
Abstract
Secretogranin III (SgIII) is a granin protein involved in secretory granule formation in peptide-hormone-producing endocrine cells. In this study, we analyzed the expression of the LacZ reporter in the SgIII knockout mice produced by gene trapping (SgIII-gtKO) for the purpose of comprehensively clarifying the expression patterns of SgIII at the cell and tissue levels. In the endocrine tissues of SgIII-gtKO mice, LacZ expression was observed in the pituitary gland, adrenal medulla, and pancreatic islets, where SgIII expression has been canonically revealed. LacZ expression was extensively observed in brain regions, especially in the cerebral cortex, hippocampus, hypothalamic nuclei, cerebellum, and spinal cord. In peripheral nervous tissues, LacZ expression was observed in the retina, optic nerve, and trigeminal ganglion. LacZ expression was particularly prominent in astrocytes, in addition to neurons and ependymal cells. In the cerebellum, at least four cell types expressed SgIII under basal conditions. The expression of SgIII in the glioma cell lines C6 and RGC-6 was enhanced by excitatory glutamate treatment. It also became clear that the expression level of SgIII varied among neuron and astrocyte subtypes. These results suggest that SgIII is involved in glial cell function, in addition to neuroendocrine functions, in the nervous system:
Keywords
Introduction
Secretogranin III (SgIII) is a member of the granin protein family that is involved in secretory granule formation in peptide-hormone-producing endocrine cells. The granin family consists of seven types of acidic secretory proteins: chromogranin A (CgA), chromogranin B (CgB), secretogranin II (SgII), and SgIII–VI.1–4 Some of the members of the granin family, such as CgA, CgB, SgII, and SgIII, may participate in the aggregation/condensation process of peptide hormones during the biosynthesis of secretory granules (SGs). 5 In this process, SgIII has been suggested to play a role as a bridge molecule between the peptide hormone/CgA core aggregates and the cholesterol-rich microdomains of SG membranes.6,7 Furthermore, SgIII has been found to bind to membrane-associated carboxypeptidase E (CPE), a prohormone-processing (end-modifying) enzyme that has been proposed as a candidate molecule for the sorting receptor of prohormones. 8 These findings suggest that the cooperative formation of a sorting platform consisting of SgIII, CgA, and CPE at the trans-Golgi network is crucial for SG formation.6,8,9 SgIII was originally identified as a neuron-specific gene 1B1075 product in mice.10–12 Immunohistochemical and in situ hybridization analyses have shown the expression of SgIII in the nervous and endocrine tissues of various organisms.6,11,13–17 However, it does not have completely understood the expression of SgIII of more detailed cell or tissue structure.
We have established a strain of SgIII gene-deficient mice via the gene trap method (SgIII-gtKO) and conducted analyses at the individual level. We found that SgIII-gtKO mice exhibited no overt abnormalities under ordinary rearing conditions. However, these animals were highly vulnerable to various loading conditions compared with normal wild-type control (WT) mice. For example, they showed marked obesity and impaired glucose tolerance when fed a high-fat/high-sucrose diet. These phenotypes were associated with increased proinsulin accumulation, accompanied by hypertrophy of pancreatic islets and decreased active insulin secretion. Furthermore, the SgIII-gtKO mice exhibited a small elevation of plasma adrenocorticotropic hormone (ACTH) content from the pituitary gland after chronic restraint stress. Thus, the lack of SgIII caused maladaptation of endocrine cells to inadequate diet and stress by impairing the proteolytic conversion of prohormones in SGs. 18
The SgIII-gtKO strain was produced using gene trapping, in which the gene trap vector is inserted into intron 11 of the SgIII genome on chromosome 9, followed by the induction of β-Geo (a fusion gene of the LacZ reporter and neomycin-resistance genes) expression by the authentic SgIII promoter. Therefore, this model has the advantage of being able to visualize the SgIII expression sites with high sensitivity, which renders it an effective tool for detailed cell/tissue expression analyses. In the central nervous system (CNS), the SgIII transcripts and protein have been found widely in brain regions and seem to be predominantly expressed in neuronal elements associated with neuroendocrine function and monoaminergic neurotransmission.11,17,19 More interestingly, it has been reported that the expression of SgIII in astrocytes is involved in the astrocytic secretory pathway and is regulated by the glial activation state. 16 Here, we studied the expression of SgIII extensively in the nervous tissues using the LacZ reporter and identified the subtypes of SgIII-expressing neurons and astrocytes. We demonstrated that LacZ expression was observed extensively in the central and peripheral nervous tissues and was pronounced in glial cells. In addition, using rat glial cell lines, we clarified the relationship between SgIII expression and the activated glial response induced by glutamate stimulation.
Materials and Methods
Gene Trapping and SgIII-gtKO Mice
The gene trapping used in this study has been published elsewhere.20,21 The structure of the gene-trapping cassette, its genomic insertion site, and the expression of a fusion gene of the LacZ reporter and neomycin-resistance genes (β-Geo) in SgIII-gtKO mice (B6;129S-Scg3tm1Lex strain; The Jackson Laboratory, Bar Harbor, ME) are shown in Fig. 1. The SgIII-gtKO and WT mice were generated in the genetic background of the C57BL/6J strain by the intercrossing of heterozygous mice. The genotypes of the offspring were determined by amplifying the SgIII alleles and the gene-trapping cassette by polymerase chain reaction using the appropriate primers, as previously reported. 18 The mice were maintained on a 12-hr light/dark cycle and given free access to tap water and standard chow (CRF-1; Oriental Yeast, Osaka, Japan). All animal experiments were performed in compliance with relevant laws and the guidelines of the Care and Use of Laboratory Animals of the Research Council of Akita Prefectural University (approval number 17-03 and 18-02).
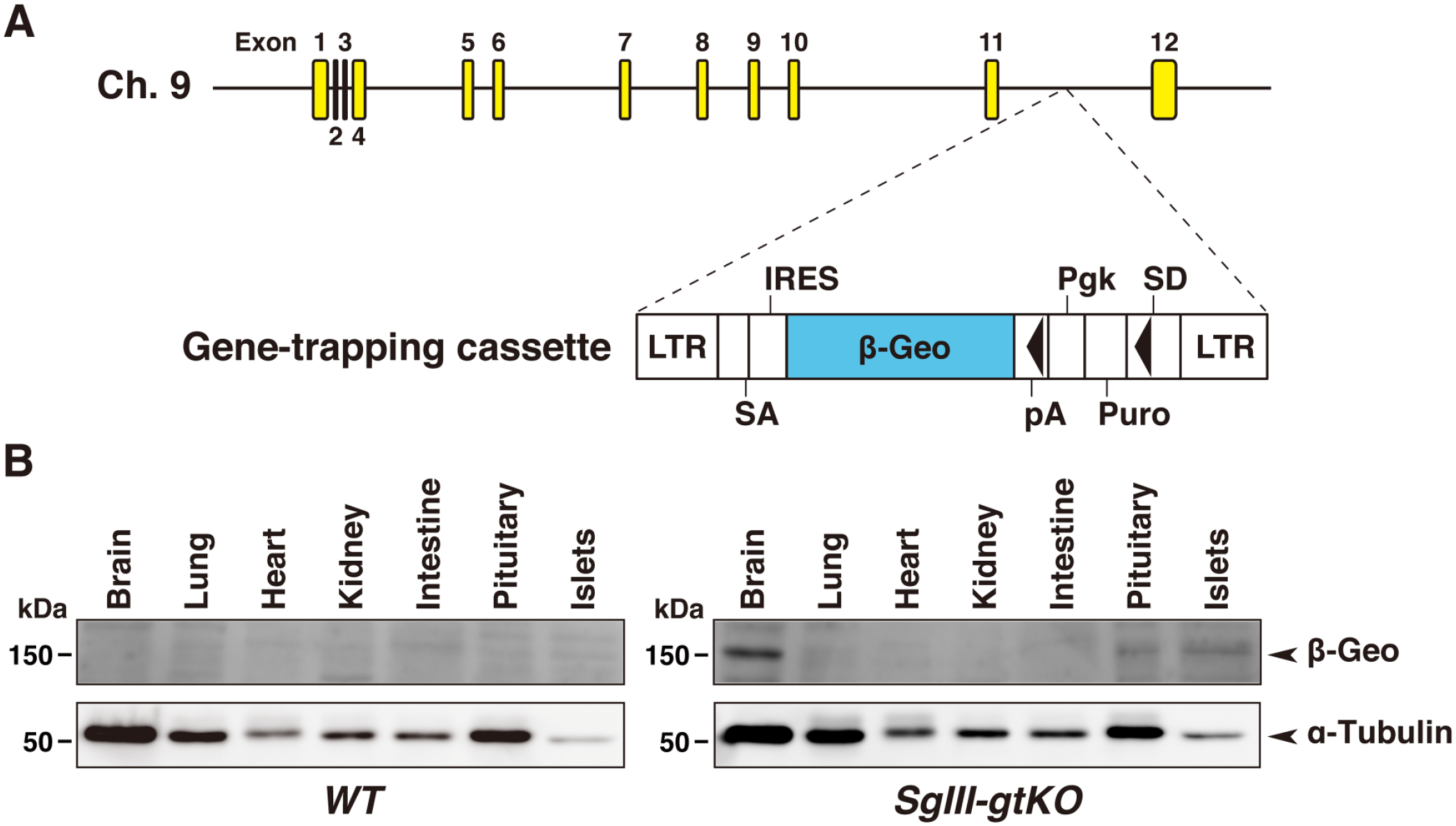
Structure of the SgIII gene trap vector and genomic insertion. (A) The gene trap vector was inserted on mouse chromosome 9, between exons 11 and 12 of the SgIII genome. 18 The gene-trapping cassette contains a splice-acceptor site (SA) positioned at the 5′ end of a promoterless reporter region, β-geo (a fusion of neomycin phosphotransferase and β-galactosidase), allowing the vector to be spliced to the endogenous gene with an internal ribosomal re-entry site (IRES) between the SA site and the reporter gene sequence. The puromycin resistance gene (Puro) is also expressed when the gene trap vector is inserted into an intron under the control of a constitutive phosphoglycerate kinase gene (Pgk) promoter, which is thought to trap a gene to be transcriptionally silent in mouse embryonic stem cells. As a result, the LacZ-reporter gene can be translated even when it is not fused to the trapped gene. 21 The black triangles in the cassette indicate the loxP sites. Abbreviations: LTR, retroviral long terminal repeat; SD, splice donor site; SgIII, secretogranin III. (B) Expression of β-Geo detected by immunoblotting using an anti-β-galactosidase antibody (upper panels). Equal amounts of protein were extracted from each tissue (20 μg) of WT or SgIII-gtKO mice. Loading controls were provided by α-tubulin immunoblotting (lower panels).
LacZ Staining
For LacZ staining, tissue processing was performed as described previously.22,23 Five-month-old male mice (n=5) were deeply anesthetized by an intraperitoneal injection of sodium pentobarbital (0.1 mg/g body weight) and fixed with 50 ml of ice-cold 3.8% formaldehyde in 0.1 M sodium phosphate buffer (pH 7.4) via cardiac perfusion for 10 min after washing out the blood with physiological saline perfusion for 3 min. The brain, spinal cord, and peripheral nervous and endocrine tissues were removed and postfixed in 1.9% formaldehyde in 0.1 M sodium phosphate buffer (pH 7.4) at 4C overnight. The isolated islets (as described below) were also fixed by immersion in 3.8% formaldehyde in 0.1 M sodium phosphate buffer (pH 7.4) at 4C overnight. Fixed brains were embedded in 2% agarose in 0.05 M sodium phosphate buffer (pH 7.4) and sliced coronally on a Microslicer DTK-1000 instrument (Dosaka EM; Kyoto, Japan) at a thickness of 400 μm. Slices were rinsed in 0.05 M sodium phosphate buffer (pH 8.0) 3× and reacted in 5-bromo-4-chloro-3-indolyl-β-
Isolation of Pancreatic Islets
Pancreatic islets were isolated by collagenase digestion methods from 5-month-old male mice (n=4) as described previously. 25 Briefly, 6 ml of collagenase solution (500 U/ml) (C-7657, type XI; Sigma-Aldrich, Inc. St. Louis, MO) was injected from the duodenal papilla of the pancreatic duct, and the pancreas was excised from the abdomen and digested at 37C for 30 min. After digestion, the tissue was dispersed by gentle pipetting and islets were isolated from exocrine tissues. The islets were handpicked under a stereomicroscope and used for LacZ staining and immunoblotting.
Primary Antibodies
The rabbit polyclonal anti-SgIII antibody raised against rat SgIII peptides 373–471 7 was used for immunohistochemistry and immunoblotting (1:2000 dilution). The mouse monoclonal anti-β-galactosidase antibody (1:2000 dilution, Z3781; Promega, Madison, WI) was used for immunoblotting to analyze the tissue expression of LacZ. The mouse monoclonal anti–glial fibrillary acidic protein (GFAP) antibodies (1:200 dilution, GF5, ab10062; Abcam, Cambridge, UK and 1:60 dilution, GF12.24; Progen Biotechnik GmbH, Heidelberg, Germany) and rabbit polyclonal anti–glutamine synthetase (GS) antibody (1:20,000 dilution, ab49873; Abcam) were used for immunohistochemistry and immunoblotting. The mouse monoclonal anti-α-tubulin antibody (1:3000 dilution, T5168; Sigma-Aldrich, Inc.), anti–glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibody (1:500 dilution, ab9484; Abcam), and anti-β-actin antibody (1:5000 dilution, A5316; Sigma-Aldrich, Inc.) were used for loading control in the immunoblot analysis.
Immunohistochemistry
Tissues of 4- to 6-month-old male mice (n=5) were excised and fixed in Bouin’s fixative for 3 days. Tissue blocks were then dehydrated and embedded in paraffin. Five-µm-thick sections were mounted on silane-coated glass slides (Muto Pure Chemicals; Tokyo, Japan) and processed for immunohistochemistry as described previously. 17 For SgIII immunohistochemistry, deparaffinized sections were immersed in 0.01 M sodium citrate buffer (pH 6.0) and processed using microwave antigen retrieval. Subsequently, the sections were treated with Dako REAL Peroxidase-Blocking Solution (Dako; Glostrup, Denmark) and blocked with 5% normal goat serum (Vector Laboratories; Burlingame, CA) in PBS. After incubation with primary antibodies overnight at 4C, the sections were rinsed in PBS and incubated with secondary antibodies using EnVision Dual Link System horseradish peroxidase (HRP)-labeled polymer reagent (Dako). The sections were then visualized using a peroxidase substrate kit (SK-4100; Vector Laboratories) and counterstained with Mayer’s hematoxylin. The control experiment in the immunohistochemistry followed the literature which is describing its guidelines. 26 As a negative control, sections were incubated with normal rabbit or mouse IgG (Santa Cruz; sc-2027 or sc-2025, 1 μg/mL) instead of the primary antibodies; we confirmed that there were no significant immunosignals in this case. Images were acquired using a BX51 microscope equipped with a DP72 CCD camera and digitized with the DP2-BSW software (Olympus).
Dual Staining of LacZ Expression and Immunohistochemistry
LacZ-stained tissues were embedded in 0.4% Agarose L (317-01182; Nippon Gene Co. Ltd., Tokyo, Japan) and 1% gelatin (16605-42; Nacalai Tesque, Kyoto, Japan) in 0.05 M sodium phosphate buffer (pH 7.4) and refixed in Bouin’s fixative for 24 hr. The tissue blocks were dehydrated and embedded in paraffin. Subsequently, deparaffinized sections were processed by antigen retrieval, reacted with primary antibodies, and incubated with secondary antibodies using the HRP-labeled polymer technique, as mentioned above.
Immunoblotting
For immunoblot analysis, tissue samples from 5-month-old male mice (n=6) were collected, frozen on dry ice, and stored at −80C until use. Frozen tissues were thawed and homogenized with lysis buffer containing 20 mM Tris, pH 7.5, 150 mM NaCl, 2.5 mM MgCl2, 1 mM EGTA, 1.0% Triton X-100, and a protease inhibitor cocktail (Complete Mini; Roche Diagnostics, Mannheim, Germany). The homogenates were spun at 20,000 × g for 10 min, the supernatants were collected, and the protein concentration was determined using a Coomassie brilliant blue protein assay reagent (Nacalai Tesque Inc.), as described previously. 17 Each protein extract was normalized for total protein content (5−40 µg), subjected to SDS-PAGE, and transferred to an Immobilon-P transfer membrane (Millipore; Billerica, MA). After blocking with 5% skimmed milk in Tris-buffered saline (pH7.4) supplemented with 0.05% Tween 20 for 1 hr, the membrane was incubated with primary antibodies. The peroxidase-conjugated secondary antibodies (Jackson ImmunoResearch Laboratory; West Grove, PA), anti-rabbit IgG (cat. no. 111-035-144), and anti-mouse IgG (cat. no. 111-035-146) were diluted (1:5000) and used. Signal detection was performed using enhanced chemiluminescent Western blotting detection reagents (PRN2106; GE Healthcare Bio-Sciences KK, Tokyo, Japan).
Glial Cell Lines and Cell Culture
The rat-glioma-derived C6 (resource no. RCB2854) and RGC-6 (resource no. RCB2783) cell lines were provided by RIKEN BRC through the National BioResource Project of the MEXT, Japan. The C6 cell line was maintained in RPMI1640 (11875-093; Thermo Fisher Scientific, Waltham, MA) supplemented with 10% fetal bovine serum (FBS; S1780-500, Japan Bioserum, Tokyo, Japan), and the RGC-6 cell line was maintained in MEM (M4655; Sigma-Aldrich Inc.) supplemented with 10% FBS. These cell lines were cultured at 37C in a humidified atmosphere of 5% CO2.
To evaluate astrocytic SgIII expression, activation of glioma cells was performed as follows. C6 and RGC-6 glioma cell lines grown in six-well plates to 80% confluency were stimulated with
Statistical Analysis
Statistical significance was evaluated using Student’s t-tests, and p<0.05 was considered significant. Statistical significance is indicated in each figure legend as follows: *p<0.05, **p<0.01.
Results
Expression of SgIII/LacZ in Canonical Endocrine Tissues
The gene trap vector was inserted between exons 11 and 12 of the SgIII genome on chromosome 9 (Fig. 1A). 18 We first examined the SgIII/LacZ expression in SgIII-gtKO mice by immunoblotting using anti-SgIII and anti-β-galactosidase antibodies in the canonical endocrine tissues, where the SgIII expression had previously been established.6,13–15,17 The expression of β-galactosidase was detected in the brain, pituitary gland, and pancreatic islets in SgIII-gtKO, but not in WT mice (Fig. 1B). In WT mice, positive signals for SgIII were detected in endocrine tissues, whereas no significant signal was detected in those of SgIII-gtKO animals (Fig. 2A). To determine whether the gene inserted in the SgIII genomic locus is expressed similarly as the endogenous SgIII, we then examined the consistency between the SgIII and LacZ expression by immunohistochemistry and histochemical X-gal staining. In immunohistochemical staining, a moderate-to-strong positive immunoreaction was observed in the glandular (anterior and intermediate lobes) and neural (posterior lobe) pituitary, adrenal medulla, and pancreatic islets in WT mice, whereas no immunoreaction was detected in SgIII-gtKO mice (Fig. 2B). Equivalently, LacZ expression in these tissues was detected in SgIII-gtKO mice (Fig. 3). The expression pattern in these endocrine tissues correlated well with that of SgIII determined by immunohistochemistry. Thus, it was revealed that the gene-trapping cassettes were well controlled by endogenous SgIII-promoter activity in SgIII-gtKO mice, and that this mouse strain is useful for the sensitive examination of SgIII expression.
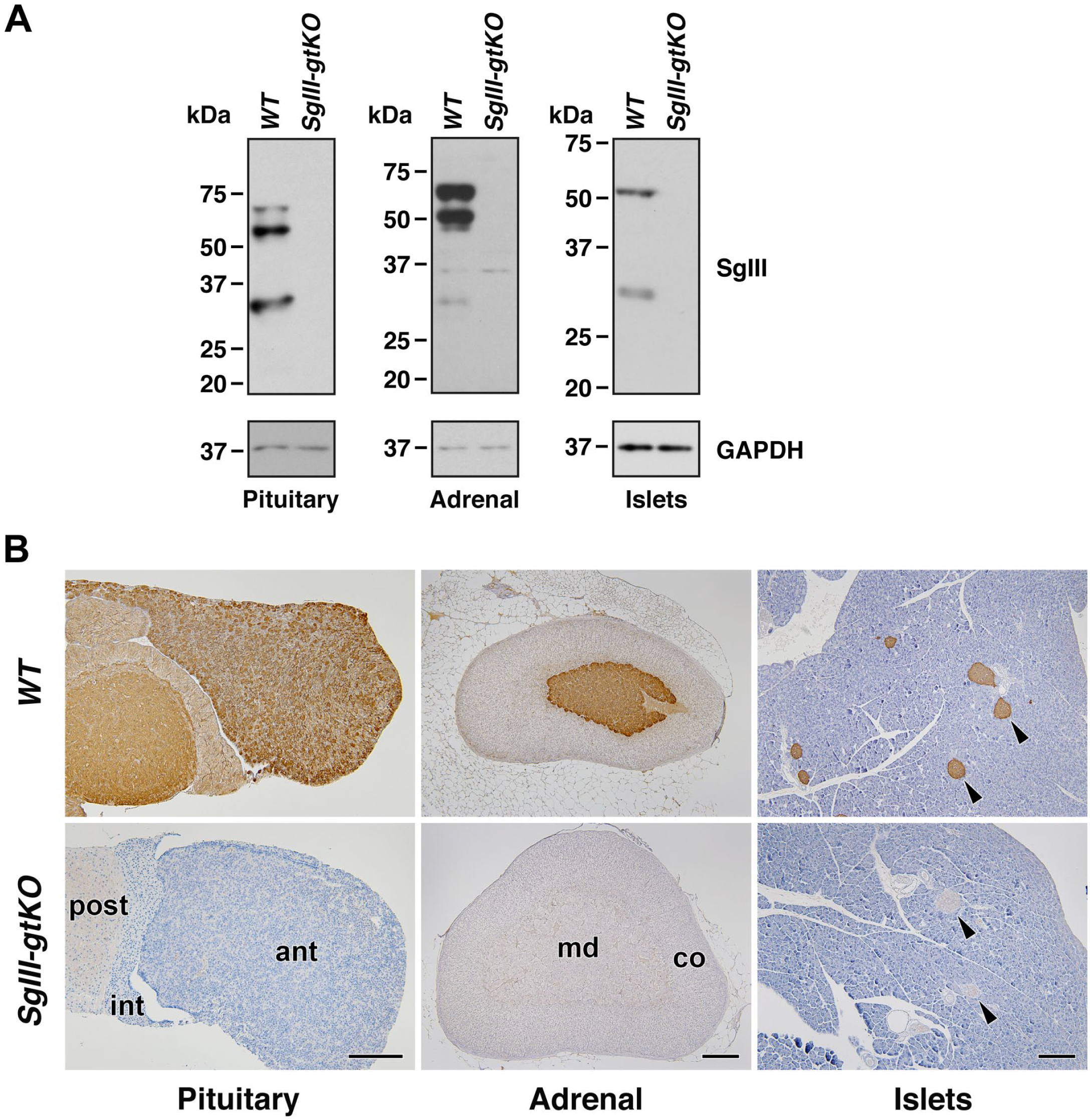
Expression of SgIII in canonical endocrine tissues. (A) Immunoblot analysis. Equal amounts of the protein extracts from the pituitary glands (10 μg), adrenal glands (10 μg), and pancreatic islets (5 μg) of WT or SgIII-gtKO mice were used for SDS-PAGE, followed by immunoblotting using anti-SgIII and anti-GAPDH antibodies. (B) Immunohistochemical analysis. Paraffin sections of pituitary glands, adrenal glands and pancreases from SgIII-gtKO and WT mice were immunostained with anti-SgIII antibody. Arrowheads indicate pancreatic islets. Sections were counterstained with hematoxylin. Scale, 50 μm. Abbreviations: ant, anterior pituitary lobe; int, intermediate pituitary lobe; post, posterior pituitary lobe; co, adrenal cortex; md, adrenal medulla; SgIII, secretogranin III; WT, wild-type; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
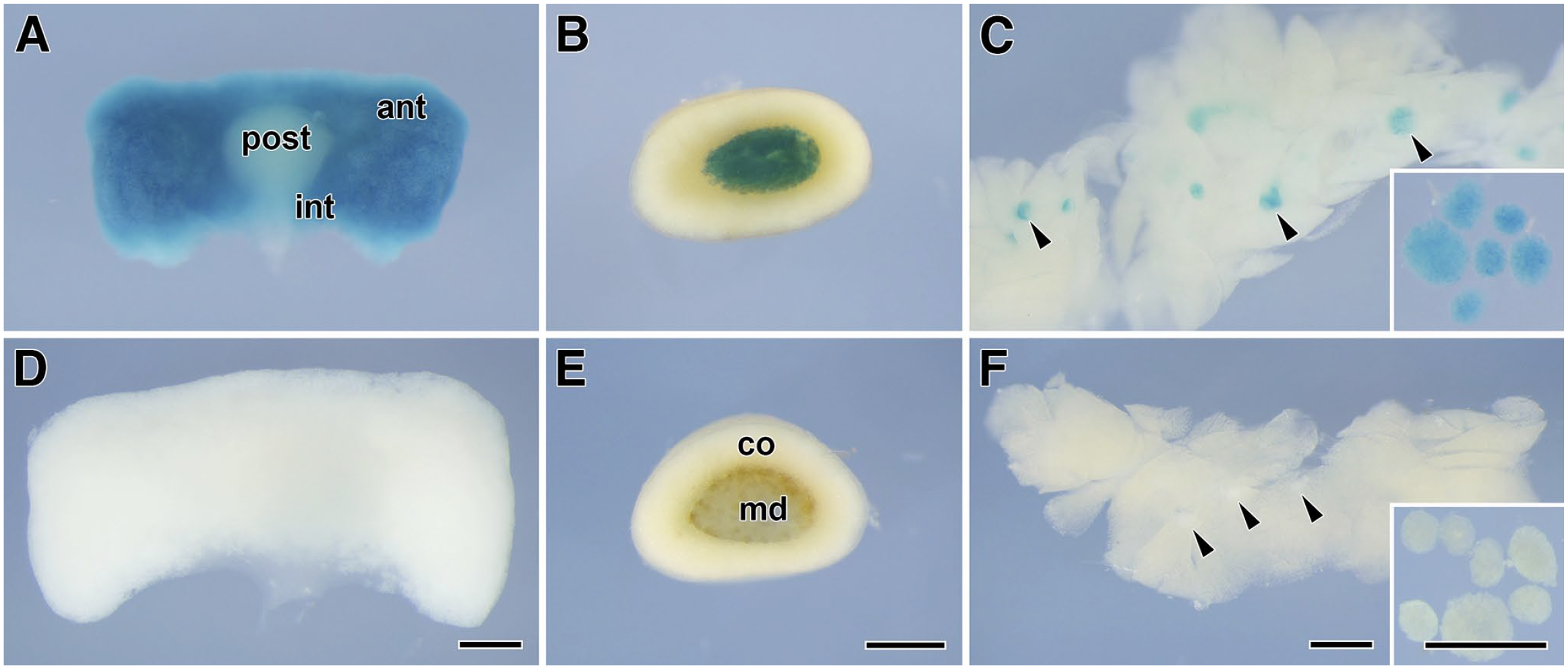
LacZ expression in endocrine tissues. Whole-mount LacZ staining of the pituitary glands (A, D), adrenal glands (B, E), and pancreases (C, F) from SgIII-gtKO (A–C) and WT (D–F) mice. Arrowheads in (C) and (F) indicate the pancreatic islets, and the inset panels show the isolated islets. Scale, 0.5 mm. Abbreviations: ant, anterior pituitary lobe; int, intermediate pituitary lobe; post, posterior pituitary lobe; co, adrenal cortex; md, adrenal medulla; SgIII, secretogranin III; WT, wild-type.
LacZ Expression in the Brain and Spinal Cord
To determine the LacZ expression in the CNS, we then conducted a comprehensive analysis using coronal tissue slices from SgIII-gtKO mice. The β-galactosidase activity was detected in various brain regions and the spinal cord, including the olfactory bulb, cerebral cortex, caudate putamen, hypothalamus, hippocampus, thalamus, amygdala, midbrain, cerebellum, pons, medulla oblongata, and gray matter of the spinal cord of SgIII-gtKO mice (Fig. 4A–E and K–O), whereas no β-galactosidase activity was detected in control WT mice (Fig. 4F–J and P–T). LacZ expression was strong in the cerebral cortex, hippocampus, and molecular layer of the cerebellum and moderate in the caudate putamen, hypothalamus, amygdala, midbrain, medulla oblongata, and dorsal (posterior) horn of the spinal cord.
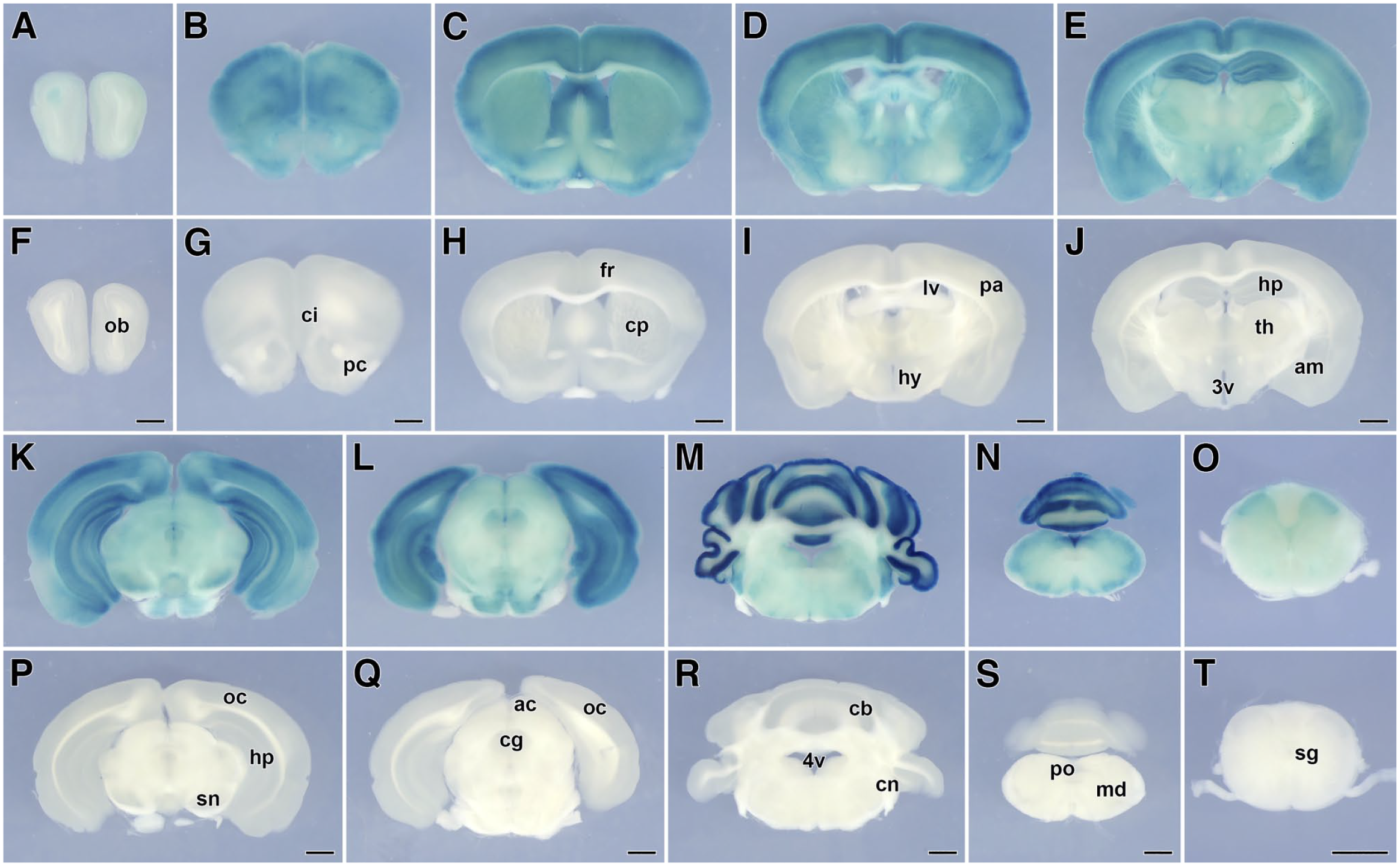
LacZ expression in slice preparations of the brain and spinal cord. The panels from (A) to (E) and (K) to (O) of SgIII-gtKO or (F) to (J) and (P) to (T) of WT mice are arranged from the rostral to the caudal. Scale, 1 mm. Abbreviations: ac, anterior colliculi; am, amygdaloid nuclei; cb, cerebellar cortex; cg, central gray; ci, cingulate cortex; cn, cochlear nuclei; cp, caudate-putamen; fr, frontal cortex; hp, hippocampus; hy, hypothalamus; lv, lateral ventricle; md, medulla oblongata; ob, olfactory bulb; oc, occipital cortex; pa, parietal cortex; pc, piriform cortex; sn, substantia nigra; sg, gray matter of the cervical spinal cord; th, thalamus; 3v, third ventricle; 4v, fourth ventricle; SgIII, secretogranin III; WT, wild-type.
In the slices of cerebral cortex and hippocampus, the cortical layers II and III and the dentate gyrus (DG) exhibited a stronger expression of LacZ (Fig. 5A and B). A moderate β-galactosidase activity was detected in other cortical areas and hippocampal cornu ammonis (CA) fields. The paraventricular nuclei (Fig. 5C and D), dorsomedial nuclei, and arcuate nuclei (Fig. 5E and F) of the rostral and intermediate hypothalamus showed a moderate expression of LacZ. Moreover, the molecular layer and piriform neuron layer of the cerebellar cortex showed a stronger expression of LacZ (Fig. 5G and H).
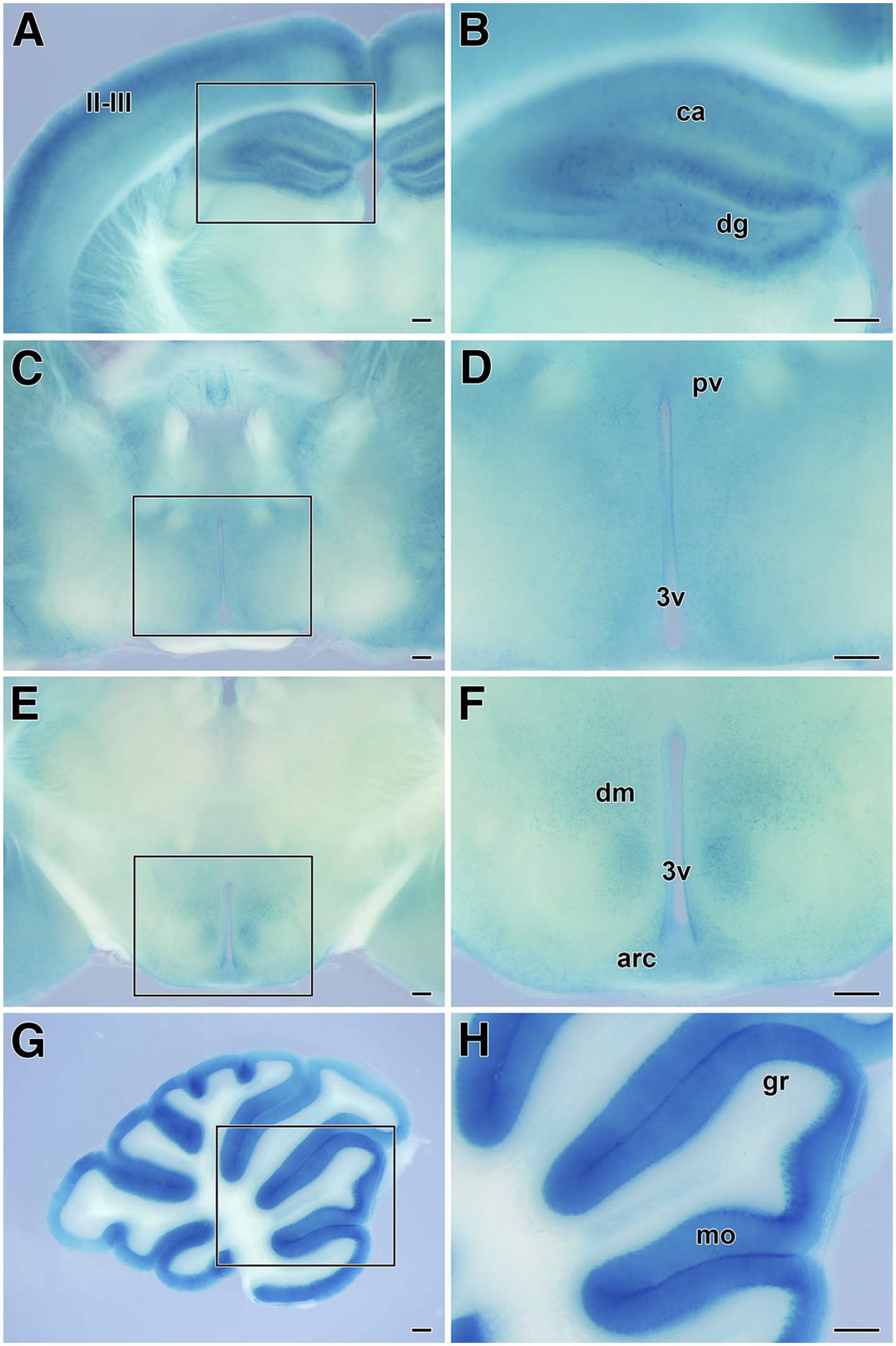
LacZ expression in brain tissues of SgIII-gtKO mice. Strong LacZ expression was observed in the slices of the cerebral cortex and hippocampus (A), rostral hypothalamus (C), intermediate hypothalamus (E), and cerebellum (G). The panels on the right (B, D, F, and H) are enlarged images of the rectangles enclosed by the line in the images on the left. Scale, 250 μm. Abbreviations: arc, arcuate nuclei; ca, cornu ammonis; dg, dentate gyrus; dm, dorsomedial hypothalamic nuclei; gr, granular layer of the cerebellar cortex; mo, molecular layer of the cerebellar cortex; pv, paraventricular hypothalamic nuclei; II/III, layers II and III of the cerebral cortex; 3v, third ventricle; SgIII, secretogranin III.
In the section-level analysis, GFAP-positive astrocytes in the hippocampal DG and cerebral cortex were the main cellular elements that expressed LacZ (Fig. 6A–D). In the hippocampus, CA pyramidal neurons and DG granular cells did not exhibit β-galactosidase activity (Fig. 6B). Conversely, some of the pyramidal neurons of the cerebral cortex expressed LacZ (Fig. 6C). The perivascular structure showed an overlapping of the LacZ blue staining and GFAP-positive perivascular astrocytes (Fig. 6D). In the cerebellar cortex, Purkinje cells, Bergmann glia (which are a GFAP-positive cerebellar-specific subtype of astrocytes), and a limited number of cells located in the granular layer near the piriform neuron layer expressed LacZ (Fig. 6E–G). In addition to these cells, LacZ expression was observed in GFAP-positive astrocytes localized in the medulla oblongata (data not shown). In the lateral ventricle, ependymal cells expressed LacZ (Fig. 6H).
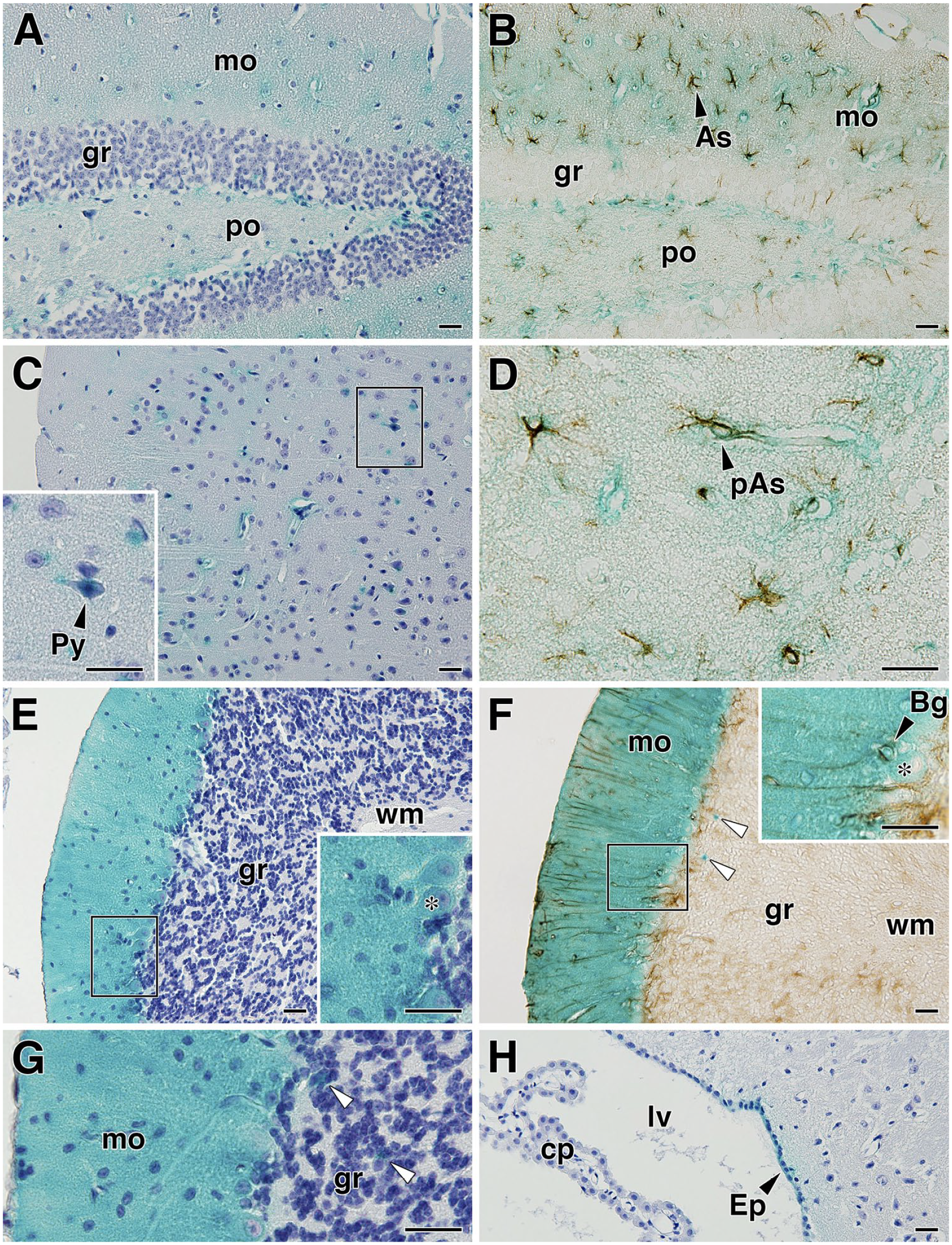
LacZ expression in cellular elements in brain tissues of SgIII-gtKO mice. After LacZ staining, paraffin sections were prepared and counterstained with hematoxylin (A, C, E, G, and H) or reacted with anti-GFAP antibodies (B, D, and F). The panels show the hippocampal DG (A and B), cerebral cortex (C and D), cerebellum (E–G), and lateral ventricle and ependyma (H). The insets in (C), (E), and (F) are enlarged images of the rectangles enclosed by the line. The inset in (C) shows large pyramidal neurons in layer V, and that in (F) shows Purkinje cells and GFAP-immunolabeled Bergmann glia and their radial processes. The asterisks on the insets in (E) and (F) indicate Purkinje cells. The open arrowheads in (F) and (G) indicate LacZ-expression-positive cells in the granular layer. Scale, 20 μm. Abbreviations: Bg, Bergmann glia; cp, choroid plexus; Ep, ependymal cell; gr, granular layer; lv, lateral ventricle; mo, molecular layer; pAs, perivascular astrocyte; po, polymorphic layer; Py, pyramidal neuron; wm, white matter; SgIII, secretogranin III; GFAP, glial fibrillary acidic protein; DG, dentate gyrus.
LacZ Expression in the Peripheral Nervous Tissues
To assess the LacZ expression in the peripheral nervous tissues, we analyzed the retina, optic nerve, and trigeminal ganglion. In the retina, LacZ expression was detected in the ganglionic cell layer, inner granular layer, and outer granular layer (Fig. 7A–C). A weak expression of LacZ was observed in the optic nerves (Fig. 7D), and expression in glial cells was inferred from transverse section images (Fig. 7E). In the trigeminal ganglion, a moderate β-galactosidase activity was detected in the assumed ganglion cells and ganglionic glial cells (Fig. 7F). No β-galactosidase activity was detected in these tissues of WT mice (Fig. 7G–I). To identify the LacZ-expressing cellular elements, we then performed a section-level analysis. In the optic nerve, GFAP-positive astrocytes expressed LacZ (Fig. 7J and K). In the trigeminal ganglion, LacZ expression was observed in some ganglion cells and surrounding satellite cells (Fig. 7L and M).
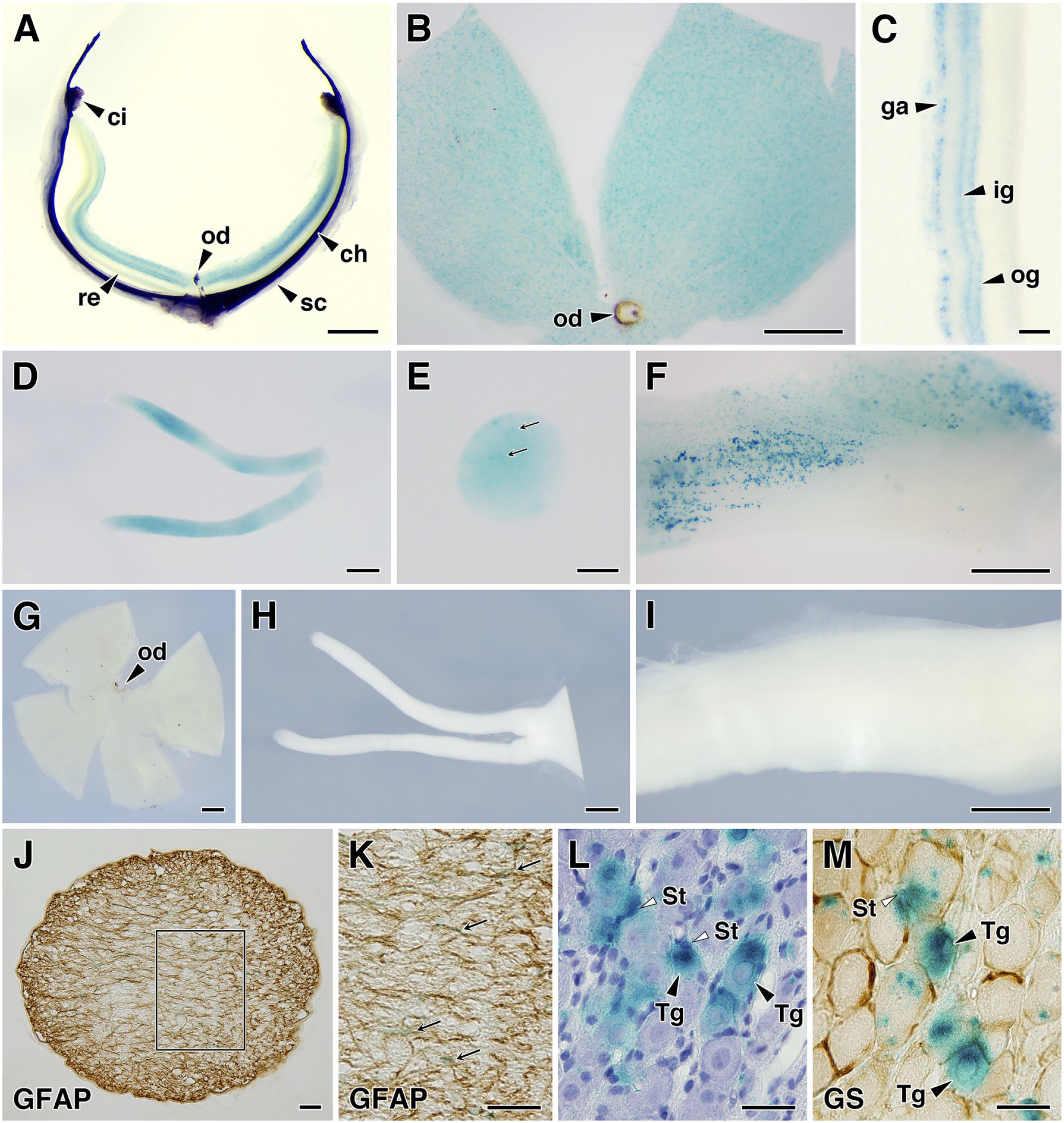
LacZ expression in the peripheral nervous tissues of SgIII-gtKO mice. In the eye, LacZ expression was observed in the retina. The panels show a horizontal slice of the eye (A), a flat-mounted retinal preparation (B), and a transverse retinal section (C). LacZ is expressed in the ganglion cell layer and inner and outer granular layers. In the peripheral nervous system, LacZ expression was observed in the optic nerves (D and E) and trigeminal ganglion (F). Panels (D) and (E) show the shape and a transverse section of the optic nerve. Panels (G)–(I) show the negative control of LacZ staining in WT mice tissues: a flat-mounted retina (G), optic nerves (H), and trigeminal ganglion (I). Panels (J) and (K) show GFAP immunolabeling of the optic nerve transverse section and (K) shows an enlarged image of the rectangle enclosed by the line in (J). The arrows indicate LacZ-positive signals in glial cells in (E) and GFAP-immunolabeled astrocytes in (K). The trigeminal ganglion sections were counterstained with hematoxylin (L) or reacted with an anti-GS antibody (M). Scales, 500 μm (A, B, D, F, and G–I), 100 μm (C and E), and 20 μm (J to M). Abbreviations: ch, choroid; ci, ciliary body; ga, ganglionic cell layer; ig, inner granular layer; od, optic disk; og, outer granular layer; re, retina; sc, sclera; St, satellite cells; Tg, trigeminal ganglion neuron; SgIII, secretogranin III; GFAP, glial fibrillary acidic protein; GS, glutamine synthetase.
Expression of SgIII in the Cerebellum
Although there are reports that SgIII is expressed at several sites in the CNS,11,16,19 knowledge about cerebellar SgIII expression has not been fully obtained. Therefore, we further analyzed the cerebellar expression of SgIII in WT and SgIII-gtKO mice. An immunoblotting analysis revealed the expression of three fragmented bands of SgIII in the cerebellum of WT mice (Fig. 8A), suggesting a similar proteolytic processing from the SgIII precursor as that observed in the adrenal gland, compared with other endocrine tissues (Fig. 2A).
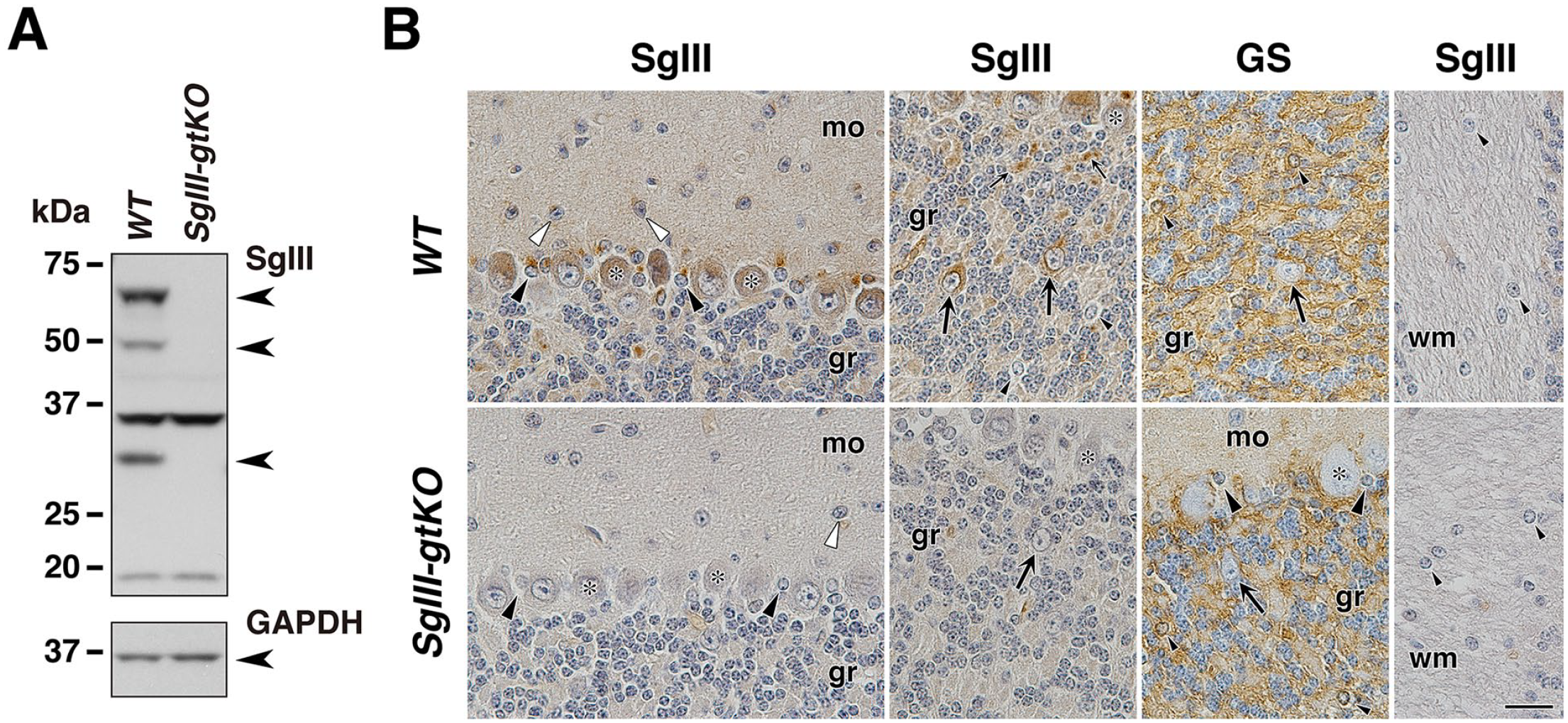
Expression of SgIII in the cerebellum. (A) Cerebellar tissue lysates (40 μg of protein extract) from WT and SgIII-gtKO mice were used for SDS-PAGE, followed by immunoblotting using anti-SgIII and control anti-GAPDH antibodies. Three SgIII-specific bands were detected (arrowheads), and two bands near 37 and 20 kDa were nonspecific. Immunohistochemistry using anti-SgIII (B, C, and E) and anti-GS (D) antibodies. The asterisks indicate Purkinje cells, and the large black arrowheads indicate Bergmann glia in the piriform neuron layer. The open arrowheads indicate basket-like cells. The large arrows indicate Golgi-like cells that do not express GS, and the small arrows indicate the neuropil surrounding the granule cell perikaryon (considered to be the cerebellar glomerulus). The small black arrowheads indicate GS-positive astrocytes in which SgIII expression was not detected in the granular layer. Scale, 20 μm. Abbreviations: gr, granular layer; mo, molecular layer; wm, white matter; SgIII, secretogranin III; GFAP, glial fibrillary acidic protein; GS, glutamine synthetase; WT, wild-type; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
We then analyzed the cellular elements that express SgIII in the cerebellum by immunohistochemistry (Fig. 8B). Purkinje cells and Bergmann glia in the piriform neuron layer and Golgi-like cells showing negative immunoreactivity of GS and localizing in the granular layer expressed SgIII strongly. The molecular layer, basket-like cells, and neuropil around the granule cells showed a moderate level of SgIII expression. Slightly stronger expression in the granular layer was observed in the assumed cerebellar glomerulus, an intertwined mass of nerve fibers consisting of granule cell dendrites and Golgi cell axon terminals surrounding mossy fiber terminals. Conversely, GS immunoreactivity-positive gray matter astrocytes in the granular layer and fibrous astrocytes located in the white matter exhibited negligible SgIII expression.
Glial Expression and Reactive Induction of SgIII
SgIII expression in glial cells, mostly interpreted as astrocytes in the CNS, has been reported. 16 Our LacZ-reporter-based analysis of SgIII expression revealed that cortical and hippocampal astrocytes and cerebellar Bergmann glia constitutively expressed SgIII. In contrast, it was extremely difficult to detect SgIII expression in cerebellar gray matter astrocytes in the granular layer and fibrous astrocytes in the white matter. Among the roles of astrocytes, synaptic glutamate clearance is one of the critical functions in regulating neurotransmitter homeostasis. 28 Therefore, we treated two rat glioma-derived cell lines with similar properties, C6 and RGC-6, with glutamate to investigate the relationship between the changes in SgIII expression level and glutamate stimulation. Although the basal level of SgIII expression was very low in these glioma cells compared with that detected in the control pituitary tissue (Fig. 9A), SgIII expression was significantly induced (1.5−2.0-fold) by glutamate treatment (Fig. 9B).
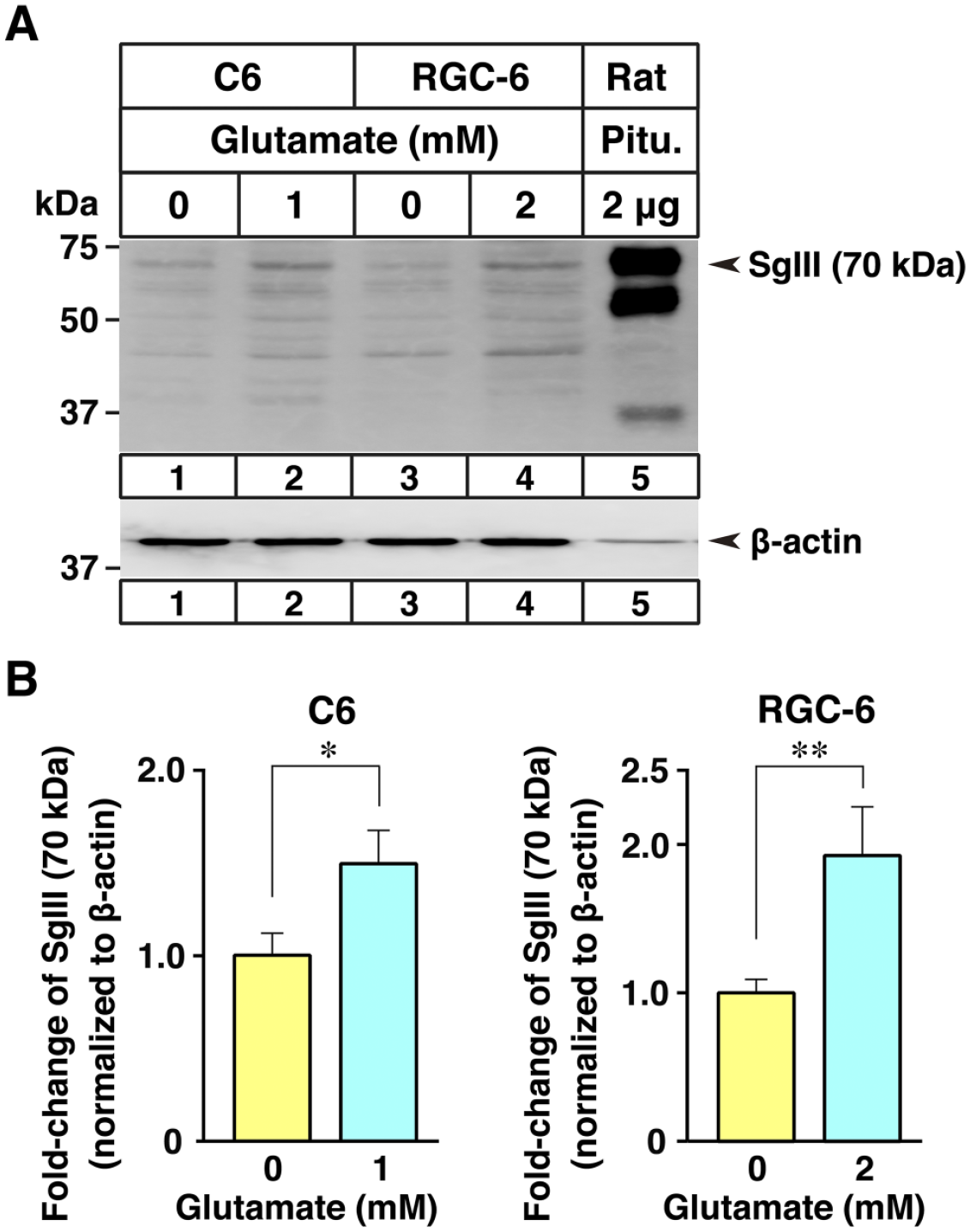
Expression of SgIII in C6 and RGC-6 glioma cells cultured with glutamate. (A) Immunoblot analysis of SgIII (70 kDa) and β-actin in the total cell lysates (20 μg) from C6 and RGC-6 cells cultured with the indicated concentration of glutamate for 24 hr and in rat pituitary lysates (2 μg). (B) Quantification of the immunoblot of SgIII in C6 and RGC-6 cells. The intensities of the immunoreactive bands on the immunoblots presented in (A) were quantified by densitometric analysis. The expression levels of each protein were normalized to that of β-actin and are expressed as arbitrary units, whereas the mean levels in 0 mM glutamate are defined as 1.0. Data are presented as mean ± SEM for each group (n=6 per group: six sample preparations from six independent experiments). *p<0.05; **p<0.01. Abbreviation: SgIII, secretogranin III.
Discussion
We recently established the SgIII-gtKO mouse line and demonstrated that these mice exhibit maladaptation to an inadequate diet and stress because of the impairment of the proteolytic conversion of prohormones in SGs, although the biogenesis of SGs and basal secretion of hormones are not impaired in ordinary conditions. After high-calorie-diet feeding, SgIII-gtKO mice showed marked obesity, which was associated with increased proinsulin accumulation in pancreatic islets and decreased active insulin secretion. Moreover, after loading with chronic restraint stress, the plasma ACTH concentration was not sufficiently increased, and the accumulation of proopiomelanocortin (POMC), which is an ACTH precursor in the pituitary gland, was increased. Based on these results, it was suggested that SgIII is of great importance in the process of conversion to active hormones by limited prohormone degradation during SG formation, especially in the peptide-hormone-secreting endocrine cells. 18
We report here the findings obtained from SgIII-gtKO mice, in which LacZ gene expression is controlled by the authentic SgIII promoter. We analyzed the LacZ expression macroscopically and identified cells/tissues that had not been analyzed to date at the microscopic level. The LacZ staining of the tissues indicated the cell types that endogenously express the SgIII protein. In the canonical endocrine tissues (pituitary glands, adrenal glands, and pancreatic islets), the expression of the SgIII protein and that of LacZ were well matched in immunoblotting and histochemical analyses, suggesting the advantages of the gene trap method. Although the details of SgIII expression verified by in situ hybridization and immunohistochemistry in the rat CNS have been significantly reported, 11 the presentation of topographical images is limited. Therefore, we planned to analyze LacZ expression as a reporter in the mouse CNS and clarify its relative expression intensity. The comprehensive analysis performed in the nervous tissues showed an extensive expression of LacZ in various brain regions, including the cerebral cortex, caudate putamen, hypothalamus, hippocampus, thalamus, amygdala, midbrain, cerebellum, pons, medulla oblongata, and gray matter of the spinal cord. The strongest expression level was observed in the cerebral cortex, hippocampus, and cerebellar cortex. The typical SgIII expression in neurons with neuroendocrine properties, such as those located in the hypothalamic nuclei, as well as in the cerebral cortical pyramidal neurons, hippocampal pyramidal neurons, and cerebellar Purkinje neurons has been reported.11,19 We recently detected SgIII expression in active amine-secreting cells in the gastrointestinal tract and adrenal medulla. 17 The somas of amine-secreting neurons are present in the hypothalamic arcuate nuclei, substantia nigra, central gray matter (catecholamine neurons), and area postrema (serotonin neurons) in the CNS, and a coherent LacZ expression in these brain regions was observed. Furthermore, we found that SgIII is expressed in the ependymal cells that constitute the ventricular wall as a new finding. We then immunohistochemically analyzed SgIII expression in the cerebellum in detail because these cellular elements have not been fully elucidated. We found that SgIII was expressed in Golgi-like cells and basket-like cells, in addition to the previously reported Purkinje cells and Bergmann glia. This was the first report that at least four different cell types express SgIII in the cerebellum. SgIII expression was also observed in presumable cerebellar glomeruli in the granular layer. In the peripheral nervous system, LacZ expression was observed in retinal neurons and in astrocytes in the optic nerves. Furthermore, we found that LacZ was expressed in subtypes of ganglion neurons and satellite cells (ganglional glial cells) in the trigeminal ganglion.
Regarding the cellular elements that express SgIII in the CNS, an important study reported that SgIII is a glial cell granin, the overexpression of which is accompanied by reactive gliosis. 16 SgIII expression in astrocytes, but not in oligodendrocytes, has been reported in the rat somatosensory cortex, hippocampal CA1, corpus callosum, optic nerve, cerebellar granular layer, and Bergmann glia. The expression levels of SgIII in astrocytes were rather remarkable compared with those detected in neurons. Interestingly, transcriptional regulation of the astrocytic expression of SgIII has been suggested to be associated with the trafficking and release of SgIII as secretions, and its upregulation was detected during reactive gliosis. 16 We also observed LacZ expression in astrocytes distributed in the cerebral cortex, hippocampal DG and CA fields, medulla oblongata, and so on. LacZ was expressed abundantly in astrocytes. However, in the cerebellum, Bergmann glia showed a constitutive SgIII/LacZ expression, in contrast to their negligible expression in astrocytes localized in the granular layer and white matter. Thus, based on the results of this study, it also became clear that the expression level of SgIII varies among astrocyte subtypes in the physiological condition.
SgIII expression is not only detected in neurons but also in astrocytes, highlighting the essential diversity of this molecule. Even in neurons, there is a diversity in the localization of intracellular proteins. 11 For example, as shown by ultrastructural immunocytochemistry, the SgIII protein was localized in the perikaryon and fibers in pyramidal cells of the cerebral cortex and seemed to be delivered at the site of presynaptic terminals, whereas SgIII was localized within the dendritic processes in the hippocampal CA3 pyramidal neurons. The intracellular localization of SgIII appeared to vary, from synaptic vesicles in the former to small organelles resembling vesicles of smooth endoplasmic reticulum in the latter. It is interesting to note that there are wide range differences from intracellular localization between neuron subtypes to varied expression levels among astrocyte subtypes. 11
Recently, several lines of research have suggested that SgIII is involved in the pathophysiology of nervous tissues. In patients with Alzheimer’s disease (AD) and an AD transgenic mouse model, a remarkable accumulation of SgIII and CPE was detected in the cortical lesion site, where dystrophic neurites surrounding amyloid plaques and plaque-associated reactive astrocytes exist. 29 Because CPE and SgIII are essential in the processing and targeting of neuropeptides and neurotrophins, their participation in the pathological progression of AD may be suggested. 30 SgIII was expressed in the retina, where it acted as an angiogenic factor in diabetic retinopathy.31,32 Furthermore, SgIII has been suggested to be involved in signal transduction in neurotoxin-induced apoptosis in dopaminergic neuroblasts. 33 Although the pathophysiological significance of the astrocytic SgIII expression is not fully understood, this protein might be involved in the secretory function of astrocytes (recently also called gliocrine). It has been noted that extremely diverse vesicles and secretory organelles, such as synaptic-like microvesicles, dense-core vesicles, lysosomes, exosomes, and ectosomes, are secreted from astrocytes.34,35 Furthermore, astrocytes are traditionally involved in the clearance of the neurotransmitter glutamate as a physiological function. 28 In this regard, we analyzed the changes in SgIII expression level after stimulation of glioma-derived cell lines with glutamate and demonstrated that exposure to glutamate increased SgIII expression. SgIII upregulation is also suggested to be involved in the parkinsonian toxin-induced astrocyte activation, and SgIII may act as a cofactor in astrocyte activation stimulated by various toxins.36,37 Thus, it seems that the action of SgIII as an active substance also has potential importance. Moreover, the fragmentation pattern of SgIII from its precursor in the nervous tissues showed a similar proteolytic processing to that detected in the adrenal gland, which was different from that of other peptide-hormone-secreting endocrine cells.
In summary, we performed an extensive analysis of LacZ gene expression in SgIII-gtKO gene-trapped mice. We demonstrated that this descriptive mapping of the SgIII-expressing cells is significant to provide a more precise localization of this protein in the central and peripheral nervous systems. The comprehensive pursuit of this reporter gene expression allowed us to clarify the previously unpredictable cell types that express SgIII with sufficient sensitivity and confirmed what is already known about the distribution of SgIII cellular expression by immunohistochemistry and in situ hybridization techniques.
Footnotes
Acknowledgements
The authors thank Mr. Ken Tsushima (Akita Prefectural University, Akita, Japan) for his help in mice breeding and RIKEN BRC for providing the C6 (resource no. RCB2854) and RGC-6 (resource no. RCB2783) glioma cell lines.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
HG designed the study, carried out the histological and biochemical experiments, performed the data analysis, and drafted the manuscript. MH and AH prepared mice samples, and performed biochemical analysis and glioma culture experiments. TY and ST contributed to the experimental design, data analysis, and reviewing of the manuscript. All authors have read and approved the final manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Grants-in-Aid from the Japan Society for the Promotion of Science (JSPS) #16K08078 and #20K06418 (to H.G.) and #18K05443 (to M.H.). It was also supported in part by the joint research program of the Institute for Molecular and Cellular Regulation, Gunma University (no.18015, to H.G. and S.T.) and Nihon University College of Bioresource Sciences Research Grant for 2019–2020 (to H.G.).