
Abstract
The Escherichia coli LacZ gene (encoding β-galactosidase) is a widely used reporter for gene regulation analysis in transgenic mice. Determination of β-galactosidase activity is classically performed using 5-bromo-4-chloro-3-indolyl-β-d-galactopyranoside/ferri-/ferrocyanide (X-Gal/FeCN) histochemistry. Uncertainty about the origin of the β-galactosidase signal is encountered in tissues containing high levels of endogenous β-galactosidase. Here, we show that reliable results can nevertheless be obtained in these tissues by performing the histochemical reaction under slightly basic pH conditions (pH 8–9). We further demonstrate that in this context, analysis of tissue sections may be advantageous over that of conventional whole-mount tissues because poor dye penetration and remaining tissue acidity are avoided in tissue sections. We also recommend that bacterial debris should always be carefully removed from the luminal surface of gastrointestinal tract specimens unless staining of resident microflora is deliberately used as an internal positive control in the assay. Finally, we show that 6-chloro-3-indolyl-β-
Introduction
Genetically engineered mice expressing the LacZ reporter gene encoding bacterial β-galactosidase (Bact β-Gal) under a variety of promoters have been widely used to determine gene expression patterns.
1
In such mice, visualizing Bact β-Gal activity is achieved by hydrolysis of the indoxyl glycoside 5-bromo-4-chloro-3-indolyl-β-d-galactopyranoside (X-Gal) to an indolyl alcohol, which then forms an insoluble blue precipitate. The reaction is easy and quick to perform, inexpensive, and applicable to small embryos, intact organs, and sectioned tissue.
2
The main disadvantage of using Bact β-Gal histochemistry (HC) in a screen for targeted gene expression is the specificity of the method, which differs dramatically between organs because of highly variable amounts of interfering mammalian endogenous β-galactosidase (Endo β-Gal). In mice, Endo β-Gal is present in a strain-dependent manner mainly in the epididymis, kidney, and gastrointestinal tract, resulting in unwanted X-Gal staining.3–5 Endo β-Gal may mask, mimic, or interfere with detection of LacZ reporter gene expression. A few previous studies have explored this potential pitfall in the phenotyping process of genetically modified mice mainly using whole mounts
6
or minced organs.
7
These studies emphasize that modulation of reaction parameters (eg, pH) may function as a tool to discriminate between Endo β-Gal activity and activity resulting from LacZ reporter gene expression.
8
Unfortunately, most of the protocols in the literature for X-Gal staining omit the need to conduct pH checks and routine pH adjustments to avoid X-Gal staining from the endogenous enzyme. In our laboratory, we routinely employ cryostat sections and pH-adjusted X-Gal staining to specifically identify Bact β-Gal-producing cells in cryostat sections of LacZ transgenic mouse organs. Our simple and robust staining approach is even effective in the study of tissues with high Endo β-Gal levels, such as the epididymis, kidney, and gastrointestinal organs. However, within the last years, an alternative, sensitive method able to detect Bact β-Gal activity has been developed in whole-mount embryos.
9
It comprises the use of 6-chloro-3-indolyl-β-
Materials and Methods
We made use of organs obtained in another study. 10 No animals were sacrificed solely for the purpose of this study. Animal maintenance and carbon dioxide euthanasia in the other study were in accordance with guidelines set by the EU Laboratory Animal Directive, German Animal Welfare Act and approved by the government agency of the Free State of Saxony, Germany (T27/11; T46/12; T07/13).
Organs used in whole-mount HC were lightly fixed in 4% formalin before histochemical processing. Organs used in cryostat section HC were snap frozen in Tissue-Tek-filled molds (VWR International, Dresden, Germany). The frozen organs were cut into sections with a thickness of 10 µm in a cryostat at −20C, mounted on glass slides, postfixed in ice cold methanol/acetone (1:1) for 1 min, and air-dried for 15 min at room temperature.
The histochemical X-Gal reaction was performed in different buffer systems depending upon the designated pH: (1) 0.1-M citric acid–0.1-M sodium citrate buffer was employed for a designated acidic pH, (2) 0.5-M HEPES buffer for a pH between 7 and 8, and (3) 0.2-M glycine–0.2-M sodium hydroxide buffer for a basic pH above 8. Fifty mg (2.5 mM) of X-Gal dissolved in 2.5-ml N,N-dimethylformamide was added followed by the addition of 15-mM NaCl, 1.3-mM MgCl2, 3-mM Fe(CN)6, and 3-mM K4(Fe(CN)6) to 50-ml final volume of the various buffer solutions. The buffer system for the S-Gal reaction consisted of 0.1% sodium deoxycholate; 0.2% octylphenoxy poly(ethyleneoxy)ethanol, branched (linear formula: (C2H4O)nC14H22O; IGEPAL CA-630; Sigma, Taufkirchen, Germany); and 2-mM MgCl2 dissolved in 0.1-M phosphate buffer, pH 7.3, if not otherwise stated. Fifty mg (3 mM) of S-Gal (Sigma) and 0.4-mM nitro blue tetrazolium chloride (NBT; Sigma) were added to 50 ml of this buffer. X-Gal staining of whole mounts was performed in tubes on a shaker overnight at room temperature. X-Gal staining of tissue sections was routinely conducted in a humidified chamber overnight at 30C. Staining of cryostat sections with S-Gal was initially performed in humidified atmosphere overnight at 30C and later for 6 hr at either 30C or 37C (see the “Results” section for further details). All specimens were thoroughly washed after substrate incubation with distilled water and in the case of cryostat sections, counterstained with nuclear fast red-aluminum salt in the case X-Gal/FeCN staining or with Mayer’s hematoxylin in the case of S-Gal/NBT incubation. Cryostat sections were embedded in Entellan (Merck Chemicals, Darmstadt, Germany) and documented with an Axioplan 2 light microscope (Zeiss, Jena, Germany) equipped with a ProgRes C3 digital camera (Jenoptik, Jena, Germany) and a digital recording system (ProgRes CapturePro 2.8.8; Jenoptik). Whole mounts were floated in PBS and analyzed with an Olympus SZ61 stereo microscope equipped with an SC30 camera and cellSens image acquisition software (all Olympus, Hamburg, Germany).
Results
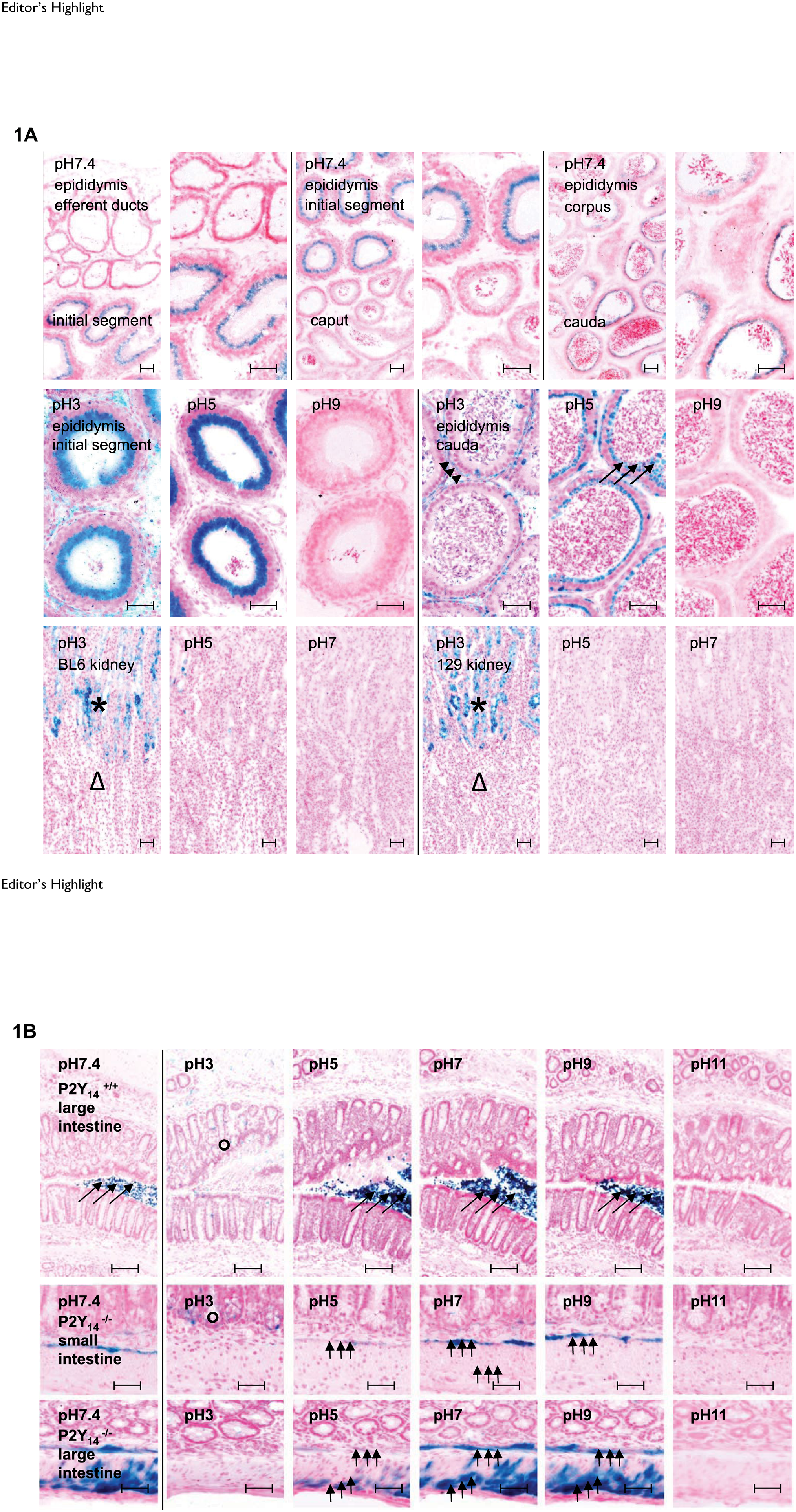
In cryostat sections of murine epididymis, the X-Gal HC of Endo β-Gal varied strongly according to the anatomic site (Fig. 1, top row) and dependent on the pH of the reaction mixture (Fig. 1, second row from the top). Endo β-Gal staining was intense in the epithelium of the initial segment of the caput and the cauda epididymis. Virtually no staining was seen in the epithelium of the corpus epididymis and in the epithelium of the efferent ducts. Lowering the pH of the reaction mixture to 3 caused a substantial shift in the staining pattern in the cauda from intraductal (arrows) to periductal cells (arrow heads). In contrast, elevating the pH of the reaction mixture above 8 prevented Endo β-Gal staining by X-Gal in epididymis sections completely. Unfortunately, Endo β-Gal preserved its potential to interfere with Bact β-Gal activity in epididymis whole mounts at a pH range between 3 and 11 probably as a consequence of a pH gradient within the tissue (Fig. 2, top row).
Editor’s Highlight
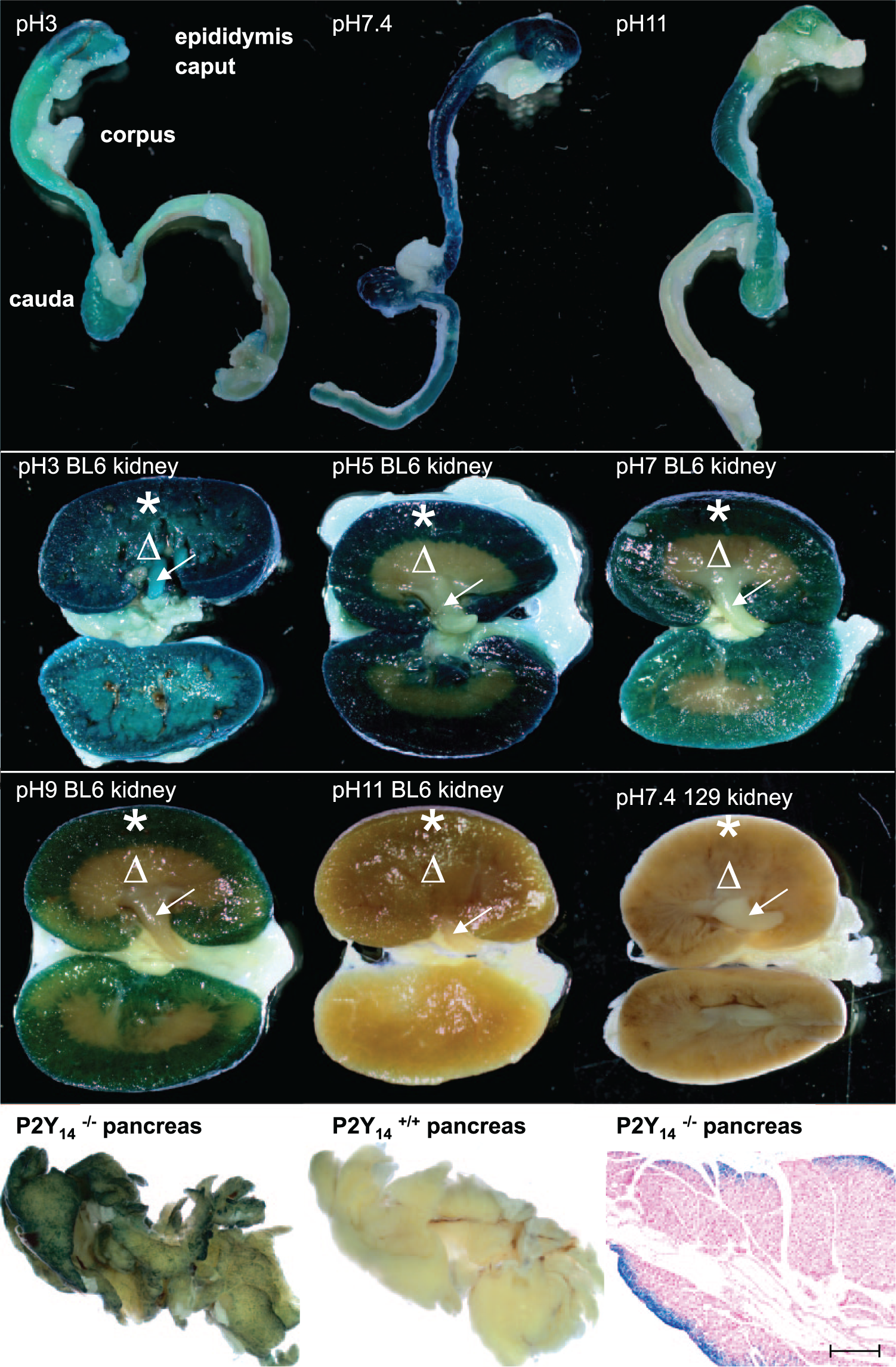
Macroscopic detection of endogenous β-galactosidase by 5-bromo-4-chloro-3-indolyl-β-d-galactopyranoside (X-Gal) whole-mount histochemistry in wild-type epididymis (top row), wild-type kidney (second and third rows from the top), and in pancreata from wild-type mice, as well as bacterial β-galactosidase in pancreata from mice with a targeted mutation of P2Y14 (fourth row from the top, first two panels from the left). Section prepared from an X-Gal-stained pancreas from a mouse with expression of the LacZ reporter gene under the P2Y14 promoter (fourth row from the top, last panel from the left). Counterstaining with nuclear fast red-aluminum sulfate solution; scale bars as indicated (see the text for further details). The scale bar represents 200 µm. From top to bottom: 2nd and 3rd rows the stars indicate renal cortex; 2nd and 3rd rows the triangles indicate renal medulla; 2nd and 3rd rows the arrows indicate renal pyramids.
In the murine kidney, Endo β-Gal activity was primarily a problem for Bact β-Gal detection in the cortex (star), but enzyme activity varied across mouse strains, tissue preparations, and individual kidneys (Fig. 1, third row; Fig. 2, second and third rows from the top). Endo β-Gal activity was negligible in the kidney medulla (triangle) and papilla (arrow in Fig. 2, second and third rows from the top) of both cryostat sections and whole mounts. In the gastrointestinal tract, Endo β-Gal activity was detectable in the crypts of Lieberkühn under acidic reaction conditions (circles in Fig. 1, fourth and fifth rows from the top). Transient and resident bacterial microflora may have been responsible for an unwanted specific staining in the lumen (large arrows). The β-Gal of the bacterial microflora catalyzed hydrolysis of X-Gal in exactly the same way and with the same pH activity profile as the Bact β-Gal originating from LacZ reporter gene expression, as demonstrated here for the smooth muscle layers (small arrows) in intestinal sections from mice with a targeted mutation of P2Y14 (Fig. 1, fifth and sixth rows from the top). 10 Therefore, bacterial debris must be carefully removed from the luminal surface of gastrointestinal tract specimens before X-Gal staining unless microfloral staining is used as an internal positive control.
In whole-mount embryos, an alternative protocol for the visualization of LacZ reporter gene expression has recently been reported. 9 We conducted this S-Gal/NBT assay in parallel with the standard X-Gal/FeCN assay on cryostat sections. The S-Gal/NBT assay worked equally well on the sectioned adult tissue as it has been shown to work in whole-mount embryonic tissue by others. 9 The obtained staining patterns were absolutely identical when compared with the ones obtained by X-Gal/FeCN HC. However, the overnight histochemical staining response was much more intense when compared with the signal obtained with X-Gal/FeCN. To convert the intensity of the S-Gal/NBT staining to the reference standard staining with X-Gal/FeCN, the incubation time for the S-Gal/NBT reaction had to be shortened from overnight to 6 hr (Fig. 3, top row). This shortening of the incubation time resulted in a marked decrease in Endo β-Gal staining. Even in cryostat sections with high interfering Endo β-Gal activity (eg, epididymis, kidney), almost no Endo β-Gal staining was observed when cryostat sections were treated with S-Gal/NBT instead of X-Gal/FeCN (Fig. 3, middle row). A dramatic reduction in the overall assay activity was noted when an acidic pH was employed (Fig. 3, bottom row).
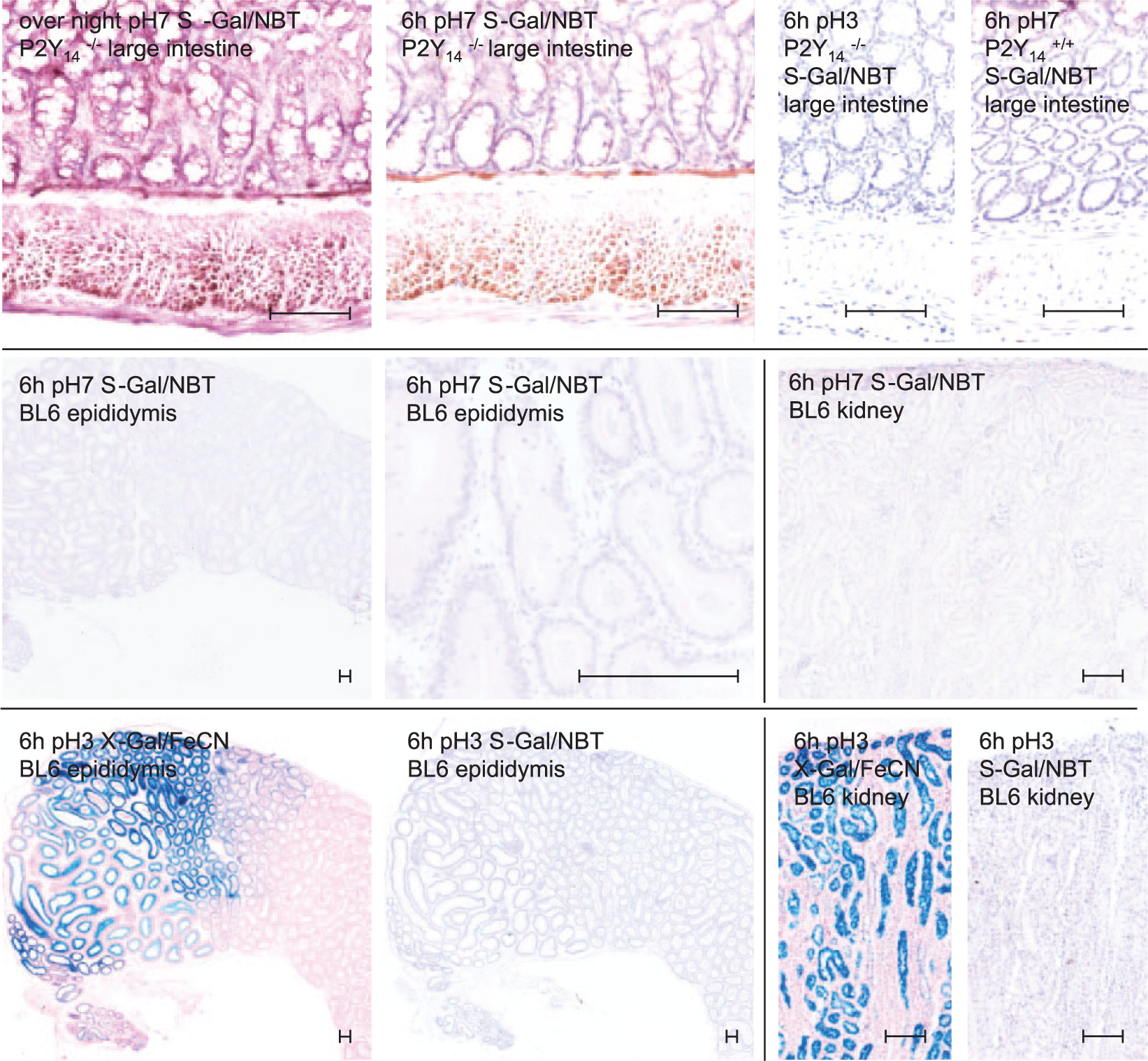
Detection of bacterial β-galactosidase (Bact β-Gal) and endogenous β-galactosidase (Endo β-Gal) by S-Gal histochemistry (HC) in comparison with X-Gal HC. Upper row: Detection of Bact β-Gal by S-Gal HC in cryostat sections of large intestines from P2Y14-deficient mice showing LacZ reporter gene expression at a physiological and an acidic pH and in comparison with a wild-type littermate as indicated. Middle row: Endo β-Gal activity tested in adult murine tissues such as epididymis (first and second sections from the left) and kidney (third section from the left). Bottom row: Comparison of X-Gal/FeCN and S-Gal/NBT staining at an acidic pH in high Endo β-Gal containing organ sections. Counterstaining with nuclear Mayer’s hematoxylin and fast red-aluminum sulfate solution; scale bars as indicated (see the text for further details). The scale bars represent 100 µm. Abbreviations: S-Gal/NBT, 6-chloro-3-indolyl-β-
Discussion
In this work, we used X-Gal staining of cryostat sections and whole mounts to demonstrate how Endo β-Gal expression in C57BL/6 mice can effect the determination of LacZ reporter gene expression in the epididymis, kidney, and gastrointestinal organs.
In view of the reported results and our extensive experience with methods to detect LacZ reporter gene expression by X-Gal staining in transgenic mice, we recommend the following procedure:
perform the enzyme HC immediately after preparing the cryostat sections;
keep the pH of the X-Gal reaction mixture between 8 and 9;
exclude or confirm interfering Endo β-Gal activity, selectively by lowering and elevating the pH of the reaction mixture, particularly when targeted gene expression is explored in a tissue that is commonly known to contain high Endo β-Gal activity; and
thoroughly rinse naso-oropharyngeal and gastrointestinal mucosal specimens before shock freezing to avoid staining by luminal microflora unless microfloral staining is intended as a natural positive control.
Screening whole-mount preparations for the expression of LacZ-reporter-bearing transgenes allows investigators to quickly obtain three-dimensional information concerning the expression of a targeted gene without the need for embedding and sectioning the tissues. The limitation of using whole-mount tissue in a screen at necropsy is that Endo β-Gal staining may override the Bact β-Gal staining (eg, Fig. 2, top row). It is important to note that X-Gal poorly penetrates into organs (even loosely organized organs, such as the murine pancreas). As a consequence, whole-mount staining may be incomplete, uneven, and has the potential for generating false-negative results (Fig. 2, fourth row from the top).
In summary, we show that cryostat sections preserve β-Gal activity to a high degree and allow for the determination of the contribution of individual cells to the total reaction. Our data indicate that the LacZ reporter gene should be used with caution in mice in which the transgene of interest must be assessed in a battery of organs known to contain high amounts of Endo β-Gal (eg, epididymis, testis, kidney, secretory glands, and intestine). Immediate enzyme HC of tissue cryostat sections is the superior choice compared with whole-mount HC in this case. If species, strain, and sex-specific divergence in Endo β-Gal activity are considered and special attention is paid to raise the reaction mixture pH to a weakly alkaline range, X-Gal staining of cryostat sections allows reliable discrimination between Endo β-Gal activity and Bact β-Gal activity (Fig. 1).
Reliable detection of Bact β-Gal in cryostat sections can also be achieved by S-Gal/NBT staining. S-Gal/NBT staining appears to be more sensitive than X-Gal/FeCN staining in sectioned tissue (this study) as well as in whole-mount tissue. 9 The higher sensitivity of S-Gal/NBT staining allows a shortening of the incubation time (Fig. 3, top row). This has the beneficial effect that interfering Endo β-Gal staining is minimized (Fig. 3, middle row). The shortening of the incubation time will certainly positively influence tissue integrity. This will be particularly advantageous if antigen detection by immunohistochemistry is intended to follow S-Gal/NBT staining.
Footnotes
Acknowledgements
We cordially thank Professor Jaime Rivera, Department of Cell and Developmental Biology, University of Massachusetts. His advice was of great help in establishing the 6-chloro-3-indolyl-β-
Author Contributions
AMR designed the study, assembled the data, and drafted the manuscript. OB critically interpreted the data, revised the draft, and wrote the final version of the manuscript with AMR. CM and AS were heavily involved in data acquisition, analysis, interpretation, and presentation. All authors have read and approved the final manuscript version.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This particular study received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors. It was jointly financially supported by the Institute of Anatomy and the Institute of Biochemistry, Medical Faculty, University of Leipzig.