
Abstract
Endothelial lipase (EL), a member of the triglyceride lipase gene family, has been shown to be a key player in HDL metabolism. Northern blots revealed that EL was highly expressed in endothelium, thyroid, lung, placenta, liver, and testis. In liver and adrenal gland, EL protein was localized with vascular endothelial cells but not parenchymal cells. EL was shown to be upregulated in tissues such as atherosclerotic plaque where it was located in macrophages, endothelial cells, and medial smooth muscle cells. The purpose of this study was to investigate the cellular localization of EL in thyroid and other tissues where EL is known to be expressed. Besides its presence in vascular endothelial and smooth muscle cells, EL protein was detected in the epithelial cells that line the follicles within the thyroid gland. EL-specific immunostaining was also found near the cell surface as well as in the cytoplasm of adipocytes. Using immunoblots, EL expression was confirmed in cultured human omental and subcutaneous adipocytes. EL expression, however, was not found in preadipocytes. These findings suggest that EL plays a role in thyroid and adipocyte biology in addition to its well-known role in endothelial function and HDL metabolism.
Endothelial lipase (EL) is a member of the triglyceride lipase gene family that includes hepatic lipase (HL) and lipoprotein lipase (LPL). The most well-defined role for EL is in HDL metabolism (Brown et al. 2009; DeSantis et al. 2008; Edmondson et al. 2011; Huang et al. 2010; Yasuda et al. 2010; Zhang et al. 2012). High plasma levels of EL have been shown to correlate with low plasma levels of HDL, and human genome–wide association studies have shown several EL polymorphisms that are highly associated with plasma HDL cholesterol (HDL-C) concentrations (Bauer et al. 2011; Edmondson et al. 2011; Edmondson et al. 2009). EL mRNA has been detected in endothelial cells as well as in the thyroid, lung, placenta, liver, and testis (Hirata et al. 1999; Ishida et al. 2003; Jaye et al. 1999). EL mRNA and protein expression has also been noted in brain capillary endothelial cells composing the blood-brain barrier (Sovic et al. 2005). In the liver and adrenal gland, EL protein was colocalized with vascular endothelial cells but not parenchymal cells (Yu et al. 2004). In fact, EL appears to be abundantly expressed in tissues that have elevated metabolic rates and are highly vascularized (Choi et al. 2002). EL has been shown to be upregulated in tissues under pathological conditions, such as inflammation, where it may facilitate delivery of fatty acids to maintain an elevated metabolic state. Unfortunately, under some circumstances, high levels of EL may in fact contribute to pathologies associated with chronic inflammation.
Several observations have led to a fairly strong case that EL plays a role in atherosclerosis and cardiovascular disease in rodents and humans (Brown and Rader 2007; DeSantis et al. 2008; Huang et al. 2010; Ishida et al. 2004a; Riederer et al. 2011; Riederer et al. 2012; Yasuda et al. 2010). EL mRNA expression was increased fourfold in HUVECs and HCAECs following exposure to TNFα and IL-1β and in the lung epithelium of lipopolysaccharide-treated mice (Hirata et al. 2000). Expression of EL message was also augmented in the heart and aorta of rats exposed to angiotension II or shear stress caused by elevated blood pressure (Shimokawa et al. 2005). Similar to LPL, EL on the surface of the endothelium has been shown to induce macrophage activation and monocyte adhesion (Kojma et al. 2004; Li and Renier 2007). Furthermore, atherosclerotic plaques contain elevated EL mRNA and protein expression, and EL was located in the plaque macrophages (foam cells), as well as the endothelial and medial smooth muscle cells (Azumi et al. 2003; Bartels et al. 2007; Brown and Rader 2007; Wu et al. 2009). Serum EL levels were significantly higher, and HDL levels lower, in dialysis patients with low serum albumin and/or high sensitive C-Reactive Protein (hsCRP) levels (Fujii et al. 2008). Therefore, the authors suggested that EL may play an important role in atherogenesis in dialysis patients.
Besides vascular pathology, overexpression of EL has been linked to other disease states, including asthma, diabetes, and stroke. EL protein was found to be overexpressed in epithelial cells, alveolar type II cells, and endothelial cells in the lung during inflammation (Otera et al. 2009). Inactivation of EL in mice was associated with an increase in plasma HDL and an attenuation of allergic inflammation in the lung, leading the authors to conclude that EL may modulate the progression of allergic asthma (Otera et al. 2009). Metabolic inflammation, as measured by high leptin levels and increased TNFα concentrations at the fetal– placental interface, was associated with an upregulation of placental EL mRNA expression in obese women with gestational diabetes mellitus (Gauster et al. 2011). EL protein expression was enhanced in podocytes 1 to 2 weeks after high glucose exposure and glomeruli from diabetic nephropathy patients (Jain et al. 2010). These results provide evidence that EL may potentiate hyperglycemic podocyte stress and possibly the pathogenesis of diabetic glomerulopathy in humans. Low-level EL mRNA expression was reported in CA3 pyramidal neurons of the hippocampus, ependymal cells in the ventral part of the third ventricle, and some cortical cell layers in mice (Paradis et al. 2004), and expression increased following kainate-induced neurodegeneration or occlusion of the middle cerebral artery. Moreover, high plasma levels of EL were correlated with central nervous system injury (e.g., stroke) (Paradis et al. 2004). Despite the fact that EL expression may normally be protective, overexpression of EL may lead to an exacerbation of the pathological response.
Thus, given the growing body of evidence for the expression and potential function of EL outside the vasculature in both normal and pathologic states, we used immunohistochemistry to investigate the localization of EL in several tissues. Besides its well-characterized expression in vascular smooth muscle and endothelial cells that line blood vessels, the data revealed that EL was expressed in the epithelial cells that line the follicles within the thyroid. EL was also expressed near the cell surface as well as in the cytoplasm of adipocytes. Using immunoblots, EL expression was confirmed in cultured human omental and subcutaneous adipocytes but not in preadipocytes. These findings provide further evidence that EL, in addition to its well-known role in endothelial function and HDL metabolism, may play an additional, more direct role in thyroid and adipocyte biology.
Materials and Methods
Reagents
Four polyclonal antibodies were used in this study: EL19–32 antibody directed against amino acids 19–32 from human EL (Cayman Chemicals, Ann Arbor, MI); EL442–456 antibody directed against mouse amino acids 442–456; EL1–500 antibody directed against purified full-length, flag-tagged mouse EL protein; and ELAAV antibody directed against human EL expressed in rabbits via AAV viral infection (generously provided by Dr. Daniel J. Rader). EL442–456 and EL1–500 antibodies were produced in rabbits and affinity purified by New England Peptide (Gardner, MA). Both of these antibodies were found to cross-react with mouse and human EL but not human HL or LPL on immunoblots as well as in immunohistochemistry (IHC) experiments (data not shown) using HEK293 cells stably expressing either mouse EL or human EL, HL or LPL (Darrow et al. 2010). Notably, EL1–500 showed higher cross-reactivity and darker staining with mouse EL than human. Human thyroid tissue lysate was purchased from ProSci, Inc. (Poway, CA). Human-cultured subcutaneous preadipocyte and adipocyte total cell extracts, as well as extracts from omental adipocytes cultured from both normal and diabetic patients, were purchased from Zen-Bio, Inc. (Research Triangle Park, NC).
Immunohistochemistry
Tissues for IHC were routinely fixed in 10% neutral buffered formalin, transferred into phosphate-buffered saline (PBS), and then routinely processed and embedded in paraffin for sectioning (5 µm) onto SuperFrost Plus microscopic slides. Slides were deparaffinized, hydrated, and processed for routine IHC as previously described (D’Andrea et al. 2005; D’Andrea et al. 2000). Briefly, slides were microwaved in Target buffer (Dako, Carpinteria, CA), cooled, placed in distilled H2O, and then treated with 3.0% H2O2 for 10 min. Afterward, the slides were rinsed in PBS (pH 7.4) and then placed in PBS. All subsequent reagent incubations and washes were performed at room temperature. Normal blocking serum (Vector Laboratories, Burlingame, CA) was placed on all slides for 10 min. After briefly rinsing in PBS, primary antibodies (EL19–32 1: 250, EL442–456 1:400, EL1–500 1:25 for human tissues or EL1–500 1:100 for mouse tissues) were placed on slides for 30 min. The slides were washed and biotinylated secondary antibodies, goat anti-rabbit (polyclonal antibodies) were placed on the tissue sections for 30 min (Vector Laboratories). After rinsing in PBS, the horseradish peroxidase conjugated avidin-biotin complex reagent (HRP-ABC, Vector Laboratories) was added for 30 min. Slides were washed and treated with the chromogen DAB (Dako) twice for 5 min each, then rinsed in deionized H2O and counterstained with hematoxylin. A monoclonal antibody to vimentin, (V6630, Sigma, St. Louis, MO) the widely conserved ubiquitous, intracellular filament protein, was utilized as a positive control to demonstrate tissue antigenicity and control reagent quality. The negative controls included replacement of the primary antibody with preimmune serum.
Immunoblot Analysis
Equal amounts of tissue lysates (20-µg protein) were separated on 8% SDS-PAGE gels and blotted onto iBlot gel transfer nitrocellulose membranes (Invitrogen, Life Technologies Corp., Grand Island, NY). Proteins were detected using either 1:200 dilution of EL19–32 primary antibody or 1:1000 dilution of ELAAV primary antibody followed by 1:5000 dilution horseradish peroxidase–conjugated anti-rabbit secondary antibody (Santa Cruz Biotechnology, Santa Cruz, CA) and enhanced chemiluminesence (ECL) Western blotting substrate (Pierce, Thermo Fisher Scientific, Rockford, IL).
Results
EL Localization in Human Placenta, Prostate, Testes, Spleen, Brain, Skeletal Muscle
Because EL has been shown to be expressed in placental tissue via Northern blots, immunoblots, and IHC, human placental samples were used as positive controls for EL protein expression using three different antibodies directed against EL (Lindegaard et al. 2006; Lindegaard et al. 2005). Similar to previous reports of EL protein localization in placental tissue, all three EL-specific antibodies (EL442–456, EL19–32, and EL1–500) immunolabeled the syncytiotrophoblasts and endothelial cells as evidenced by brown punctate staining (black arrows, Fig. 1A–C). In the prostate gland (Fig. 2A, B) and seminal vesicles of the testes (Fig. 2C), EL immunostaining was observed in the smooth muscle cells (black arrows). Little to no EL immunoreactivity, however, was detected in the epithelial cells in these tissues (open arrows, Fig. 2A–C). In the spleen, EL-specific immunolabeling was localized to vascular smooth muscle and endothelial cells (black arrows, Fig. 2D) but not to the leukocytes (open arrow, Fig. 2D). In the brain, the majority of the immunostaining was again localized to vascular smooth muscle and endothelial cells (black arrow, Fig. 2E), and little to no staining was noted in the neurons (open arrow, Fig. 2E). Similarly, in the skeletal muscle, the majority of EL immunostaining was localized to vascular smooth muscle and endothelial cells of the arterioles (top arrow, Fig. 2F) and venules (bottom arrow, Fig. 2F), and little to no staining was observed in the myocytes (open arrow, Fig. 2F). Other than some differential labeling in the medial smooth muscle cells within the various tissues studied, the labeling patterns were generally similar among the three EL antibodies. In addition, the expression patterns were consistent with previously published data depicting EL localization. Note that we did not observe any detectable brown immunolabeling in the negative controls (data not shown). Also, vimentin-positive immunolabeling was observed in all tissues assayed (data not shown).
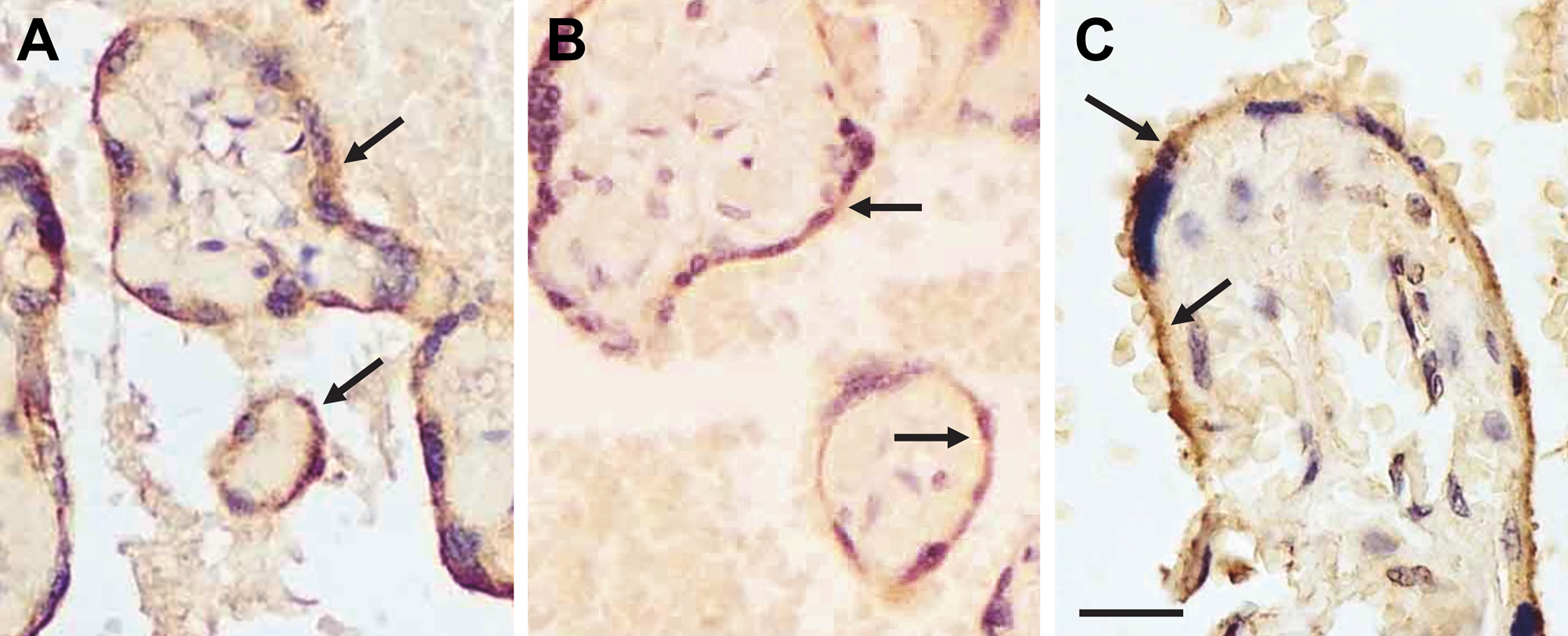
Endothelial lipase (EL) protein expression was localized to syncytiotrophoblasts and endothelial cells in the placenta as evidenced by dark brown punctate staining with anti-EL antibodies: (A) EL442–456, (B) EL19–32, and (C) EL1–500. EL immunostaining is indicated by black arrows. Bar = 50 µm (A–C).
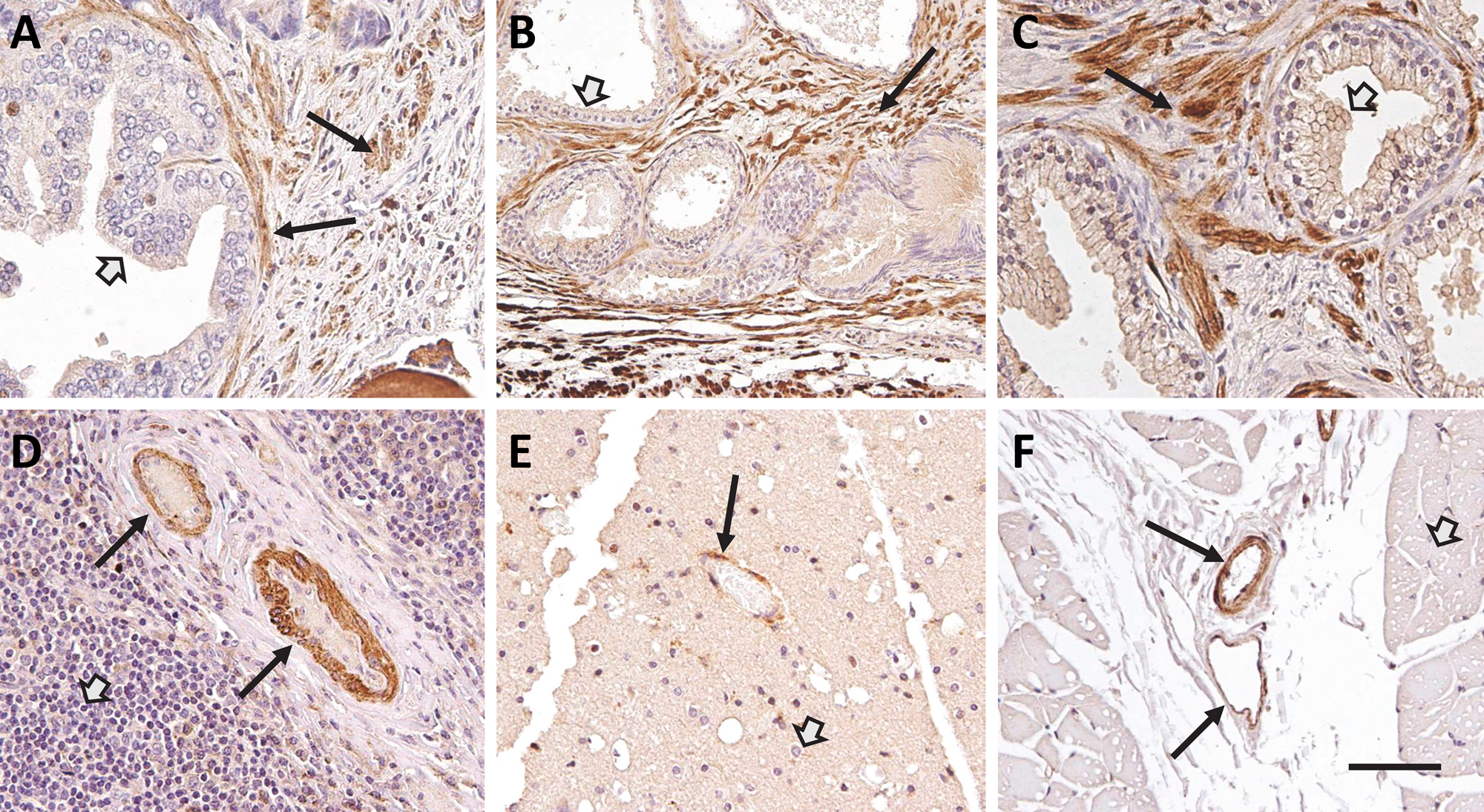
Endothelial lipase (EL) protein was localized to endothelial and smooth muscle cells in human prostate (A and B), testes (C), spleen (D), brain (E), and skeletal muscle (F). EL protein expression was revealed by brown punctate staining with anti-EL antibodies: (A and B) EL442–456 showed EL expression in smooth muscle cells (black arrows) but not the glandular epithelium in the prostate gland (open arrow); (C) EL442–456 showed EL expression in smooth muscle cells (black arrow) but not the epithelial lining (open arrow) of the seminal vesicles; (D) EL442–456 showed EL expression in endothelial and vascular smooth muscle cells (black arrows) of the blood vessels but not the immunocytes (open arrow) within the spleen; (E) EL19–32 showed EL expression in endothelial cells (black arrow) but not the neurons (open arrow) in the brain; and (F) EL1–500 showed EL expression in the endothelial and smooth muscle cells of arterioles (black arrow, top vessel) and venules (black arrow, bottom vessel) but not the myocytes (open arrow) in skeletal muscle. Bar = 100 µm (A, C, D), 200 µm (B, E, F).
EL Localization in Human Thyroid Gland
The thyroid gland comprises numerous follicles (Fig. 3). The lumens of these follicles are filled with colloid, an acidophilic substance that consists of thyroglobulin, the storage form of the thyroid hormones. The most prominent cell types of the thyroid follicle are the epithelial cells that encase the colloid material. The function of the epithelial follicular cells is to synthesize and secrete the iodine containing hormones T3 and T4. The follicles of actively secreting thyroid glands tend to be small with little colloid, and the follicular epithelial cells that line each follicle are tall and cuboidal, reflecting active hormone synthesis and secretion. Conversely, the follicles of less active thyroid glands are larger, filled with colloid and lined with epithelial cells that appear flattened against the follicular basement membrane.
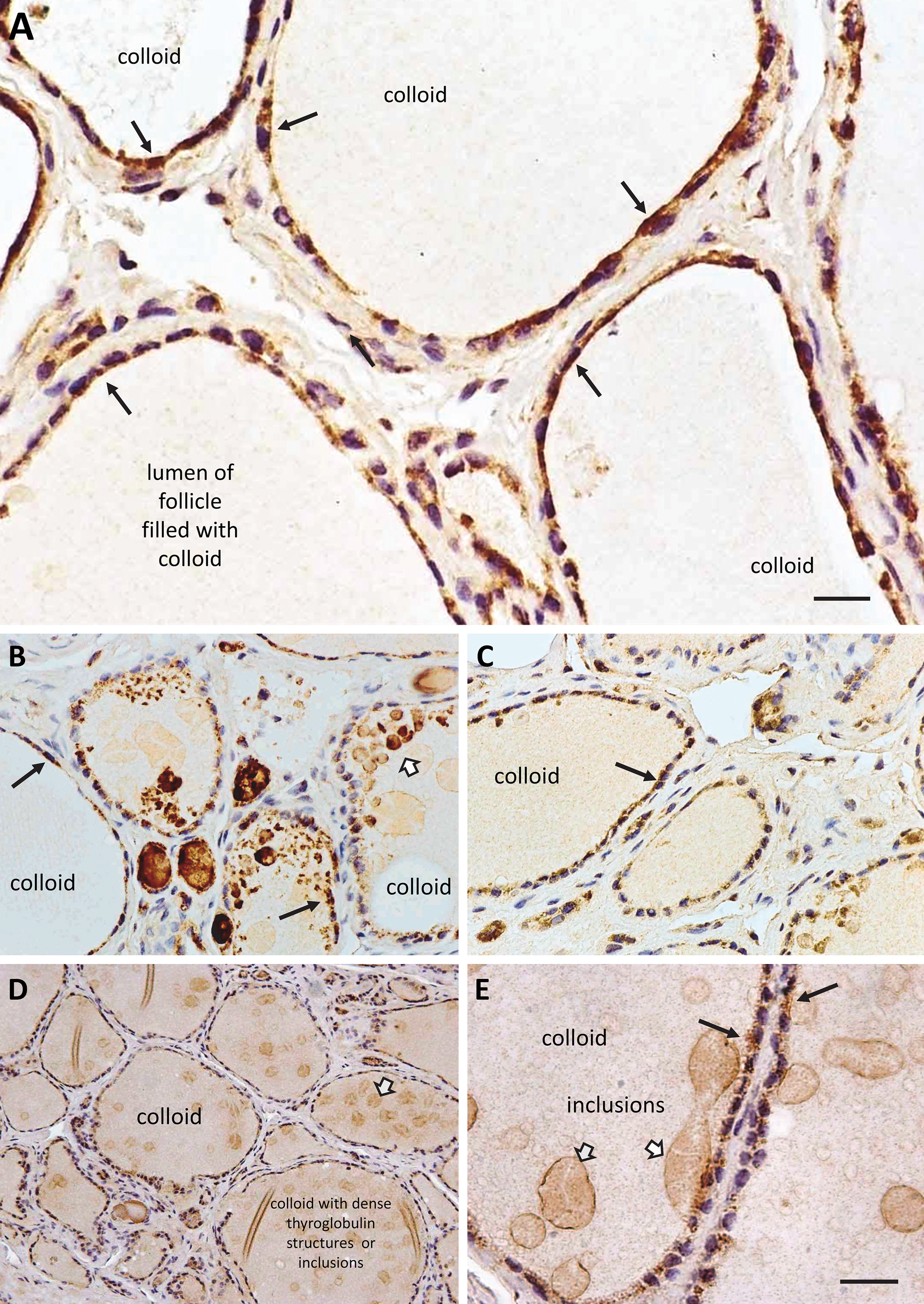
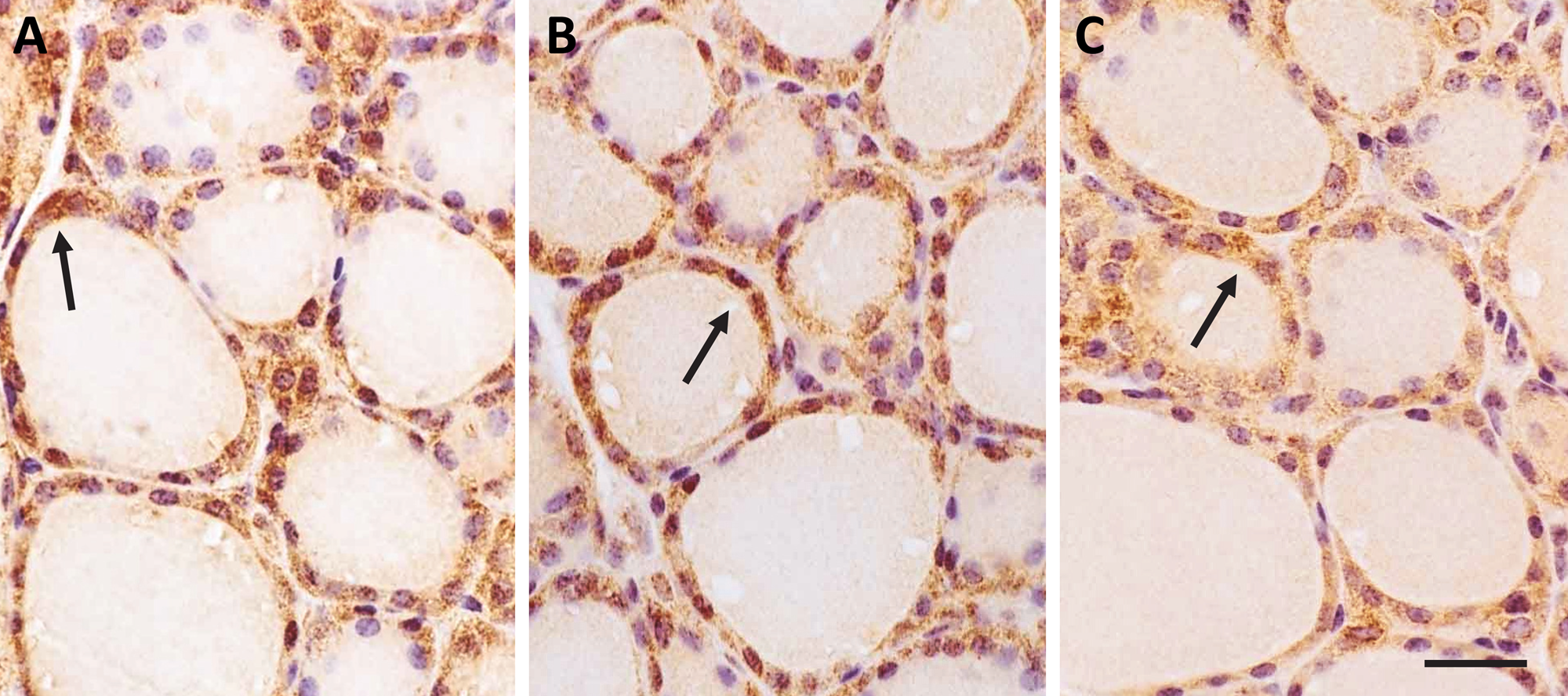
Distribution of endothelial lipase (EL) protein in the follicles of the human thyroid gland. EL protein expression was visualized by dark brown punctate staining (black arrows) in the cuboidal epithelial cells that line the lumen of each follicle within the thyroid. (A) EL442–456 antibody, (B) EL19–32 antibody, (C) EL1–500 antibody, (D) EL19–32 antibody (magnification ×200), and (E) EL19–32 antibody (magnification ×1000). Many of the follicles within the thyroid gland were filled with colloid, some showing inclusions (open arrows), and the follicular epithelium often appeared flat, presumably due to low glandular activity. Bar = 25 µm (A), 50 µm (B, C), 100 µm (D), 25 µm (E).
There was moderate to strong, punctate EL immunolabeling in the follicular epithelial cells that was observed with all three EL antibodies (EL442–456, EL19–32, and EL1–500) (black arrows, Fig. 3A–D). Higher magnification indicated that the cytoplasmic labeling, in many of the epithelial cells, tended to be denser at the apical (luminal) side than the basal side (black arrows, top right Fig. 3A). In addition, there was a periodicity to the labeling pattern along many of the cells that was interrupted by the nuclei, as the thickness of these cells is not much thicker than that of the nuclei in many cases. Thus, the labeling is located close to the nuclei on either side (lower left arrow, Fig. 3A, and black arrows, Fig. 3D), areas where the Golgi apparatus is often prominent, suggesting active EL synthesis in these cells and not just cell surface attachment. Of particular note was the fact that even within the same follicle, not all follicular epithelial cells appeared to express the same levels of EL: some with high levels, some with low, and even a few without detectable EL (Fig. 3A–E). Generally, there were many more follicular epithelial cells with EL immunolabeling than without. Moreover, many follicles within the thyroid gland were filled with colloid, some showing inclusions or “globules” comprising dense thyroglobulin (open arrows, Fig. 3B, D). These types of inclusions are observed in follicles of older humans and rodents, and are often indicative of decreased thyroid activity and a reduced ability of the follicular epithelial cells to reabsorb old and resecrete newly synthesized thyroglobulin (Gerard et al. 2004; Salabe 2001). The fact that the follicular epithelial cells are flat in appearance and the follicles are filled with colloid also suggests that the thyroid samples were from older individuals with somewhat inactive thyroid follicles. Given that these inclusions are typically made up of thyroglobulin, the apparent EL immunostaining in these inclusions is probably an artifact due to entrapment of the antibodies in the regions of dense, almost insoluble protein material in spite of the fact that we did not observe immunolabeling in our negative controls.
The thyroid gland is highly vascularized, and it was not unexpected to find EL protein immunostaining in the endothelial and smooth muscle cells of the blood vessels. All three antibodies immunolabeled the endothelial cells moderately (black arrows, Fig. 4A–D); however, there was differential EL labeling in the surrounding smooth muscle cells. Whereas EL442–456 labeled the medial smooth muscle cells moderately to intensely (open arrow, Fig. 4A), the other two antibodies, EL19–32 and EL1–500, produced much lighter smooth muscle staining (open arrows, Fig. 4B–D) within the thyroid. The reason for this particular difference between the antibody staining is not clear.
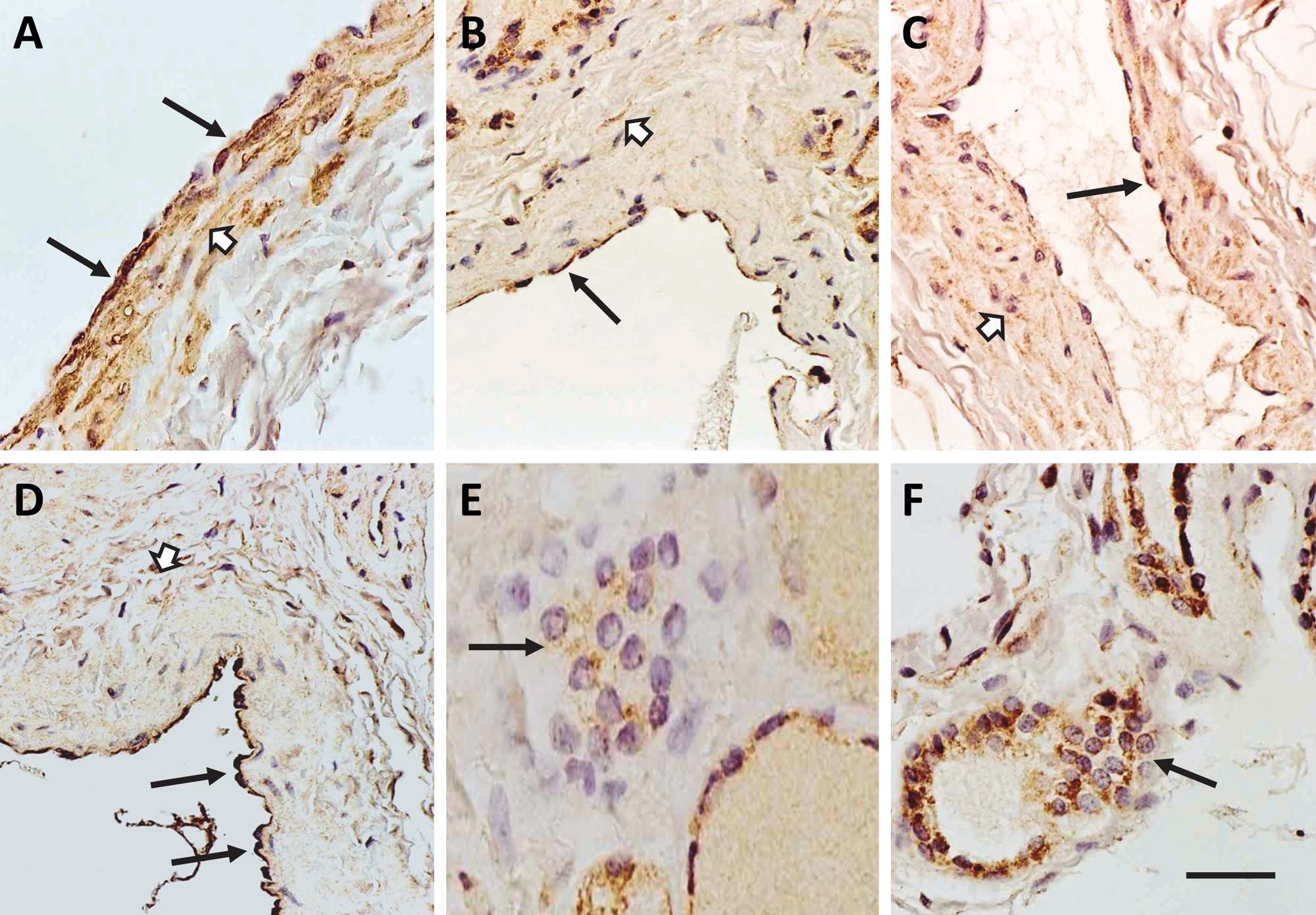
Endothelial lipase (EL) immunoreactivity was localized in the blood vessels but not the parafollicular cells of the human thyroid. EL protein, indicated by brown punctate staining, was found to be expressed in the endothelial (black arrows) and smooth muscle cells (open arrows) of the vasculature within the thyroid gland using: (A) EL442–456 antibody, (B, C) EL19–32 antibody, and (D) EL1–500 antibody. However, little to no EL-specific immunostaining was noted within the parafollicular cells (black arrows) within the thyroid gland using (E) EL442–456 antibody and (F) EL19–32 antibody. Bar = 100 µm (A–D, F), 50 µm (E).
The parafollicular cell is another cell type in the thyroid that resides outside of the follicles. These cells are known to synthesize and secrete the hormone calcitonin, which lowers blood calcium levels in the body by reducing the number and activity of osteoblasts in bones, thereby lowering bone resorption and calcium release. They appear as single cells or as clumps on the periphery of the follicles. The parafollicular cells tended to express minimal to no levels of EL protein (black arrows, Fig. 4E, F). This relative expression is consistent with EL’s primary role in the follicular epithelial cells as compared with the parafollicular cells.
EL Localization in Human Adipose Tissue
Fortuitously, there was adipose tissue present in the thyroid gland tissue samples. While the localization in the thyroid was analyzed, it was noted that all three antibodies (EL442–456, EL19–32, and EL1–500) presented moderate EL labeling in the adipocytes (black arrows, Fig. 5A–C). The labeling appeared to be diffuse throughout the minimal amount of cytoplasm found in adipocytes. There were some areas of the adipose tissue that had little to no immunostaining. However, the majority of adipocytes exhibited brown labeling indicating EL immunoreactivity.
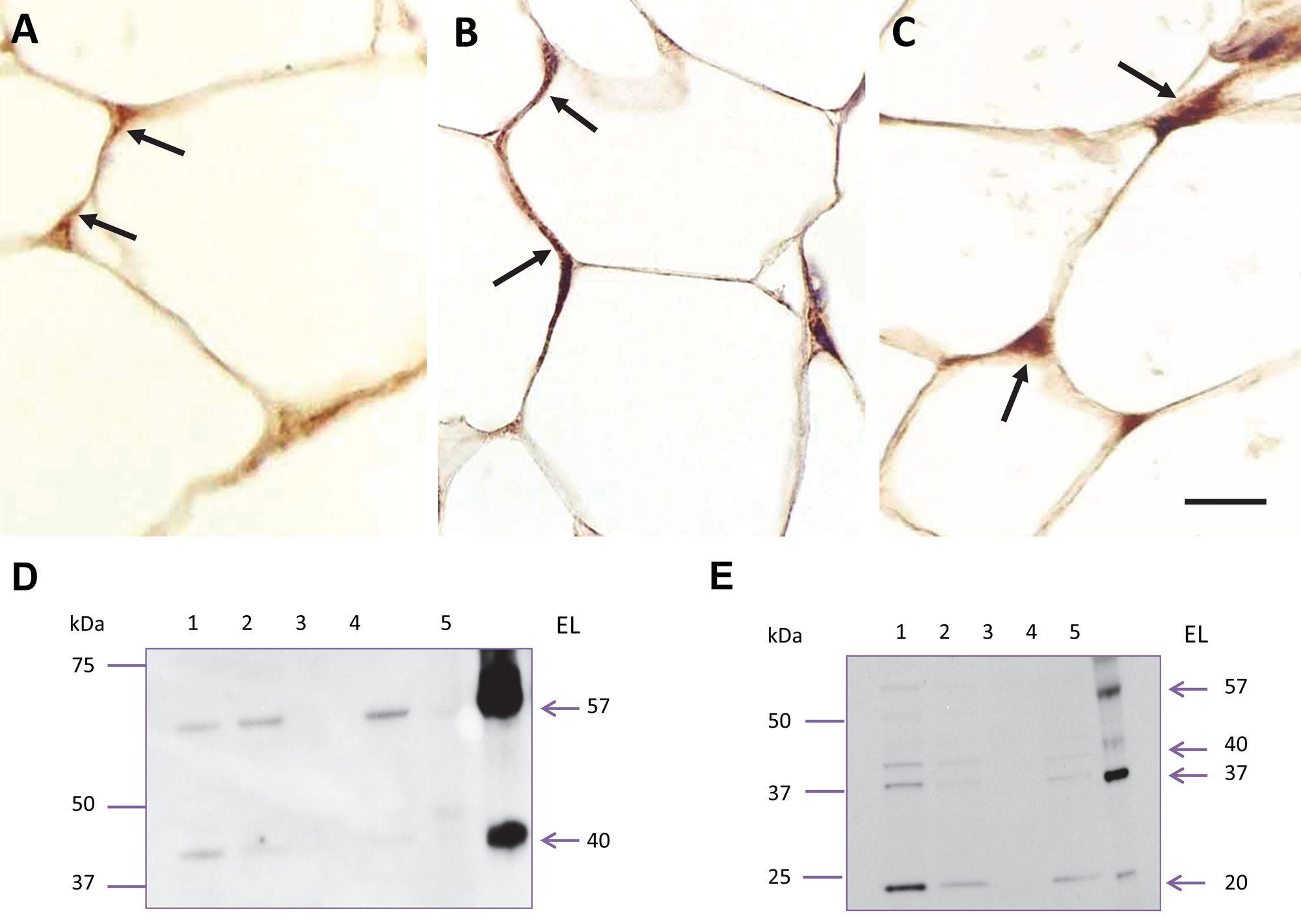
Endothelial lipase (EL) protein expression in human adipose tissue. EL-specific immunostaining was noted in the adipocytes (black arrows) within the tissue that surrounds the thyroid gland. This expression was evident with all three antibodies: (A) EL442–456 antibody, (B) EL19–32 antibody, and (C) EL1–500 antibody. EL protein was also detected on immunoblots of lysates from human omental adipocytes cultured from normal (lane 1) and diabetic (lane 2) patients and from human subcutaneous preadipocytes (lane 3) and differentiated adipocytes (lane 4); these levels were compared with either purified mouse EL protein (lane 5, D; EL19–32 antibody) or human thyroid gland tissue lysates (lane 5, E; ELAAV antibody). Bar = 50 µm (A–C).
Because EL expression in adipose tissue was previously reported using only Northern blots and PCR (Kratky et al. 2005; Paradis et al. 2006; Villaret et al. 2010), immunoblots were used to confirm EL protein expression in adipocytes. Lysates from human omental adipocytes (isolated from both normal and diabetic patients) as well as preadipocytes and differentiated adipocytes (from subcutaneous tissue) were electrophoresed and immunoblotted with the well-characterized, commercially available EL antibody EL19–32. The results indicated that while EL was not expressed in subcutaneous preadipocytes (lane 3, Fig. 5D), it was expressed in omental adipocytes from both normal (lane 1, Fig. 5D) and diabetic (lane 2, Fig. 5D) patients, as well as in differentiated subcutaneous adipocytes (lane 4, Fig. 5D). This observation was confirmed with another well-characterized antibody that has been used frequently for ELISA detection of circulating plasma EL, ELAAV (Fig. 5E). Either purified mouse EL protein (lane 5, Fig. 5D) or human thyroid tissue lysate (lane 5, Fig. 5E) was used as a positive control.
Note that the banding pattern on the immunoblots is different between the two antibodies. These differences are due to the portion of EL that was used to generate the antibodies (a peptide corresponding to amino acids 19–32 from the amino terminal end of EL versus the full-length EL protein expressed in an adenoviral vector in rabbits) and the fact that EL is proteolytically cleaved to smaller forms. The predominant protein bands corresponding to EL protein were similar to those previously reported: 37, 40, and 57 kDa. Of further note is the fact that full-length mouse EL protein was expressed with a flag tag at the C-terminal end and purified using an anti-flag M2 affinity column. The resulting bands on a Coomassie Blue–stained gel were sequenced, and the bands around 57 and 40
EL Localization in Mouse Thyroid Gland and Adipose Tissue
To explore whether EL was also expressed in the mouse thyroid gland and adipocytes, tissue samples from thyroid as well as subcutaneous adipose tissue from mice on a high-fat diet were obtained and processed for IHC. After staining, there was moderate to strong, punctate EL immunolabeling in the follicular epithelial cells that was observed with all three EL antibodies (EL442–456, EL19–32, and EL1–500) (black arrows, Fig. 6A–C). Notably, the mouse follicular epithelial cells were more cuboidal in shape than the follicular epithelial cells found in human thyroid tissue. Additionally, the EL-specific staining was more uniform, as was the size and shape of the follicles. As for the mouse adipose tissue, there were areas of strong, dark brown punctate staining in the adipocytes (black arrows, Fig. 7A, B). As previously noted in the human adipose tissue samples, there were areas of the mouse adipose tissue sections that had significantly less EL-specific staining. In other words, there were areas of little staining and areas of very dark staining that appeared to be areas that were rich in macrophages. These areas were more numerous in adipose from mice that were fed a high-fat diet (Fig. 7A, B) than those fed chow (data not shown). This may be related to the higher amount of tissue inflammation that has been described in adipose from mice fed a high-fat diet (Sun et al. 2011). This is particularly evidenced by the presence of crownlike structures composed of macrophages (open arrows, Fig. 7A, B) that are typically found in metabolically active, inflammation-enriched adipose tissue (Murano et al. 2008). These areas of high inflammation revealed EL staining in both the adipocytes and what appear to be the macrophages. As expected, EL immunoreactivity was also present in the endothelial cells (black arrows) and vascular smooth muscle cells (open arrow) within the adipose tissue vasculature.
Distribution of EL protein in the follicles of the mouse thyroid gland. EL protein expression was visualized by dark brown punctate staining (black arrows) in the cuboidal epithelial cells that line the lumen of each follicle within the thyroid: (A) EL442–456 antibody, (B) EL19–32 antibody, and (C) EL1–500 antibody (magnification ×200). Bar = 75 µm (A–C).
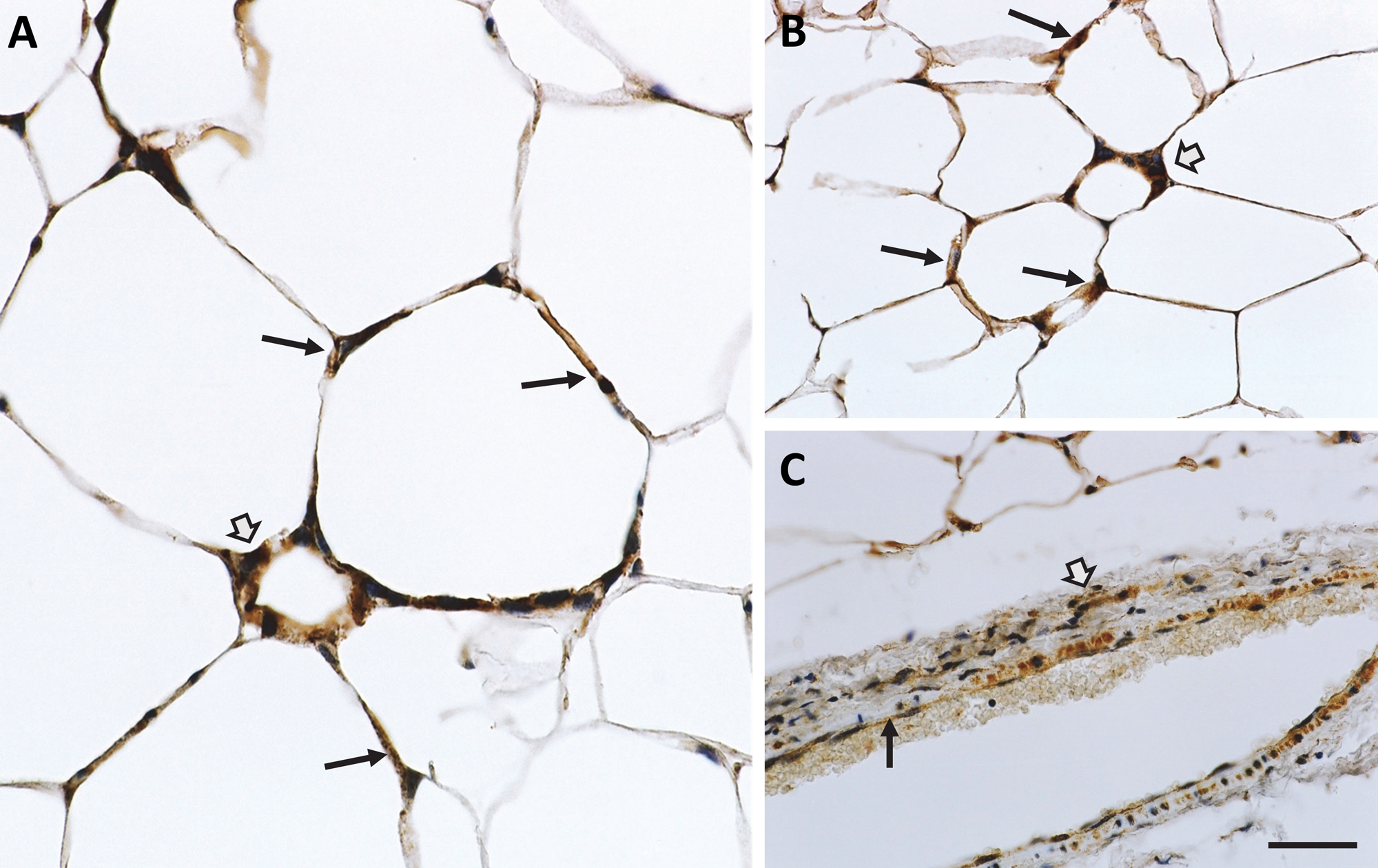
Endothelial lipase (EL) protein expression in mouse adipose tissue. The presence of EL immunoreactivity (black arrows) was detected in subcutaneous adipocytes and the macrophages of the crownlike structures (open arrows) of mice fed a high-fat diet: (A) EL442–456 antibody and (B) EL19–32 antibody. (C) EL19–32 antibody, EL immunoreactivity was also present in the endothelial cells (black arrow) and vascular smooth muscle cells (open arrow) in the adipose tissue vasculature. Bar = 50 µm (A), 100 µm (B), 200 µm (C).
Discussion
Initially, EL was thought to be predominantly expressed in and secreted from endothelial cells. Similar to HL, secreted EL protein was reported to be associated with heparan sulfate proteoglycans on the cell surface (Fuki et al. 2003). Evidence supports the fact that EL is also expressed in a variety of other tissues and cell types and can be found intracellularly as well as associated with the cell surface (Ishida et al. 2004b; Nielsen et al. 2009). Similar to the expression pattern of EL in the lung, where EL protein was found to be expressed in epithelial cells, alveolar type II cells, and endothelial cells (Otera et al. 2009), EL expression in the thyroid gland was found in follicular epithelial cells as well as in the endothelial and smooth muscle cells of the blood vessels. EL labeling in the follicular epithelial cells showed possible membrane labeling that was more prominent on the luminal, apical surfaces of these epithelial cells. Significant levels of EL expression were not detected in the parafollicular cells.
The most compelling finding from our study was the detection of intense EL-positive immunolabeling in human and mouse adipose tissue. Similar to the follicular epithelial cells, there appeared to be both cell surface and intracellular EL immunostaining in the adipocytes. There have been reports of cytoplasmic expression of isoforms of EL that are expressed in most cell types (Ishida et al. 2004b). Some of the bands detected by ELAAV antibody appear to be similar in size to the non-secreted isoforms of EL that have been reported. In addition, it may be that EL is being actively synthesized and processed by the thyrocytes and adipocytes, producing what appears to be cytoplasmic localization. Paradis et al. demonstrated that plasma EL levels were positively correlated with body mass index and a proatherogenic lipid profile (Paradis et al. 2006). Moreover, the authors showed that EL mRNA levels were similar in subcutaneous and omental adipose tissue and that EL message levels were much lower than LPL message levels in these tissues. However, because the levels of EL message were very low, much lower than those of LPL, they concluded that the correlation between elevated plasma EL levels and increased visceral adiposity was significant but unlikely to be causal (Paradis et al. 2006). Our data suggest that while EL mRNA may be low in human adipose tissue, EL protein levels are fairly high and may contribute to plasma circulating EL in patients that are obese and/or categorized as having metabolic syndrome. Furthermore, EL protein expression in mouse adipose tissue appeared to be higher in areas of the adipose tissue that had a higher number of macrophages and dead adipocytes (i.e., crownlike structures). EL immunoreactivity was also detected in what appear to be the macrophages. Notably, areas dense with macrophages or crownlike structures were not observed in the human adipose tissue samples that were investigated for this study. These regions of high inflammation were also not observed when adipose tissue samples were obtained from patients that were obese or diagnosed with metabolic syndrome. EL expression patterns may differ somewhat in human adipose tissue samples that reflect a high inflammatory state.
It has been hypothesized that EL is highly expressed in tissues that have high metabolic rates and are highly vascularized (Choi et al. 2002). Since the thyroid gland and lung are metabolically active and are unable to store fatty acids, these organs may use exogenous fatty acids hydrolyzed via EL’s phospholipase activity to maintain their high rates of metabolism (Choi et al. 2002). EL expression in adipocytes may also be related to the level of inflammation in the particular adipose tissue depot. EL appears to be expressed in several adipose tissue depots: subcutaneous, omental, as well as adipose tissue surrounding the thyroid gland. Despite the fact that EL expression may be protective under some conditions, it is clear that overexpression of EL can lead to an exacerbation of the pathological response. EL presence in the arterial wall is clearly one of these cases where overexpression of EL has been strongly associated with increased monocyte adhesion and increased atherogenesis. Similarly, in adipose tissue, EL overexpression leads to or is a consequence of a higher inflammatory state, which may result in enhanced secretion of EL into the circulation. Increased plasma EL is associated with decreased HDL. Therefore, understanding EL function, in follicular epithelial cells of the thyroid, adipocytes, and macrophages in inflamed adipose tissue, may be key to understanding the potential role that EL plays in the dyslipidemia that is associated with metabolic diseases, such as metabolic syndrome, obesity, and type II diabetes.
Footnotes
Acknowledgements
We thank the Rader laboratory for generously providing the anti-human EL antibody, Thomas Garrabrant for coordinating development of EL antibodies at New England Peptide, Sharon L. Burke for characterization of the EL antibodies, and Danielle Lawrence and Michael Alicknavich for the IHC. We also kindly acknowledge Andrew L. Darrow for his intellectual input and overall support of the EL project.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: All authors were employees of Janssen Research and Development during the conduct of these studies.