
Abstract
Cell adhesion to extracellular matrix is a complex process involving protrusive activity driven by the actin cytoskeleton, engagement of specific receptors, followed by signaling and cytoskeletal organization. Thereafter, contractile and endocytic/recycling activities may facilitate migration and adhesion turnover. Focal adhesions, or focal contacts, are widespread organelles at the cell-matrix interface. They arise as a result of receptor interactions with matrix ligands, together with clustering. Recent analysis shows that focal adhesions contain a very large number of protein components in their intracellular compartment. Among these are tyrosine kinases, which have received a great deal of attention, whereas the serine/threonine kinase protein kinase C has received much less. Here the status of protein kinase C in focal adhesions and cell migration is reviewed, together with discussion of its roles and potential substrates.
Background
Ever since the discovery of the protein kinase C (PKC) family as the major phorbol ester-inducible tumor promoters in the 1980s (Blumberg 1988), these serine/threonine kinases have been extensively studied. They represent a major family involved in multiple signaling pathways with well-established roles in essential cellular processes, such as proliferation, differentiation and regulation of gene transcription (Marte et al. 1994; Way et al. 2000). In addition, some members are implicated in the control of cell adhesion and migration, including responses to extracellular matrix macromolecules.
Prominent junctional organelles involved in cell-matrix adhesion are focal adhesions, also referred to as focal contacts. These are signaling organelles at the cell-matrix interface and nearly always lie at the termini of actin-containing microfilament bundles (Hotulainen and Lappalainen 2006). Some microfilament bundles are contractile (Pellegrin and Mellor 2007; Vallenius 2013) so that focal adhesions can be essential sensors of cellular tension. Signaling to the cytoskeleton is integral to focal adhesions, but they can also regulate proliferation and survival.
The major transmembrane components of focal adhesions are integrin receptors (Zaidel-Bar et al. 2007). However, the complexity of the intracellular compartment of these organelles is extraordinary, with over 160 components identified from previous literature (Zaidel-Bar and Geiger 2010). Three more recent proteomics studies using various cellular models and preparation techniques have, however, changed the landscape completely (Humphries et al. 2009; Kuo et al. 2011; Schiller et al. 2011). Combining their data yields 1683 proteins associated with integrin-mediated adhesion (Geiger and Zeidal-Bar 2012). Moreover, there are rather few proteins common to all three analyses. However, as expected, matrix proteins, cytoskeletal and adaptor proteins, receptors, endocytic components, phosphatases and kinases feature strongly. Among these, tyrosine kinases, such as focal adhesion kinase (FAK) and Src have received much attention (Brown et al. 2005; Gardel et al. 2010; Webb et al. 2004; Westhoff et al. 2004). Originally believed to be essential for their assembly, it seems more likely that FAK is involved in the turnover of these structures (Geiger et al. 2001; Ilic et al. 1995). Serine/threonine kinases have, on the other hand, received much less attention, and few representatives appear to be localized to focal adhesions. However, members of the PKC family, most notably PKCα, have been recorded on several occasions.
In this review, we focus on the PKC family and its possible roles in focal adhesion assembly and dynamics. Since focal contacts and focal adhesions are key components for adhesion and migration (Chrzanowska-Wodnicka and Burridge 1996; Kirfel et al. 2004; Ridley et al. 2003) both in vitro and in vivo, their functions are important to understand. Moreover, they are reduced or absent and may have different compositions in tumor cells, which provides further rationale for their study.
PKC Structure and Activation
The PKCs are sub-grouped into three classes, based upon structural homology and cofactor requirements for activation. The classical PKCs (cPKCs) are the α, βI, βII and γ isoforms, which are activated by Ca2+, diacylglycerol (DAG) and phosphatidylserine (PS). PKC δ, ϵ, η and θ comprise the novel PKCs (nPKCs), which lack the Ca2+ binding property, whereas the atypical PKCs (aPKCs) ζ and ι/λ respond only to PS for activation (Fig. 1). This family of 65-79 kDa kinases all have a conserved N-terminal regulatory domain, containing one or two membrane-targeting motifs and a pseudosubstrate that blocks the catalytic site of the C-terminal catalytic domain of the non-activated kinase (Newton 2001). A conserved and essential threonine phosphorylation site is present at their activation loop and threonine or serine phosphorylation sites at the turn- and hydrophobic motifs. The aPKCs are slightly different, having a negatively charged glutamic acid in place of serine or threonine at the hydrophobic site (Keranen et al. 1995).
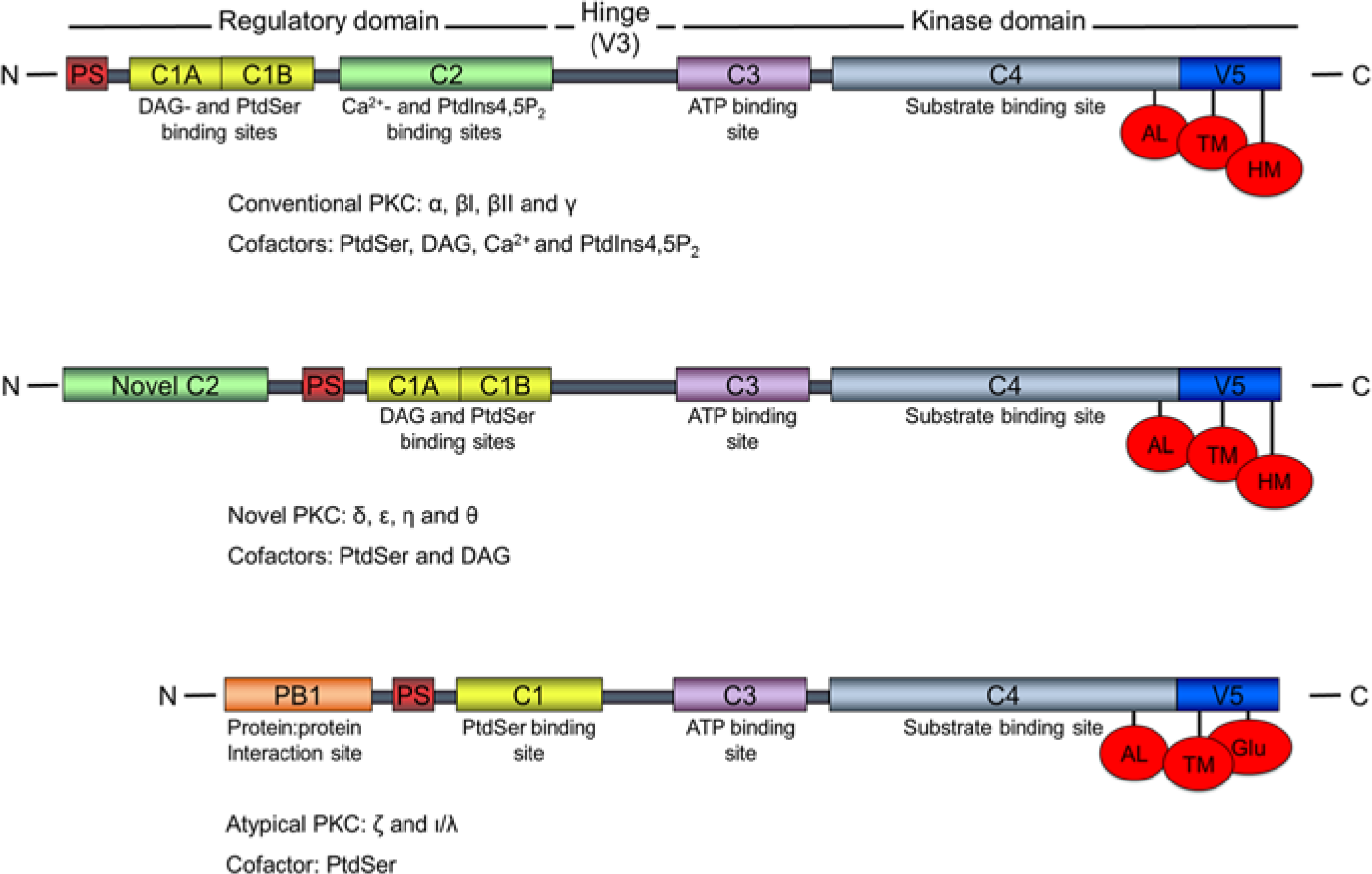
PKC family structures. Subfamily nomenclatures, corresponding regulatory- and kinase domain regions, specific cofactor requirements and binding sites, and the three priming sites are indicated. The figure is not to scale. N: N-terminus; C: C-terminus; PS: pseudosubstrate; C1, C2…: conserved region 1, 2…; V: variable region; hinge: hinge region; DAG: diacylglycerol; PtdSer: phosphatidylserine; PtdIns4,5P2: phosphatidylinositol 4,5-bisphosphate; AL: activation loop; TM: turn motif; HM: hydrophobic motif.
According to the classical view of PKC activation, the newly synthesized protein localizes at the plasma membrane (Borner et al. 1989), where it becomes phosphorylated at the activation loop. This PDK-1–dependent phosphorylation (Dutil et al. 1998) is followed by a rapid phosphorylation at the turn- and hydrophobic motifs of classical and novel PKCs (Borner et al. 1989; Keranen et al. 1995). This primed and ready-to-signal kinase now localizes in the cytosol until activated (Keranen et al. 1995). Upon activation, the kinase is recruited to specific subcellular compartments to phosphorylate downstream substrates. PDK-1–mediated phosphorylation takes place without any cofactor requirements (Dutil et al. 1998) and seems to occur as soon as PKC is expressed, as PDK-1 is a constitutively active enzyme (Mora et al. 2004), and the rate limiting factor of PKC priming phosphorylation seems to be the dissociation of PDK-1 (Gao et al. 2001). Consequently, most PKC species are found in their primed state, and dephosphorylation is probably a pathway either for their degradation or rephosphorylation. Therefore, their subcellular localization and temporal activity are considered to be the most important regulators of PKC activity.
In a canonical process for the activation of primed PKC, second messengers are produced through ligand binding to cognate receptors, such as growth factor receptors, with downstream activation of phospholipase Cγ (PLCγ). This cleaves the membrane lipid phosphatidylinositol 4,5-bisphosphate (PtdIns4,5P2) to yield the co-factors diacylglycerol and inositol trisphosphate (IP3), which can trigger Ca2+ flux into the cytosol from intracellular stores (Parekh et al. 2000). Canonical PKC activation is possible at adhesions, since phospholipase Cγ1 is listed as a focal adhesion component. This may be an indicator of convergence of growth factor/tyrosine kinase signaling with that of integrins, since integrins have no intrinsic kinase activity. Early studies implicating phospholipase in this pathway used fibronectin-coated beads as a proxy for focal adhesion formation (Plopper et al. 1995; Miyamoto et al. 1995).
In terms of focal adhesions, however, an additional mechanism of PKCα activation has been documented (Lim et al. 2003; Oh et al. 1998). The transmembrane proteoglycan receptor, syndecan-4, is frequently associated with focal adhesions, and in the presence of Ptdins4,5P2, PKCα is activated in a ternary complex (Oh et al. 1997, 1998). Consistent with this non-canonical PKCα activation, discrete binding sites for PtdIns4,5P2 have been localized in this protein (Corbalán-García et al. 2003). Furthermore, in neuroblastoma cells, PKCα was insensitive to stimulation by DAG in vivo (Raghunath et al. 2003). PtdIns4,5P2 is important for focal adhesion assembly for several reasons in addition to a potential role in PKC signaling; for example, mediating conformational changes in vinculin is required for its binding to actin and other focal adhesion components (Gilmore and Burridge 1996).
The Life Cycle of Focal Adhesions
At the leading edge of migrating cells, actin fibers polymerize and protrude through the lamellum to form nascent adhesions (Beningo et al. 2001). These very small, rounded structures contain talin, paxillin and tyrosine phosphorylated proteins (Zaidel-Bar et al. 2003, 2004). They are the first structures to monitor the extracellular matrix and, upon receiving a migratory signal, they develop into focal complexes, which additionally contain α-actinin, vinculin and FAK (Zaidel-Bar et al. 2003, 2004) and are located between the lamellum and lamellipodium (Geiger et al. 2009). Like nascent adhesions, focal complexes may govern cell migration or further mature into the larger focal adhesions or focal contacts (Beningo et al. 2001; Geiger et al. 2001) with longer turnover rates (Ivaska 2012). Focal adhesions form at the end of F-actin stress fibers and are sites where the cells experience mechanical force and molecular signaling cues from the ECM, and from where this information is transmitted into the cell interior to induce signaling cascades (Geiger et al. 2001). Large focal adhesions bind strongly to their substratum and mediate cell spreading by transducing a large contractive load onto the stress fibers, whereas smaller adhesions at the leading edge mediate traction forces required for cell migration (Pellegrin and Mellor 2007). As the trailing edge focal adhesions disassemble, the cell body is able to translocate (Kirfel et al. 2004) in the direction of cell migration. In addition to the cellular distribution of PKCs serving as important means to control focal adhesions, the temporal control of individual PKC isoform activation also represents a control mechanism.
Cell Spreading and Focal Complex Formation
When anchorage-dependent cells are seeded onto fibronectin, an interaction between integrin α5β1 and the central integrin-binding domain of fibronectin induces an outside-in activation of the integrin, leading to cell attachment through the activation of PKCϵ (Besson et al. 2001; Disatnik and Rando 1999; Disatnik et al. 2002). This was shown in mouse skeletal muscle cells lacking α5 integrin, which were unable to attach or spread onto the RGD-containing domain, as were α5-expressing cells treated with a PKC inhibitor (Disatnik and Rando 1999). PKC in this system was also potentially able to mediate inside-out activation of α4β1 integrin, initiating cell spreading through an α5β1-independent mechanism. PKCα and δ activities were not required in order for these cells to spread; these isoforms seemed to be activated later and were involved in the formation of adhesion organelles (Disatnik et al. 2002).
In the case of PKCα, this delayed activity probably reflected its importance in focal complex formation during cell spreading. Accordingly, it was reported in MEF cells that PKCα activity was required in order for these cells to form focal complexes during cell spreading (Bass et al. 2007a). This effect of PKCα was mediated through an early activation of Rac1 in cells seeded onto the α5β1-binding domain of fibronectin. Once again, cells were able to spread without PKCα engagement; however, for focal complexes to form, interaction of syndecan-4 with fibronectin to recruit PKCα-mediated Rac1 activation was required, as knock down or drug-mediated inhibition of this kinase failed to activate Rac1 and induce the maturation of focal complexes (Bass et al. 2007b).
Focal Adhesion Formation
Once cells have attached and started to spread, focal complexes start to mature into focal adhesions. In various contexts, PKCα has been discussed as a regulator.
Historically, PKCα was first recorded as a focal adhesion component by Jaken’s group in REF52 cells (Fig. 2) (Jaken et al. 1989). Subsequently, we showed this was true of primary fibroblasts (Woods and Couchman 1992). However, in 3T3 cells, focal adhesions did not stain strongly for PKCα and, in transformed cells, it was noted that PKCα was more frequently localized to the nucleus (Leach et al. 1989). More recent proteomics work with mouse fibroblasts identified PKCα, but no other family members, in the integrin adhesome, interestingly in a pool that was dependent on myosin II activity (Schiller et al. 2011). A similar study with human fibroblasts also identified the kinase, but not reproducibly (Kuo et al. 2011).
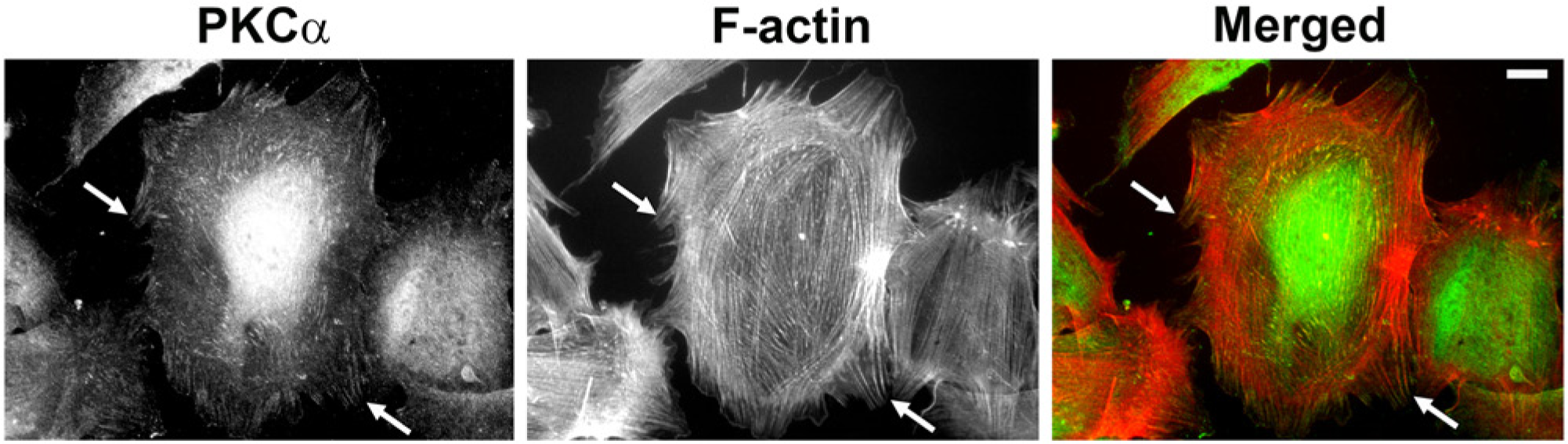
PKCα localizes to focal adhesions. Endogenous PKCα was detected in rat embryonic fibroblasts by indirect immunofluorescence microscopy. Cells were fixed in 4% paraformaldehyde containing 0.1% Tween 20 in phosphate-buffered saline followed by sequential incubations with anti-PKCα (clone M4, Millipore, Billerica, MA) and Alexa488-labelled anti mouse IgG (green; Molecular Probes, Carlsbad, CA). Filamentous actin (F-actin) was visualized using Phalloidin-Alexa568 (red; Molecular Probes). Images were captured on a Zeiss Axioplan epifluorescence microscope, and intensity and contrast were enhanced using Adobe Photoshop CS6 (Adobe Systems Inc., San Jose, CA). Arrows show co-localization of PKCα with termini of actin stress fibers. Bar = 10 μm.
Integrin β1 and PKCα have been suggested to interact directly in a single report, this being in breast carcinoma cells (Ng et al. 1999). In primary fibroblasts, however, a syndecan-4–mediated mechanism has been proposed (Bass et al. 2007a; Horowitz et al. 1999; Oh et al. 1998; Woods and Couchman 1992). These cells seeded onto the RGD-containing central domain of fibronectin will spread but form minimal focal adhesions and stress fibers (Bass et al. 2007a; Woods et al. 1986). However, the addition of the HepII domain of fibronectin, which binds to the heparan sulfate chains of syndecan-4 proteoglycan promotes recruitment to focal complexes and focal adhesions (Bass et al. 2007a; Saoncella et al. 1999; Woods et al. 1986, 2000). Clustering of the proteoglycan depends on binding of PtdIns4,5P2 (Koo et al. 2006; Oh et al. 1997). This complex is then able to recruit and activate PKCα (Oh et al. 1998), inducing a signaling cascade in which phosphorylation of RhoGDIα (on Ser34) releases its binding to the small GTPase RhoA (Dovas et al. 2006, 2010). In its GTP-bound and active state, RhoA binds ROCKII to release its auto-inhibition, leading to the phosphorylation of the myosin light chain of myosin II through myosin light-chain kinase and phosphatase activation and inhibition, respectively (Yoneda et al. 2005). Phosphorylated myosin II is able to bind actin, which activates the intrinsic ATPase activity of myosin II, leading to contraction of the cytoskeleton, and formation of stress fibers and focal adhesions (Chrzanowska-Wodnicka and Burridge 1996; Citi and Kendrick-Jones 1987; Riento and Ridley 2003).
PKCα-mediated regulation of Rho GTPase activation is, however, quite complex, and other scenarios have been reported. PKCα also seems to mediate RhoA inhibition during focal complex formation in the early stage of cell spreading. Accordingly, in the initial α5β1- and syndecan-4-mediated spreading of immortalized mouse fibroblasts onto their respective fibronectin domains, focal complex formation required a p190ARhoGap (p190A)-mediated down-regulation of RhoA, localizing this small GTPase in the cytosol (Bass et al. 2008). The ability of focal complexes to mature into focal adhesions, however, required relocation and activation of RhoA at these structures at later time-points when cells were spread. The interaction of active PKCα with syndecan-4 was required for p190A subcellular localization and for its inhibition of RhoA activity (Bass et al. 2008). Thus, spatiotemporal regulation of PKCα is essential for the formation of focal complexes and, later, their maturation into focal adhesions. In a similar way, CHO cells expressing β1 or β3 integrin do not form focal adhesions in response to cognate antibodies used as ligands but PKC and Rho activity were required additionally (Defilippi et al. 1997). Elfenbein et al. (2009) suggested a different scenario with respect to syndecan-4/PKCα and RhoGDI. In this case, evidence was presented that, in response to FGF2 binding to the heparan sulfate of syndecan-4, PKCα phosphorylated Ser96 of RhoGDI, causing release of RhoG (a Rac homolog), which, in turn, led to the polarized activation of Rac1, and cell migration (Elfenbein et al. 2009).
PKCα Substrates and Interactome
As a serine/threonine kinase, PKCα has many potential substrates. Related to the integrin adhesome, however, the list of known substrates is modest, but almost certainly incomplete. The adhesome website (www.adhesome.org) currently lists 26 interactions, though many of these relate to components that may be peripheral to focal adhesions. Others do not clearly involve the α isoform. Table 1 lists the known substrates and interacting partners with respect to focal adhesions. Prominent substrates are focal adhesion proteins; e.g., filamin A and C, vinculin, VASP and talin. Other potential substrates are tyrosine phosphatases and kinases, but overall the impact of PKC interaction and phosphorylation has yet to be fully evaluated.
PKCα Interacting Partners and Substrates.
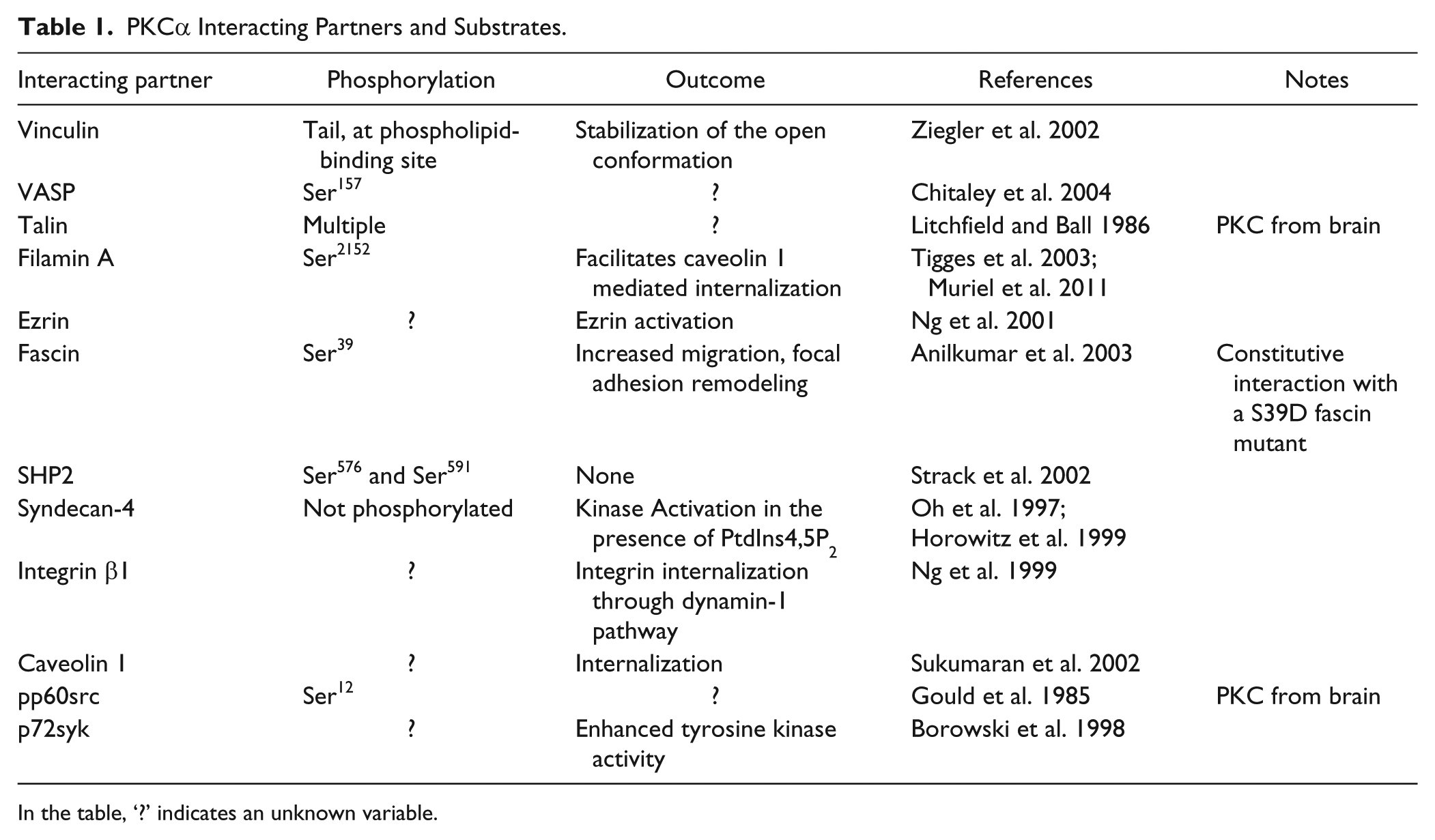
In the table, ‘?’ indicates an unknown variable.
Whereas PKCα may be a focal adhesion component, the counteracting serine/threonine phosphatases have also been reported to be part of focal adhesions. In a number of different cell types, PP1δ was reported as a focal adhesion component, which could be immunoprecipitated with FAK and αV integrin (Villa-Moruzzi et al. 1998). PP1α was reported not to be a focal adhesion component, but later Eto et al. (2007) reported on the appearance of PP1α in very late-staged adhesions that contained tensin. In endothelial cells, PP2A activity was reported to oppose cell migration (Young et al. 2002). With its inhibition, paxillin serine phosphorylation increased, while its tyrosine phosphorylation decreased. This caused dissolution of FAK/Src/paxillin complexes, with increased Src activity leading to enhanced migration. One of the fibroblast adhesome studies (Kuo et al. 2011) reported the involvement of several serine/threonine phosphatases, which will be interesting to follow up. In contrast, given the important roles for FAK and Src, tyrosine phosphatases are well known modulators of integrin-mediated adhesion (Stoker 2005; Cohen and Guan 2005; Broussard et al. 2008).
In the context of focal adhesion assembly, PKCα substrates may not be limited to regulators of small G-proteins of the Rho family. Raf-1 is a direct target of PKCα, which, when activated, signals through the ERK/MAPK cascade (Kolch et al. 1993). In some cells, phosphoERK localizes to focal adhesions (Besson et al. 2001), but in these glioma cells, PKCϵ activity, not PKCα, localized phosphoERK at focal adhesions. This may be an important facet of survival signaling, a function of focal adhesions additional to theirs roles in cytoskeletal organization (Meredith and Schwartz 1997). Whereas the tyrosine kinase FAK is reported not to be a direct substrate for PKCα (Sinnett-Smith et al. 1993; Vuori and Ruoslahti 1993), in some way its activity might be regulated by PKC (Tu et al. 2001). However, the molecular basis for this is unclear. Focal adhesions contain a plethora of cytoplasmic protein components, many of which could be substrates for PKCα. The major focal adhesions adaptor protein paxillin is serine phosphorylated upon cell adhesion onto extracellular matrix substrates and PKCs may directly or indirectly mediate paxillin phosphorylation at adhesion sites (Nichilo and Yamada 1996; Bellis et al. 1997).
The interaction between PKCα and caveolin 1 is worthy of mention, as this hints at a possible role for the kinase in adhesion turnover. PKCα phosphorylation of filamin A at Ser2152 is required for caveolin 1 internalization (Muriel et al. 2011). This may be related to β1 integrin endocytosis in a dynamin- and caveolin-dependent manner, in response to syndecan-4/PKCα activation by ligand binding to the heparan sulfate chains of the syndecan (Bass et al. 2011).
Focal Adhesion Disassembly
In addition to their role in focal adhesion assembly, PKCs might therefore be implicated in the disassembly of these structures. Phosphorylation of paxillin at Ser178 is known to be important for cell migration and an Ala178 non-phosphorylatable mutant of this protein induced focal adhesions of increased size in NBTII cells, implicating a possible role for paxillin phosphorylation in focal adhesion disassembly and cell migration (Huang et al. 2003). In hepatocellular carcinoma cells, hepatocyte growth factor binding to its receptor c-Met induced cell migration through ERK-mediated paxillin phosphorylation, while also stimulating PKC activation (Hu et al. 2013). Using a PKC isotype-specific shRNA screen and drug treatments in this cell system, PKCα and δ isoforms were found to inhibit ERK-mediated phosphorylation of paxillin at Ser178, probably through mediating endocytosis and degradation of the c-Met receptor. Yet, the expression of these isoforms was still required for actin polarization and migration of this cell line. This apparent discrepancy between PKC-mediated inhibition of ERK activity and the simultaneous promotion of cell migration once again demonstrates the general theme of subtle spatiotemporal regulation of PKC activity in cell migration and focal adhesion dynamics. Accordingly, time-course experiments showed that both the membrane level and activity of PKC fluctuated three times within 24 hours, matching the phosphorylation levels of its downstream signaling proteins, as well as a temporal actin polarization (Hu et al. 2013).
Although not strictly related to cell adhesion, Violin et al. (2003) provided new insight into PKC and substrate dynamics. In a series of FRET microscopy studies, they showed an oscillatory phosphorylation/dephosphorylation of membrane-associated PKC substrates that immediately followed similar cytoplasmic Ca2+ oscillations. Moreover, a YFP-PKCβII probe also showed similar oscillations in membrane association with cycling times of minutes. These studies were carried out in HeLa cells, but whether these observations can be extrapolated to PKC in adhesions is not known. Possibly, this spatiotemporal regulation of PKC, and substrate phosphorylation, is necessary to provide focal adhesion turnover that is timed to meet the specific requirements of cell migration. It would be interesting to know whether the serine phosphorylation of paxillin or other PKC substrates in adhesions exists to the same degree at both the leading and trailing edges of cells, and whether cell migration influences the spatial regulation of PKC-dependent, focal adhesion component phosphorylation.
Connecting Focal Adhesions to the Cytoskeleton
Vinculin is one of the focal adhesion proteins known to mediate tension and, in its phosphorylated state, it is active and able to bind F-actin to connect focal adhesions with the actin cytoskeleton (Grashoff et al. 2010; Johnson and Craig 1995; Massoumi and Sjölander 2001), and possibly transduce tension monitored at focal adhesions into signaling cascades throughout the cytoskeleton. Besides its important role in focal adhesions, vinculin is also present at cell-cell contacts. Phorbol ester-induced activation of PKCα causes vinculin to dissociate from its cell-cell interaction partner, α-catenin, in intestinal epithelial cells, and induces its accumulation with PKCα at focal adhesions, leading to increased cell adhesion (Massoumi and Sjölander 2001). Although not investigated, this might also induce a migratory phenotype, which requires cell-cell contacts to become weaker and focal adhesions to grow at the leading edge of migrating cells. PKCα-mediated phosphorylation of vinculin was also seen in HeLa cells spreading on collagen (Ziegler et al. 2002). In this system, PtdIns4,5P2 was suggested to recruit vinculin to focal complexes—this interaction exposing a docking site for PKCα to phosphorylate the vinculin tail (Ziegler et al. 2002). As discussed, PtdIns4,5P2 is important for PKCα recruitment to some focal adhesion protein complexes. In this setting, however, an additional role for PtdIns4,5P2 probably exists, where it disrupts the closed intramolecular head-tail conformation of vinculin and induces a parallel oligomerization to expose its PKCα binding site (Johnson and Craig 1995; Ziegler et al. 2002).
Vinculin is one of the first components to be localized at cellular adhesion sites and is a commonly used marker of both the initial focal complexes as well as focal adhesions (Geiger et al. 2001). The interaction between PKCα and vinculin was transient and only observed at early time points, suggesting a role for PKCα in initiating focal adhesion formation, at least in regards to vinculin phosphorylation (Ziegler et al. 2002). An interaction between PtdIns4,5P2 and vinculin has been shown to inhibit the association between vinculin and F-actin, while promoting vinculin interactions with focal adhesion proteins (Steimle 1999). This might explain the observed transient nature of the PKCα/vinculin interaction, in that PKCα would then be transiently recruited to phosphorylate vinculin, which then dissociates from PtdIns4,5P2, and also PKCα, to be free for its interactions with F-actin in its now phosphorylated state. Therefore, PtdIns4,5P2-mediated temporal control of PKCα localization near its vinculin substrate has important implications in connecting focal adhesions to the cytoskeleton.
Focal Adhesions and Cell Migration
When cells receive a migratory stimulus, they polarise to migrate in the stimulatory direction (Li and Gundersen 2008), a process that often involves the atypical family of PKCs (Etienne-Manneville and Hall 2001; Iden and Collard 2008). The migrating cells undergo repeated cycles of lamellipodial protrusion of their leading edge, produced through F-actin polymerization (Pellegrin and Mellor 2007), to anchor this site onto the substrate (Abercombie et al. 1971), enabling contraction of actomyosin fibres throughout the cell body (Pellegrin and Mellor 2007). The generated traction force displaces the cell body in the forward direction, due to retraction of the trailing edge and the concomitant disassembly of the trailing edge focal adhesions, releasing cells from their substratum at the cell rear (Kirfel et al. 2004; Pellegrin and Mellor 2007).
In contrast to the initial cell adhesion and spreading, during which primarily the ϵ isoform of PKC is activated (Disatnik et al. 2002), an early activation of PKCδ is required for migration of many cell types (Chaudhuri et al. 2005). This isoform phosphorylates syndecan-4 at Ser183 (Murakami et al. 2002), inhibiting syndecan-4 interaction with PtdIns4,5P2 and subsequently oligomerization (Koo et al. 2006) as well as PKCα binding and activation (Koo et al. 2006; Murakami et al. 2002). Therefore, PKCα and δ appear to have contrasting roles in cell adhesion and migration. Possibly, the PKCδ isoform is required for the initiation of migration, whereas PKCα is important for an initiated migration to proceed, where an earlier activation would promote cell spreading. Accordingly, in endothelial cells, prolonged drug-mediated activation of PKCδ negatively regulated cell migration by binding and serine phosphorylating syndecan-4 to inhibit its PKCα association and activation (Chaudhuri et al. 2005). One function of PKCδ might then be to inhibit PKCα’s effects on focal adhesions and cell spreading (Chaudhuri et al. 2005) during the initiation of migration. Additionally, PKCδ-mediated inhibition of PKCα could be required for cells to form the leading edge protrusion, which is required for cells to migrate. Interestingly, both the α and δ isoforms of PKC have been identified at nascent adhesions (Jockusch et al. 1995). Fibronectin-mediated activation of PKCα inhibits the formation of nascent adhesions and cell protrusions by phosphorylating fascin (at Ser39) to block its association with actin and thereby its functioning in F-actin polymerization (Adams 2004). A dynamic activity of PKCα is probably required in this context. Accordingly, both Ser39 phosphorylated and dephosphorylated fascin species are important for carcinoma cell protrusion and migration on laminin (Hashimoto et al. 2007). Contrasting this, though, the effect of Ser39 has been suggested to be ECM substrate-specific, as phosphorylation of this residue was required for focal adhesion formation and cell spreading specifically on fibronectin (Adams et al. 1999), with reduced cell migration (Anilkumar et al. 2003). The interaction between wild-type fascin and PKCα was, however, temporal (Anilkumar et al. 2003), which does support an involvement of PKCα in focal adhesion formation during cell spreading, but does not exclude its importance in the regulation of focal adhesion dynamics during the later stages of cell migration.
An important aspect of cell migration is the replenishment of integrin at the leading edge of cells, to control the dynamics of focal adhesions and cell polarity, through integrin-mediated regulation of downstream adhesion components (Caswell et al. 2009). The exact mechanism of integrin recycling in cell migration is, however, still a matter of investigation. A controversy seems to exist between a model where integrins from the trailing edge are internalized to be recycled at the leading edge focal adhesions or, alternately, that most of the internalization and recycling occurs adjacent to the leading edge (Bretscher 1996; Caswell and Norman 2006). As the formation of adhesions and their turnover is very dynamic at the leading edge of cells, it seems plausible that integrins at these sites would be internalized and recycled in order to regulate such dynamics.
Integrins are internalized into early endosomes to be sorted for degradation or recycling through a long or short pathway loop. Short-loop recycled integrins, such as αvβ3, are, upon stimulation, rapidly brought back to the plasma membrane through a Rab4-dependent mechanism, whereas other integrins are transported to the perinuclear recycling compartment for relocation to the membrane through a Rab11-mediated process of long-loop internalization (Caswell and Norman 2006).
PKCs are known to regulate cell-surface integrin levels in several cell types. Since the earlier studies on cell adhesion were conducted, it has more recently been suggested that PKCϵ is a key kinase in β1 integrin recycling, an essential part of the spreading and migratory process (Ivaska et al. 2002, 2005), while Besson et al. (2002) proposed an interaction between these two proteins mediated by RACK1. Evidence that PKCϵ locates to nascent or mature focal adhesions is lacking, but this is not inconsistent with the key roles for the kinase in trafficking, rather than signaling, at adhesions.
Focal Contacts and Focal Adhesions
The terms focal contact and focal adhesion are both used interchangeably to describe the same structures. However, a distinction between them was proposed many years ago (Couchman and Rees 1979): rapidly migrating cells established small, short-lived structures that would eventually give way to larger, more stable adhesions that were more compatible with anchorage. The principle that focal adhesion size is inversely related to migration speed has been revisited several times (Lauffenburger and Horwitz 1996; Huttenlocher and Horwitz 2011). In the intervening period, however, no clear biochemical distinctions between these two potential classes of adhesions have been described (Geiger et al. 2009). This underlies the continuing lack of knowledge on what governs the turnover rate of focal adhesions. In some cells, moreover, sliding of these structures with respect to the matrix-coated substrate has been reported (Goldyn et al. 2009; Smilenov et al. 1999) but, once again, it is not known how this property relates to the molecular constitution of adhesions. Whether PKC isoforms have roles to play in the stability or half-life of adhesions is unknown. Great strides have been made recently by proteomics analyses that have identified a huge number of potential components (Geiger and Zeidal-Bar 2012); some may be dispensable for focal adhesion assembly (Ilic et al. 1995), whereas others, such as, vinculin and talin (Geiger et al. 2009), appear essential.
Atypical PKCs and Migration
There has been much recent focus on the atypical members of the PKC family. These are PKCζ and PKCλ in the mouse and PKCζ and PKCι in human. The λ and ι isoforms are, in fact, about 98% identical, while PKCζ has approximately 70% identity to the other isoforms (Newton 1997). They cannot be activated by phorbol esters, but are activated by phospholipids such as phosphatidylinositol 3,4,5 trisphosphate and phosphatidic acid (Nakanishi et al. 1993; Limatola et al. 1994; Xiao and Liu 2013). As with other PKC enzymes, full activity also requires PDK-1 activity (Hirai and Chida 2003). Their importance for cell migration lies in part from complexes formed with members of the Rho family; e.g., Cdc42 and Rac, together with PAR6 and PAR3 (Joberty et al. 2000; Lin et al. 2000).These are directed to the leading edge of motile cells and regulate polarity. Since Cdc42 and Rac direct protrusive activity, these polarity complexes localize this activity for efficient locomotion. PAR6 interacts with the PB1 domain at the N-termini of the atypical PKCs, a domain that is unique to these isoforms (Xiao and Liu 2013). Linkage with integrins is also provided through an interaction between PAR complexes and Tiam1, a Rac guanine exchange factor. Tiam 1 also binds talin directly, and so indirectly with integrins (Wang et al. 2012). In this way, it is proposed that Rac activity is directed to integrin-mediated adhesions, consistent with adhesion turnover in cell migration.
In non-small cell lung cancer, PAR6 has been shown to associate with, and be phosphorylated by, the TGF-β receptor II (Gunaratne et al. 2013). In this way the atypical PKC/PAR complex promotes the TGF-β–driven epithelial-mesenchymal transformation widely reported to be a key aspect of cell motility acquired by carcinomas (Heldin et al. 2012). Many tumor cell types and some normal cells assemble adhesion complexes known as podosomes or invadopodia (Gimona et al. 2008). Even though they contain some cell-surface and cytoskeletal components in common with focal adhesions, they are nevertheless distinct. Moreover, invadopodia are sites of not only extracellular matrix adhesion, by virtue of integrins, but also matrix degradation, mediated largely by metalloproteinases. Atypical PKCs are not required for assembly of these organelles, where roles for PKCα could be important (Xiao et al. 2013), but PKCζ can be recruited by PKCδ, for example (Xiao et al. 2010, Xiao and Liu 2013). PKCζ is involved in the recruitment, release and activation of MMP9, an important matrix-degrading proteinase (Xiao and Liu 2013). In total, therefore, multiple PKC isoforms may be involved in the establishment and activity of podosomes and invadopodia, key organelles of extracellular matrix invasion.
In Vivo Adhesions and Disease
Structures analogous or homologous to focal adhesions have been described in vivo on many occasions (Kano et al. 1996; Turner et al. 1991). More recently, advanced microscopy has identified these structures in cells grown in three-dimensional matrices (Cukierman et al. 2001; Kubow and Horwitz 2011). However, others suggest that focal adhesions are not visible in cells grown in three-dimensional matrices (Fraley et al. 2010). Previous electron microscopy identified structures known as fibronexus in granulation tissue, associated with what most likely were myofibroblasts (Singer 1979). Dense bodies of smooth muscle are often proposed to be in vivo focal adhesion equivalents (Turner et al. 1991). However, these structures, much like the costameres of skeletal muscle, are very stable, in contrast to the transient adhesions of migratory cells. A proteomic comparison would be interesting but, so far, PKCα, although present in focal adhesions of tissue cultured cells, has not been shown as a component of costameres; in comparison, PKCϵ has been shown to be involved (Vanwinkle et al. 2002). Syndecan-4, on the other hand, which binds PKCα, is a costamere component (Vanwinkle et al. 2002), perhaps interacting with α-actinin in these structures (Okina et al. 2012), consistent with known interactions mapped in fibroblasts and by in vitro studies (Choi et al. 2008; Greene et al. 2003; Okina et al. 2012). Overall, however, it does suggest that PKCα may be associated with dynamic structures rather than very stable ones.
Knockout mice lacking PKCα have been produced and have at least two interesting phenotypes. They have skin wounding defects (Hara et al. 2005), reminiscent of both syndecan-1 (Stepp 2002) and -4 (Echtermeyer et al. 2001) knockout mice, which also show defects in postnatal repair. In addition, PKCα null mice, or those subject to pharmacological inhibition of the enzyme, show dramatic protection from cardiac failure (Liu and Molkentin 2011). Whether this relates to cell-matrix interactions is less clear, but it is interesting that recent data suggest that syndecan-4 is essential for compensatory hypertrophy in the pressure-overloaded heart (Finsen et al. 2011). This was linked to signaling through the phosphatase calcineurin and nuclear factor of activated T-cell (NFAT), which is reduced in the absence of the proteoglycan. This, and related data suggest that syndecan-4 can be a tension sensor independent of other focal adhesion components (Li and Chaikoff 2002). Further work is required to dissect this pathway, but it will be interesting to ascertain whether syndecan-4 is unique among the family in regard to this property.
Conclusions
Protein kinase C has central roles in signaling in response to many extracellular ligands, and can influence many aspects of cell behavior. Its association with focal adhesions, particularly with regard to PKCα, has been recorded several times but, although its interactions are known, the specific responses and impact on adhesion dynamics still await elucidation. Within adhesions there are a plethora of potential substrates, and it will take much further work to understand its molecular functions.
Footnotes
Declaration of Conflicting Interest
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: Support from the Danish National Research Foundation, Lundbeck Fonden, the Novo Nordisk Fonden, and the department of Biomedical Sciences at the University of Copenhagen are gratefully acknowledged.