
Abstract
PRMT5 has been reported to be involved in the processes of tumor progression at various steps. The aim of this study was to examine the role of PRMT5 in epithelial ovarian cancer (EOC). In this study, PRMT5 and Ki-67 expression were examined by immunohistochemistry (IHC) in cohorts of normal, benign, and cancerous ovarian tissues. PRMT5 overexpression was observed in 83.1% (98/118) of EOCs, and it was significantly associated with serous type, poor differentiation, advanced tumor stage, lymph node invasion, presence of residual tumor, and high expression of Ki-67 (p<0.05, respectively). Moreover, overexpression of PRMT5 was an independent prognostic marker for decreased overall survival and progression-free survival in univariate survival analysis and multivariate Cox regression analysis. In ovarian cancer cell lines A2780 and SKOV3, PRMT5 knockdown by siRNA inhibited cell growth/proliferation and induced apoptosis via upregulation of E2F-1. These results suggest that overexpression of PRMT5 correlates with an aggressive malignant phenotype and may constitute a novel prognostic factor for EOC. Thus, PRMT5 may represent a clinically effective new target for therapy of ovarian cancer.
Ovarian cancer is one of the leading causes of death from gynecological malignancies. Despite the improvement in surgical techniques and the advent of more effective therapeutics, the overall 5-year survival has increased only from 36% to 44% during the past three decades (Siegel et al. 2012). More than 80% of ovarian cancers are of epithelial origin, which are highly invasive, respond poorly to therapies, and are usually detected at advanced stages, resulting in poor prognosis (Auersperg et al. 2002; Jordan et al. 2006). To improve better prognostic and predictive assays and develop new therapeutic strategies for epithelial ovarian cancer (EOC), we must understand the molecular mechanisms underlying EOC.
Arginine methylation is becoming more acknowledged as an important type of posttranslational modification found in both nuclear and cytoplasmic proteins (Bedford and Richard 2005; Bedford and Clarke 2009). The posttranslational modification of protein arginine methylation is widely appreciated, playing a vital role in cellular function. The methylation of arginine residues is catalyzed by the protein arginine methyltransferases (PRMTs), which can catalyze the addition of one or two methyl groups to the guanidine nitrogen atoms of arginine. To date, PRMT family have been found 11 members in humans (including PRMT1-11), and most PRMTs, with the exception of PRMT2, PRMT10, and PRMT11, have been shown to have enzymatic activity and can catalyze arginine methylation (Bedford and Richard 2005; Pal and Sif 2007). Based on the primary protein sequence and the specificity for distinct substrates, PRMTs are mainly classified as type I and type II enzymes. These two types of enzymes catalyze the formation of a mono-methylated (MMA) intermediate; subsequently, type I PRMTs (PRMT1, 3, 4, 6, and 8) further catalyze the production of asymmetric dimethylation of arginine residues (aDMA), and type II PRMTs (PRMT5, 7, and 9) catalyze the formation of symmetric dimethylation of arginine residues (sDMA) (Bedford 2007). Arginine methyltransferase 5 (PRMT5), the first identified type II enzyme (Branscombe et al. 2001), is 637 amino acids long and maps to in human chromosome 14q11.2 (Krause et al. 2007) and is involved in a variety of biological processes, including ribosome biogenesis (Ren et al. 2010), assembly of the Golgi apparatus (Zhou et al. 2010), cellular differentiation (Dacwag et al. 2007, 2009; Mallappa et al. 2011), cellular proliferation (Scoumanne et al. 2009), apoptosis (Cho et al. 2012), and germ cell specification (Ancelin et al. 2006; Eckert et al. 2008). PRMT5 can methylate H3R8 and H4R3 to induce silencing of tumor suppressor genes, such as ST7 and NM23 (Pal et al. 2004). In addition, it has been shown to interact with many other genes, such as p53 (Scoumanne et al. 2009), E2F-1 (Cho et al. 2012), IL-2 (Richard et al. 2005), cyclin E1 (Fabbrizio et al. 2002), TRAIL receptor (Tanaka et al. 2009), the CDK4 complex (Aggarwal et al. 2010), and E-cadherin (Hou et al. 2008), to participate in many cellular processes. In humans, PRMT5 is widely expressed in many organs (such as heart, muscle, and testis) at different levels (Pollack et al. 1999). Aberrant expression of PRMT5 is found in a wide variety of human diseases, especially in many cancers, including leukemia and lymphoma (Pal et al. 2007; Wang et al. 2008), gastric cancer (Kim et al. 2005), breast cancer (Powers et al. 2011), and colorectal cancer (Cho et al. 2012), and is related to poor outcome in patients with breast cancer and colorectal cancer (Powers et al. 2011; Cho et al. 2012).
However, little is known about the role of PRMT5 in ovarian cancer. In this study, we investigated the expression profile of PRMT5 protein in ovarian cancer tissue by immunohistochemistry (IHC) and determined its correlation with clinicopathological features and its possible prognostic significance. Then we further confirmed the action of PRMT5 in regulating cell growth/proliferation and apoptosis in ovarian cancer cell lines A2780 and SKOV3.
Materials and Methods
Patients and Samples
EOC (n=118), benign ovarian tumor (n=20), and normal ovary (n=12) samples were collected from the Department of Gynecology, Qilu Hospital of Shandong University, between January 2005 and December 2008. The age of the patients ranged from 24 to 86 years, with a median of 59 years. All patients with only gynecological tumors were treated without any preoperative therapy. Twelve normal ovarian samples were from women subjected to hysterectomy with oophorectomy due to non-ovarian pathologies. Tumor stage was in accordance with the International Federation of Gynecology and Obstetrics (FIGO) criteria, whereas tumor grade and histological type were determined following World Health Organization standards. The clinicopathological parameters are shown in Table 1. The study was approved by the Institutional Medical Ethics Committee of Qilu Hospital, Shandong University.
Relationship between the Expression of PRMT5 and Clinicopathological Characteristics or Ki-67 in Human Epithelial Ovarian Cancers
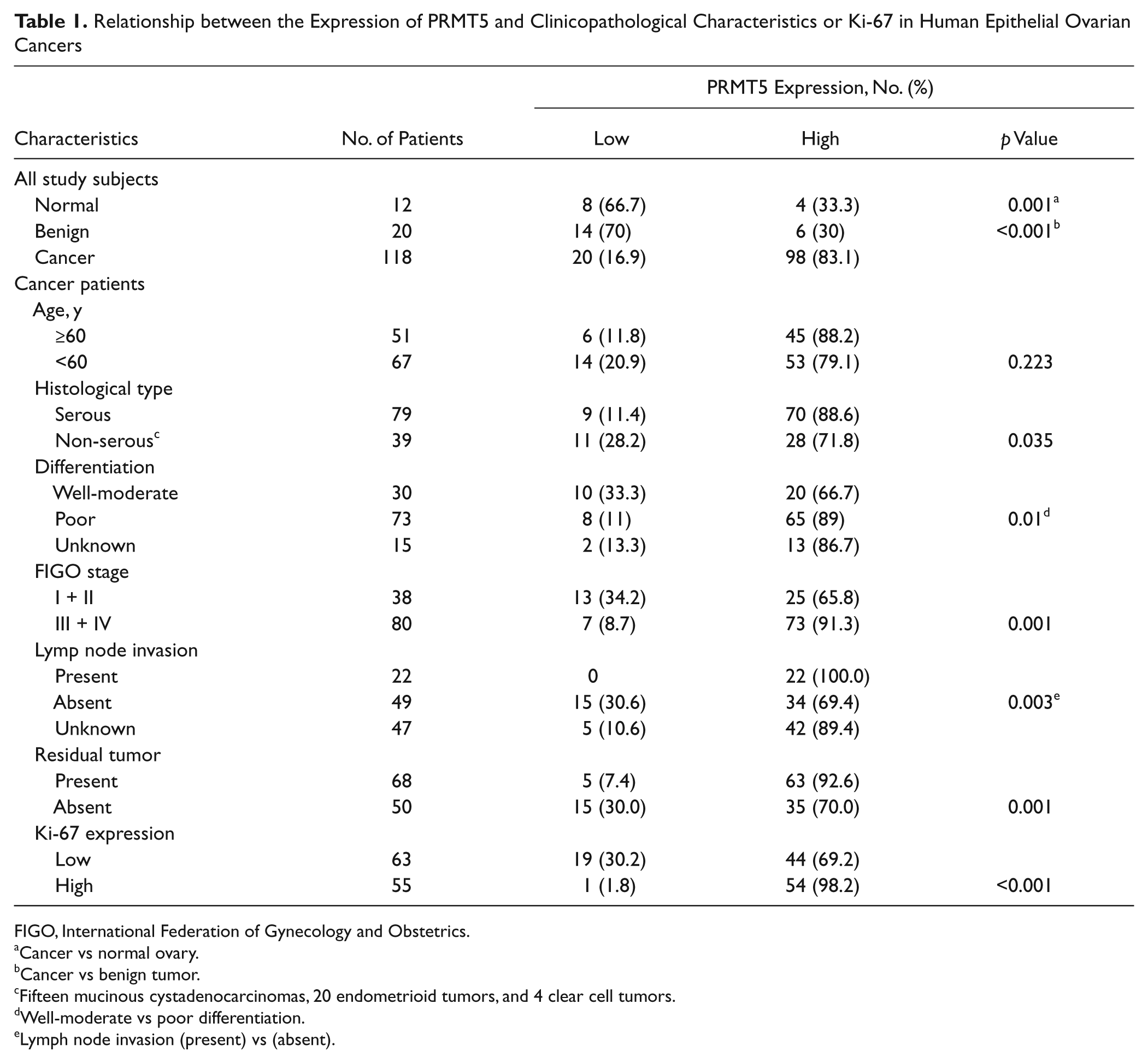
FIGO, International Federation of Gynecology and Obstetrics.
Cancer vs normal ovary.
Cancer vs benign tumor.
Fifteen mucinous cystadenocarcinomas, 20 endometrioid tumors, and 4 clear cell tumors.
Well-moderate vs poor differentiation.
Lymph node invasion (present) vs (absent).
All EOC patients received standard adjuvant chemotherapy based on platinum postoperatively, and the clinical outcomes were available. All patients had been followed up from the date of surgical treatment to January 2012. The follow-up time ranged from 3 to 81 months, with a median of 32 months. Follow-up information included overall survival (OS) and progression-free survival (PFS). OS was calculated from the date of surgery to the date of death or last follow-up, and PFS was from the date of surgery to the date of documented disease progression.
Immunohistochemistry
IHC staining was performed with the standard streptavidin-peroxidase complex method. Sections (4 µm) were stained with anti-PRMT5 (1:200; EIAab Science, Wuhan, China) and anti–Ki-67 (1:100; Dako, Glostrup, Denmark) overnight at 4C, incubated with biotin-labeled secondary antibody and developed with a horseradish peroxidase–labeled streptavidin reagent, and then stained for 3 min with 3,3′-diaminobenzidine (Histostain-plus kit; Zhongshan Golden Bridge Biotechnology, Beijing, China). Sections were lightly counterstained with hematoxylin and mounted with neutral balsam. The primary antibody was replaced with 1 × PBS as a negative control.
Evaluation and Quantification of Immunostaining
Evaluation of staining of PRMT5 and Ki-67 was independently done by two pathologists in a blinded manner. PRMT5 was scored for both nuclear and cytoplasm staining. It was scored semiquantitatively based on a staining intensity of 0 = no staining, 1 = mild, 2 = moderate, and 3 = strong expression, as well as the following distribution: 0 = less than 5%, 1 = 6% to 20%, 2 = 21% to 50%, and 3 = greater than 50% staining. The immunoscore was expressed as a combined score of staining intensity and distribution, with a maximal combined immunoscore of 6. Tumors with a final staining score ≥4 were defined as having a high expression of PRMT5 (Liang et al. 2007).
To measure Ki-67 expression, nuclei from at least 1000 tumor cells were counted from the fields, and the labeling index was calculated as the percentage of labeled nuclei of the total number of tumor cells that were counted. The median was used to dichotomize into low- and high-expressing groups for Ki-67 (Ali-Fehmi et al. 2003).
Cell Culture
Human ovarian cancer cell lines SKOV3 and A2780 were obtained from the Institute for Biological Sciences, Chinese Academy of Sciences, Shanghai. A2780 was grown in RPMI-1640, and SKOV3 was grown in McCoy’s 5A containing 10% fetal bovine serum (FBS) in a humidified atmosphere at 37C with 5% CO2.
siRNA Transfection
The two human PRMT5 siRNA sequences (siRNA-1 [siP1], 5′-GGGACUGGAAUACGCUAAUTT-3′ and 5′- AUUAGCGUAUUCCAGUCCCTT-3′; siRNA-2 [siP2], 5′-GGACCUGAGAGAUGAUAUATT-3′ and 5′-UAU AUCAUCUCUCAGGUCCTT-3′) and the scrambled siRNA sequence (5′-UUCUCCGAACGUGUCACGUTT-3′ and 5′-ACGUGACACGUUCGGAGAATT-3′) were designed and synthesized by Genepharma Co., Ltd (Shanghai, China). Cancer cells were incubated in culture without antibiotics for 24 hr prior to transfection, resulting in 30% to 50% confluence. Then they were transfected with PRMT5 siRNA (siP1 and siP2) or control scrambled siRNA (siC) in Opti-MEM (Invitrogen; Carlsbad, CA) using the Lipofectamime 2000 transfection reagent (Invitrogen) in accordance with the manufacturer’s instructions. The ratio of Lipofectamime 2000 to siRNA was 1:1. Silencing of PRMT5 was assayed for mRNA and protein expression level at 48 hr and 72 hr after transfection, respectively. Each transfection was repeated three times. Untransfected ovarian cancer cells were the normal control.
Quantitative Real-Time PCR
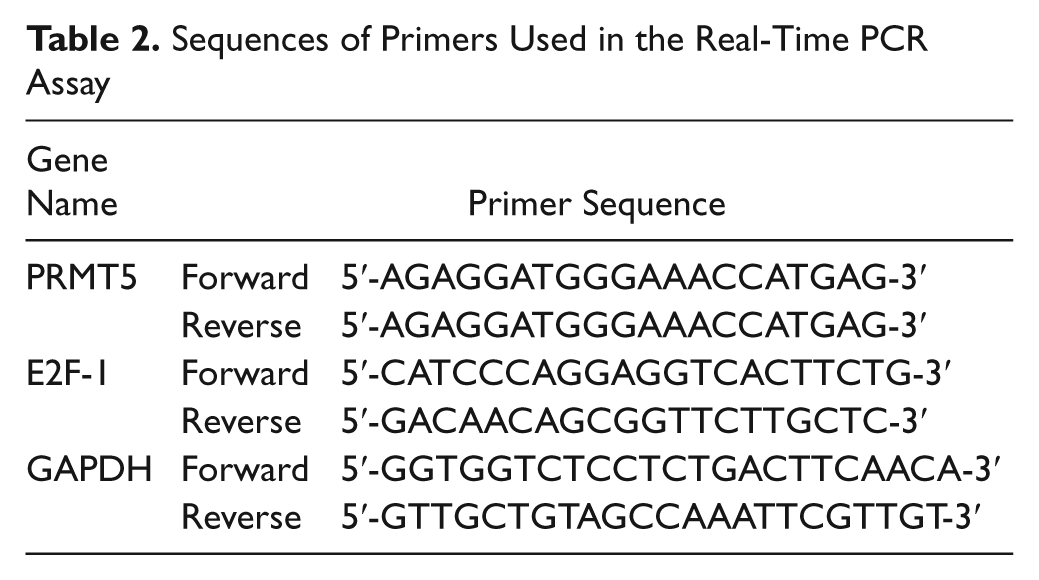
At 48 hr posttransfection, total RNA was extracted using TRIzol reagent (Invitrogen) in a standard procedure. Reverse transcription was carried out using the PrimeScript RT reagent kit (Takara; Dalian, China). Real-time PCR was then performed using SYBR Green Realtime PCR Master Mix (TOYOBO; Osaka, Japan) in the LightCycler 2.0 PCR instrument (Roche Applied Science, Mannheim, Germany) according to the manufacturer’s instructions. The PCR reaction conditions were as follows: denaturation at 95C for 30 sec, followed by 45 cycles of 95C for 5 sec, 10 sec at 58C, and 15 sec at 72C; and a final extension at 65C for 15 sec. All primer sequences (Biosune; Shanghai, China) are listed in Table 2. The relative mRNA level was normalized on the basis of GAPDH and to a normal control. The control was included in each run, and its normalized mRNA level was set at a value of 1.0.
Sequences of Primers Used in the Real-Time PCR Assay
Western Blot Analysis
At 72 hr after transfection, protein lysates were extracted in RIPA buffer according to the manufacturer’s instruction (Bytotime, Jiangsu, China). Denatured protein samples were equally (70 µg) loaded on 10% SDS-PAGE gels and transferred to PVDF membranes using the Bio-Rad (Hercules, CA) electrotransfer system. The membranes were blocked in 5% non-fat milk in Tris-buffered saline containing Tween 20 (TBST; 10 mmol/L Tris-HCl, 150 mmol/L NaCl, 0.1% Tween 20) for 1 hr at room temperature, followed by incubation overnight at 4C with the following primary antibodies: rabbit anti-PRMT5 and anti–E2F-1 (1:1000; Cell Signaling Technology, Danvers, MA) and mouse anti–β-actin (1:1000; Zhongshan Golden Bridge Biotechnology, Beijing, China). HRP-conjugated secondary antibodies (anti-mouse and anti-rabbit [1:4000; Zhongshan Golden Bridge Biotechnology]) were added for 1 hr at room temperature. Detection was performed by enhanced chemiluminescence (Millipore, Billerica, MA) and recorded by Kodak molecular imaging software (Kodak; Rochester, NY). Western blot was performed at least three times for each sample. The band intensity was measured by Image J (National Institutes of Health; Bethesda, MD), and the relative protein expression was normalized to β-actin.
CCK8 Assay
Cells were seeded in a 96-well plate at a density of 3000 cells per well, allowed to adhere overnight, and then subjected to transfection with siC and siP1 or no transfection (normal control). At the indicated time points, 10 µl CCK8 (Jingmei Biotech; Shanghai, China) and 90 µl culture medium were added and then incubated at 37C for 2 hr. The blank well was the control. The absorbance was read at 450 nm using a microplate reader. All assays were performed in triplicate and repeated three times. Cell viability (%) = (absorbance of the transfection wells – absorbance of the blank control wells)/(absorbance of the normal control wells – absorbance of the blank control wells) × 100%.
Proliferation Assay by Labeling Ki-67
Ovarian cancer cells were seeded onto glass coverslips at 30% to 50% confluence overnight and then were transfected with siP1 or siC. After 72 hr, the cells were rinsed in PBS and fixed in cold acetone for 10 min. The cells on the coverslips were permeabilized using PBS buffer with 0.5% Triton X-100. Then a standard immunohistochemistry procedure of streptavidin-peroxidase complex as described previously (Vital-Reyes et al. 2006) was performed to determine the expression of Ki-67 and PRMT5. The IHC assay for each condition and cell line was repeated at least twice. For Ki-67, the percentage of stained nuclei was estimated by counting positive and negative nuclei on 10 random fields for each coverslip undergoing each treatment.
Apoptosis Analysis
At 72 hr after transfection, cells were harvested by trypsinization, washed in cold PBS, and resuspended in binding buffer (HEPES-buffered PBS supplemented with 2.5 mM CaCl2) at a density of 1 × 106 cells/ml. The cells were doubled stained with the Annexin V–FITC and propidium iodide (PI) apoptosis detection kit (Jingmei Biotech), and incubated at room temperature for 15 min in the dark. The cells were then analyzed by flow cytometry with an FACScan flow cytometer (BD, San Jose, CA) using a 488-nm laser line. Data were analyzed using WinMDI 2.8 software (Trotter 2004). The cells positive for Annexin V (AnnexinV+ PI– + AnnexinV+ PI+) were counted as apoptotic cells.
Statistical Analysis
Quantitative data are expressed as the mean ± SD, except where indicated. The significance of the difference between groups was evaluated with the two-tailed Student’s t-test. IHC data were analyzed using the χ2 test or Fisher’s exact test. The Spearman test was used to analyze the relationship between PRMT5 and Ki-67 scores. Univariate Cox regression analysis was used to evaluate the prognostic significance of variables of EOC. Survival probabilities of patients were determined using Kaplan-Meier curves and compared using the log-rank test. Parameters that showed significant prognostic value in the univariate analysis were evaluated by multivariate Cox regression analysis. All calculations were performed using SPSS 13.0 for Windows (SPSS, Inc, an IBM Company, Chicago, IL). A p value <0.05 was considered statistically significant.
Results
PRMT5 Is Highly Expressed in EOC and the Expression Level Is Correlated with the Proliferation Activity
Positive staining for PRMT5 was observed both in the nucleus and cytoplasm, and the staining of Ki-67 was in the nucleus of ovarian tumor cells (Fig. 1). Using the criteria described earlier, high expression of PRMT5 was observed in 4 of 12 (33.3%) normal ovaries, in 6 of 20 (30%) benign tumors, and in 98 of 118 (83.1%) EOCs, respectively. The level of PRMT5 in EOCs was the highest (p<0.05), and there was no significant difference between normal ovaries and benign tumors. In normal and benign tumor cases with a high expression of PRMT5, the expression was mostly localized to the epithelium, and most of the scores for positive tissue staining were 4 to 5 and classified as mild to moderate staining. Interestingly, there was stronger staining of PRMT5 in epithelial inclusion cysts and stromal cells of normal ovaries in some cases (Fig. 1E). The relationship between the expression of PRMT5 in EOCs and the clinicopathological features of the patients is summarized in Table 1. High expression of PRMT5 was significantly associated with serous subtype, advanced FIGO stage, poor differentiation, lymph node invasion, and the presence of residual tumor (p<0.05, respectively).
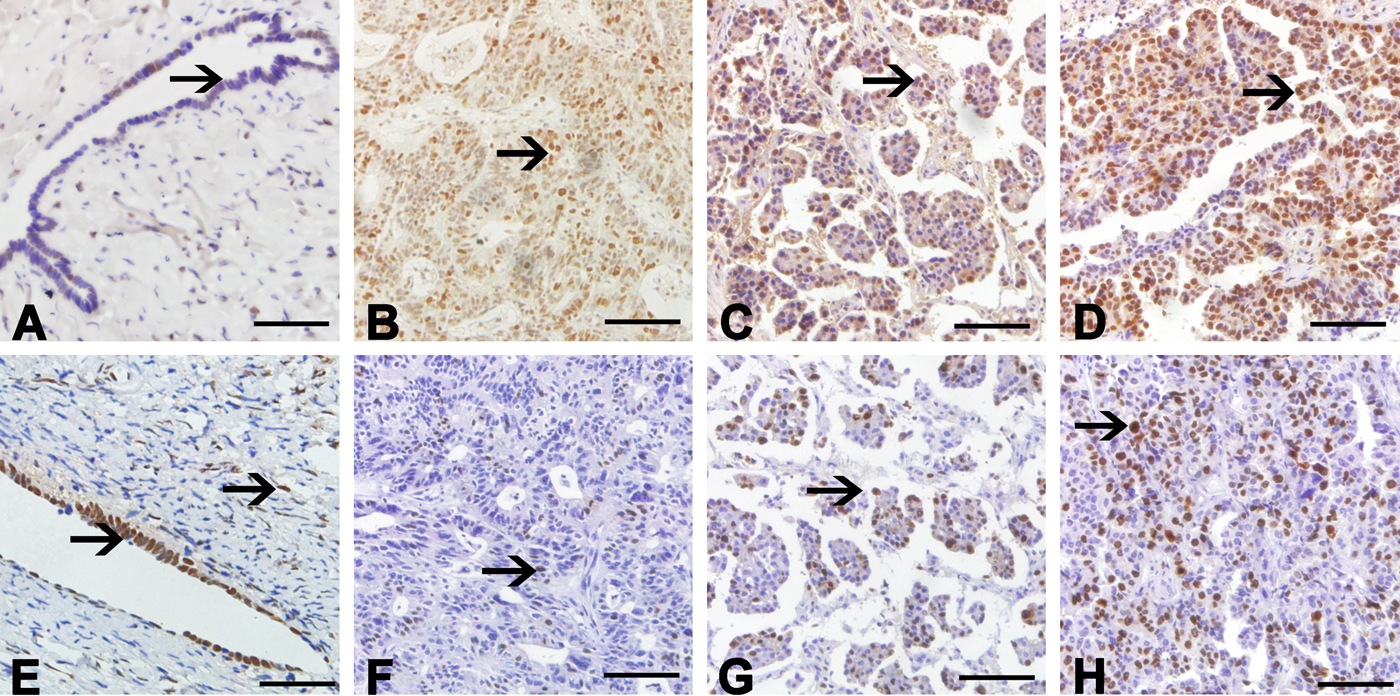
Representative images for PRMT5 and Ki-67 immunohistochemical (IHC) staining. (A) Benign ovarian tumor tissue showed a negative staining of PRMT5 (arrow). (E) Epithelial inclusion cysts and stromal cells showed positive staining of PRMT5 (arrows). (B–D) IHC staining of PRMT5, from left to right, shows mild, moderate, and strong staining. (F–H) Ki-67 staining of the same set of epithelial ovarian cancer specimens. Positively stained cells of PRMT5 and Ki-67 are shown (arrows). Bars: A, E = 50 µm; B–D, F–H = 100 µm.
Because PRMT5 has been implicated in promoting cell proliferation, we stained the same set of EOC specimens with Ki-67, a cell proliferation marker that has been used as a measure of tumor proliferative activity. The median for the Ki-67 labeling index in the entire cohort was 25%, and we used this as a cutoff for the high versus low Ki-67 labeling index. A significant positive correlation was noted between PRMT5 expression in tumor cells and Ki-67 expression (r = 0.377, p<0.001), in which tumors with high Ki-67 expression had a significantly higher expression of PRMT5 (69.2% vs 98.2%, p<0.001). Together, we conclude that PRMT5 is significantly increased in primary EOC, and its expression correlates with a high proliferation index, as revealed by Ki-67 staining.
Level of PRMT5 Is Negatively Correlated with Survival of Ovarian Cancer Patients
At the end of the follow-up period, of the 118 ovarian cancer patients, 63 (53.4%) had died, with 61 patients dying from cancer-related causes and 2 patients from other causes. The median OS time of patients was 40 months (95% confidence interval [CI], 19.1–60.1), and median PFS time was 20 months (95% CI, 16.8–23.2). To evaluate the impact of elevated PRMT5 expression on OS and PFS, we first performed a univariate analysis of traditional clinicopathogical variables for prognosis (Table 3). In the OS analyses, serous subtype, PRMT5 expression, age, FIGO stage, and tumor differentiation, but not other clinicopathological features, were significant predictors of OS. In the PFS analyses, all clinicopathological variables, except lymph node invasion, were significant predictors of PFS (Table 3). Univariate analysis by log-rank test indicated that patients with high PRMT5 expression had a significantly poorer OS and shorter PFS than those with low PRMT5 expression (p<0.001, each; Fig. 2).
Univariate and Multivariable Analyses of Several Clinicopathological Parameters in Relation to Overall Survival and Progression-Free Survival of Patients with Epithelial Ovarian Cancers
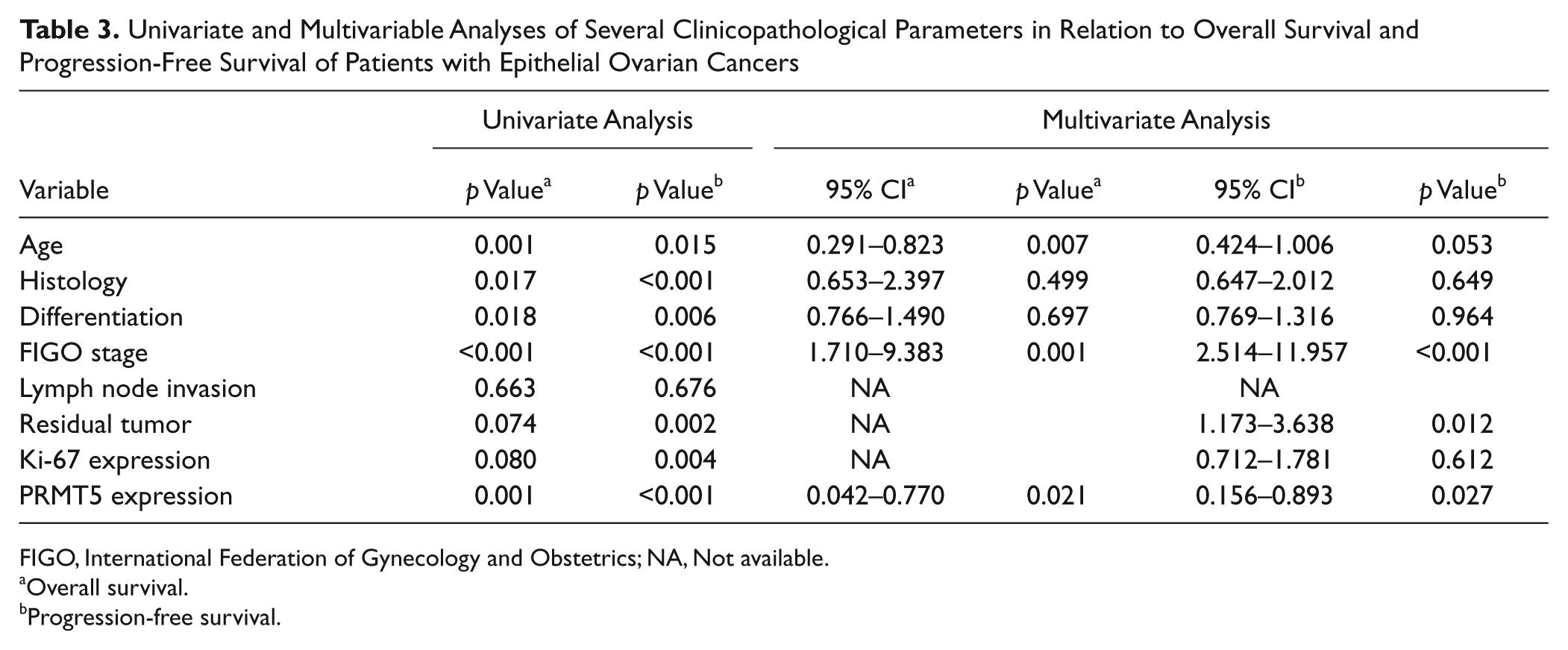
FIGO, International Federation of Gynecology and Obstetrics; NA, Not available.
Overall survival.
Progression-free survival.
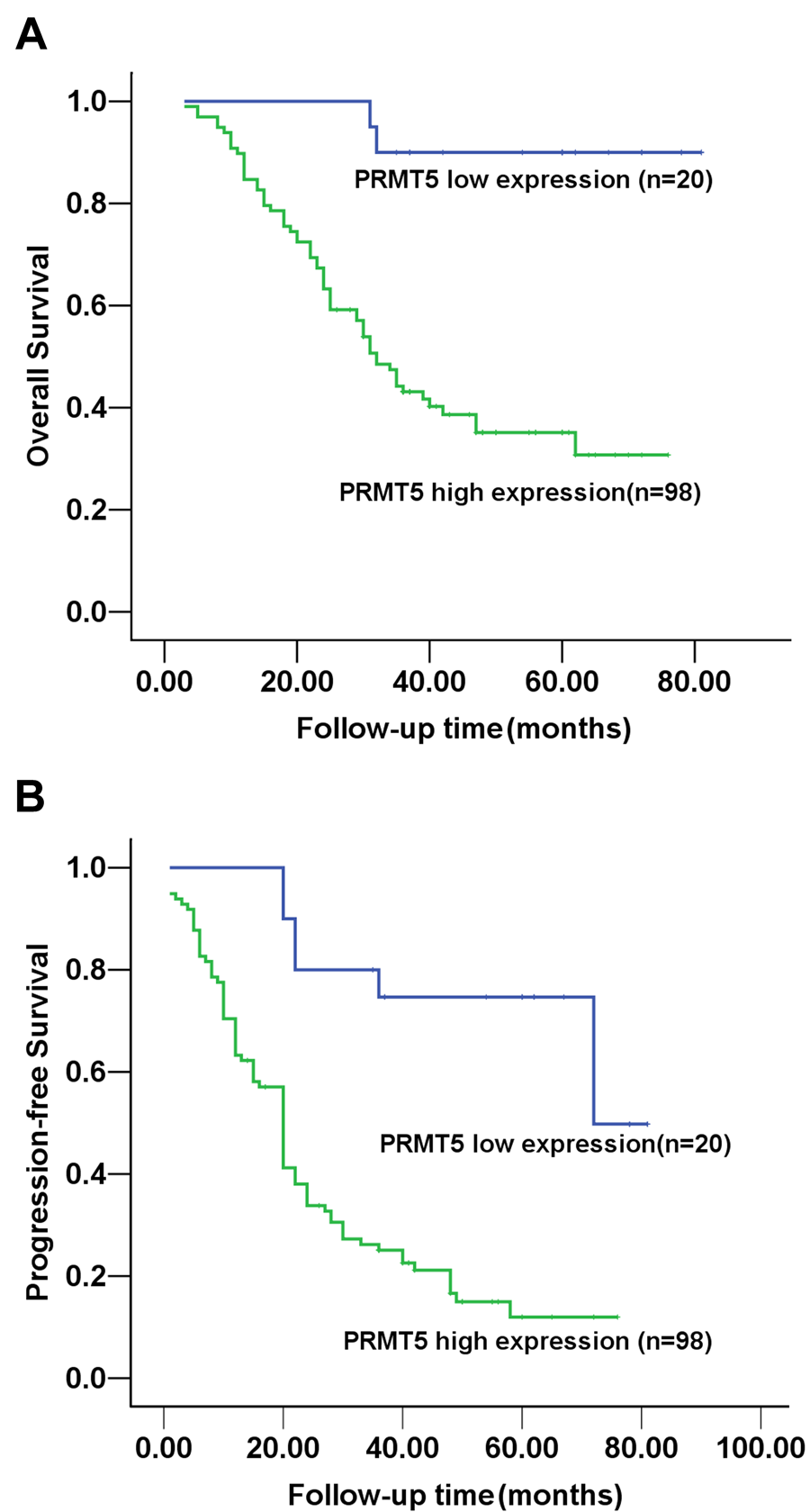
Kaplan-Meier survival curves according to the immunoexpression of PRMT5 protein. (A) Overall survival. (B) Progression-free survival.
To further determine the relationship between survival and clinicopathological features, a multivariate Cox regression model adjusted for statistically significant prognostic factors was performed. The expression of PRMT5 and other clinicopathological features that were significant in the univariate analysis were examined in the multivariate analysis. Only the high expression of PRMT5, older age, and advanced stage retained their significance as independent prognostic factors of poorer OS (Table 3). The results for PFS were similar: the high expression of PRMT5 was found to be an independent prognostic factor for shorter PFS, as were advanced stage and the presence of residual tumor (Table 3).
Knockdown of PRMT5 by siRNA Inhibits the Growth and Proliferation of Ovarian Cancer Cells In Vitro
Because PRMT5 expression positively correlated with Ki-67 expression, we sought to investigate the effects of PRMT5 knockdown on growth and proliferation of human EOC cell lines A2780 and SKOV3. We used PRMT5 siRNA (siP1 and siP2) to transfect ovarian cells, scrambled siRNA (siC) as the control, and untransfected ovarian cancer cells as the normal control. The knockdown efficacy of siRNA was confirmed by real-time PCR at 48 hr and Western blot at 72 hr posttransfection. Compared with normal control or cells transfected with siC, in A2780 cells, downregulation of PRMT5 mRNA (~83%, p<0.001) and protein (~55%, p<0.001) expression was observed in the cells with siP1, but there was no significant change in PRMT5 expression in the cells with siP2 (Fig. 3A–C); in SKOV3 cells, downregulation of PRMT5 mRNA (~90% with siP1, ~75% with siP2, p<0.001, each) and protein (~60% with siP1, ~44% with siP2, p<0.001, each) expression was observed (Fig. 3E–G). According to the results, siP1 was the most efficient and specific sequence to silence the expression of PRMT5 in the two cell lines and was used to finish the subsequent experiments.
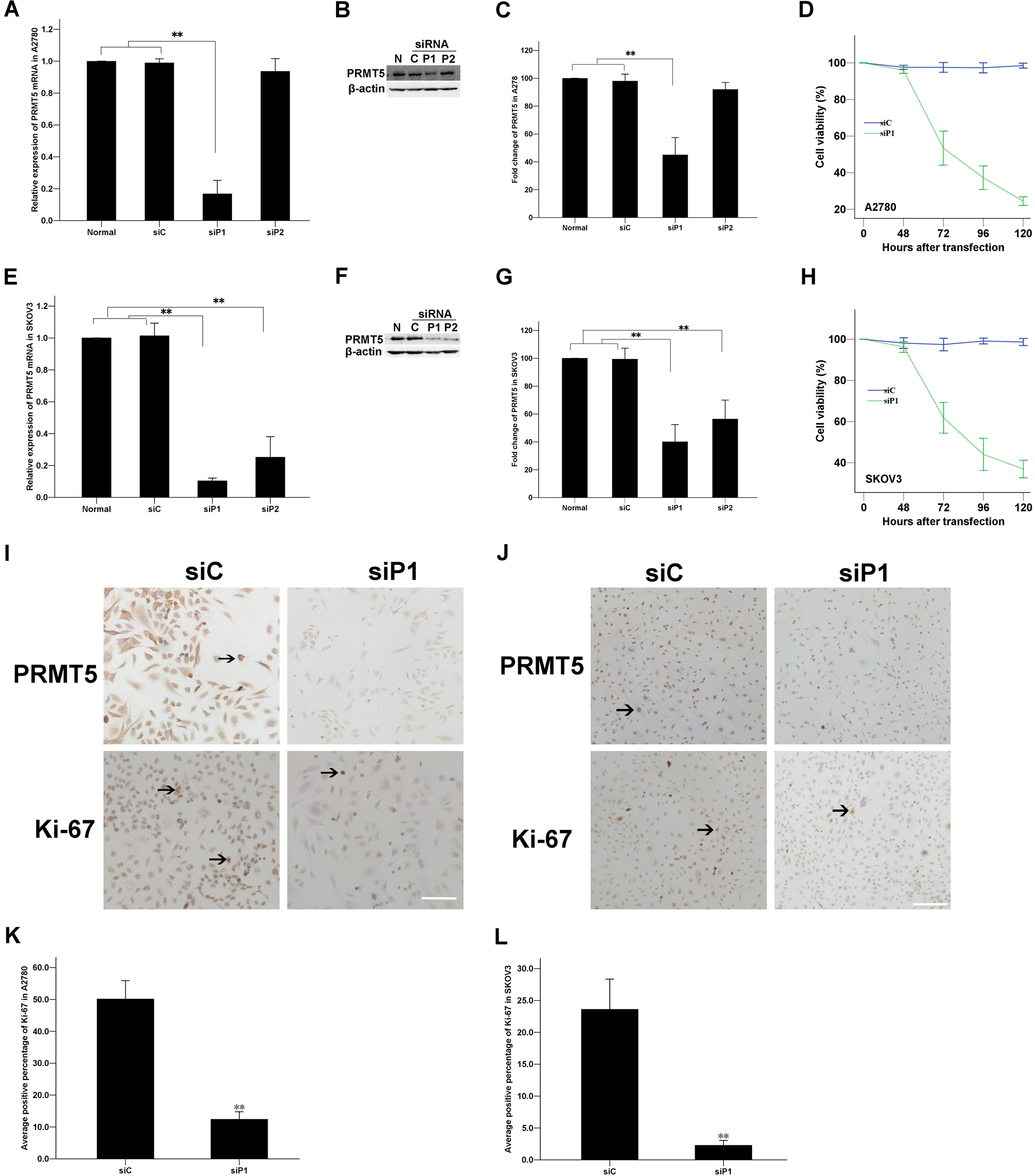
Involvement of PRMT5 in the growth of ovarian cancer cell lines, A2780 and SKOV3. (A–C, E–G) Inhibition of PRMT5 expression in ovarian cancer cells by siRNA. Cells were transfected with PRMT5 siRNA (siP1 and siP2) and control siRNA (siC). Real-time PCR and Western blot revealed a significant reduction in PRMT5 mRNA and protein expression in ovarian cancer cells transfected with siP1 compared with normal control and cells transfected with siC. Results shown are representative of three independent experiments. A–C = A2780 cells; E–G = SKOV3 cells. (D, H) Effects of PRMT5 siRNA on the growth of ovarian cancer cells. At 48, 72, 96, and 120 hr posttransfection with siP1 or siC, cell viability was determined by the CCK8 assay. (I–L) At 72 hr posttransfection (siP1 and siC), cells were labeled with Ki-67 by immunohistochemistry. (I, J) Representative images of PRMT5 and Ki-67 staining of ovarian cancer cells on coverslips. (I) A2780, bar = 100 µm; (J) SKOV3, bar = 200 µm. (K, L) Average percentage of Ki-67–positive nuclei is shown. Data are shown as the mean ± SD; n=3; two-tailed unpaired Student’s t-test; **p<0.001.
To investigate whether downregulation of PRMT5 resulted in the inhibition of cell growth in ovarian cancer cells, we used the CCK8 assay to determine cell viability. At 72 hr after transfection, cell viability started to decrease in the cells transfected with siP1 as compared with the cells transfected with siC or in the normal control (p<0.05; Fig. 3D, H). Moreover, the inhibition of growth was in a time-dependent manner. Similar results were observed in both A2780 and SKOV3 cells lines.
Next, cell proliferation activity was detected by the Ki-67 labeling assay. At 72 hr after transfection, compared with the control siC-transfected cells, cells transfected with siP1 showed a significantly decreased positive percentage of Ki-67 (12.43 ± 3.27% vs 50.16 ± 8.01% in A2780 and 2.29 ± 1.04% vs 23.62 ± 6.61% in SKOV3, respectively) (p<0.001; Fig. 3I–L).
PRMT5 Regulates Ovarian Cancer Cell Growth and Apoptosis Dependent on E2F-1
We next examined whether downregulation of PRMT5 could induce apoptosis in ovarian cancer cells. At 72 hr after transfection, we detected apoptosis induced by transfection with siP1 in A2780 cells stained with Annexin V–FITC and PI and assessed by flow cytometry. The percentage of apoptosis in siP1-transfected A2780 cells was higher than that in the siC-transfected A2780 cells, with 29.37 ± 0.90% vs 16.27 ± 0.84% (p<0.001; Fig. 4A, B). A similar result was observed with SKOV3 cells; the percentage of apoptosis was 28.93 ± 0.60% after siP1 transfection and 14.13 ± 1.36% after siC transfection (p<0.001; Fig. 4C, D).
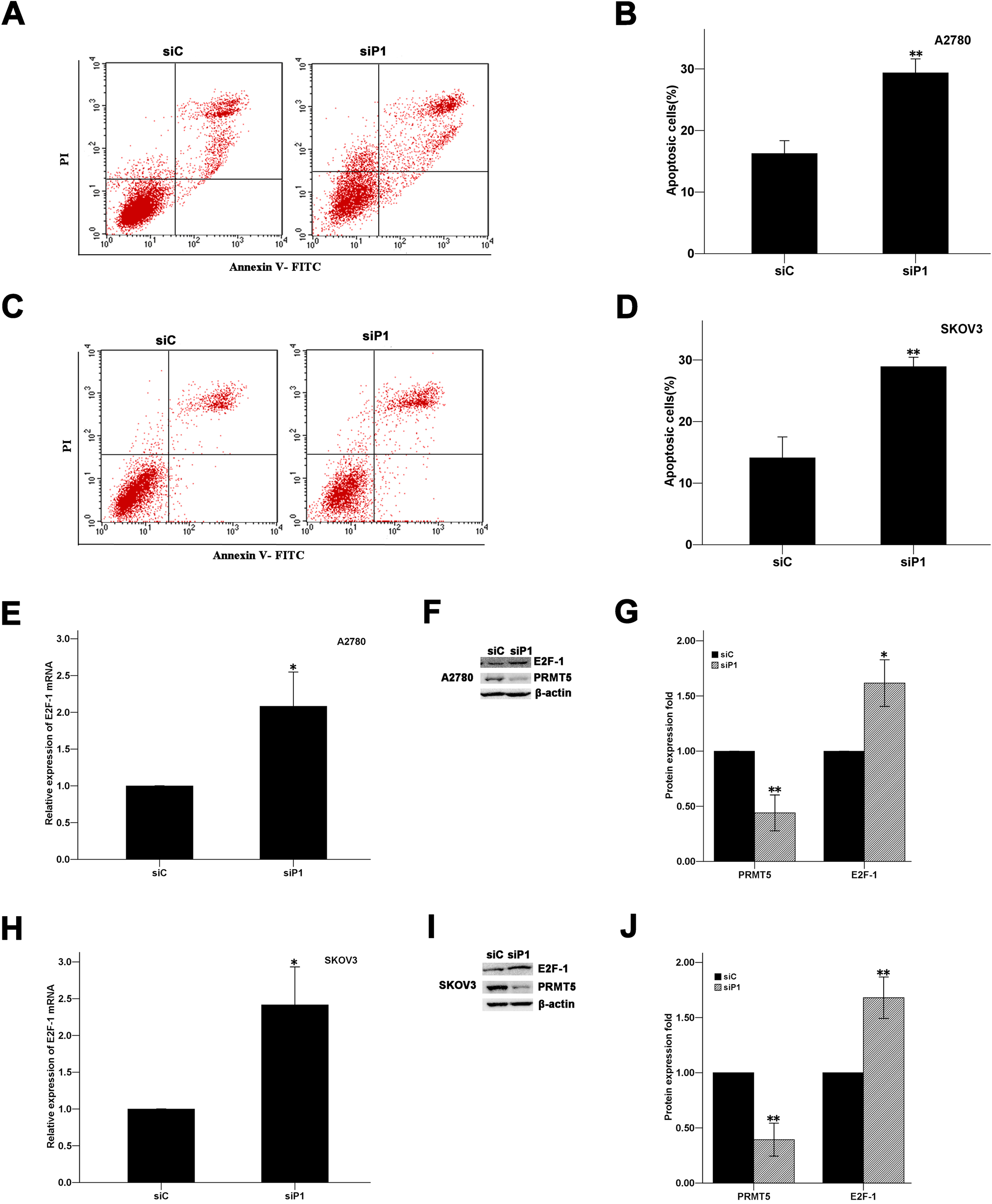
Knockdown of PRMT5 induced apoptosis via an upregulation of E2F-1 in ovarian cancer cell lines. (A–D) Ovarian cancer cells were transfected with siP1 and siC; 48 hr after transfection, cells were harvested and processed for Annexin V–FITC and propidium iodide (PI) staining and analyzed by flow cytometry. (A, C) Representative flow cytometry plots are shown. (B, D) Percentage of apoptotic cells. (E–J) Real-time PCR and Western blot analyses showed that E2F-1 was significantly upregulated following the inhibition of PRMT5 in ovarian cancer cells. A, B, E–G = A2780 cells; C, D, H–J = SKOV3 cells. Data are shown as the mean ± SD; n=3; two-tailed unpaired Student’s t-test; *p<0.05, **p<0.001.
Many genes have been shown to be involved in the regulation of cell growth and apoptosis in cancer cells, such as eIF4E in breast cancer (Scoumanne et al. 2009) and E2F-1 in colorectal cancer (Cho et al. 2012). To determine how PRMT5 regulates cell growth and apoptosis in ovarian carcinoma, we first investigated the mRNA level of E2F-1 using real-time PCR in ovarian cancer cells transfected with siRNA against PRMT5. Our study revealed that knockdown of PRMT5 resulted in a significant upregulation of E2F-1 mRNA in A2780 (2.1-fold, p=0.016) and in SKOV3 (2.4-fold, p=0.009) cells (Fig. 4E, H). To verify whether the alternation of the E2F-1 gene at the level of transcription ultimately results in its altered translation, we conducted Western blotting for the E2F-1 protein. Western blot analysis showed that the protein level of E2F-1 was also upregulated in A2780 (1.6-fold, p=0.006) and SKOV3 (1.7-fold, p<0.001) cells (Fig. 4F,G, I, J). The results showed that ariginine methylation by PRMT5 could regulate cell growth and apoptosis via the upregulation of E2F-1 in EOC.
Discussion
In this study, we measured the expression of PRMT5 in different types of human ovarian tissues. The highest level of PRMT5 was observed in EOC. However, the staining of PRMT5 is significantly stronger in inclusion cysts than in normal ovarian surface epithelium, which are thought to be the precursor lesion of a subset of EOC (Salazar et al. 1996). This suggests that overexpression of PRMT5 is an early event during EOC development. Furthermore, the expression of PRMT5 was increased along with the progression of the disease. These results demonstrate that PRMT5 may play a crucial role as an oncogene in the development of EOC. Different histologies of EOC have distinct pathogenetic profiles (Kobel et al. 2008). Serous ovarian cancer, the most common subtype of EOC, is a predominant advanced-stage carcinoma (Gilks 2004). In our study, the level of PRMT5 expression was higher in the serous subtype than in the non-serous subtype. Because the number of the serous subtype was larger than that of the non-serous subtype, the difference of PRMT5 expression between histological differentiation was uncertain, we should add the number of non-serous subtype cases to verify the difference in the further study.
The cellular localization of PRMT5 has not been identified in different types of cells. In most primary and immortalized cells, PRMT5 is mainly in the cytoplasm, with a small amount in the nucleus, whereas in transformed cells, this distribution is reversed (Pal et al. 2004, 2007). In the nucleus, PRMT5 may promote cell proliferation and transformation by transcriptionally silencing cell cycle controllers and tumor suppressors (Pal et al. 2004). In our study, PRMT5 existed in both the nuclei and cytoplasm, but mainly in the nuclei, of ovarian cancer cells. This result was similar to the study of Cho et al. (2012) and further indicated that PRMT5 was involved in the regulation of ovarian cancer growth and proliferation. Many studies have shown that PRMT5 is involved in the regulation of major signaling pathways that affect cell death and malignant transformation (Scoumanne et al. 2009; Tabata et al. 2009; Yang et al. 2009; Aggarwal et al. 2010; Liu et al. 2011). To verify the function of PRMT5 to regulate the growth of ovarian cancer cells, we analyzed the expression of Ki-67, a proliferation-related nuclear protein. Ki-67 has a low level of expression in resting cells and a high level in proliferating cells and has been routinely used as a measure of tumor proliferative activity. In our study, we also found that the expression of PRMT5 was positively correlated with the expression of Ki-67, in which tumors with a high Ki-67 expression had a significantly higher expression of PRMT5. These findings suggest a possible role for PRMT5 in promoting ovarian cancer cell proliferation and in the progression of ovarian carcinoma.
In the survival analysis, a high level of PRMT5 was identified as an independent factor for poor prognosis in patients with EOC. Recently, PRMT5 was identified as a cofactor that greatly alters the function of programmed cell death protein 4 (PDCD4) in breast cancer, and overexpression of PRMT5 was found to correlate with a worse outcome within a cohort of breast cancer patients whose tumors contained a higher expression of PDCD4 (Powers et al. 2011). Another study also showed that high levels of PRMT5 coincide with low levels of E2F-1 and reflect a poor clinical outcome in colorectal cancers (Cho et al. 2012). These studies showed that PRMT5 may be a good prognostic marker for survival in patients with epithelial ovarian carcinoma.
We have found that PRMT5 was correlated with the expression of Ki-67 in EOC, which indicates that PRMT5 is able to promote cell proliferation in vivo, but the regulatory action of PRMT5 in ovarian cancer cells is not clear in vitro. To verify this idea, we investigated the role of PRMT5 in cell proliferation and apoptosis in EOC cell lines A2780 and SKOV3. Indeed, we found that downregulation of PRMT5 had an effect on growth inhibition of ovarian cancer cells in CCK8 and Ki-67 labeling assays. We also revealed that downregulation of PRMT5 could induce apoptosis in A2780 and SKOV3 cells using flow cytometry.
Multiple genes have been implicated in PRMT5 knockdown-induced growth inhibition and apoptosis (Fabbrizio et al. 2002; Pal et al. 2003, 2004; Wang et al. 2008; Scoumanne et al. 2009; Cho et al. 2012). To further address the mechanism through which PRMT5 regulates growth and apoptosis in EOC, we investigated whether PRMT5 regulates the expression of E2F-1. Our results demonstrated that the depletion of PRMT5 by siRNA could upregulate E2F-1 mRNA and protein expression, both in A2780 and SKOV3 cells. E2F is a complex family of transcription factors implicated in different cells and involved in the regulation of cell proliferation and cell cycle progression (Polager and Ginsberg 2008). E2F-1, as a member of this gene family, is crucial for E2F-dependent apoptotic programming (Lazzerini Denchi and Helin 2005). E2F-1 overexpression also has a growth-promoting effect in EOC cells and is thought to be a useful prognostic indicator (Suh et al. 2008). E2F-1 is directly methylated by PRMT5, and this arginine methylation plays an important role in regulating the biochemical function of E2F-1, which affects E2F-1–dependent growth control. Downregulation of PRMT5 has been shown to increase E2F-1 protein levels in U2OS and HCT116 cells, which caused a decreased growth rate and the induction of apoptosis (Cho et al. 2012). In our study, we also found that the E2F-1 expression coincides with PRMT5 in ovarian cancer cells, suggesting that PRMT5 could regulate growth and apoptosis through E2F-1.
In summary, our results showed that PRMT5 was expressed in ovarian cancers at variable levels, and that expression was related to tumor phenotype and poor prognosis. Our results further provided in vitro evidence in support of the role of PRMT5 as an oncogene in ovarian cancer cells and provided new possible molecular mechanisms for the tumorigenesis of ovarian cancer. Depleting PRMT5 by siRNA was able to substantially inhibit growth and proliferation of ovarian cancer cells and induced apoptosis in vitro; thus, the downregulation of this gene could be a potential therapeutic target in ovarian cancer. More research is needed to elucidate the relevance of PRMT5 and the molecular pathogenesis of ovarian cancer.
Footnotes
Acknowledgements
We thank Prof. Tingguo Zhang, Alei Feng, Qingjie Wang, Qianqian Shao, and Jintang Sun (Qilu Hospital, Shandong University) for their technical guidance. We also thank Dr. Baojin Ding (University of Massachusetts Medical School) for the manuscript initial review and suggestions.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Xingsheng Yang received support by Science and Tech- nology Development Planning of Shandong Province (No. 2011GSF12121).