
Abstract
Phocine distemper virus (PDV) caused thousands of deaths among harbor seals (Phoca vitulina) from the North Sea in 1988 and 2002. To examine the effects of different factors on the pathology of phocine distemper, we performed necropsies and laboratory analyses on 369 harbor seals that stranded along the Dutch coast during the 2002 PDV epidemic. Diagnostic tests for morbillivirus infection indicated a differential temporal presence of morbillivirus in lung and brain. Seals of 3 years or older were significantly more often IgG positive than younger seals. The most frequent lesions in PDV cases were bronchopneumonia, broncho-interstitial pneumonia, and interstitial emphysema. Extra-thoracic emphysema was rare in <1-year-olds compared with older seals, even though severe pneumonia was more common. PDV cases generally had empty stomachs and less blubber than by-caught seals from before the epidemic. In PDV cases involving older animals, lung, kidney, and adrenal weights were significantly increased. Bordetella bronchiseptica was isolated from lungs in two thirds of the PDV cases examined. Our results indicate that brain should be included among the tissues tested for PDV by RT-PCR; that either phocine distemper has a longer duration in older seals or that there are age-related differences in immunity and organ development; that dehydration could play a role in the course and outcome of phocine distemper; and that bacterial coinfections in lungs are more frequent in PDV cases than gross lesions suggest. These results illustrate how quantitative analysis of pathology data from such epidemics can improve understanding of the causative disease.
Introduction
Phocine distemper virus (PDV), a morbillivirus affecting marine mammals, caused thousands of deaths in the harbor seal (Phoca vitulina) populations of the North Sea in 1988 and again in 2002. 18, 24, 27, 45 The PDV epidemics occurred in the summer, during the harbor seal birth, molt and breeding season. 24, 58 At this time of the year, harbor seals show relative site fidelity, hauling out on (mostly intertidal) sandbanks in the Wadden Sea (3,600 counted in 2001) or in the Delta area (150 counted in 2001). 46, 59, 61, 63 Pups are born between the end of May and the beginning of July, and weaned approximately 24 days after birth. 38 The mating season is from July to August. 63 The exact timing of molt during the summer varies with seal age and sex. 56 During molt and breeding activities, seals may partly or completely reduce their food intake. 15, 59 Besides harbor seals, there were approximately 500 gray seals (Halichoerus grypus) in the Wadden Sea, hauling out mainly on a sandbank between Terschelling and Vlieland. 60 PDV-associated deaths were not observed in this species. 45
During both epidemics, seal carcasses that stranded along the North Sea coasts during the 2 epidemics were necropsied, and the pathology of PDV was described in harbor seals from the UK, Denmark, Sweden, and Germany. Broncho-interstitial pneumonia, interstitial pneumonia, or suppurative bronchopneumonia, and lymphocytic depletion in lymphoid tissues were the most consistent lesions. Nonsuppurative encephalitis, sometimes with demyelination, was found occasionally. Coinfections with parasites (lice, worms) and bacteria (in particular Bordetella bronchiseptica) were common. 3, 7, 26, 29, 39, 40
Up to now, the pathology of seals stranding along the Dutch coast during the PDV outbreaks has not been described. Moreover, although the pathology of phocine distemper has been described in general, many aspects are poorly understood. For example, we do not know the effect of storage method or choice of tissue on the outcome for morbillivirus diagnostic assays; the effect of seal age on the character of the lesions or the outcome of diagnostic tests; the relationship between PDV infection and nutritional state; or the relationship between bacterial coinfections and the observed lesions.
The aim of this study was therefore to describe the pathology of PDV in harbor seals that stranded in The Netherlands in 2002, and to increase our understanding of the pathology of PDV infection in harbor seals in general by a quantitative approach to questions, such as those mentioned in the previous paragraph. We used a sample of 369 seals that were necropsied during the 2002 PDV epidemic in The Netherlands. Morbillivirus diagnostic assays were performed on these seals and effects of storage method and sample type assessed. Excluding the 137 non-confirmed cases, we then used the 232 confirmed cases for describing lesions, state of nutrition, and coinfections, and for determining correlations between assay results, lesions, age, and coinfections. A subset of 29 confirmed PDV cases was used for histologic analysis and a subset of 36 for bacteriologic analysis. The systematic collection of seal carcasses and the consistent necropsy procedure applied before and during the outbreak provided the opportunity to compare organ weights, nutritional state, and bacterial coinfections of PDV-infected seals with those of pre-epidemic seals that died of other causes.
Materials and Methods
The PDV epidemic sample
More than half of the 2,284 seals that stranded along the Dutch coast during the 2002 phocine distemper epidemic were necropsied. Most carcasses were stored in −20°C freezing containers placed on the Dutch Wadden Sea islands or the mainland for the duration of the epidemic. Several mass necropsy sessions were organized to examine these carcasses, subsequently referred to as “frozen seals.” In a minority of cases, necropsy was performed soon after death or euthanasia of the seal and these carcasses were stored at +4°C until necropsy. These are referred to as “cooled seals” in this paper.
Life history data were collected from all carcasses. Pathologic information and biological samples were collected almost exclusively from carcasses in which the organs had retained their shape and most of their color (fresh or mildly decomposed carcasses). Seals taken into rehabilitation before their death were excluded. The final number of seals from the 2002 PDV epidemic discussed in this paper consisted of 369 harbor seals (54 cooled, 315 frozen).
Diagnostic assays for morbillivirus
Samples from the carcasses of the 369 seals were tested by at least 1 and up to 4 assays to detect morbillivirus infection. The samples were taken as described in Kuiken and Baker (1991). 32
First, reverse transcriptase-polymerase chain reaction (RT-PCR) was performed to detect morbilliviral RNA in lung tissue of 361 (51 cooled, 310 frozen) seals, as described elsewhere. 27 Kidney, urinary bladder, and brain tissues were also tested in cooled seals, and kidney and urinary bladder tissues in a number of frozen seals. The samples were preserved in lysis buffer (6 M Guanidine-HCL, 10 mM Urea, 10 mM Tris-HCl, 20 % Triton-X100 (v/v), pH 4.4) and stored at −20°C or room temperature until analysis.
Second, an indirect avidin–biotin–complex immunohistochemistry (IHC) test was performed to detect morbillivirus antigen in formalin-fixed, paraffin-embedded tissues of 83 seals (38 cooled, 45 frozen), as described elsewhere. 16, 22, 29 The primary antibody used was polyclonal rabbit anti-measles virus serum. The tissues tested were lung, spleen, liver, kidney, urinary bladder for all seals, and additionally cerebrum and cerebellum for the cooled seals.
Third, serum samples of 82 seals (46 cooled, 36 frozen) were tested for morbillivirus-specific immunoglobulin M (IgM) antibody by an antibody-capture enzyme-linked immunosorbent assay (ELISA). Plates were coated with goat anti dog-IgM for capturing IgM, and horseradish-peroxidase–labeled CDV antigen was used to detect bound IgM. 27 Serum samples were centrifuged at 10,000 g for 5 minutes, and stored at −20°C until use.
Finally, the sera of 326 seals (53 cooled, 273 frozen) were tested for morbillivirus-specific immunoglobulin G (IgG) antibodies by indirect ELISA. 42 Plates coated with CDV antigen were incubated with seal serum and subsequently with horseradish-peroxidase–conjugated protein A to detect bound IgG. Sera were centrifuged and stored as previously described for IgM serology.
Though these assays were specific for morbillivirus rather than for PDV, it had previously been established that PDV was the morbillivirus causing the epidemic. 27 Cases positive in 1 or more of these 4 assays are referred to as “PDV cases” in this paper.
Necropsy findings in PDV cases
Necropsies followed a standard protocol. 32 Body length was measured as the straight distance from the nose tip to tail tip measured alongside the carcass with the carcass lying flat on the back. Blubber thickness was measured half-way along the length of the sternum. Pneumonia was called extensive when more than 50% of the lung volume was estimated to be firm. The lungs, liver, heart, and spleen of 35 cooled seals (17 males, 18 females) were weighed as were the kidneys, brain, and adrenals of 34 of these. For logistic reasons, organs were not weighed and brains were not examined in the frozen seals.
Formalin-fixed, paraffin-embedded samples of lung, spleen, kidney, urinary bladder, liver, cerebrum, cerebellum, adrenal, and occasionally other tissue sections of 29 cooled PDV cases were sectioned at 5 μm, stained with hematoxylin and eosin (HE) following standard procedures, and examined by light microscope. Additionally, lung tissue sections of 131 frozen PDV cases were stained (HE) and examined for numbers of neutrophils.
The age of a seal was determined by counting the number of cementum layers in 1 of the upper canine teeth. 19 Three age classes were used in the paper: <1-year-olds, 1- and 2-year-olds, and 3-year-olds and older. The <1-year-olds are pups, 1- and 2-year-olds are subadults, and 3-year-olds and older are mostly adults (F. Read, personal communication).
Tissues were screened for coinfections with phocine herpes virus 1, bacteria, worms, and lice (Echinophthirius horridus). To detect coinfection with phocine herpes virus 1, lung tissue samples preserved in lysis buffer of 169 frozen seals were tested by PCR. 35 Bacterial culture was performed on lung and tracheo-bronchial lymph node samples of 36 seals (26 cooled, 10 frozen); on liver, kidney, and spleen samples in 26 cooled seals; and on samples of other organs if gross lesions suggested that these were infected with bacteria. The samples were kept cool at +4°C until culture on aerobic blood agar, anaerobic blood agar, and Mac Conkey agar, and additionally on blood agar with increased CO2 for Brucella sp. In the 26 cooled seals, external iliac lymph node, and the uterus or testis samples were also cultured specifically for Brucella sp. Finally, lung-, heart- and stomach-worm burdens and numbers of lice were estimated and recorded as 1 of 4 levels (0; 1–10; 11–100; >100) in both cooled and frozen seals. Worms from cooled seals were fixed in 70% ethanol for identification. 25, 30, 52, 54
The harbor seals used for comparison with PDV cases: the non-PDV cases
Organ weights, state of nutrition, and bacteriology results of PDV cases were compared with those of nonepidemic seals stranded dead on the coast of The Netherlands. These are referred to as “non-PDV cases” in this paper.
For organ weights, the comparison group consisted of all 16 nonepidemic seals for which organ weights were available. The deaths of these 16 seals were associated with by-catch and other accidents (n = 6), mesenteric torsion (n = 2), lung- or heartworm infection (n = 2), bacterial infection (n = 3), or undetermined cause (n = 3). The weights of lung and heart were recorded for all 16 cases, liver and spleen in 15 of them, kidney in 12, adrenals in 8, and brain in 6.
For state of nutrition, the comparison group consisted of all 25 nonepidemic seals for which stomach contents and blubber thickness had been recorded and that had stranded in the same time period as the epidemic (16 June to 29 November) in 1 of the 2 years preceding the epidemic (2000 and 2001). The last 2 criteria were applied to minimize the influence of seasonal and between-year variation on the results. The deaths of the 25 seals were associated with by-catch (n = 5), foreign body (n = 2), mesenteric torsion (n = 2), lung- or heartworm infection or both (n = 3), bacterial infection (n = 4), or undetermined cause (n = 9).
For bacteriologic results, the comparison group consisted of all 8 seals that stranded in the time period detailed in the previous paragraph and from which specific bacteria had been cultured. The deaths of these 8 cases were associated with a flipper or prostate abscess (n = 3), by-catch (n = 2), lymphadenitis (n = 1), heartworm related hemorrhage (n = 1), or an undetermined cause (n = 1).
Statistics and models
Statistical tests were performed using Excel 2000 (MS Office 2000) and R. 44 All P values used were 2-sided, and P ≤ .05 was considered significant.
Models were fitted in R. The amount of variability in the response value explained by a model was calculated as the difference between the total deviance and the residual deviance, divided by the total deviance.
Results
Diagnosis of morbillivirus infection
Out of 369 seals, 232 (63%) tested positive for morbillivirus infection. The proportion of seals testing positive was 168/361 (47%) by RT-PCR, 45/83 (54%) by IHC, 30/82 (37%) by IgM serology, and 87/326 (27%) by IgG serology.
The remaining seals (137/369, 37%) tested negative by RT-PCR or other assays even though the gross lesions were usually consistent with PDV infection (data not shown). We had already minimized the negative effect of autolysis on assay results by largely excluding carcasses in a moderate or advanced state of autolysis from our study. Therefore, we hypothesized a negative effect of the process of freezing and thawing on assay results. However, analysis of our data did not support this hypothesis: significantly fewer cooled than frozen seals were positive in the lungs by RT-PCR (12/51 cooled seals positive vs. 150/310 frozen seals, X2 = 9.958, df = 1, P = .002), and no significant effect of storage conditions was observed in the other tests (IHC in lung sections: 11/38 cooled seals positive vs. 16/45 frozen seals, X2 = 0.164, df = 1, P = .685; IgM serology: 18/46 cooled seals positive vs. 12/36 frozen seals, X2 = 0.096, df = 1, P = .757; IgG serology: 19/53 cooled seals positive vs. 68/273 frozen seals, X2 = 2.185, df = 1, P = .139).
The types of tissue testing positive by RT-PCR differed among seals. Brain tissue was frequently positive by RT-PCR (Table 1) when lung tissue was negative, and vice versa (Table 2). Assuming that seals testing positive in brain tissue had neurologic signs, this indicates that a number of neurologic cases did not have detectable viral RNA in their lungs at the time of death. Additionally, although comparable proportions of IgM-positive (7/16) and IgM-negative (11/28) seals tested RT-PCR positive in 1 or more tissues (X2 = 0.250, df = 1, P = .617), the tissue distribution among these 2 groups differed significantly: IgM-positive seals were generally RT-PCR positive in the lung, while IgM-negative seals were generally RT-PCR positive in the brain (Table 2; Fisher exact test, P = .015).
Tissue distribution of morbilliviral RNA and morbilliviral antigen in phocine distemper virus-infected harbor seals stored either cooled or frozen.
In cerebrum; nt = not tested.
Comparison of diagnostic methods for morbillivirus infection in harbor seals infected with phocine distemper virus.∗
+ = positive; - = negative.
Cerebrum; nt = not tested.
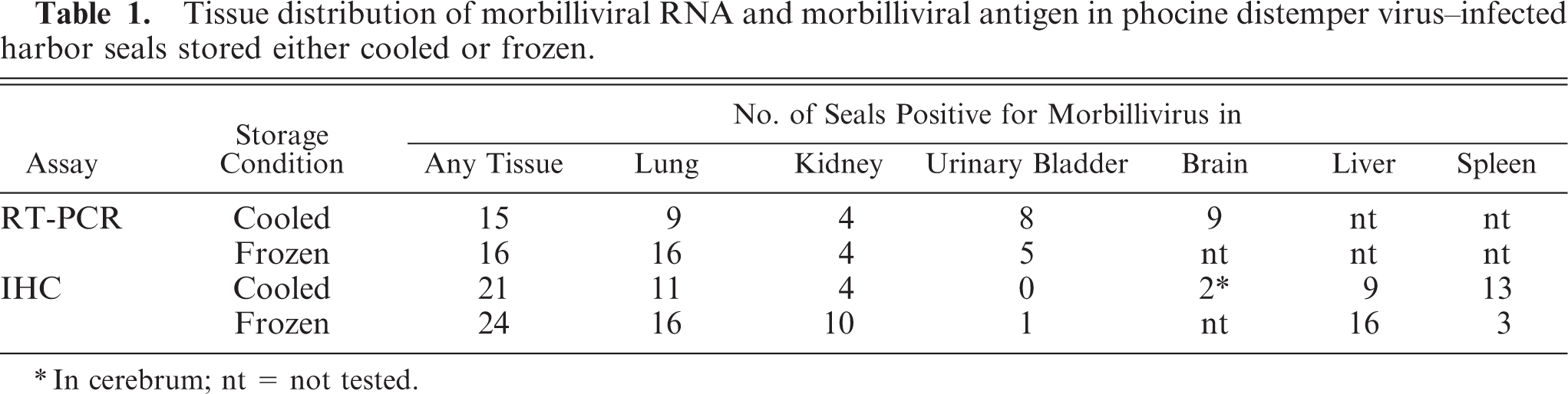
Morbilliviral antigen expression by IHC was most frequently detected in lung and liver (Table 1). The cell types expressing morbilliviral antigen were bronchial epithelial cells (Fig. 1), alveolar epithelial cells, alveolar macrophages, and syncytia in lung sections, bile duct epithelial cells in liver sections, mononuclear cells in spleen sections, pelvic epithelial cells in kidney sections, neurons and mononuclear cells in cerebrum sections, and transitional epithelial cells in urinary bladder sections. The tissue distribution of morbilliviral antigen expression by IHC often differed from that of viral RNA detected by RT-PCR (Table 2).
Lung; harbor seal No. 17. Morbillivirus antigen in bronchial epithelial cells. Immunoperoxidase histochemistry and hematoxylin, 400×.
Seals that had IgG to morbillivirus but were negative by IgM, RT-PCR, and IHC may in principle have been infected before the 2002 PDV epidemic. Because PDV occurs as an epizootic disease in harbor seals in the North Sea and the only recorded occurrence of PDV prior to 2002 in this population was in 1988, seals that were exclusively IgG-positive had to be older than 13 years to have been infected with PDV in 1988 rather than in 2002. For IgG-positive seals that were 13 years or younger, we assumed that they had been infected in 2002 even though we did not have paired serum to prove seroconversion. In total, age was determined in 190/232 PDV cases (82%), of which 2% (3/190) were older than 13 years. Only 1 of these, a 16-year-old female, was positive by IgG and negative by the other 3 tests, and could thus have been infected in 1988.
The proportion of PDV cases with IgG to morbillivirus increased significantly with age (3 levels: <1-year-old, 1- or 2-year-old, ≥3-year-old; Fisher exact test, n = 173, P = .003). The proportion of PDV cases with RT-PCR–positive lung tissue did not vary significantly with age, though it tended to decrease (Fisher exact test, n = 173, P = .218). The relation of IgM and IHC results to age could not be determined in this sample because too few seals were tested by these methods.
Main gross lesions
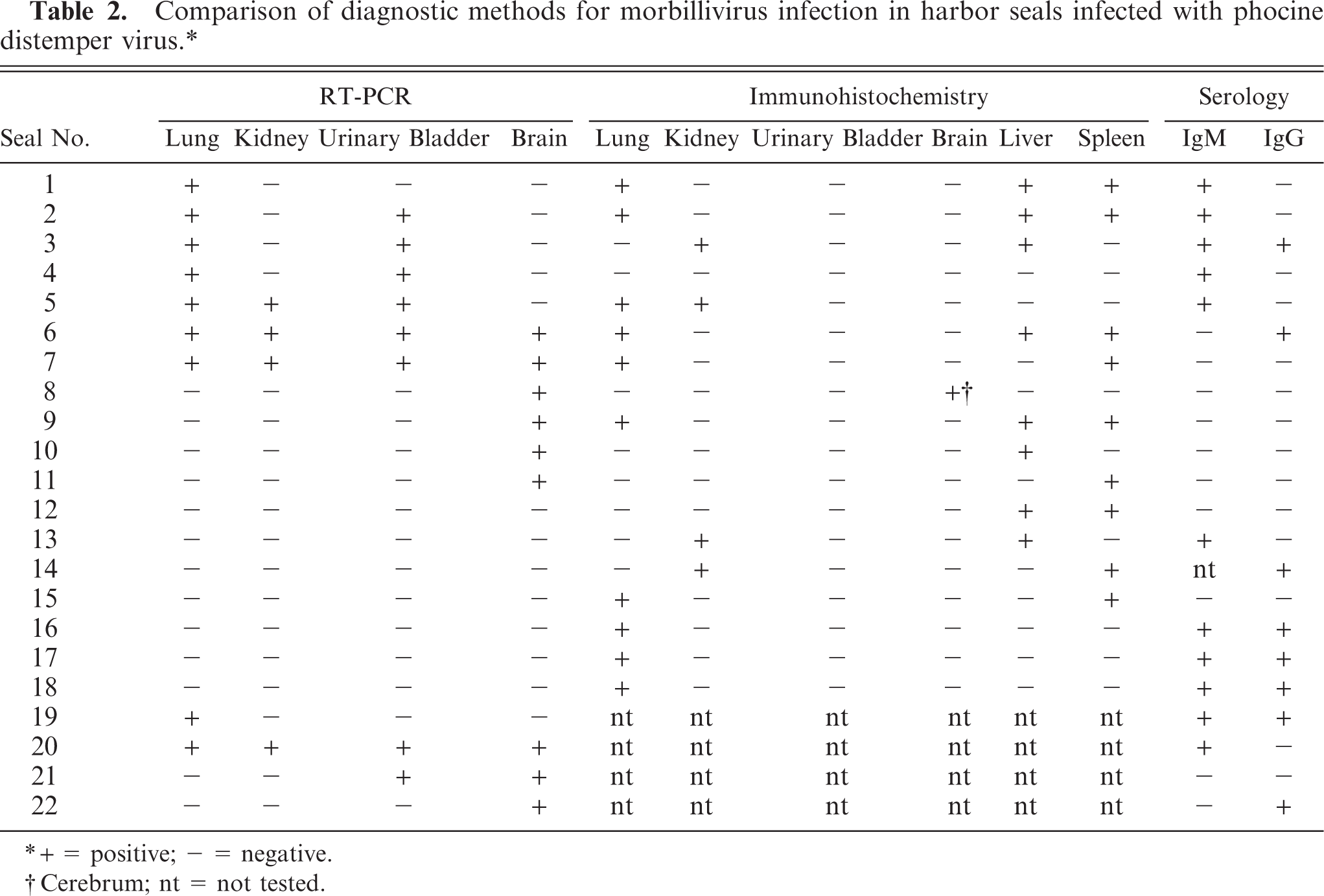
The main gross lesions were pneumonia and interstitial emphysema (Fig. 2). Pneumonia was characterized by raised consolidated areas that were bright red to dark-purple red. Both severity and distribution of pneumonia were determined in 210 of the 232 PDV cases. Pneumonia occurred in 198/210 cases (94%). The pneumonia was severe (174/198, 88%) or moderate (24/198, 12%), never mild. The distribution of the pneumonia was restricted to the ventral parts of the lungs in 69/198 (35%) cases (cranioventral, n = 30; caudoventral, n = 5; both, n = 34) and was more widespread in 129/198 (65%) cases (multifocal, n = 6; extensive, n = 123). In 12/210 (6%) cases there was no pneumonia: lungs were atelectatic (n = 1), edematous (n = 2), congested (n = 2), both edematous and congested (n = 2), or had no lesions (n = 5).
Thoracic cavity; harbor seal No. 23. Pulmonary and mediastinal emphysema; histologically, there was severe broncho-interstital pneumonia, associated with combined phocine distemper virus and Streptococcus equi subsp. zooepidemicus infections.
The presence and distribution of emphysema were determined in 226/232 cases. The vast majority of PDV cases had emphysema (195/226, 86%). Emphysema was pulmonary in 85/195 (44%; Fig. 2), mediastinal in 178/195 (91%; Fig. 2), pericardial in 152/195 (78%), retro-peritoneal in 102/195 (52%), and subcutaneous in 132/195 (68%). Retro-pharyngeal emphysema was observed in 2 cases. The distribution of the emphysema was variable. The most frequent distribution (46/226, 20%) was a combination of mediastinal, pericardial, retroperitoneal, and subcutaneous emphysema. Emphysema was rarely limited to the lungs (8/195, 4%).
Emphysema occurred significantly more often in cases with pneumonia than in those without (Fisher exact test, n = 210, P < .001). However, the extent of emphysema (3 levels: absent, thoracic only, thoracic and extra-thoracic) did not vary significantly with the severity of pneumonia (2 levels: moderate, severe; Fisher exact test, n = 198, P = .07), or with the distribution of pneumonia (2 levels: ventral only, throughout the lungs; X2 = 1.015, n = 198, df = 2, P = .603).
Age (3 levels: <1-year-old, 1- or 2-year-old, ≥3-year-old) had a significant effect on the extent of emphysema (Fisher exact test, n = 178, P ≤ .001), with extra-thoracic emphysema occurring only very rarely in <1-year-olds while being very common in older seals (Fig. 3). Seal age also had a significant effect on the severity of the pneumonia (3 levels: absent, moderate, severe; Fisher exact test, n = 175, P = .023) and on the distribution of pneumonia (3 levels: absent, ventral only, throughout the lungs; Fisher exact test, n = 175, P = .038). In particular, when pneumonia occurred in <1-year-olds, it was always severe and mostly extensive (Fig. 3).
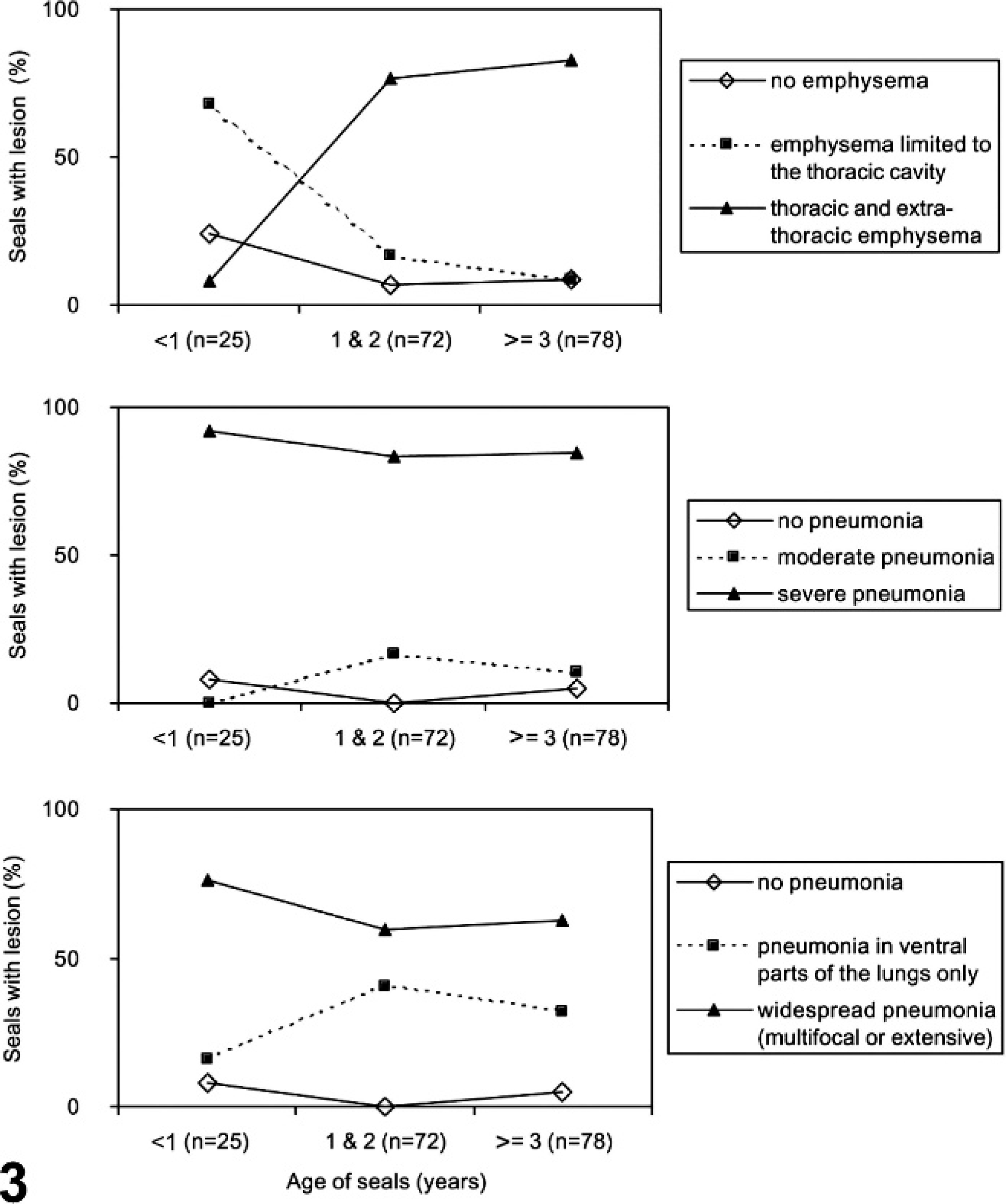
The relationship between age and extent of emphysema, severity of pneumonia, and distribution of pneumonia in harbor seals infected with phocine distemper virus.
Other gross lesions
There were 21/232 (9%) PDV cases with other respiratory tract or thoracic cavity lesions: suppurative bronchitis (12/232, 5%), pulmonary abscesses (3/232, 1%), hemopurulent pleuritis (1/232, <1%), fibrinous pleuritis (1/232, <1%), pulmonary infarct (1/232, <1%), pulmonary hematoma (1/232, <1%), and hydrothorax (2/232, 1%). Of these cases, 19/19 (100%) also had pneumonia, 17/21 (81%) also had emphysema, 12/19 (63%) were RT-PCR positive in the lungs, and 9/21 (43%) were IgG positive.
There were 16/232 (7%) cases that had other significant gross lesions in other organ systems. These were purulent cystitis (1/232, <1%), otitis externa (2/232, 1%), phlegmon and fasciitis (1/232, <1%), hepatic atrophy possibly caused by porto-caval shunt (1/232, <1%), intestinal volvulus (2/232, 1%), perforation of internal organs by corpus alienum (2/232, 1%). Lymphadenopathy occurred in 6/232 cases (3%), affecting multiple lymph nodes (n = 4) or single nonthoracic lymph nodes (n = 2). In addition there was a neonate that had starved. Of these cases, 8/14 (58%) had pneumonia, 6/14 (43%) had emphysema, 4/16 (25%) were RT-PCR positive in the lungs, and 12/15 (80%) were IgG positive. Compared with the remainder of the PDV cases, these cases had significantly less pneumonia and emphysema (Fisher exact test, P ≤ .001), were significantly less RT-PCR positive (Fisher exact test, P ≤ .001) and significantly more IgG positive (Fisher exact test, P = .002).
Microscopic lesions
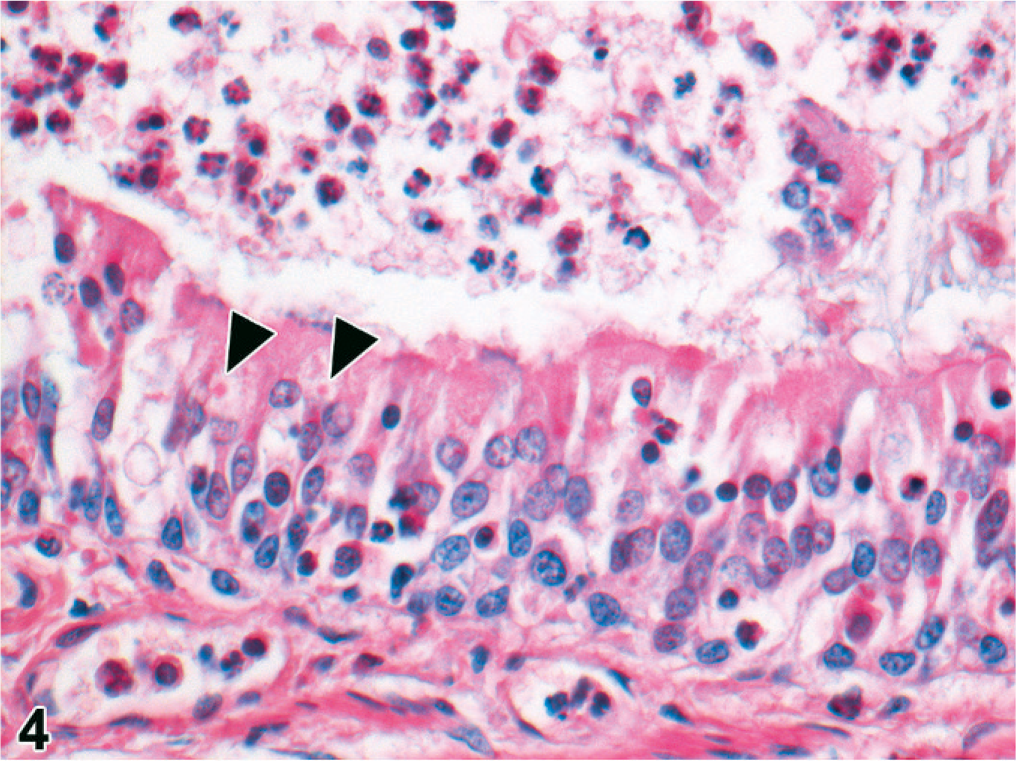
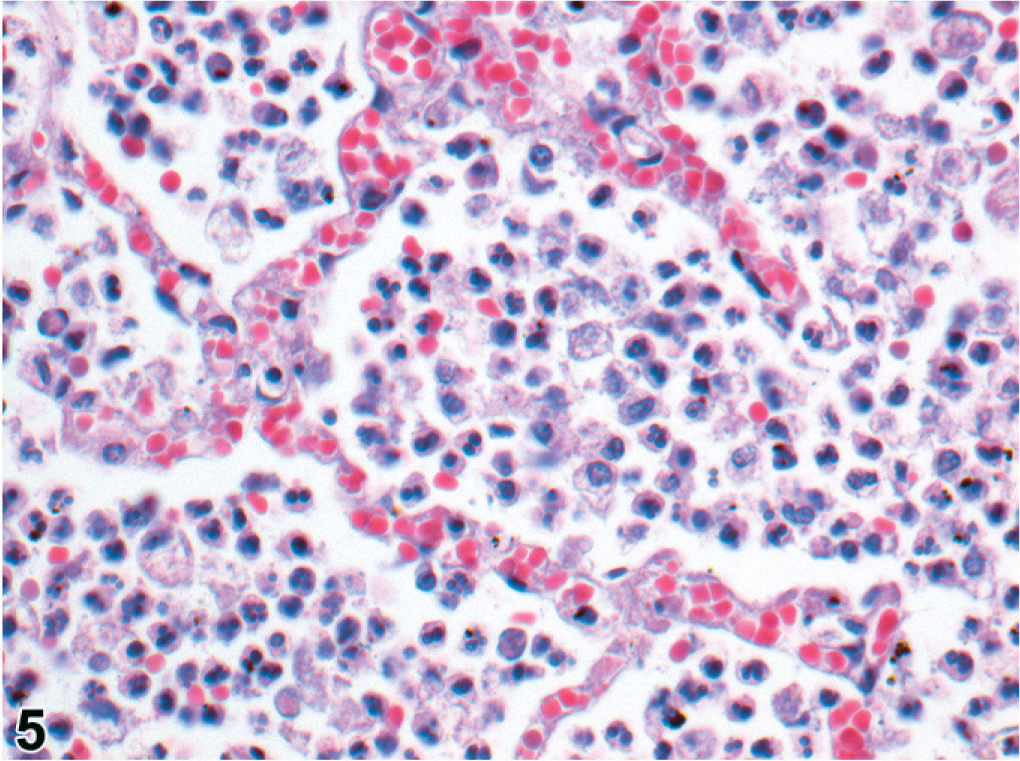
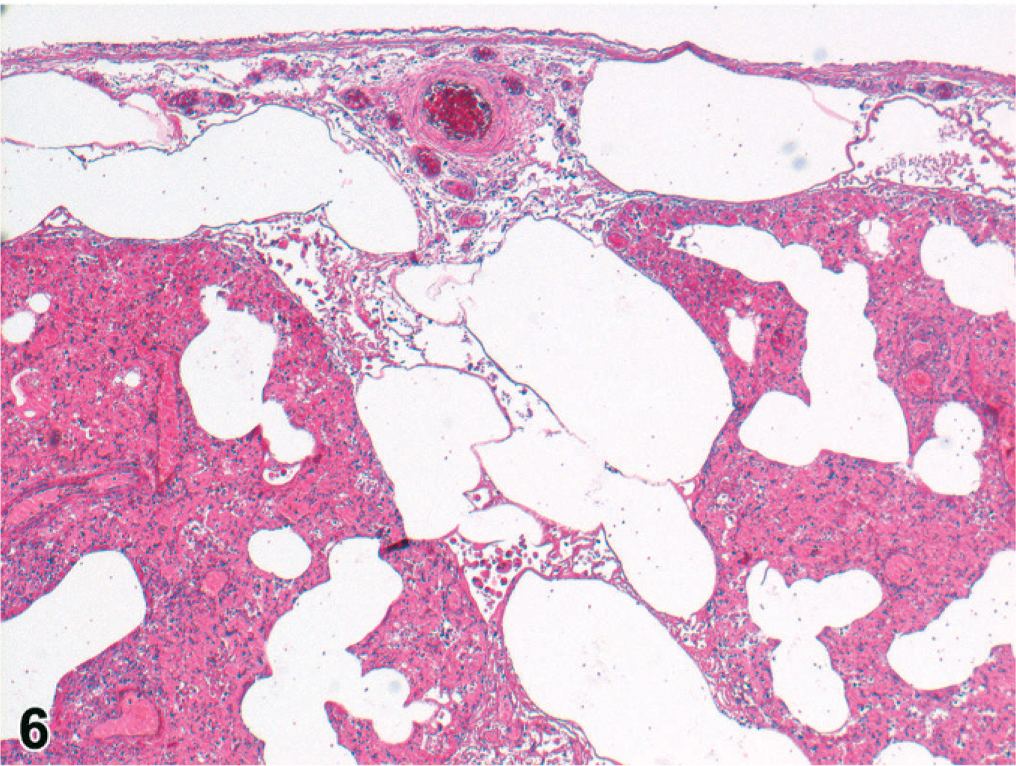
The 29 cooled seals examined histologically were 6 <1-year-olds, 9 1- or 2-year-olds, 10 ≥3-year-olds, and 4 of undetermined age. Microscopically, pneumonia occurred in 25/29 (86%) of the cooled PDV cases, characterized as bronchopneumonia (15/25, 60%) or broncho-interstitial pneumonia (10/25, 40%). Bronchopneumonia was generally marked, subacute, diffuse, and suppurative (Figs. 4, 5). In 2/14 cases, there were distinct, round, 3- to 10-μm-diameter, intracytoplasmic and intranuclear inclusion bodies in bronchial epithelium (Fig. 4); in 4/14 cases, there was pulmonary emphysema (alveolar, interlobular, subpleural, or combinations of these) (Fig. 6). Broncho-interstitial pneumonia was generally moderate, chronic, multifocal, and histiocytic (Fig. 7). Worms (probably Parafilaroides gymnurus) were observed in the alveoli and bronchiolar lumina of 12/29 (41%) seals, all 0- or 1-year-olds. Diffuse acute moderate to severe congestion of the lung was nearly always observed (28/29, 97%; Figs. 5–7).
Lung; harbor seal No. 6. Seal with suppurative broncho pneumonia associated with combined phocine distemper virus and Bordetella bronchiseptica infections. There are many neutrophils in the lumen and fewer among the bronchial epithelial cells. Note the eosinophilic intracytoplasmic inclusion bodies, characteristic for morbillivirus infection, in several bronchial epithelial cells. HE, 400×.
Lung; harbor seal No. 3. Seal with suppurative bronchopneumonia associated with combined phocine distemper virus and Bordetella bronchiseptica infections. Neutrophils fill the alveolar lumina. HE, 400×.
Lung; harbor seal No. 24. Seal with marked suppurative bronchopneumonia associated with combined phocine distemper virus and Bordetella bronchiseptica infections. There is alveolar, interstitial and subpleural emphysema, and marked pulmonary congestion. HE, 100×.
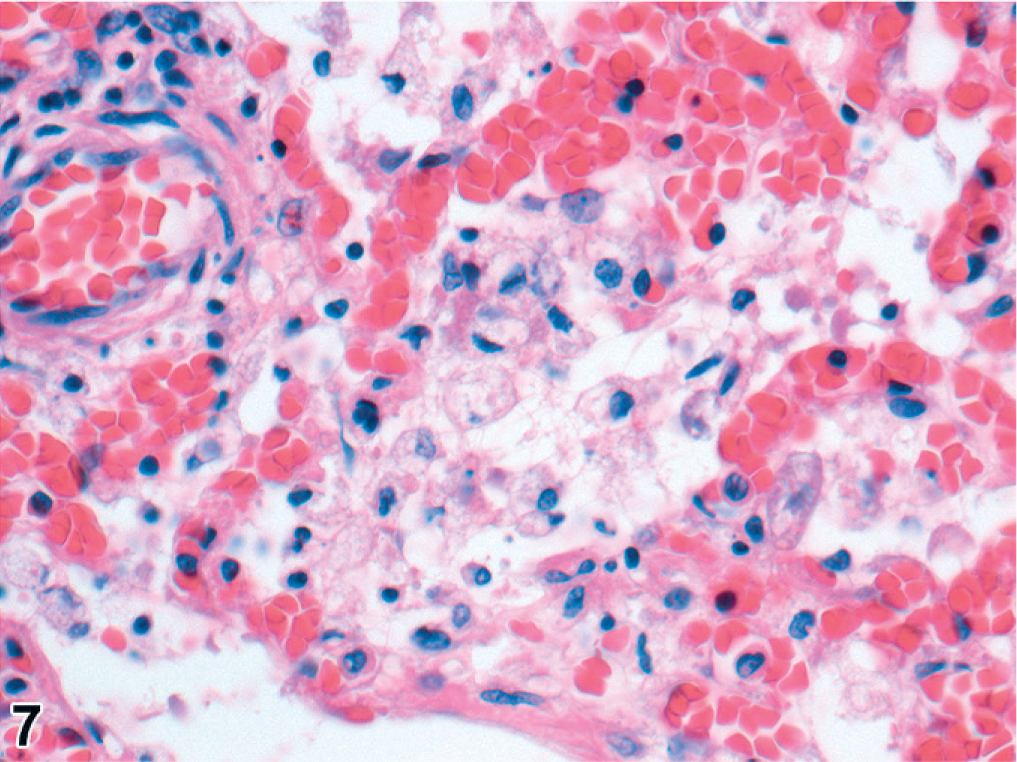
Lung; harbor seal No. 23. Histiocytic broncho-interstitial pneumonia, associated with combined phocine distemper virus and Streptococcus equi subsp. zooepidemicus infections, with marked congestion. HE, 400×.
The 2 above-mentioned cases with inclusion bodies in bronchial epithelium also had intracytoplasmic inclusion bodies in the urinary bladder epithelium, and in 1 case also in renal pelvic epithelium and bile duct epithelium. The latter case had a mild necrotizing cystitis, pyelitis, and cholangitis. Splenic lymphoid depletion occurred in 5/29 (17%) cases, all ≥3 years old, splenic lymphoid hyperplasia in 1 case (1/29, 3%), and extramedullary erythropoiesis in 1 case (1/29, 3%). One seal (1/29, 3%) had marked adrenocortical atrophy. Another (1/29, 3%) had multifocal adrenocortical necrosis and multifocal necrotizing hepatitis. No lesions were observed in cerebrum or cerebellum.
The lung sections of 52/131 frozen PDV cases were interpretable. High to very high numbers of neutrophils occurred in 29/52 (56%) of these, corresponding with moderate to marked suppurative bronchopneumonia. These neutrophils were located in bronchi and sub-bronchial glands, alveoli and sometimes in interlobular spaces.
Organ weights
The consolidated lungs frequently felt heavy. To test whether PDV infection had a significant effect on the weight of lungs or other organs, we fitted 2 simple models using all the organ weight data available (PDV cases and non-PDV cases together). The weights of the 2 units of paired organs were summed and considered as 1. Organ weight generally increases with body size (growth), so the first model we created regressed organ weight (response variable) over body length (predictor). The correlation between organ weight and body length was positive for all organs examined, and the amount of variation in organ weight explained by this model was 68% for heart, 61% for kidneys, 55% for both lungs and liver, 48% for adrenals, 40% for brain, and 17% for spleen. By analysis of variance (ANOVA), we then compared this first model to a second model in which both body length and having PDV or not (PDV, 2 levels: 0 = other cause of death, 1 = PDV-case) were the predictors of organ weight. Adding PDV did not significantly improve the prediction of brain, heart, liver and spleen weight. However, it did significantly improve the model for lung, adrenal, and kidney weight (lungs, F = 16.791, P < .001, 67% lung weight variability explained; adrenals, F = 14.231, P < .001, 62% adrenal weight variability explained; kidneys, F = 7.561, P = .008, 67% kidney weight variability explained). Both predictors in these second models were positively correlated with the organ weights, indicating that PDV significantly increased the weight of lungs and adrenals and to lesser extent that of kidneys (model details not shown). The plotted data showed that this effect of PDV is most manifest in larger and therefore older seals (Fig. 8).
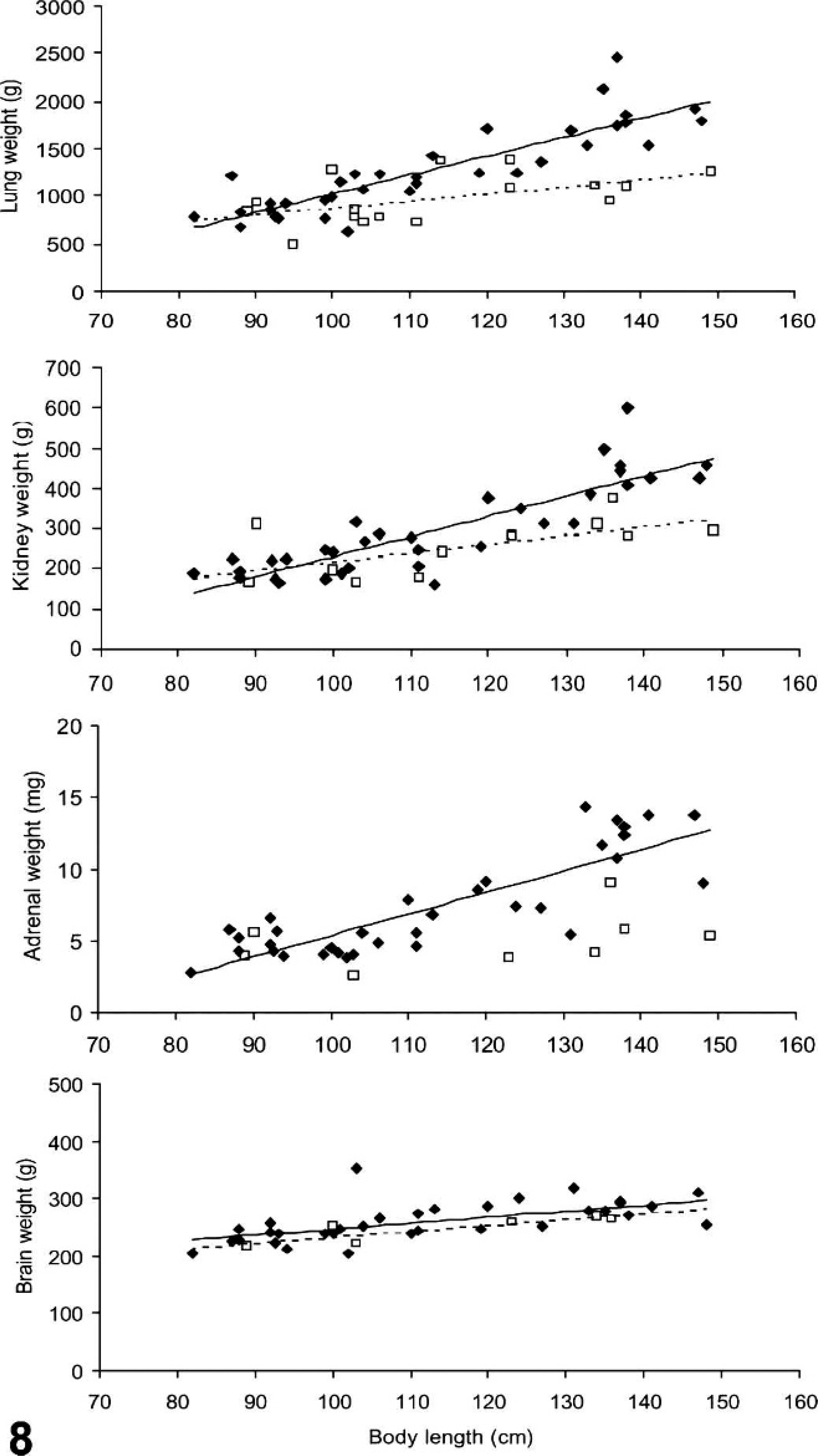
Weight of lungs, kidneys, adrenals, and brain of harbor seals infected with phocine distemper virus (closed diamonds) and noninfected seals (open squares) regressed over seal body length (cm) shows that the first 3 organs are heavier in infected seals, in particular the longer (older) seals.
We hypothesized that suppurative bronchopneumonia would result in heavier lungs. However, there was no association between moderate or high numbers of neutrophils and lung weight in PDV cases (the ratio lung weight over body length compared between 18 PDV cases with high numbers of neutrophils and 13 PDV cases with no or few neutrophils; 2-sample t-test assuming equal variances, P = .964).
State of nutrition
Stomach contents were recorded for 210 PDV cases. The majority had empty stomachs (195/210, 93%). The remaining 15 had 1 or more corpora aliena (9/210, 4%), fish bones (3/210, 1%), or fish (i.e., a recent meal, 3/210, 1%). The corpora aliena were pieces of plastic (n = 3), wire booms (n = 2, 1 spring boom and 1 French boom, both parts of pater noster rigs used in sea angling), a spring (n = 1), an iron thread (n = 1), foil (n = 1), a string (n = 1), stones (n = 2), and a piece of wood (n = 1). The 3 seals with fish in their stomach were 0, 3, and 16 years of age. The <1-year-old was positive by RT-PCR in the brain and bladder, the 3-year-old was positive by RT-PCR in the lungs (only tissue tested), and the 16-year-old was the PDV-case only positive for IgG as previously mentioned.
For comparison, the 7 non-PDV cases whose deaths were associated with worm or bacterial infections, and 1 mesenteric torsion case, had empty stomachs. The 2 cases whose deaths were associated with corpora aliena and the other mesenteric torsion case had corpora aliena. The 5 cases whose deaths were associated with by-catch had fish in their stomachs. In the 9 cases for which the cause of death was undetermined, any of these 3 stomach content situations was found.
The average blubber thickness of PDV cases (n = 231) was 13.5 mm (range 0 to 26 mm). It differed significantly from that of the non-PDV cases whose deaths were associated with by-catch (31.8 mm, range 15 to 45 mm; n = 5; Mann-Whitney test, P = .001), and seemed more comparable with those of non-PDV cases that died from other causes than by-catch.
Blubber thickness increases with age and changes seasonally with molt and reproductive activities. 23, 50, 64 Therefore, we regressed blubber thickness (mm) over age (years) and then over date of stranding, the latter also for different sex-age classes (female <1-year-olds, female 1- to 2-year-olds, female ≥3-year-olds; male <1-year-olds, male 1- to 2-year-olds, male ≥3-year-olds). There was a significant trend for blubber thickness to decrease with age (r2 = 0.039, n = 189, P = .007). Blubber thickness also decreased significantly over time during the epidemic (r2 = 0.089, n = 189, P < .001) among <1-year-olds (female, y = −0.174x + 61, r2 = 0.429, n = 13, P = .015; male, y = −0.145x + 50, r2 = 0.445, n = 15, P = .007) and seals ≥3-year-olds (female, y = −0.141x + 46, r2 = 0.299, n = 35, P ≤ .001; male, y = −0.141x + 46, r2 = 0.185, n = 49, P = .002). There was no effect of date of stranding on blubber thickness of 1- or 2-year-olds (females, r2 = 0.004, n = 47, P = .689; males, r2 = 0.003, n = 30, P = .950).
Phocine herpes virus 1 coinfection
Nine of 169 (5%) PDV cases tested were positive by PCR for phocine herpes virus 1 infection. There were 5 females (a 3- and a 4-year-old, 2 1-year-olds, and 2 100-cm-long female) and 4 males (a 5-year-old, 2 4-year-olds, and a 3-year-old).
Bacterial coinfections
Fourteen bacterial genera or species were cultured from the PDV cases examined (Table 3, Fig. 9). B. bronchiseptica was the most frequently isolated bacterium (24/36, 66%). It was always cultured from lung tissue (24/24) and sometimes additionally from other tissues examined (15/24; Table 3). The seals with B. bronchiseptica infection were <1-year-old (n = 5), 1-year-old (n = 5), 3 years or older (n = 6), or of undetermined age (n = 7). In 8/36 (22%) cases, B. bronchiseptica was cultured from the lungs (and other organs) in association with high numbers of other bacteria; in 4/36 (11%) cases, it was the only specific bacterium cultured from the lungs; and in 12/36 (33%) cases, it was the only specific bacterium cultured from all organs examined.
Tissue distribution of bacteria isolated from harbor seals infected with phocine distemper virus.∗
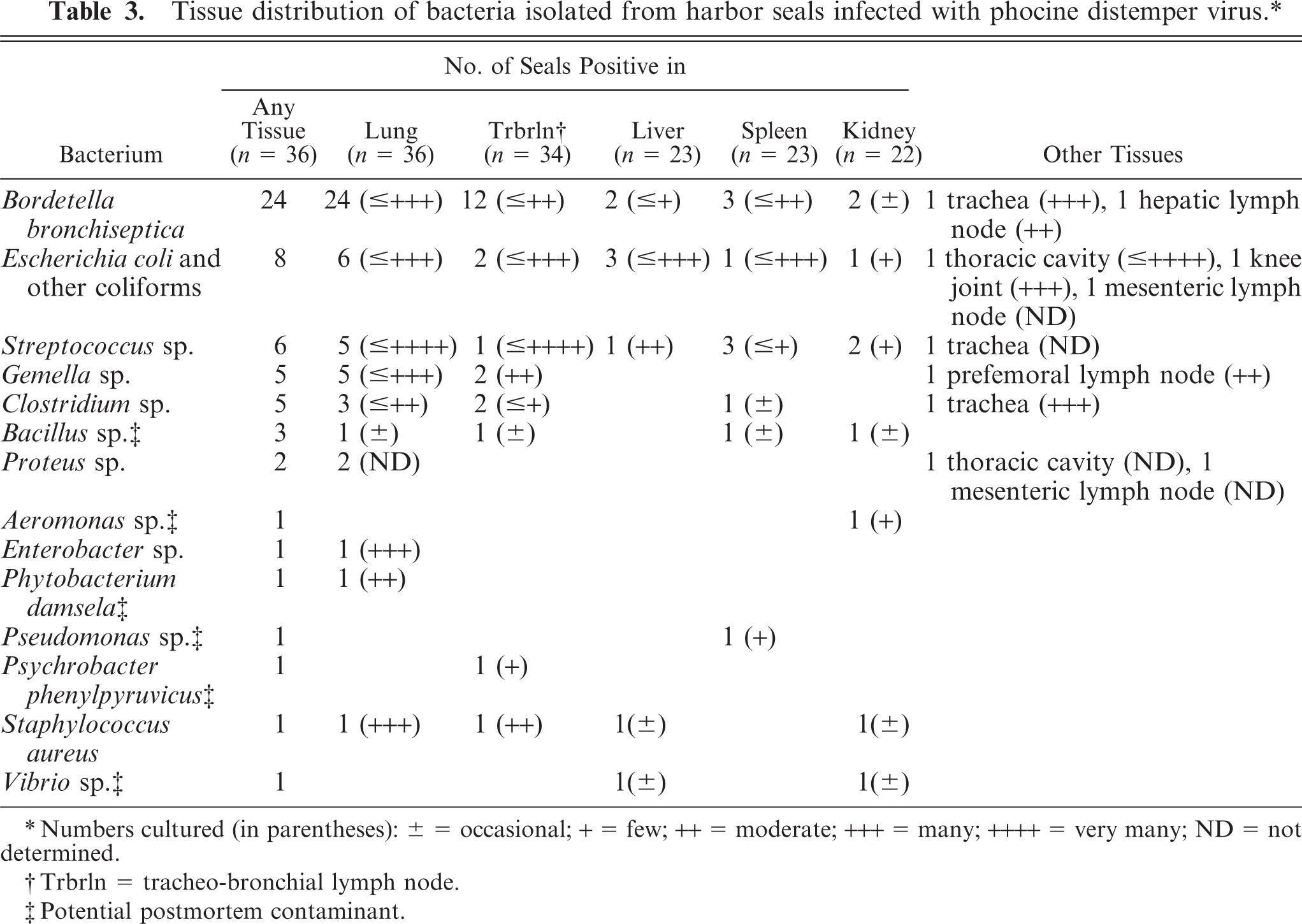
Numbers cultured (in parentheses): ± = occasional; + = few; ++ = moderate; +++ = many; ++++ = very many; ND = not determined.
Trbrln = tracheo-bronchial lymph node.
Potential postmortem contaminant.
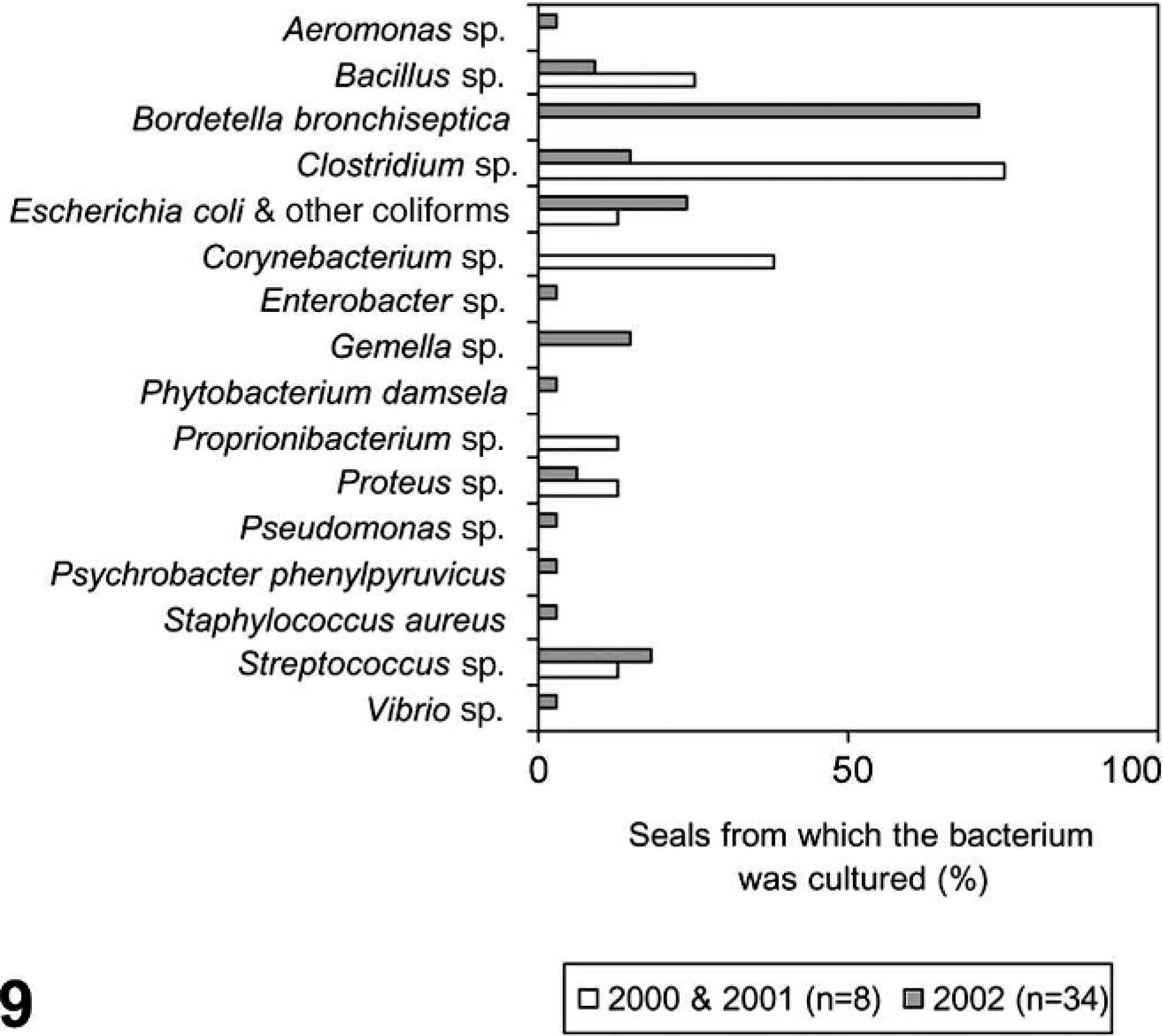
Specific bacteria isolated from harbor seals infected with phocine distemper virus (n = 34) during 2002 (gray bar) compared with bacteria isolated from noninfected seals (n = 8) over the same time period in the 2 previous years (white bar).
In the remaining 12/36 (34%) cases, B. bronchiseptica was absent. In these cases, other specific bacteria were isolated in significant numbers in lungs and other organs (5/36, 14%, including a case of Staphylococcus aureus septicemia), in small or undetermined numbers in one or few organs (5/12, 14%), or were absent (2/36 unspecified mixed cultures, 6%).
Six PDV cases with other gross lesions were among those from which bacteria were cultured: 2 bronchopneumonia cases (B. bronchiseptica with Streptococcus equisimilis or with Clostridium perfringens, from respiratory tract); a pulmonary abscess case (S. equi subsp. zooepidemicus, from multiple organs); a pulmonary hematoma case (B. bronchiseptica with S. zooepidemicus, both from multiple organs); a generalized lymphadenopathy case (Gemella sp., from lung and multiple lymph nodes), and the hepatic atrophy case (Escherichia coli, from lung, tracheo-bronchial lymph node and liver; Enterobacter sp. from lungs, Aeromonas sp. from kidney).
When B. bronchiseptica was the only specific bacterium isolated from the tissues examined, there was a straightforward relationship with the number of neutrophils in lung sections: there were no or few neutrophils when B. bronchiseptica was isolated from lung tissue only; there were moderate to large numbers of neutrophils when B. bronchiseptica was isolated from the lungs and other tissues. However when other bacteria were isolated in moderate to large numbers from the lungs, with or without B. bronchiseptica, no consistent pattern was detected with regard to neutrophil numbers.
The species of bacteria isolated from the PDV cases in 2002 only partially overlapped with those isolated from non-PDV cases in 2000–2001 (Fig. 9). Notably, B. bronchiseptica was not isolated from the non-PDV cases. In contrast, Corynebacterium sp. was not isolated from the PDV cases, whereas it was isolated from 3/8 non-PDV cases where it was twice associated with cause of death. Brucella sp. was not isolated.
Parasitic coinfections
Macroscopically, lungworms (identified as Otostrongylus circumlitus and Parafilaroides gymnurus) were observed in 25/226 (11%) of the PDV cases, heartworms (identified as Acanthotheilonema spirocauda) in 42/221 (19%), and stomach worms (identified as Pseudoterranova decipiens and Contracaecum osculatum) in 150/227 (66%). The frequency distributions of the parasites were highly aggregated as expected for macroparasitic infections (Fig. 10). 65 The distributions of lung- and heartworms were quite comparable, and different from that of stomach worms (Fig. 10).
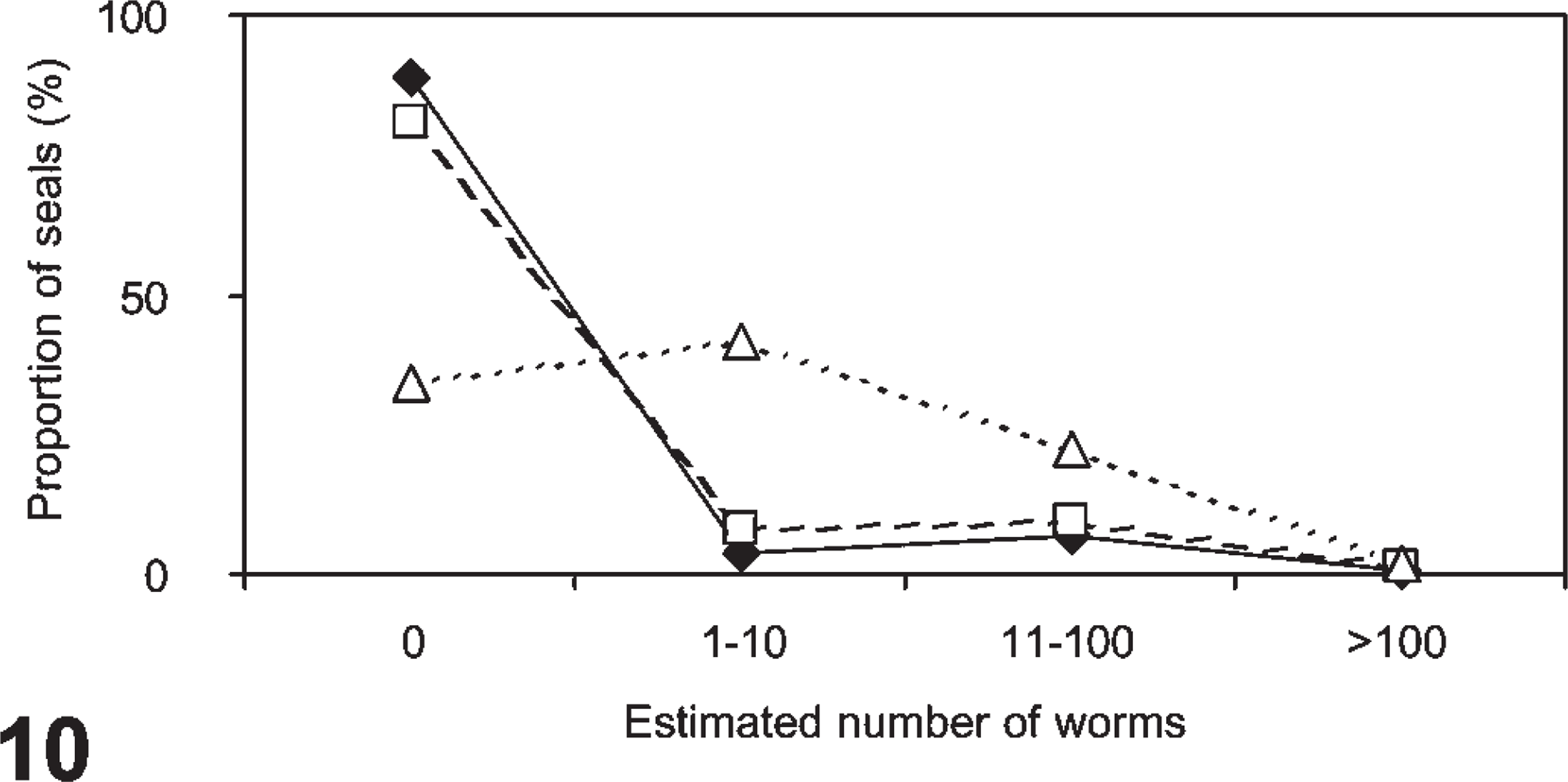
The frequency distributions of worm burdens in harbor seals infected with phocine distemper virus (lungworms, closed diamonds, n of seals = 226; heartworms open squares, n of seals = 221; stomach worms, open triangles, n of seals = 227).
All 3 worm burdens and seal age were determined for 179/232 seals. Lungworms were observed in 21 of these, heartworms in 28 and stomach worms in 150. The first <1-year-old in this sample stranded on 14 August 2002, the first <1-year-old with lungworms on 29 August, with heartworms on 27 August, and with stomach worms on 8 September 2002. Lungworms were observed most frequently and with the highest burdens in <1-year-olds, and heartworms most frequently and with the highest burdens in 1- and 2-year-olds (Fig. 11). Stomach worms were seen in all age categories, but the highest burdens occurred in seals ≥3 years old (Fig. 11). Stomach worm burden (4 levels: no worms; 1–10 worms; 11–100 worms; >100 worms) increased significantly with age in males but not in females (males, r2 = 0.172, n = 87, P < .001; females, r2 = 0.007, n = 92, P = .420).
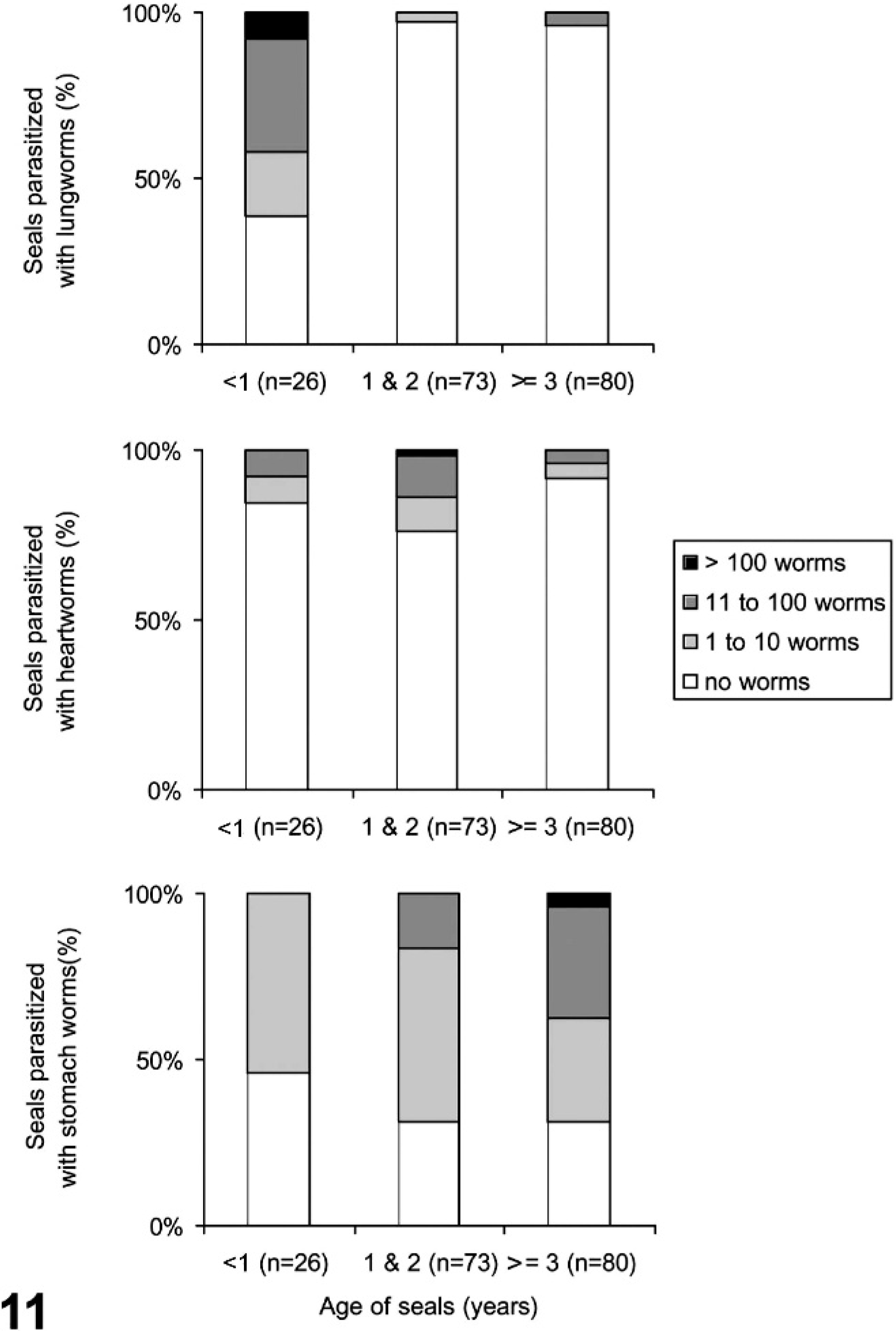
The distribution of lungworm, heartworm and stomach worm burdens in harbor seals infected with phocine distemper virus, classified according to age.
The presence and number of lice were reported in 212/232 cases. Lice were present in 29/212 (14%) of the cases (16 cases with 1–10 lice, 10 with 11–100 lice, and 3 with ≥100 lice). Lice were most frequent and with the highest burdens in 1- and 2-year-olds. However, lice can drop off carcasses once the seal is dead, and therefore this may be an underestimation of the number of seals with lice as well as of louse numbers.
Discussion
Correlating pathology findings and related laboratory diagnostic analyses in PDV cases and comparing these with those of non-PDV cases have provided further insight into the pathology of PDV infection in free-living harbor seals. Because our study was limited to seals found dead or moribund, these findings are not necessarily representative of the disease process as it may occur within the overall population. Also, although levels of contaminants occurring in seals may be immunosuppressive and therefore affect the disease process, we did not investigate this during the current study. 17 We sequentially discuss a number of points raised by the PDV assay results, the lesions, and lastly the coinfections.
Analysis of the PDV assay results showed differential temporal presence of virus in lung and brain, and age-related differences in disease course.
First, the results suggested differential periods of presence of virus in lungs and brains. Clearance of virus from host tissues occurs in morbillivirus infections, but clearance from the brain is not efficient in canine distemper cases with brain involvement in dogs. 1, 21, 43, 48 Such cases also tend to have a longer disease course than average. 1 In this study, seals positive by RT-PCR in the brains tended not to be positive by RT-PCR in the lungs, which would fit with an efficient virus clearance from most tissues including the lungs, but not from the brains. These results indicate that both lung tissue and brain tissue need to be tested by RT-PCR when PDV is suspected.
Second, associating PDV assay results to seal age provided support for age-related differences in disease course. Indeed, the proportion of seals with IgG antibodies increased significantly with age. There was also an indication for such a trend for IgG in seals infected during the 1988 epidemic, and evidence for such a trend for neutralizing antibodies. 12, 57 For the latter, 4 possible explanations were discussed. 57 The first was that juvenile seals were less exposed to the virus; this is inconsistent with the PCR-results in the present study. The second was that older seals would have more anamnestic responses; this is inconsistent with the present study because only a few dead adult seals were old enough to have been alive during the 1988 epidemic. The last 2 explanations were greater mortality among young seals and age-related compromised immune responses in young seals; these are not inconsistent with the results of our study. A variant of the latter is a lengthier disease course in older seals.
Analysis of PDV-associated lesions showed influences of the seal's anatomy and physiology on the development of emphysema, an effect of age on disease course, evidence of dehydration, an effect of age on blubber thickness, and different disease characteristics in cases with severe gross nonrespiratory tract lesions. First, emphysema is likely to develop as a result of inflammation and airway obstruction, a concept supported in this study by the positive correlation between the occurrence of pneumonia and emphysema in the PDV cases. Pneumonia is a frequent feature of morbillivirus infections in all host species. However, to our knowledge, emphysema has been associated with morbillivirus infection in seals and cattle but not in dogs. 1, 7, 26, 28, 33, 36, 39 A possible explanation is that seals and cattle—but not dogs—have well-developed interlobular septa that prevent collateral ventilation in case of airway obstruction, resulting in interstitial emphysema. 5, 13, 20, 47, 53, 62
Second, linking pneumonia and emphysema to seal age suggested age-related differences in the development of lesions. In particular, the extent of emphysema varied significantly with age class rather than with severity and distribution of pneumonia: most <1-year-olds had severe and extensive pneumonia, but rarely had extra-thoracic emphysema. In contrast, extra-thoracic emphysema in older seals was common regardless of the extent and severity of the pneumonia. A possible explanation for this difference is a lengthier disease course in older seals. This may be analogous to cattle with protracted interstitial pneumonia, where emphysema also spreads from the lungs to the mediastinum and the skin of the back. 13 Another explanation could be that physiological diving responses and oxygen storage capacity are not fully developed in <1-year-olds, so <1-year-olds perform shorter dives than older seals. 8, 10 Normally pinnipeds exhale before diving, and the air that remains in the alveoli is squeezed out and into the bronchi and trachea as pressure causes the lungs collapse. 8, 31 If the airways are obstructed, as in seals with PDV infection, air in the alveoli might rupture the alveolar walls during long dives, resulting in emphysema.
Third, the effect of PDV on nutritional state and organ weights suggests a role for dehydration. The empty stomachs of PDV cases indicate that feeding and associated water intake from prey was impeded. Further evidence for anorexia was that blubber reserves of PDV cases were on average reduced compared to those of by-caught seals (this study) or culled seals. 23 Presumably PDV cases have higher water requirements than normal, because they excrete large amounts of fluid via oro-nasal and lacrimal secretions and diarrhea. 28, 40, 41 Therefore, it is likely difficult for these seals to maintain their water and electrolyte balances. The increased kidney and adrenal weights that we observed in PDV cases fit with dehydration. Water-deprived rodents also show significantly increased renal and adrenal weights, possibly caused by the hypertrophy of the renal medulla. 2, 9, 49 Future studies of stranded seals might address the question of dehydration through measurement of urea nitrogen in vitreous humor fluid. 34
Fourth, we next found that blubber thickness of the PDV-infected seals decreased with age, and with date of stranding in seals ≥3 years old and in <1-year-olds. In seals ≥3 years old, these results suggest either lengthier disease or coincidence with physiological fat loss (e.g., because of reproductive activities), or a combination of these. 11, 14 In <1-year-olds, postweaning fat loss seems a more plausible explanation for the observed time trend than an effect of maternal antibodies on disease duration because antibodies to PDV were virtually absent in the population. 27, 38, 55
Fifth, PDV cases with macroscopic evidence of significant nonrespiratory tract lesions were significantly more RT-PCR negative and IgG positive than the other PDV cases, and fewer had pneumonia and emphysema. These findings suggest these PDV cases had a different and lengthier disease course than most PDV cases.
Analysis of coinfections showed that bacterial pneumonia occurred frequently and provided evidence for an immunosuppressive effect of PDV. First, suppurative pneumonia indicative of bacterial coinfection was commonly diagnosed by histologic analysis of lung tissues. This was surprising, because macroscopic evidence of suppurative pneumonia was rare. A possible explanation for this discrepancy might be that a high proportion of the bacterial infections in the lung were peracute to acute and had not yet led to macroscopically visible changes.
Second, the indications for immunosuppression were the frequent presence of B. bronchiseptica in the lungs, and the occurrence of lungworm infections in older seals. B. bronchiseptica is often isolated from lungs of seals and other carnivores during PDV or canine distemper virus (CDV) epidemics, but not outside these epidemics. 4, 26, 33, 39, 51 In dogs B. bronchiseptica is often chronically present in the upper respiratory tract, infection occurring early in life and followed by a certain resistance to severe disease. 6, 66 It is thought that when hosts are immunosuppressed, the bacterium can invade and colonize the lower respiratory tract and cause bronchopneumonia. The same principle applies for lungworms, as these tend to infect and cause disease in harbor seals early in life, after which surviving hosts usually have a certain degree of immunity to reinfection and disease. 37
Footnotes
Acknowledgements
We are grateful to the people who helped with the collection and necropsy of the seal carcasses stranding along the Dutch coast. We also thank Barry Ziola, Department of Microbiology and Immunology, Saskatoon, Saskatchewan, Canada, for providing the rabbit serum for the detection of morbillivirus antigen by immunohistochemistry, Herman Cremers for identifying macroparasites, and Frank van der Panne for his assistance with Figs. 1, 2, and 4–![]() .
.