. 37 In support, our present study revealed that DROSHA protein expression in the luminal epithelium of the fertile endometrium is highly expressed during the progesterone-driven early and mid-secretory phases compared with the estrogen dominant proliferative phase. The lower DROSHA protein expression in the luminal epithelium of the infertile endometrium at the early and mid-secretory phases compared with their respective fertile groups could be due to lower progesterone levels in the infertile group, at least in some women although this remains to be determined. On another note, a previous study demonstrated that Dicer ablation reduced progesterone receptor expression in the uterine luminal epithelium, suggesting that Dicer deletion could have directly or indirectly caused the depletion of progesterone receptors, 19 and affect subsequent progesterone signaling downstream pathways. Along these lines, the present study showed that the lower DICER protein expression in the infertile early secretory phase luminal epithelium compared with the fertile group may be one of the underlying factors for infertility in these patients. Together, the current and previous studies have provided evidence that a reciprocal interaction may exist between progesterone and the miR biosynthesis network to facilitate embryo implantation and subsequently pregnancy.
Overall, our findings have demonstrated that the major miR regulators DROSHA and DICER are spatiotemporally regulated across the phases in the menstrual cycle. Our study also presented the possibility that reduced expression of these regulators during the secretory phases could be associated with infertility. However, functional studies are required to provide a better insight into the role of miR machinery factors in receptivity and implantation in the endometrium.
Supplemental Material
DS_10.1369_0022155419854064 – Supplemental material for MicroRNA Biogenesis Machinery Is Dysregulated in the Endometrium of Infertile Women Suggesting a Role in Receptivity and Infertility
Supplemental material, DS_10.1369_0022155419854064 for MicroRNA Biogenesis Machinery Is Dysregulated in the Endometrium of Infertile Women Suggesting a Role in Receptivity and Infertility by Hannah Loke, Kate Rainczuk and Evdokia Dimitriadis in Journal of Histochemistry & Cytochemistry
Footnotes
Acknowledgements
The authors thank SRN Judi Hocking for collection of human endometrium.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
HL and KR performed the experiments and analyzed the data. ED designed the study. HL and ED wrote and revised the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by funding for a project grant and senior research fellowship from the National Health and Medical Research Council (NHMRC) of Australia to ED. This work was supported by the Victorian Government’s Operational Infrastructure Support Program.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.