
Abstract
C-terminal amidation is a post-translational processing step necessary to convey biological activity to a large number of regulatory peptides. In this study we have demonstrated that the peptidyl-glycine α-amidating monooxygenase enzyme complex (PAM) responsible for this activity is located in the medullary stellate epithelial cells of the thymus and in cultured epithelial cells bearing a medullary phenotype, using Northern blot, immunocytochemistry, in situ hybridization, and enzyme assays. Immunocytochemical localization revealed a granular pattern in the cytoplasm of the stellate cells, which were also positive for cytokeratins and a B-lymphocyte-associated antigen. The presence of PAM activity in medium conditioned by thymic epithelial cell lines suggests that PAM is a secreted product of these cells. Among the four epithelial cell lines examined, there was a direct correlation between PAM activity and content of oxytocin, an amidated peptide. Taken together, these data provide convincing evidence that thymic epithelial cells have the capacity to generate amidated peptides that may influence T-cell differentiation and suggest that the amidating enzymes could play an important role in the regulation of thymic physiology.
Keywords
C-
PAM enzymes have been co-localized with ami-dated regulatory peptides inside secretory granules (Martínez et al. 1993b). The localization of PAM gene expression in a particular cell population is indicative of processing of biologically active regulatory peptides (Cuttitta 1993), although in some instances the identification of PAM has preceded that of the peptide hormone (Martínez and Treston 1996).
The thymus has a crucial role in the development and organization of the immune system. Maturation of primitive incompetent precursor cells to immunocompetent T-lymphocytes takes place in the thymus, and accumulating evidence points to the involvement of a secretory system of regulatory substances in this process (Nabarra and Adrianarison 1987; Geenen et al. 1992; Faas et al. 1993). A number of amidated peptide hormones have been reported to be involved in the lymphocyte maturation process (Nabarra and Adrianarison 1987; Ogata et al. 1989; Robert et al. 1991; Geenen et al. 1992; Faas et al. 1993; Friend et al. 1994).

Structure of the pre-pro-enzyme, showing the localization of the two regions (CC and PAL2) against which antisera were raised and the probes used for Northern blot (hPAM 800) and for in situ hybridization (hPAM 1200) in this study. The main functional regions of the molecule are labeled: signal peptide (sp), cleavage site (cs), transmembrane site (tms), and cytoplasmic domain (cd).
A previous study (Braas et al. 1989) showed that the thymus as a whole produces very low amounts of PAM mRNA and that the enzyme specific activity is also low. We planned this study to determine more accurately the expression of PAM in individual thymic cell populations and the possible physiological implications of amidation as part of the post-translational activation of regulatory substances in the development of T-lymphocytes.
Materials and Methods
Twelve female Wistar rats weighing from 150-200 g and four Balb/C mice were used in this study.
Northern Blot Analysis
Three rats and two mice were sacrificed by cervical dislocation and several tissues, including thymus, were dissected out and total mRNA was prepared, after homogenization, using the guanidine isothiocyanate-cesium chloride method of Glisin et al. (1974). Total RNA (10 μg) was electrophoresed on a denaturing 1.2% agarose gel and transferred to a nitocellulose membrane. For fragment size determination, an RNA ladder was fractionated in an adjacent lane. Integrity of RNA and equivalence of loading were checked with ethidium bromide under
Immunocytochemistry
Animals were anesthesized with an overdose of sodium pentobarbital and perfused through the left ventricle with PBS and 4% paraformaldehyde in PBS. The thymuses were excised and immersed for 3 additional hr in fixative. Tissues were embedded in paraffin or in epon.
Paraffin (4-μm-thick) and semithin plastic (1-μm-thick) sections were mounted on glass slides and immunocytochemical staining was performed using the avidin-biotin method (Hsu et al. 1981). To prevent nonspecific background, the sections were pretreated with 3% H2O2 in methanol for 30 min and with normal swine serum, 1:20 in PBS, for 30 min, and incubated overnight at 4C with the primary antisera.
Polyclonal antisera were raised against synthetic fragments of the predicted sequence of human PAM; CC (PAM 288–310 with a Val for Ala conservative substitution at position 11) and PAL2 (PAM 527–546) as previously described (Martínez et al. 1993a,b; Figure 1). They were used at an 1:800 dilution. Other antibodies used include a monoclonal antibody against cytokeratins (AE1/AE3; Boehringer-Mannheim, Indianapolis, IN), and an antibody against the B-cell common marker CD-20 (Dako; Carpinteria, CA).
The sections were incubated with biotinylated secondary antibodies (Dakopatts; Glostrup, Denmark) and then with avidin-biotin-peroxidase complex (Dakopatts). The bound antibodies were visualized with 3–3′-diaminobenzidine tetrahydrochloride (Sigma; St Louis, MO). In some cases the reaction was enhanced with a nickel solution (Shu et al. 1988).
As specificity controls, antisera to PHM (CC) and PAL (PAL2) were preincubated with their respective synthetic antigens for 2 hr at RT at a concentration of 20 nmol of pep-tide/ml of optimally diluted antiserum, before application to tissue sections.
In Situ Hybridization
Paraffin-embedded thymus tissue was processed for in situ hybridization as previously described (Martínez et al. 1993a; Steel et al. 1994). In summary, sections were mounted on Vectabond (Vector Laboratories; Burlingame, CA)-coated slides. Labeled probes from the hPAM1200 plasmid (Glauder et al. 1990; Figure 1) were prepared using digoxigenin-11-UTP (Boehringer) and T7 or T3 RNA polymerases (Boehringer) to synthesize sense and anti-sense RNA transcripts, respectively. Hybridization was performed in a moist chamber at 46C for 20 hr in a 15-μl volume containing 0.5 ng/μl of probe. Stringency washes included different treatments with SSC and sodium dodecyl sulfate (SDS), incubation in 2 X SSC containing 10 μg/ml RNase at 37C for 15 min, and further rinses in 2 X SSC.
Visualization of digoxigenin was performed with a monoclonal antibody coupled to alkaline phosphatase (Boehringer) 1:500, acting for 2 hr at RT. Nitroblue tetrazolium chloride and 5-bromo-4-chloro-3-indolyl-phosphate (both from Sigma) were used as substrates for the alkaline phosphatase.
Controls included the use of the sense probe and digestion with RNase before the hybridization.
Cell Culture
Four thymic cell lines of mouse origin, previously characterized as epithelial stromal cells of the medulla (Farr et al. 1989,1993; Friend et al. 1994) were used in this study: TE-71, Z-172, Z-199R, and Z-210R. They were maintained in DMEM with added 20 μM
Cell Extracts
Whole-cell extracts were prepared from approximately 5 × 106 cells homogenized in 2 ml of TMT (20 mM TES, pH 7.4, 10 mM mannitol, 1% Triton X-100) containing a protease inhibitor cocktail of phenylmethylsulfonylfluoride, bacitracin, aprotinin, benzamidine, and leupeptin. The homogenate was then sonicated at high power with a Branson ultrasonic probe and ultracentrifuged at 100,000 X g for 30 min. Media that had been conditioned for 24 hr by exponentially growing cells were also collected for enzymatic activity assays of secreted PAM.
PAM Enzyme Assay
This assay was done according to a procedure previously described (Treston et al. 1993). In summary, the synthetic glycine-extended peptide substrate, acYFG (N-acetyl-tyrosyl-phenylalanyl-glycine), is converted to the α-amidated product acYF# (N-acetyl-tyrosyl-phenylalanylamide) by the sequential action of the PHM and PAL enzymes present in the sample. The assay mix contains concentrations of Cu2+ and ascorbate adjusted to yield maximal enzyme activity (a range of copper concentrations approximately 1 μM Cu2+ for each 0.5 μg of analyte protein, and 0.5 mM ascorbate). The enzyme activity is measured with a trace amount of radioiodinated substrate (20,000 cpm [125I-Tyr]-acYFG). Five copper concentrations were analyzed and the activity quoted was calculated from the percent conversion at the maximal concentration. Specific activities were calculated from enzyme activities and protein concentrations determined using the Pierce bicinchoninic acid reagent in the microtiter plate format. Modifications of the assay are used to determine individual PHM and PAL activities, as we have described (Treston et al. 1993).
RT-PCR Analysis
Total RNA was reverse-transcribed and amplified in a single solution as previously described (Myers and Gelfand 1991). The rTth DNA polymerase kit (Perkin-Elmer; Norwalk, CT) was used for this assay according to the manufacturer's specifications. Primer sets for amplification of PHM and PAL specific regions were as follows: PHM sense primer (PAM 255–273), 5′-AGC-CCA-CTT-TCT-GTC-TTT-A-3′; PHM anti-sense primer (PAM 637–655), 5′-TCT-CCT-CCA-ACT-CTG-AAT-C-3′; PAL sense primer (PAM 1812–1828), 5′-AGC-AAG-TTT-GTT-TAC-CA-3′; PAL anti-sense primer (PAM 2190–2206), 5′-GAC-TCT-TCT-CCC-CAC-TG-3′. Expected sizes for the PCR products are 401 and 395
Oxytocin Radioimmunoassay
The presence of oxytocin in the conditioned media was tested using the oxytocin RIA kit from Peninsula Laboratories (Belmont, CA) according to the manufacturer's specifications: 100 ml of conditioned medium was passed through a Sep-pak cartridge (Millipore; Bedford, MA), eluted in 3 ml of 60% acetonitrile-0.1% trifluoroacetic acid, freeze-dried, and reconstituted in RIA buffer. A total of 100 μl of each sample was incubated overnight with 100 μl of antioxytocin antibody. The next day, [125I]-oxytocin was added to the solution, and on the third day 100 μl of goat anti-rabbit IgG and normal rabbit serum were added before centrifugation. The radioactivity left in the pellet was measured in a γ-counter and the cpm values were corrected according to the total counts and the nonspecific binding.
Results
The Northern blot analysis showed a very faint, almost imperceptible band for the thymus compared with other tissues known to express high quantities of PAM mRNA (Figure 2).
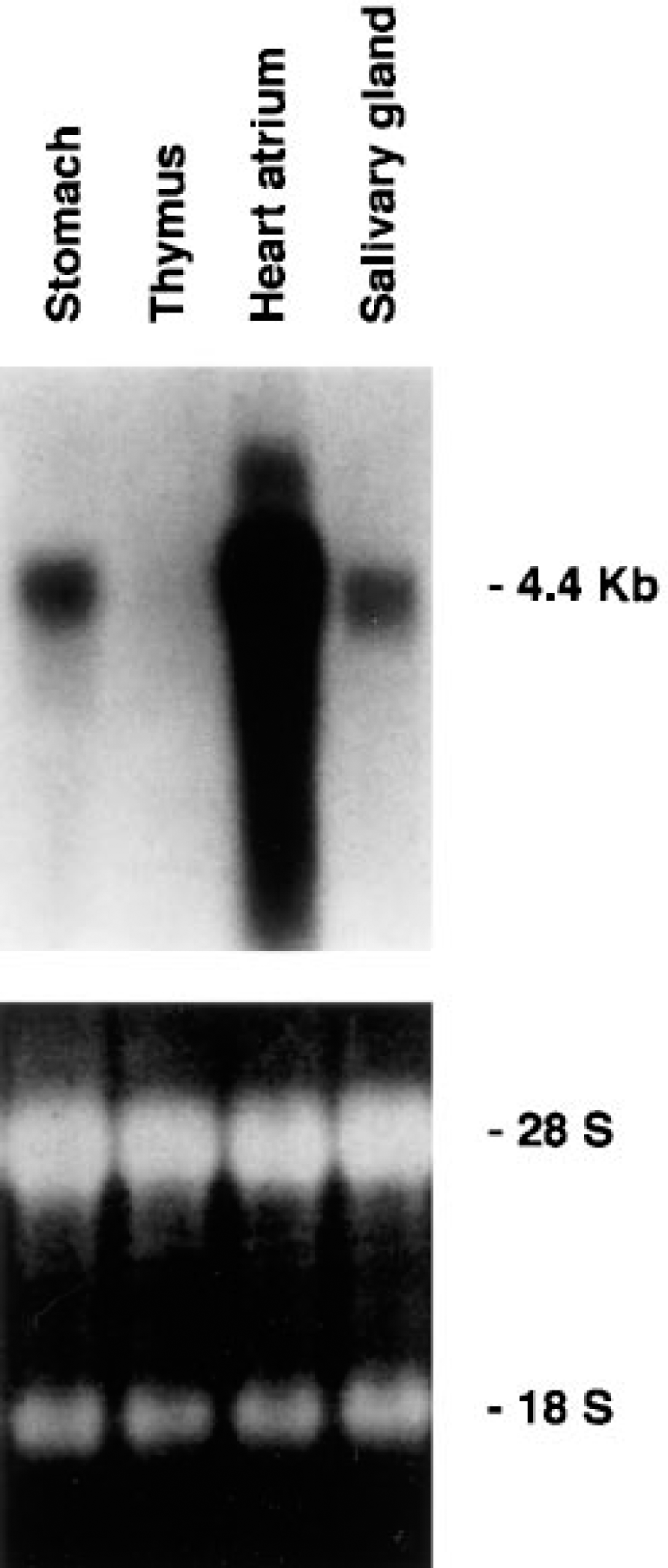
Northern blot analysis of rat total RNA extracted from stomach, thymus, heart atrium, and salivary glands probed for expression of PAM gene (
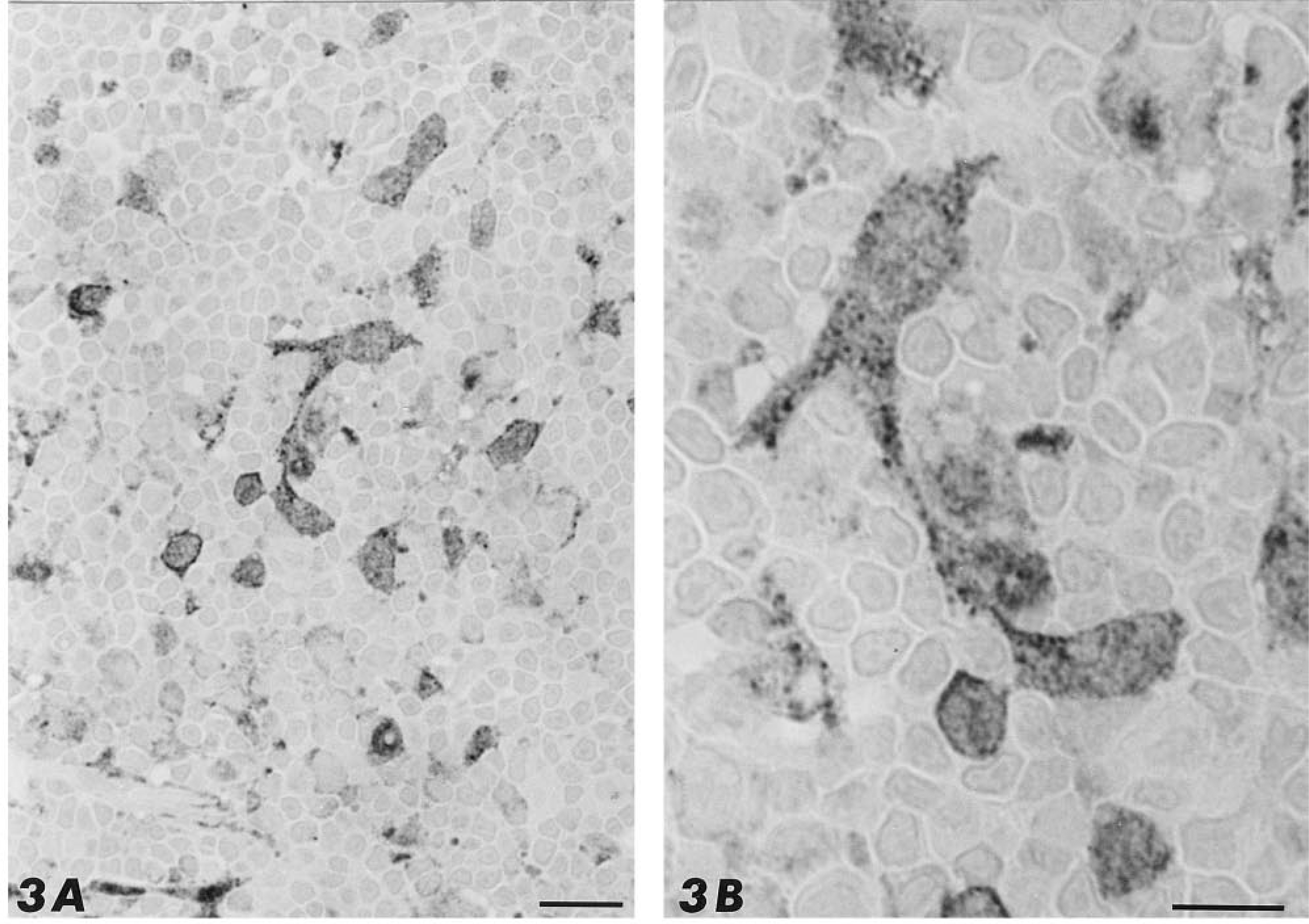
Immunocytochemical analyses demonstrated that thymus tissue reacted with antibodies directed against PHM but not PAL. The labeling was restricted to stromal stellate cells of the thymic medulla (Figure 3). These cells are characterized by a large, pale nucleus and the presence of several cytoplasmic processes (Figure 3A). The immunoreactivity was located in the cytoplasm of the medullary stellate cells displaying a patent granular pattern (Figure 3B), whereas thymocytes were absolutely devoid of staining. Preabsorption of the antibody with its synthetic peptide completely quenched the immunostaining.
Semithin section of rat thymus immunostained for PHM. Large, immunoreactive cells bearing long cytoplasmic processes can be observed among the unstained thymocytes. Bar = 20 μm. (
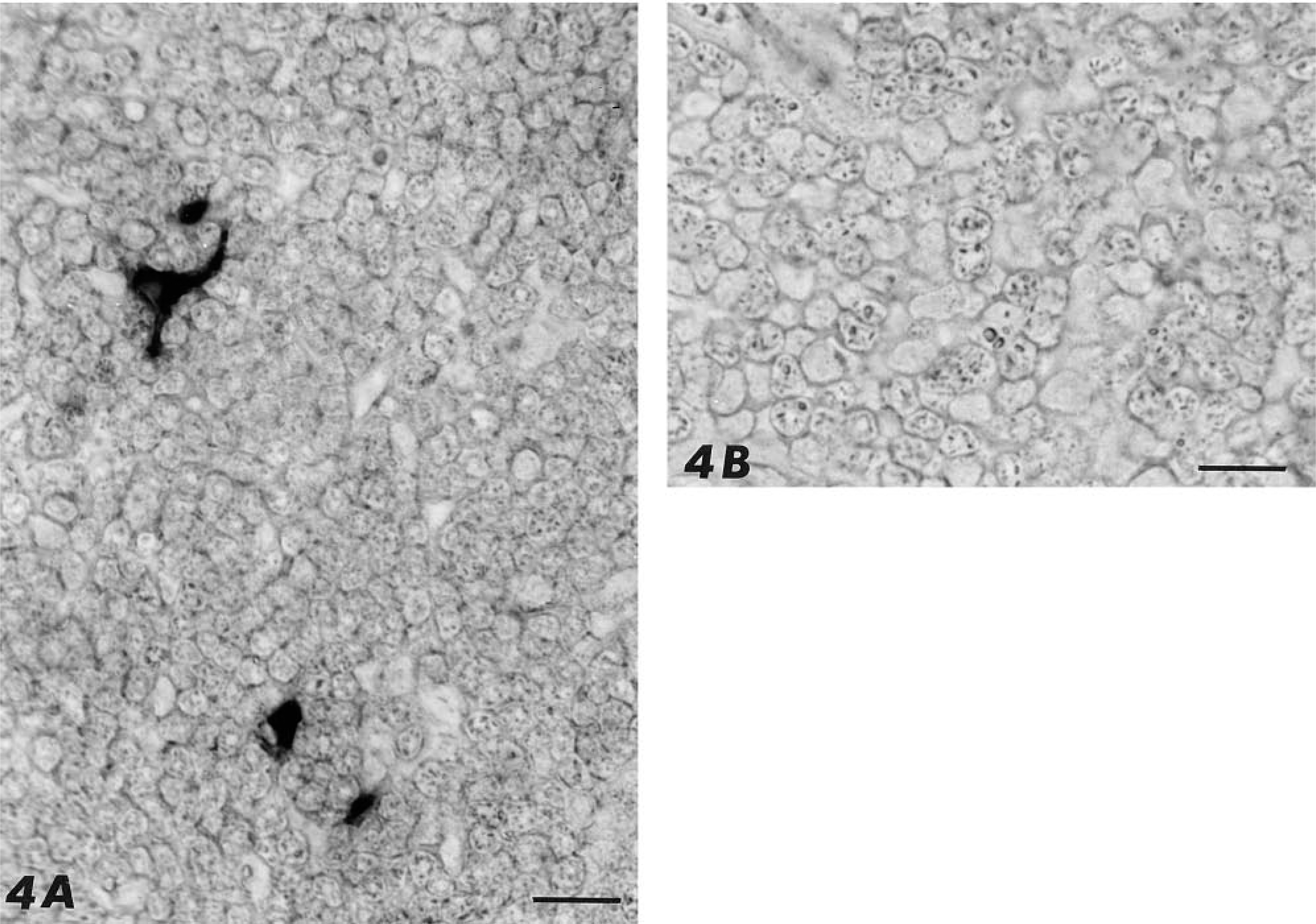
Paraffin sections of rat thymus labeled by the in situ hybridization technique. (
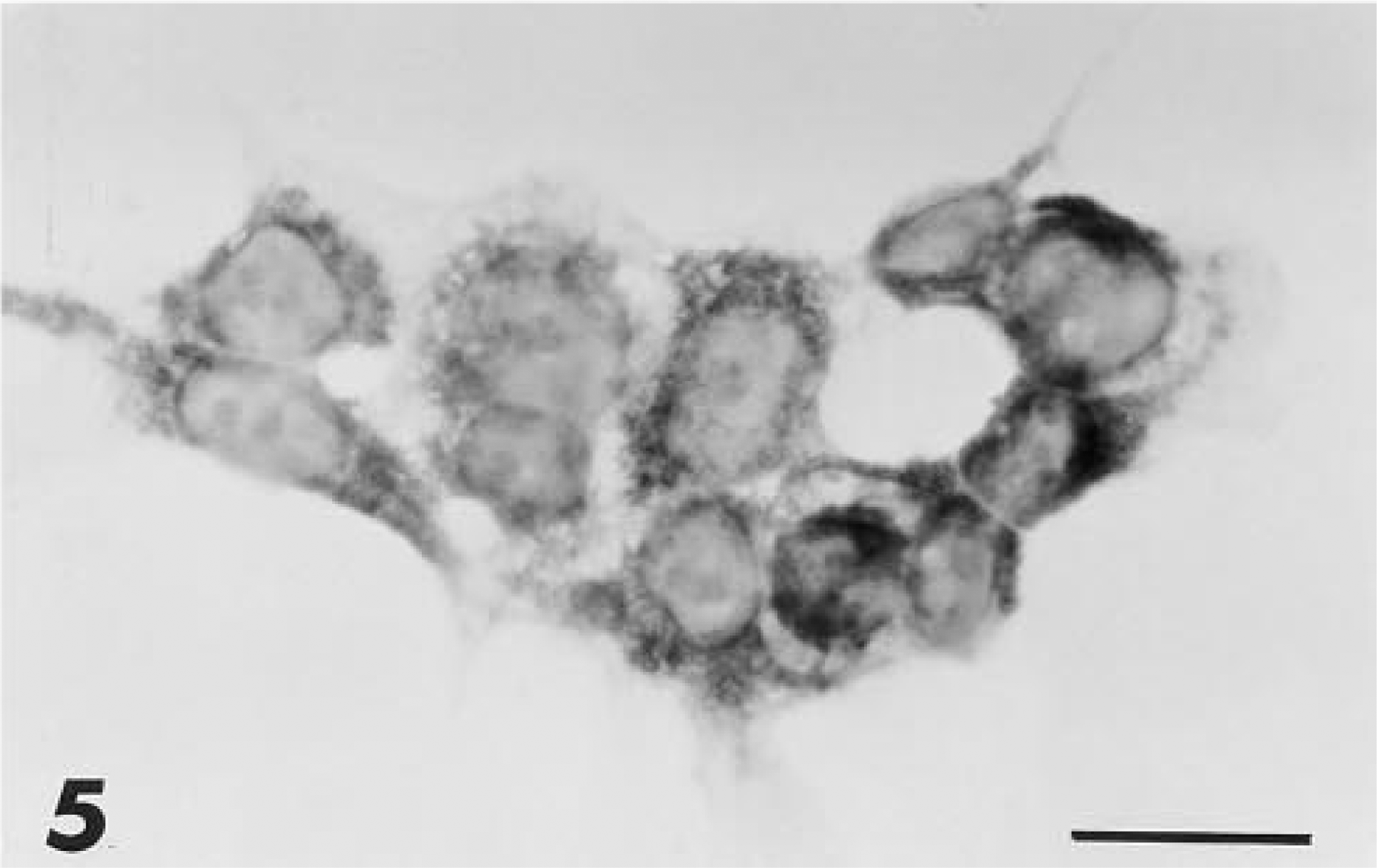
PHM immunoreactivity in the cytoplasm of the cell line Z-199R. Bar = 10 μm.
The expression of PAM mRNA in thymus was determined by in situ hybridization (Figure 4). Distribution of the positively hybridized cells was restricted to the thymic medulla and application of the sense probe gave no staining at all (Figure 4B). Staining for the mRNA was homogeneously distributed throughout the stellate cytoplasm and the number of positive cells was lower (around 20%) compared to the PHM-immunore-active cells (compare Figure 3A to Figure 4A).
To further characterize the cell type producing PAM in the thymus, a study of four medullary thymic stromal lines was performed. Similar to the results of the in situ studies, cytoplasmic labeling of thymic epithelial cell lines was observed with anti-PHM but not with anti-PAL antiserum (Figure 5). Labeling of serial sections allowed a more complete characterization of the PAM-positive cells of the thymus (Figure 6). They were also immunoreactive to antibodies against cytokeratins, thus confirming their epithelial origin, and to B-lymphocyte associated antigens, a characteristic previously reported for stellate cells in human thymic medulla (Hofmann et al. 1988; Taubenberger et al. 1991).
Northern blot analysis showed that all of the medullary epithelial cell lines contained PAM mRNA (Figure 7). In all instances, PAM mRNA levels were lower than described previously for neuroendocrine lung tumor cell lines (Treston et al. 1993), as seen in the last two lanes of Figure 7.
RT-PCR analysis was performed to investigate the differences in expression between PHM and PAL. Other organs, including stomach, heart, and salivary glands, expressed both regions of the message. Conversely, the thymus extracts were positive only for PHM and not for PAL (Figure 8), indicating that the major spliced form of the PAM mRNA that is expressed in the thymus lacks the PAL domain.
PAM enzymatic activity (Table 1) was lowest in the TE-71 epithelial cell line, highest in the Z172 line, and intermediate in the remaining two. The majority (95%) of either PHM or PAL activity recovered from stromal cell cultures was found in the conditioned medium, indicating the secretory nature of the processed PAM gene products.
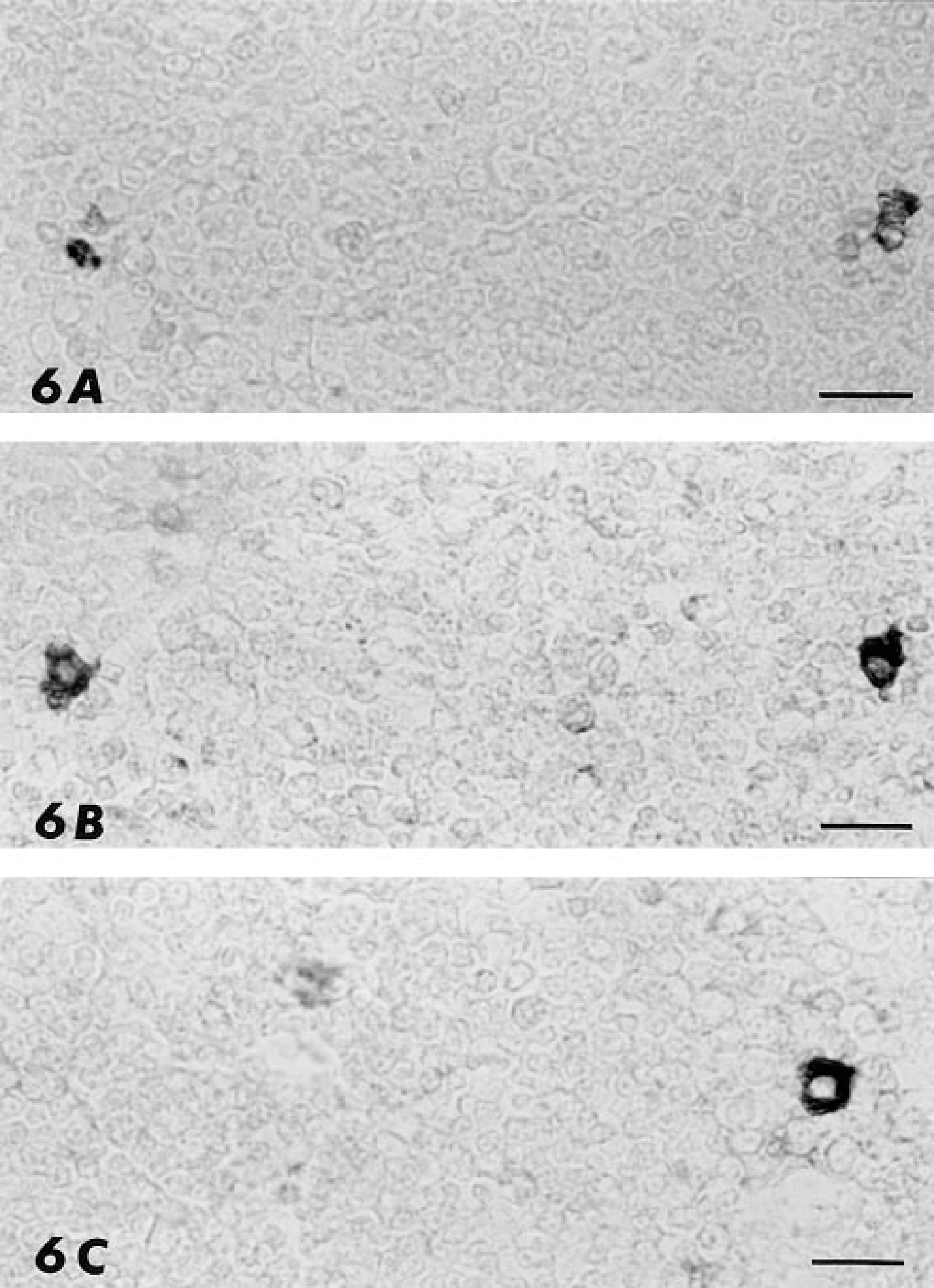
Serial sections of mouse thymus immunostained with antibodies against cytokeratin-14 (
To identify amidated peptides in these cell lines, we performed a radioimmunoassay for oxytocin, one of the amidated peptides described in thymus (Robert et al. 1991), and found a positive correlation between oxytocin secretion and PAM activity (Table 1).
Discussion
This study shows for the first time that the amidating enzymes are expressed in the stellate epithelial cells of the thymic medulla. The epithelial thymic cells form a meshwork in the organ parenchyma. Cortical epithelial cells are very reticular in shape, whereas the medullary cells comprise two types: stellate and large, vacuolated elements (Ushiki 1986; Nabarra and Adrianarison 1987). The interpretation of the images found in this study, together with their immunoreactive pattern, suggests that only the stellate epithelial cells of the medulla express PAM.
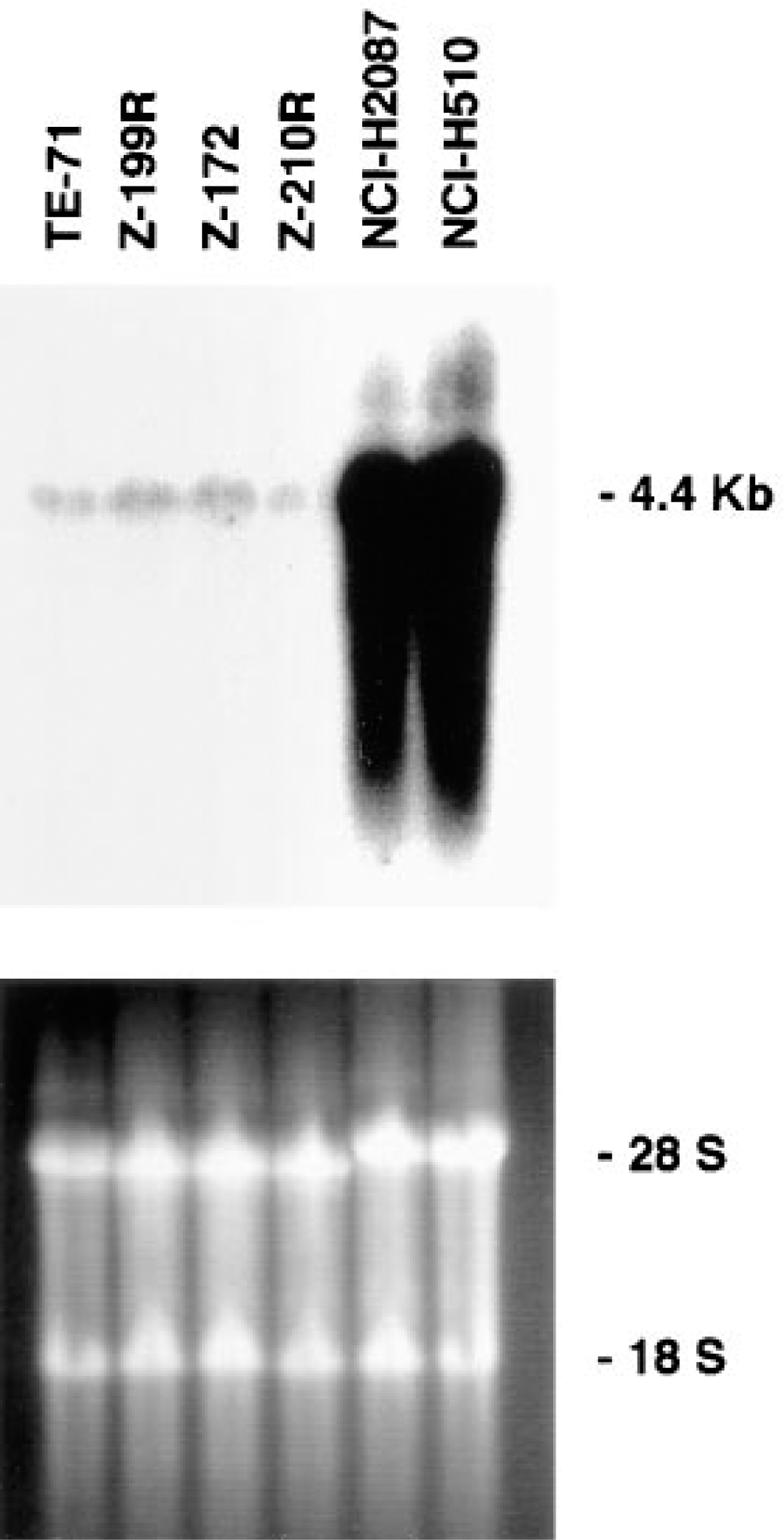
Northern blot analysis of total RNA extracted from six cell lines, which include four stromal epithelial cell lines of the thymic medulla and two non-small-cell lung carcinomas (NCI-H2087 and NCI-H510) (
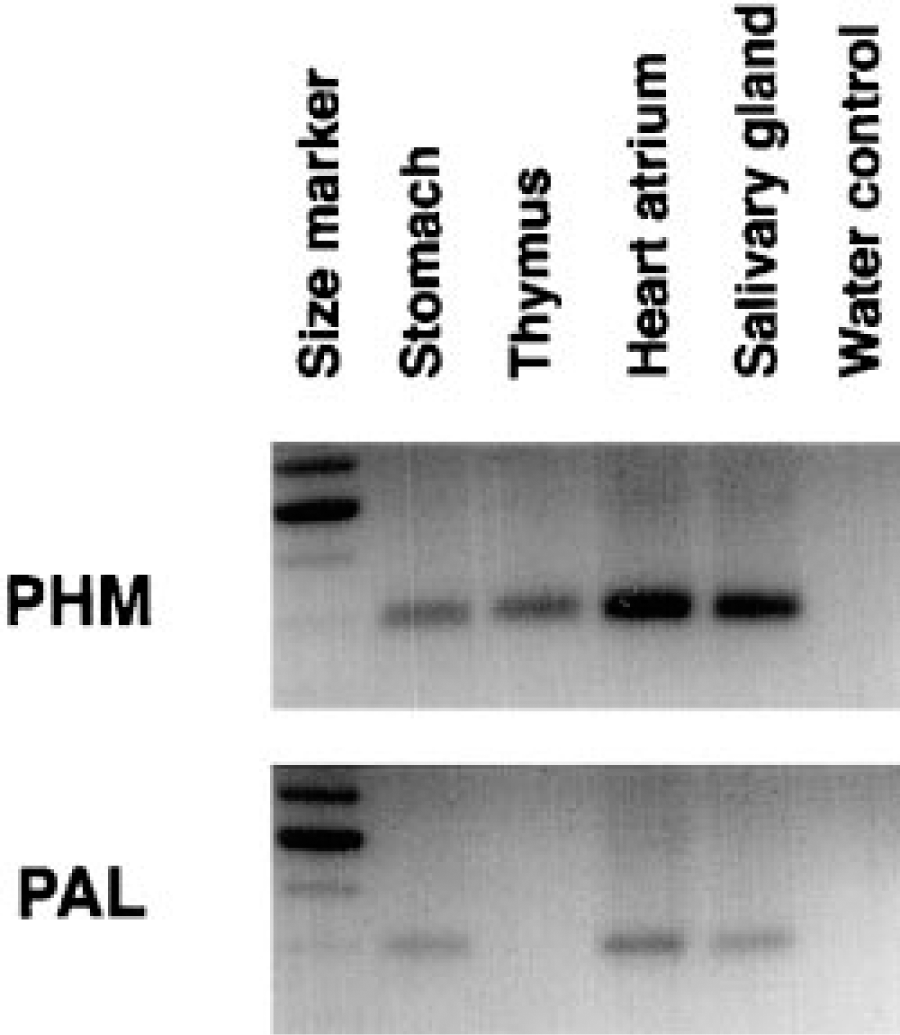
RT-PCR analysis performed on total RNA extracted from different rat organs. PHM was found in the four organs studied, but PAL was lacking in the thymus.
The low amount of mRNA revealed by Northern blot analyses of thymus RNA in this study and in a previous general survey (Braas et al. 1989) could be explained by the low frequency of the medullary stellate cells relative to other cell components of the thymus. Consistent with this explanation, relatively few cells in the thymus were found to express substantial levels of PAM mRNA and immunoreactive PHM. Furthermore, analyses of thymic epithelial cell lines indicated that PAM mRNA in the thymic epithelial cells was not as abundant as in lung tumor cell lines.
PAM activity and oxytocin levels in thymic medullary cell lines in culture; mean ± SD of four repeats are shown a
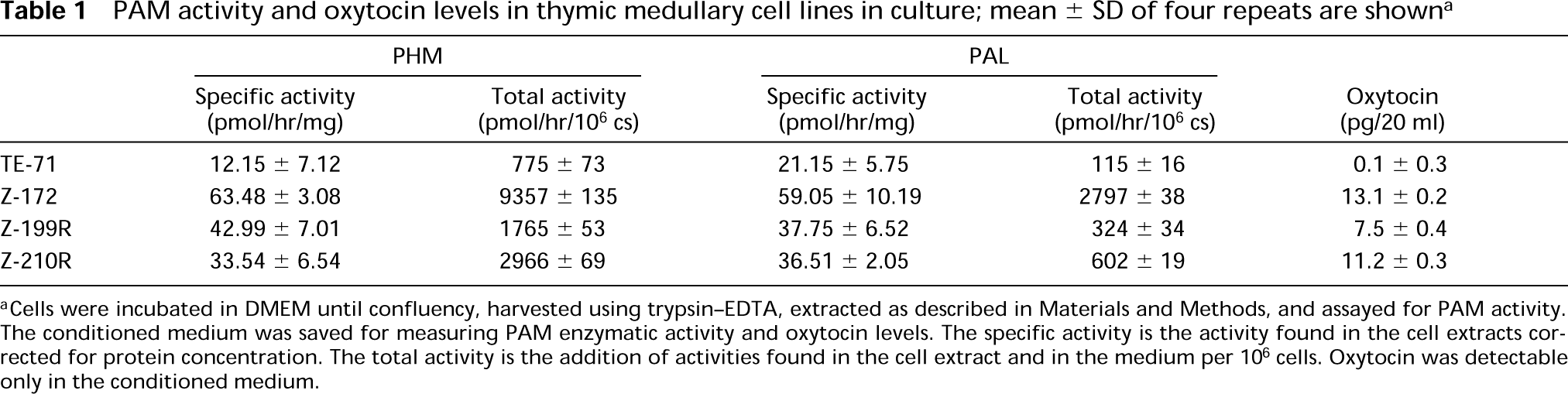
aCells were incubated in DMEM until confluency, harvested using trypsin-EDTA, extracted as described in Materials and Methods, and assayed for PAM activity. The conditioned medium was saved for measuring PAM enzymatic activity and oxytocin levels. The specific activity is the activity found in the cell extracts corrected for protein concentration. The total activity is the addition of activities found in the cell extract and in the medium per 106 cells. Oxytocin was detectable only in the conditioned medium.
The finding here that some PAM-producing cells are positive for the CC antiserum and not for PAL2 has been previously reported (Martínez et al. 1993a). Comparison of PHM and PAL activities in extracts of thymic epithelial cell lines or in media conditioned by these cells indicates that PAL activity is significantly lower than PHM activity. This pattern is the reverse of what has been observed in other cell lines (Treston et al. 1993), suggesting that PHM and PAL enzyme levels can be differentially regulated by different cell types. These facts, together with the RT-PCR experiments, suggest that thymic stellate cells may express high levels of a mRNA form lacking the PAL domain, similar to the PAM4 variant that was isolated from rat cDNA libraries (Eipper et al. 1992). The relatively low levels of PAL activity exhibited by thymic epithelial cells in vitro and the inability to amplify the PAL region or to demonstrate immunoreactive PAL in situ or in vitro, together with the finding that PHM can mediate complete amidation in the absence of PAL under some conditions (Noguchi et al. 1989), suggest that the PAL enzyme may not contribute significantly to amidation reactions occuring within the thymic environment.
The lower number of positive cells found with the in situ hybridization method, as compared with immunohistochemistry, suggests that the expression of the PAM gene is not constitutive in the thymus but follows an intermitent pattern, possibly related to physiological conditions. A similar pattern has been observed for PAM expression in the human pituitary (Steel et al. 1994). Alternatively, storage of PAM protein products within the cells, or different sensitivities of the two techniques, could explain these results.
Over the past years, evidence has accumulated that the thymus secretes biologically active factors and that these factors are secreted by the epithelial cells of the thymic reticulum (Nabarra and Adrianarison 1987). The secreted substances reported in the literature include, among others, tumor necrosis factor (TNFα), interferon γ, several interleukins, the hormone thymulin, neurohypophysis-related peptides, and tachykinins (Nabarra and Adrianarison 1987; Robert et al. 1991; Geenen et al. 1992; Faas et al. 1993; Friend et al. 1994). These cells also express the major histocompatibility complexes MHC I and MHC II (Faas et al. 1993). The secretion and interactions of these substances generate a microenvironment for T-lymphocyte differentiation and selection (Pircher et al. 1993; Rieker et al. 1993). The expression of the amidating enzymes in the stellate epithelial cells and the production of at least one amidated peptide by thymic epithelial cells indicate the potential for this class of regulators to contribute to the creation of the thymic environment.
It has been shown that several amidated regulatory peptides, including substance P, cholecystokinin 8 (CCK8), and vasoactive inhibitory peptide (VIP), among others, regulate the growth and differentiation of T-lymphocytes (Flageole et al. 1992; Ottaway 1992; Anton et al. 1993; Tang et al. 1993). It is possible that some of these substances and others are secreted by the thymic stromal cells as a component of the specific micro-environment, and therefore thymic epithelial cells require the amidating machinery to complete the maturation of the regulatory peptides. Interestingly the cell lines found to express higher levels of PAM activity and of oxytocin secretion also support lymphopoiesis in vitro (Friend et al. 1994). The contribution of amidated regulatory peptides to this activity has not yet been determined.
In summary, PAM is expressed by medullary stellate epithelial cells of the thymus and is probably involved in the post-translational maturation of regulatory peptides secreted by these cells. This study opens a new avenue in the understanding of thymic regulation.
Footnotes
Acknowledgements
Supported by a grant from Spain's Government (Ministerio de Educación y Ciencia, pf9316525626) to A.M. and by grants from the National Institutes of Health (Ag 04360, AI 24137) and the Department of Energy (DE-F006-86-ER60409) to A.F.
We gratefully thank Prof J. Engels for the gift of the PAM probes. This study was initiated by A. Martínez while at the Department of Cytology and Histology, University of Navarra, Pamplona, Spain.