
Abstract
Pulmonary fibrosis and interstitial lung disease are poorly understood in horses; the causes of such conditions are rarely identified. Equine herpesvirus 5 (EHV-5) is a γ-herpesvirus of horses that has not been associated with disease in horses. Pathologic and virologic findings from 24 horses with progressive nodular fibrotic lung disease associated with EHV-5 infection are described and compared with 23 age-matched control animals. Gross lesions consisted of multiple nodules of fibrosis throughout the lungs. Histologically, there was marked interstitial fibrosis, often with preservation of an “alveolar-like” architecture, lined by cuboidal epithelial cells. The airways contained primarily neutrophils and macrophages. Rare macrophages contained large eosinophilic intranuclear viral inclusion bodies; similar inclusion bodies were also found cytologically. The inclusions were identified as herpesviral-like particles by transmission electron microscopy in a single horse. In situ hybridization was used to detect EHV-5 nucleic acids within occasional macrophage nuclei. With polymerase chain reaction (PCR), the herpesviral DNA polymerase gene was detected in 19/24 (79.2%) of affected horses and 2/23 (8.7%) of the control horses. Virus genera–specific PCR was used to detect EHV-5 in all of the affected horses and none of the control horses. EHV-2 was detected in 8/24 (33.3%) of affected horses and 1/9 (11.1%) of the control horses. This disease has not been reported before, and the authors propose that based upon the characteristic gross and histologic findings, the disease be known as equine multinodular pulmonary fibrosis. Further, we propose that this newly described disease develops in association with infection by the equine γ-herpesvirus, EHV-5.
Interstitial lung disease is a poorly understood entity in adult horses (Equus caballus). 35 As with all veterinary species, a variety of toxic or infectious agents are implicated in targeting the alveolus and interstitium to induce interstitial pneumonia in horses, but these agents are rarely identified at the time of histologic diagnosis. 35 Interstitial fibrosis has been reported as a major component of interstitial lung disease in individual horses, as well as in a retrospective study of idiopathic interstitial lung disease in 20 horses in Florida by Buergelt et al. 6, 8, 36 Additionally, Kleiboeker et al. 13 reported on a single donkey (Equus asinus) that had pulmonary interstitial fibrosis among a group of donkeys with asinine herpesvirus-associated interstitial pneumonia. Interstitial fibrosis is a prominent feature of silicate pneumoconiosis in horses from the Monterey-Carmel peninsula of California, 27 but there is little evidence of other factors associated with fibrotic lung disease in the horse.
Herpesviridae is a large family of DNA viruses that is subdivided into three subfamilies (α, β, and γ) based upon genetic, biologic, and morphologic characteristics. 17 Eight herpesviruses have been identified in equids: 5 in the α-herpesvirinae subfamily and 3 in the γ-herpesvirinae subfamily. Of those 8 viruses, 5 naturally infect the domestic horse and 3 infect the donkey. 22 In horses, the α-herpesviruses equine herpesvirus 1 (EHV-1) and EHV-3 clearly cause clinical disease. The role in disease for the remaining α-herpesvirus, EHV-4, is unclear, although it has been implicated in upper respiratory infections. 22 EHV-2 and EHV-5 are closely related γ-herpesviruses of horses. 1, 23, 30 Whereas evidence of infection in horses with EHV-2 and EHV-5 is reported as relatively common in parts of Europe, Argentina, Australia, and New Zealand, association between infection with these viruses and disease is inconclusive at this time. 3, 7, 20, 25, 33
Herein we provide the first description of the gross pathology, histopathology, and virology findings of 24 horses with a newly recognized fibrotic lung disease that is associated with infection with EHV-5. Whereas this association does not imply causation, to our knowledge, this report is the first to document an association between a viral infection and lung fibrosis in any veterinary species and the first to describe this disease. Based on the characteristic gross and histologic findings of this disease, the authors propose that the disease be termed equine multinodular pulmonary fibrosis (EMPF). Further, we propose that this newly described disease develops in association with an infection with the γ-herpesvirus EHV-5.
Materials and Methods
Tissue Collection and Processing
All of the EMPF animals underwent euthanasia after a period of severe respiratory disease that did not respond to therapy (Table 1). When the clinical history was available, the animals were described as being mildly febrile (100.5–102.8°F), with clinical signs referable to pulmonary disease; numerous discrete to coalescing nodular densities were visible within the lungs when thoracic radiographs were taken. The horses underwent necropsy examination, and a standard series of tissues was collected in 10% neutral-buffered formalin for histopathology. Frozen lung was collected from 11 EMPF-suspect horses at the time of necropsy (Table 2); for the remaining 13 cases, histologic and virologic analyses were performed on formalin-fixed, paraffin-embedded lung (Table 2). Tissues from 23 age-matched control horses (Table 3) were collected at the time of necropsy for analysis (Table 4). The control group consisted of 18 horses obtained from cases submitted to Michigan State University, University of Pennsylvania, and University of Kentucky for pathologic examination. These animals were selected for inclusion in the study based upon similar age to the affected horses and having previously been diagnosed with pulmonary fibrosis of unknown cause. An additional 5 control horses were from a long-term study of recurrent airway obstruction at Michigan State University. Routine evaluation of the control lung tissues for viral agents at the time of diagnostic workup did not identify any significant pathogens; chronic bacterial pneumonia was present in 2 of the control horses.
Characteristics of EMPF horses.∗
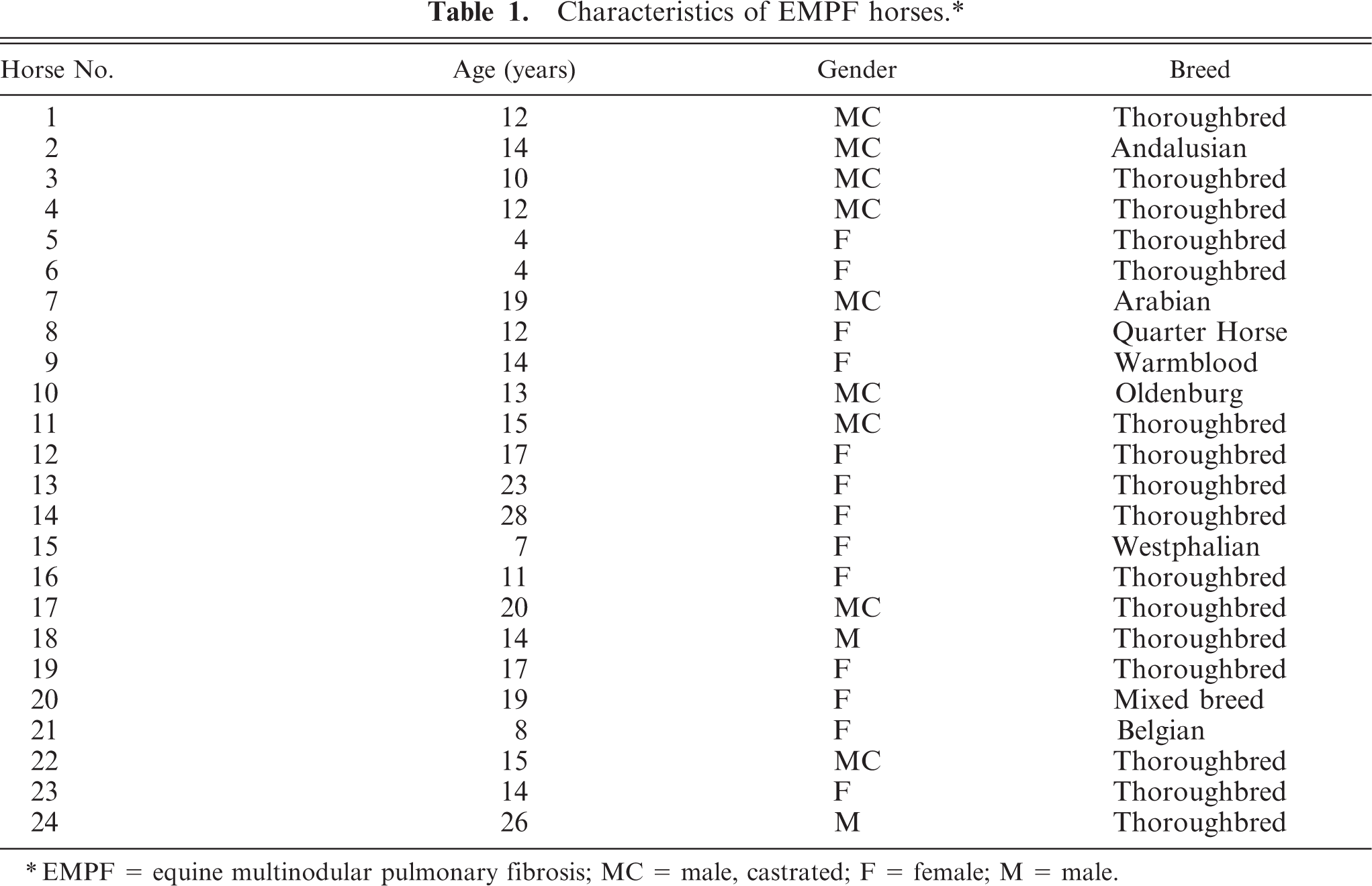
∗EMPF = equine multinodular pulmonary fibrosis; MC = male, castrated; F = female; M = male.
Equine herpesvirus PCR data: EMPF horses.∗
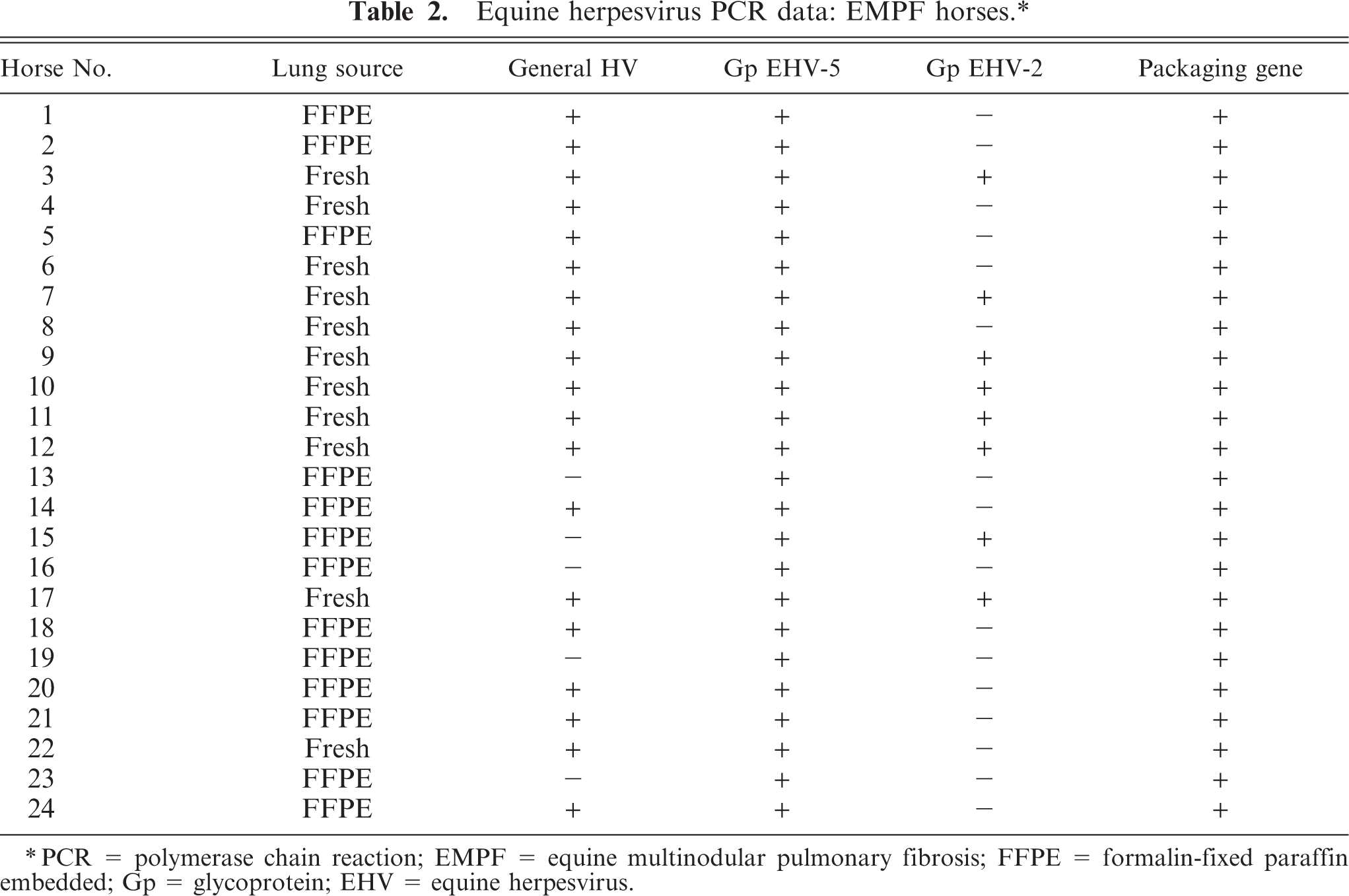
∗PCR = polymerase chain reaction; EMPF = equine multinodular pulmonary fibrosis; FFPE = formalin-fixed paraffin embedded; Gp = glycoprotein; EHV = equine herpesvirus.
Characteristics of control horses.∗
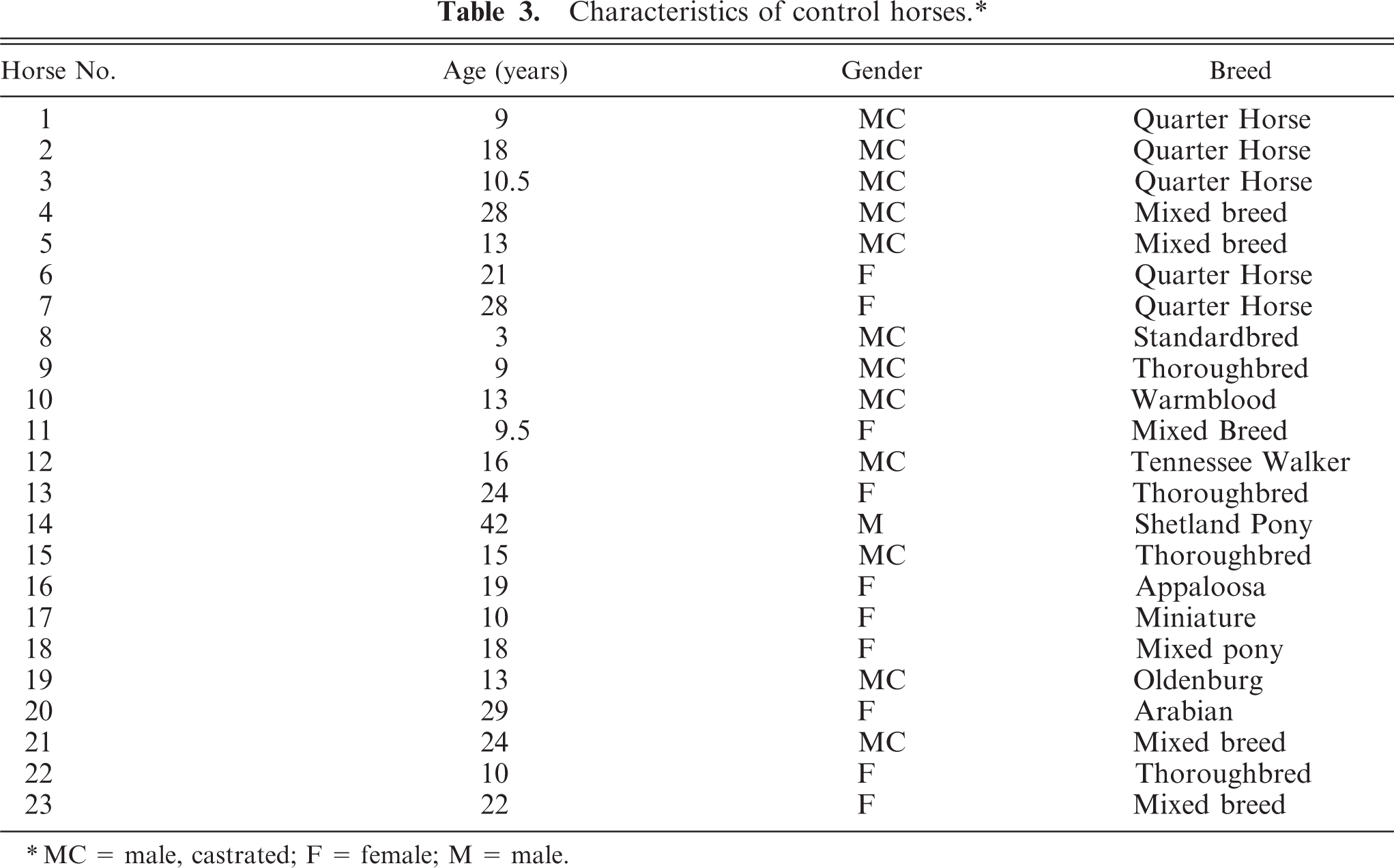
∗MC = male, castrated; F = female; M = male.
Equine herpesvirus PCR data: control horses.∗
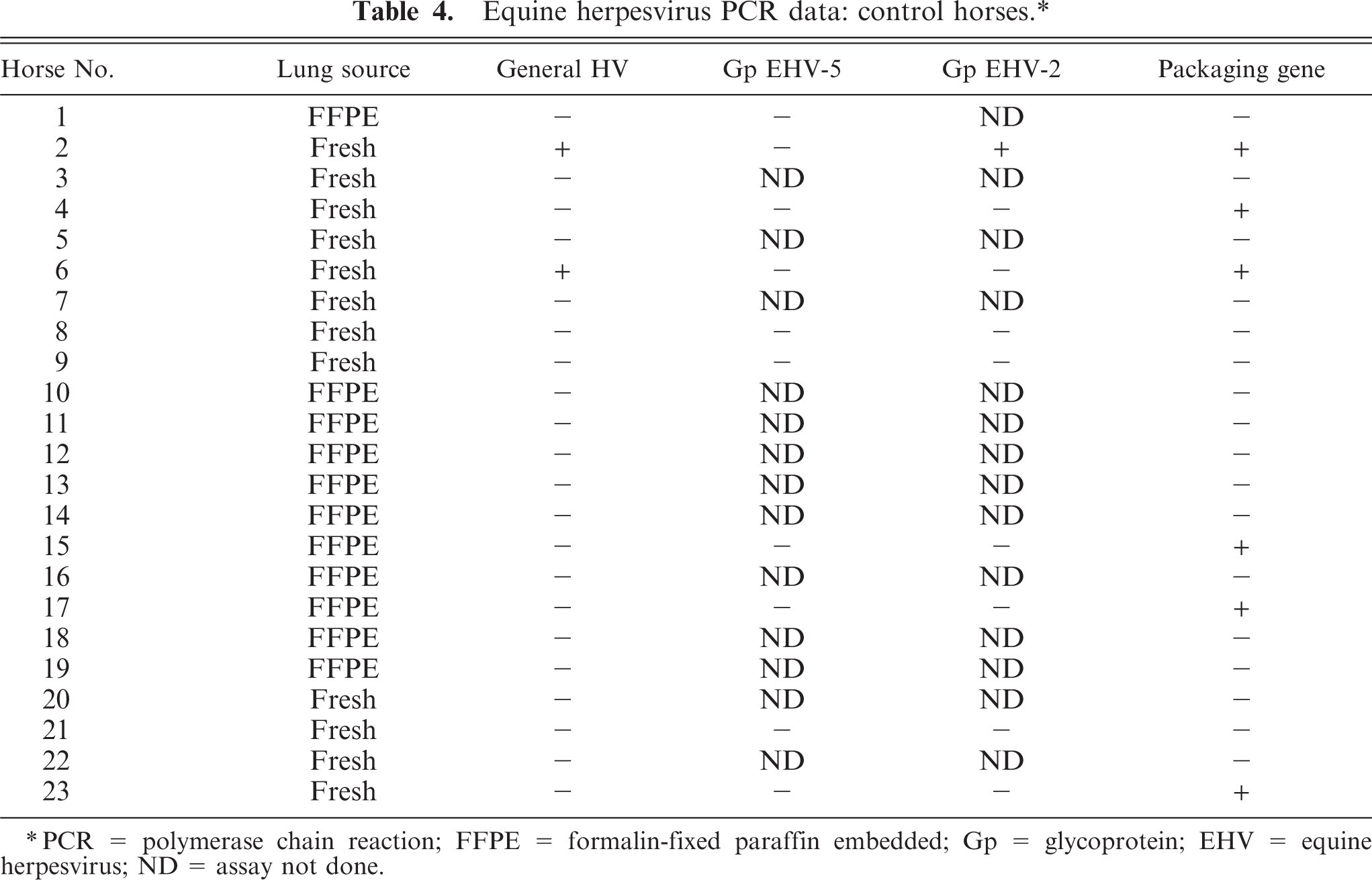
∗PCR = polymerase chain reaction; FFPE = formalin-fixed paraffin embedded; Gp = glycoprotein; EHV = equine herpesvirus; ND = assay not done.
Tissues for histopathology were routinely processed and embedded in paraffin for sectioning and placement on glass slides. All tissue sections were stained with hematoxylin and eosin. Masson trichrome staining was performed to visualize the extracellular matrix within the lung. Impression smears of the lung from horse No. 4 of the affected group were prepared on glass slides immediately after euthanasia for cytologic examination and were treated with Wright-Giemsa stain.
Transmission Electron Microscopy
For examination of tissues for viral particles, fresh lung was obtained from horse No. 4 and fixed in 4% glutaraldehyde. The postfixation processing consisted of rinsing in 0.1 mol/liter phosphate buffer and placement in osmium tetroxide (Electron Microscopy Sciences, Fort Washington, PA). The tissue was rinsed again in the phosphate buffer, transferred into 2% uranyl acetate, and rinsed in a graded series of ethanol. The tissues were placed in propylene oxide (Electron Microscopy Sciences) before being embedded in 2,4,6-tris(dimethylaminomethyl)phenol and araldite 501 (Electron Microscopy Sciences). Sections were cut at 1-μm thickness using an LKB ultramicrotome (Bromma, Sweden) and stained with toluidine blue. Tissues of interest were sectioned at 60 nm, stained with uranyl acetate and lead citrate (Electron Microscopy Sciences), and examined on a Phillips 301 electron microscope (Atlanta, GA).
Polymerase Chain Reaction (PCR) Assays
Extraction of DNA
DNA was extracted from either fresh-frozen lung or formalin-fixed, paraffin-embedded lung (Tables 2, 4). For fresh-frozen lung, approximately 400–500 mg of tissue with ≥1 clearly visible fibrotic regions was macerated with a razor blade, suspended in 4 ml of M199 cell culture medium, and placed in a 15-ml conical tube containing 5 copper-coated ball bearings that were 4.5 mm in diameter. The contents of the tube were then agitated in a vortex at maximum power for 1 minute. After centrifugation at 1,700 × g for 10 minutes, a 200-μl aliquot of supernatant was removed and placed in a 1.5-ml microcentrifuge tube. Twenty microliters of a 0.5% proteinase K solution that contained 10 mM Tris buffer (pH 8.3), 50 mM KCl, and 0.5% v/v polyoxyethylene sorbitan monolaurate was added to the tube, and the mixture was incubated at 56°C overnight. For formalin-fixed paraffin embedded lung, three 20-μm–thick sections of tissue were placed in a 1.5-ml microcentrifuge tube. The formalin-fixed tissue was digested by addition of 200 μl of ATL buffer, followed by the addition of 20 μl of proteinase K solution (DNeasy tissue kit; QIAGEN Inc., Valencia, CA). This mixture also was incubated at 56°C overnight. After digestion, the mixture from the fresh-frozen or the formalin-fixed tissue was centrifuged at 18,000 × g for 3 minutes. For paraffin-embedded tissues, the floated paraffin layer was removed with a toothpick. Next 200 μl of AL buffer was added to the microcentrifuge tubes, and the instructions for the DNeasy Tissue Kit (Qiagen Inc.) were followed through elution of the DNA in 50 μl of nuclease-free water.
PCR assays
Initially, all samples of DNA were tested using a nested PCR that detects most members of the Herpesviridae family of viruses by targeting conserved regions within the herpesviral DNA–directed DNA polymerase gene. 24, 31 The primer sequences and reaction conditions for this general herpesvirus PCR assay and all other PCR assays used during this study are given in Table 5. The 25-μl PCR mixture used to detect the DNA polymerase gene consisted of PCR buffer with 1.5 mM MgCl2, 200 mM each dNTP, 1.25 U of Taq polymerase (QIAGEN Inc.), 1.0 μM each primer, 2.5 μl of DNA for the primary amplification (1 μl of the primary amplification product for the secondary amplification), and molecular biology–grade water to volume. The final amplification product was approximately 210–250 bp. Amplification products (10 μl) were analyzed in ethidium bromide–stained 1.5% agarose gels after electrophoresis in sodium borate buffer (5 mM disodium borate decahydrate, pH adjusted to 8.5 with boric acid). 4 Gel images were captured with a Kodak EDAS 290 imaging system (Eastman Kodak Co., Rochester, NY).
Primers and reaction conditions for PCR assays.

PCR = polymerase chain reaction; EHV = equine herpesvirus.
∗PCR primers that target the viral DNA polymerase gene and detect most herpesviruses.
†PCR primers that detect the equine γ-herpesvirus DNA packaging gene.
‡PCR primers that detect the EHV-2 glycoprotein H gene.
§PCR primers that detect the EHV-5 glycoprotein H gene.
∥PCR primers that detect the EHV-1 glycoprotein B gene.
#TaqMan probe for the EHV-1 glycoprotein B gene.
¶PCR primers that detect the EHV-4 UL24 gene homolog.
Samples of DNA were further tested using several PCR assays specific for genera and subfamilies of Herpesviridae. Those PCR assays targeted the viral DNA packaging protein gene of equine γ-herpesviruses, 13 the viral glycoprotein H (gH) gene of EHV-2 or EHV-5, 20 the viral glycoprotein B (gB) gene of EHV-1, 11 and the viral UL24 homolog gene of EHV-4 (in-laboratory development at Michigan State University). The PCR mixture for the DNA packaging gene was the same as above, and the product of the PCR was an amplicon of approximately 297 bp. A hemi-nested set of PCR primers was used to detect the gH gene of EHV-2, and a single set of primers was used to detect the gH gene of EHV-5. 20 The PCR amplification products were 354 bp for EHV-2 and 344 bp for EHV-5. The PCR mixtures for those assays contained 2X reaction buffer with 1.5 mM MgCl2, 200 mM each dNTP, 0.625 U of Taq polymerase (Promega, Madison, WI), 0.3 μM each primer, 2.5 μl of DNA (1 μl of DNA for the secondary amplification of EHV-2), and molecular biology–grade water to a final volume of 25 μl.
Real-time PCR assays specific for the EHV-1 gB gene or EHV-4 UL24 gene homolog were used to detect the presence of these equine α-herpesviruses. The PCR assay for EHV-1 used a TaqMan probe as a detector of PCR amplification, 11 whereas the PCR assay for EHV-4 used SYBR Green followed by standard postamplification melt-curve analysis. The reaction mixture for EHV-1 was made from the Taq PCR master mix kit (QIAGEN Inc.) with 500 nM each primer, 240 nM probe, 2.5 μl of 25 mM MgCl2, 7 μl of DNA, and molecular biology–grade water to a final volume of 25 μl. The reaction mixture for EHV-4 was made from 25 μl of Quantitect SYBR Green PCR mastermix (QIAGEN Inc.) with 500 nM each primer, 5 μl of DNA, and molecular biology–grade water to a final volume of 50 μl. The amplification product was 302 bp, and the peak melting temperature was 84.5°C. The iCycler iQ5 real-time detection system (Bio-Rad Laboratories Inc., Hercules, CA) was used for both of these PCR assays. Positive controls consisted of DNA from known isolates from EHV-1, EHV-2, EHV-4, and EHV-5; negative controls consisted of the reaction mixture with molecular-grade water and no DNA.
In Situ Hybridization
Complementary hybridization probes were designed from the EHV-5 gH gene sequences derived in this study, using the Clone Manager Professional Suite version 7.11 software (Scientific & Educational Software, Cary, NC). Specificity of the probes was checked against aligned sequences of the EHV-2 gH gene. The probes were labeled on the 5′ end with digoxigenin. The probe sequences were synthesized by Integrated DNA Technologies (Coralville, IA). A similarly labeled nonsense probe specific for the ISMAP02 gene of Mycobacterium avium subspecies paratuberculosis was used as a control. In situ hybridization was performed on 6 EMPF horses and 5 controls essentially as described previously. 21, 34 Briefly, deparaffinized slides were hybridized with the digoxigenin-labeled oligonucleotide probes specific for the gH gene sequences. Then lung sections were incubated with alkaline phosphatase–conjugated anti-digoxigenin antibodies, followed by the NBT/BCIP detection system to amplify the chromogen signal. The digoxigenin-labeled probes used were:
IS MAP02: 5′-GCC ACC ACC GTC TTG ACC GCC TCA AAG ATC -3′
Glycoprotein H-1F: 5′-TCT GAA AGC AAG CTG GTT TCT AAC CGC ACC -3′
Glycoprotein H-1R: 5′-GGT GCT GTT AGA AAC CAG CTT GCT TTC AGA -3′
Virus Isolation
Virus isolation was performed on tissues from 7 of the EMPF horses and 12 control horses, and previously reported standard techniques were followed for EHV-5. 28 Bovine viral diarrhea virus–free rabbit kidney (RK-13) cells were grown in Eagle minimum essential medium supplemented with 10% fetal bovine serum, 0.29 mg/ml glutamine, 100 IU/ml penicillin, and 100 μg/ml streptomycin. Semiconfluent monolayers were inoculated with a lung homogenate from 1 of the affected horses, first clarified by low-speed centrifugation and then passed through a filter with an average pore diameter of 0.45 μm. The inoculum was adsorbed onto the cultures in a small amount of medium for 1 hour at room temperature. The cells were washed, re-fed, and incubated at 37°C in an atmosphere of 5% CO2. The inoculated cells were observed daily for the appearance of cytopathic changes. Five passages of the inoculated cell cultures were at an interval of 7 days between passages. Confirmation of viral infection of the cells was accomplished using the PCR methods outlined above.
Results
Summary of Cases
A total of 24 EMPF horses (Table 1) and 23 age-matched control horses (Table 3) were included in the study. The average age of the EMPF horses was 14.5 years; 11 were males and 13 were females. There were 16 Thoroughbreds and 1 of each of the following breeds: Andalusian, Arabian, Belgian, mixed breed, Oldenburg, Quarter Horse, Standardbred, and Westphalian (Table 1). The average age of the control horses was 17.5 years; 13 were males and 10 were females (Table 3). There were 5 Quarter Horses, 5 mixed-breed horses, 4 Thoroughbreds, 3 ponies, and one of each of the following breeds: Standardbred, Warmblood, Tennessee Walker, Appaloosa, Oldenburg, and Arabian (Table 3).
Gross Pathology
The most significant gross lesions were restricted to the lungs and involved all lung regions. There were 2 distinct gross manifestations of EMPF, the more common being numerous coalescing nodules of fibrosis (Figs. 1, 2). The individual nodules ranged from <1 to 5 cm in diameter. The nodules were pale tan-white and moderately firm; the borders were discrete where they met unaffected lung. Little unaffected lung is usually present when this form of the disease is recognized. The less common gross lesion consisted of multiple discrete nodules separated by grossly unaffected lung; these nodules may be confused with a neoplastic process (Figs. 3, 4). These masses of fibrosis were larger than the coalescent form of the disease (up to 10 cm), pale tan-white, and firm, and larger areas of grossly normal lung were between the nodules. In each form of the disease, the cut surface of the nodules was uniformly colored, and the foci of fibrosis bulged slightly from the surrounding tissue. Coincident with the lung lesions, the bronchial lymph nodes in the affected horses were markedly enlarged in 12/24 EMPF horses (not shown). The gross lesions of EMPF were distinctive and did not resemble the lesions of any of the control horses. In the control horses, fibrosis was not the primary gross manifestation of disease in most cases and did not involve the extent of the lung field as seen with EMPF (see histologic description below). In 5 of the control animals (Nos. 1, 10, 11, 12, and 16), nearly diffuse fibrosis was prominent as part of a granulomatous inflammatory process; unlike EMPF, discrete granulomas with mineralization were found in these 5 animals.
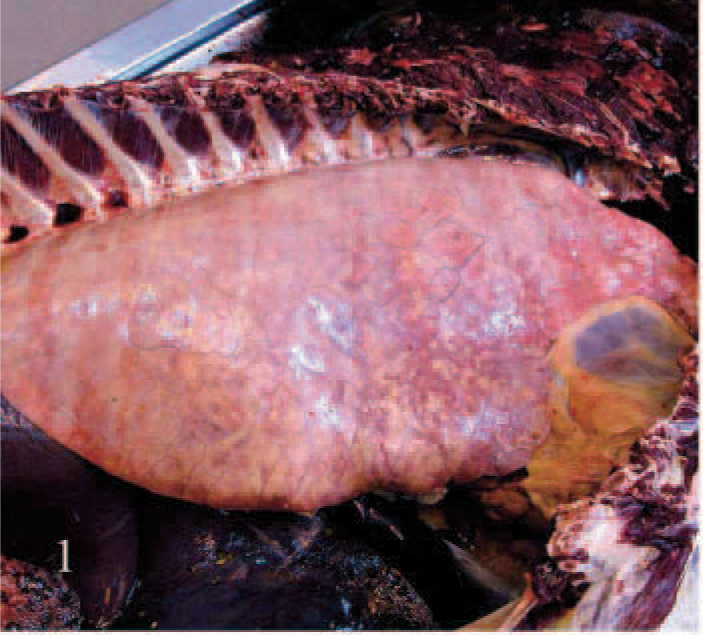
EMPF lungs; horse No. 4. Gross pathology of EMPF, diffuse nodular form. The pulmonary parenchyma fails to collapse, and rib impressions are visible on the pleural surface.
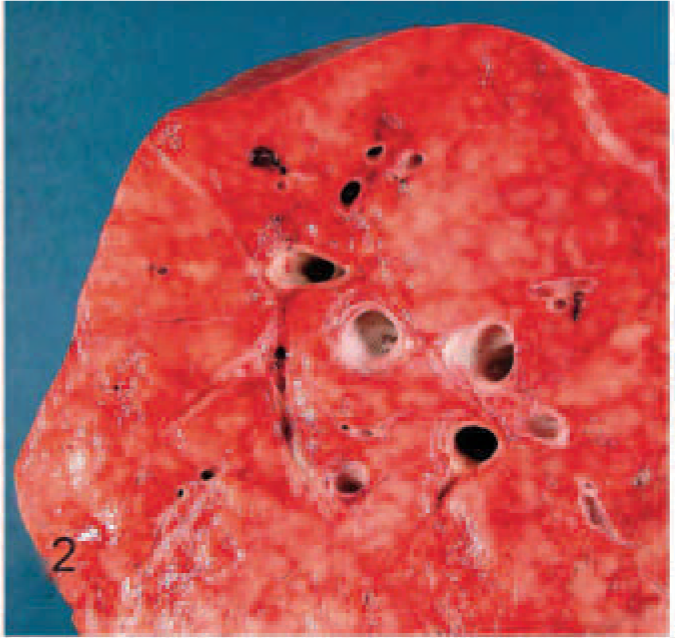
EMPF lungs; horse No. 4. Gross pathology of EMPF, diffuse nodular form, lung cut section. The parenchyma is nearly replaced by coalescent, variably sized, pale tan, moderately firm nodules of fibrosis.
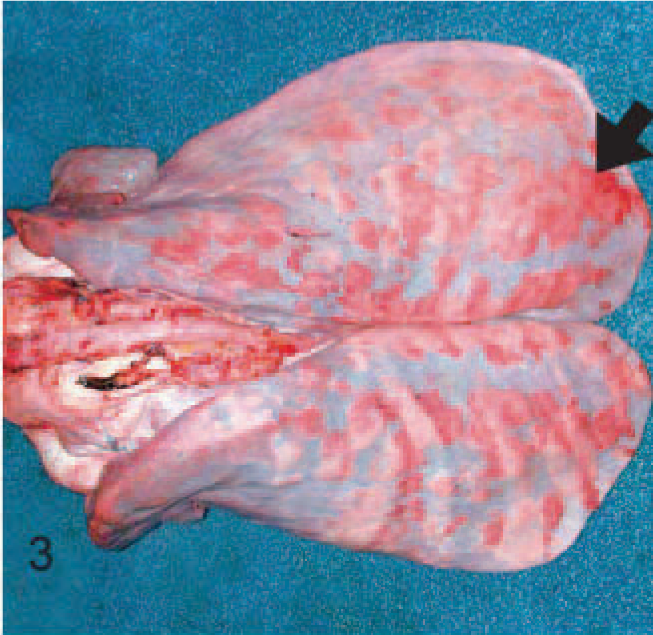
EMPF lungs; horse No. 5. Gross pathology of EMPF, discrete nodular form. Discrete nodules of fibrosis can be seen scattered throughout the lung beneath the pleural surface (arrow).
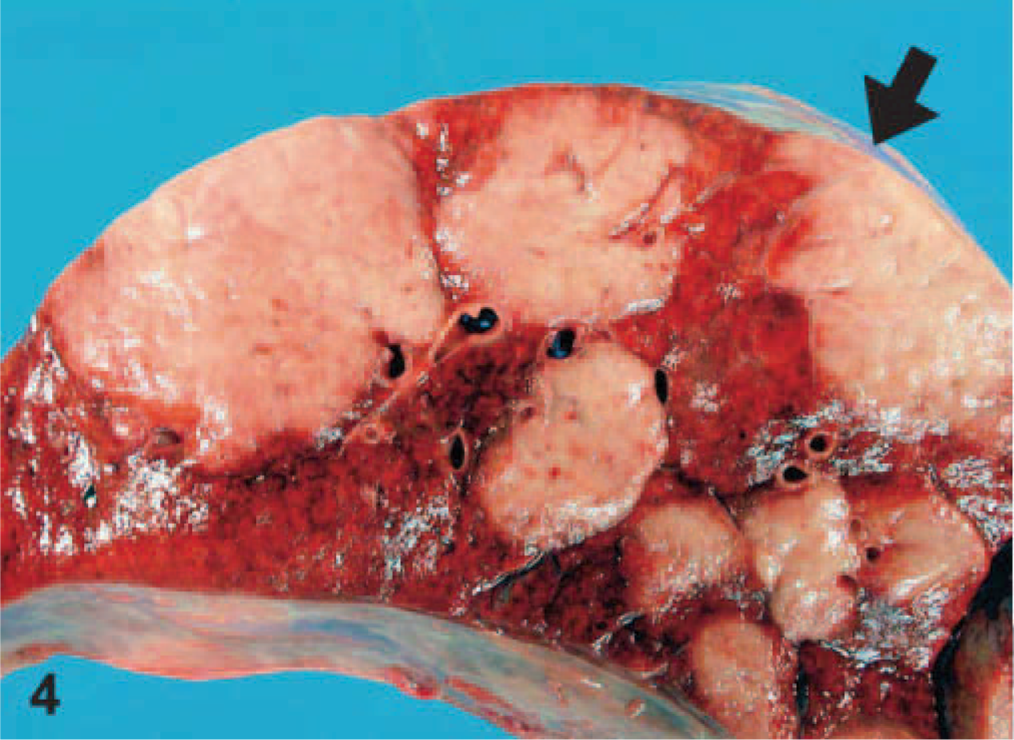
EMPF lungs; horse No. 5. Gross pathology of EMPF, discrete nodular form, lung cut section. The discrete nodules of fibrosis are large and have sharp borders between adjacent nodules and grossly normal lung (arrow).
Histopathology and Cytology
Significant histologic findings in EMPF were restricted to the lung and bronchial lymph nodes. The lung lesions were largely confined to the alveolar parenchyma and were similar regardless of the gross pathology. Nodules were sharply demarcated from adjacent less affected lung (Fig. 5). The nodules consisted of marked interstitial expansion by well-organized mature collagen (Figs. 6– 8). The interstitium was infiltrated by variable numbers of mixed inflammatory cells, consisting primarily of lymphocytes, with smaller numbers of macrophages, neutrophils, and occasional eosinophils. There was preservation of an “alveolar-like” architecture, with the lumen of the spaces lined by cuboidal cells (Figs. 6– 8). Less commonly, the fibrosis was less well organized, with the collagen deposition arranged in broad interlacing bundles that did not preserve the “alveolar-like” architecture (Fig. 9). The lumen contained moderate numbers of neutrophils and macrophages, and rarely there were large macrophages with abundant eosinophilic cytoplasm and a large oval eosinophilic intranuclear viral inclusion body (Fig. 10). The histology of the bronchial lymph nodes consisted of lymphoid hyperplasia, often with sinus histiocytosis and occasional multinucleate giant cells (not shown); viral inclusion bodies were not noted within cells within the lymph nodes.
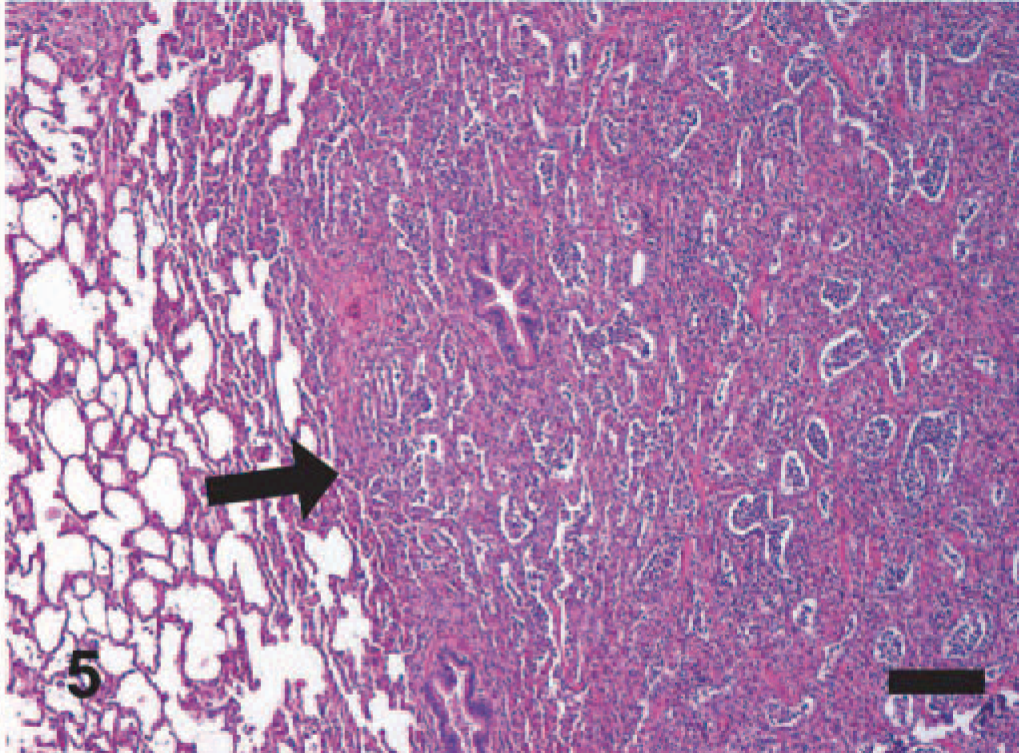
EMPF histopathology; horse No. 6. Histologically, there is a discrete border between the foci of interstitial fibrosis and the relatively unaffected lung (arrow). HE. Bar = 200 μm.
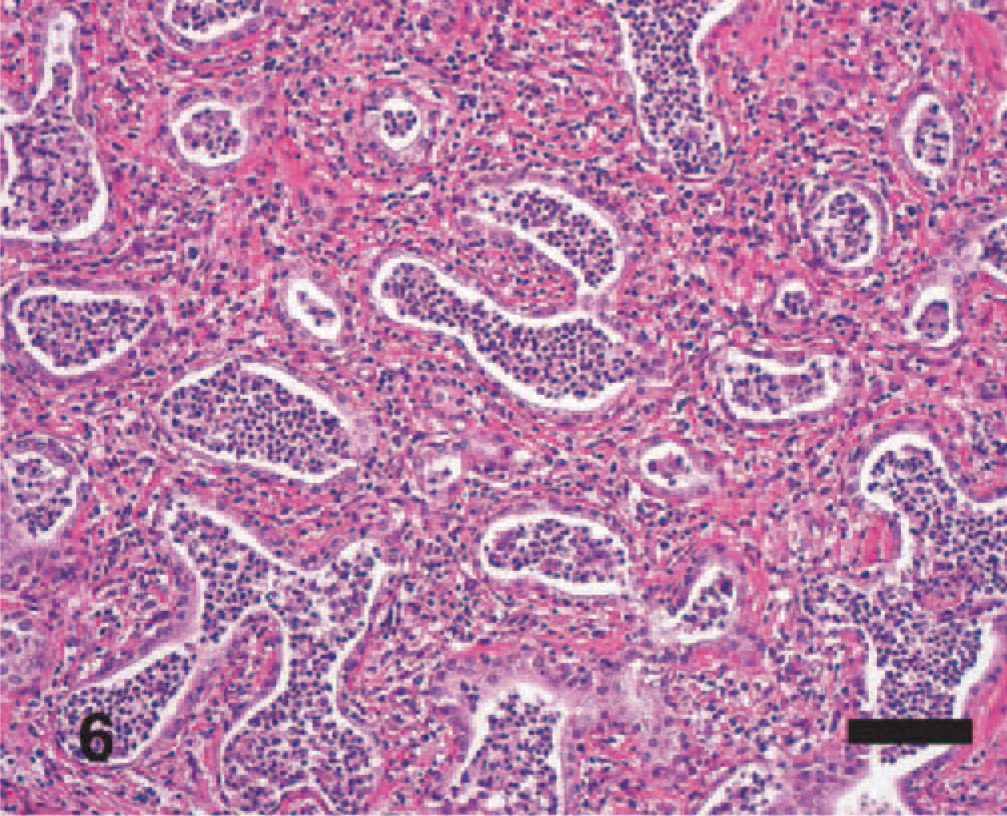
EMPF histopathology; horse No. 6. In most cases, there is preservation of an “alveolar-like” architecture, which is lined by cuboidal epithelial cells. HE. Bar = 100 μm.
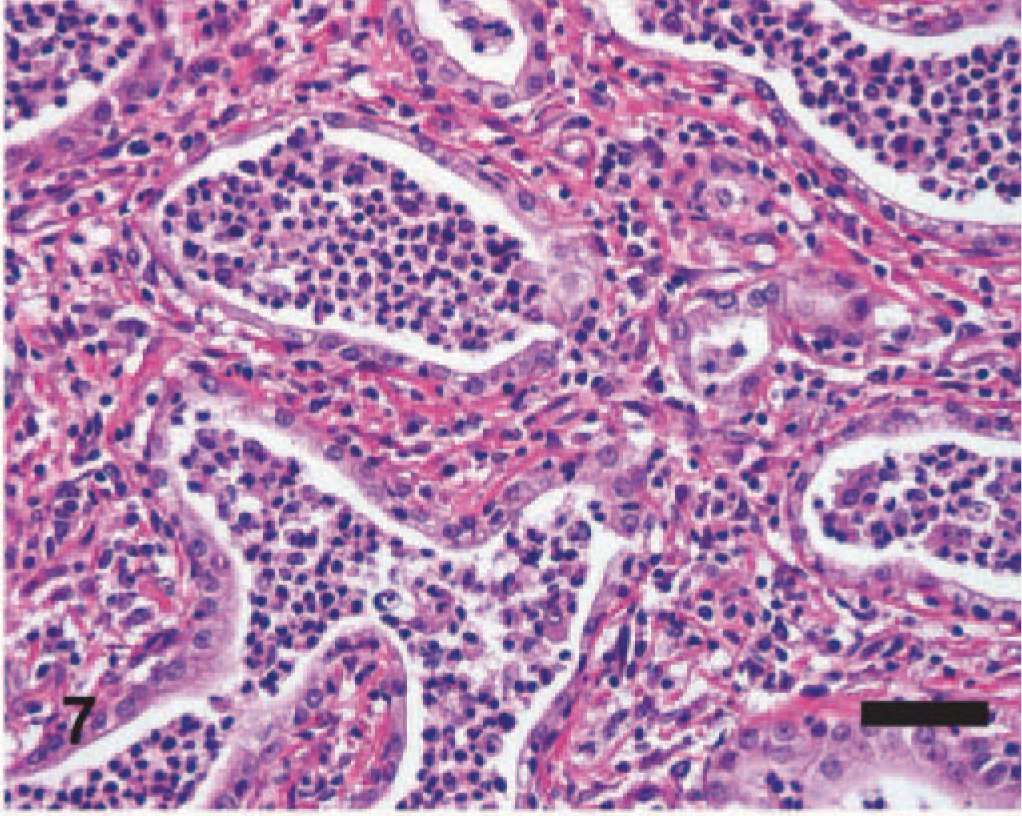
EMPF histopathology; horse No. 6. Higher magnification of interstitial fibrosis with inflammatory cells within lumen of the airspace; the inflammatory cells are primarily neutrophils and macrophages. HE. Bar = 50 μm.
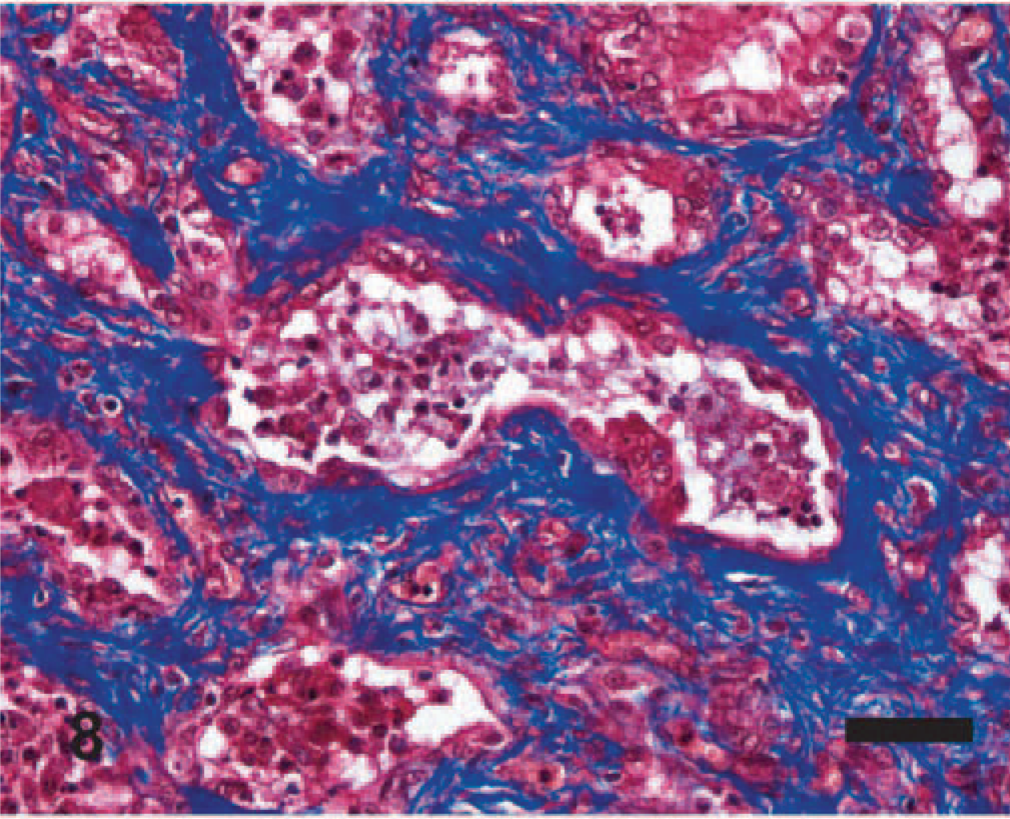
EMPF histopathology; horse No. 6. Histochemical staining of interstitial fibrosis. The fibrosis (blue) in most animals is well organized around the airspaces. Masson trichrome. Bar = 50 μm.
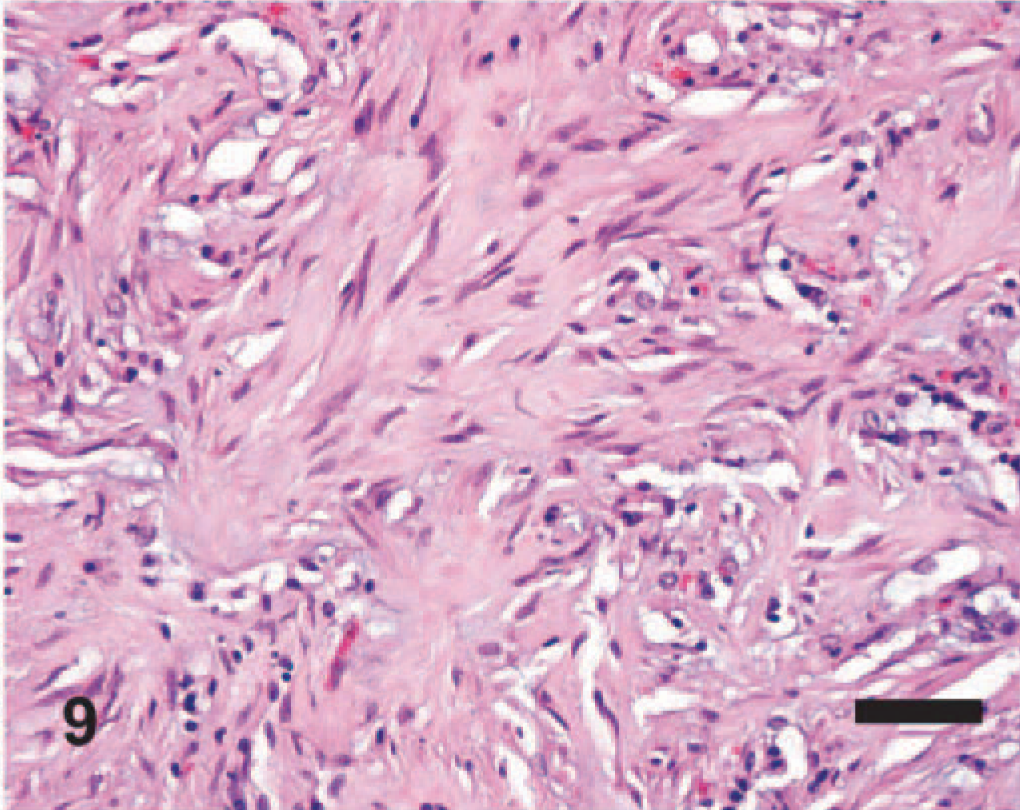
EMPF histopathology; horse No. 22. The fibrosis can be arranged in unorganized bands, without appreciable preservation of “alveolar-like” airspaces. HE. Bar = 50 μm.
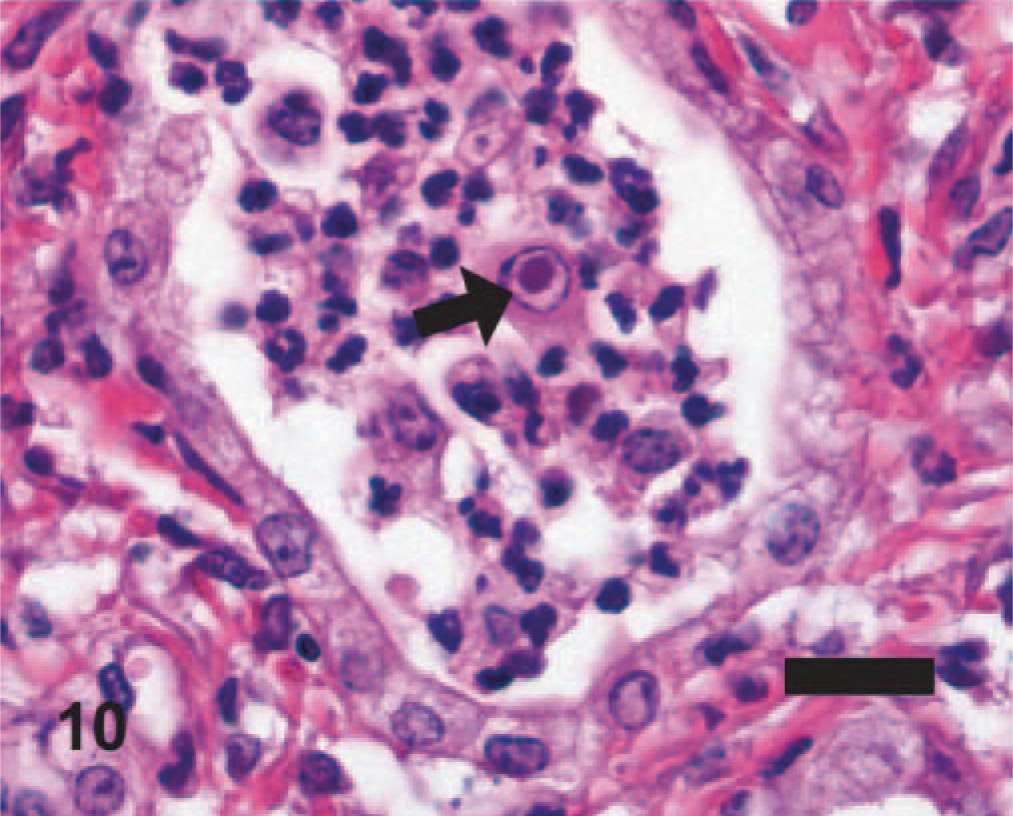
EMPF histopathology; horse No. 6. Large eosinophilic intranuclear viral inclusion body (arrow) in a large macrophage within the inflammation in the airspaces of the nodular lesions. HE. Bar = 20 μm.
The imprints of fresh lung from horse No. 4 of the affected group of horses were highly cellular; intact cells were mostly neutrophils, macrophages, and rafts of relatively uniform epithelial cells that often appeared cuboidal and had moderately basophilic cytoplasm. There were fewer mast cells and rare small clusters of spindle cells. Macrophages were often vacuolated, and low numbers were multinucleated. Low numbers of macrophages were enlarged (approximately 40 μm versus 20 μm for uninfected) and had large nuclei containing a finely granular, centralized eosinophilic intranuclear inclusion (Fig. 11); nucleoli were often located peripherally. Bacteria or other infectious agents were not found.
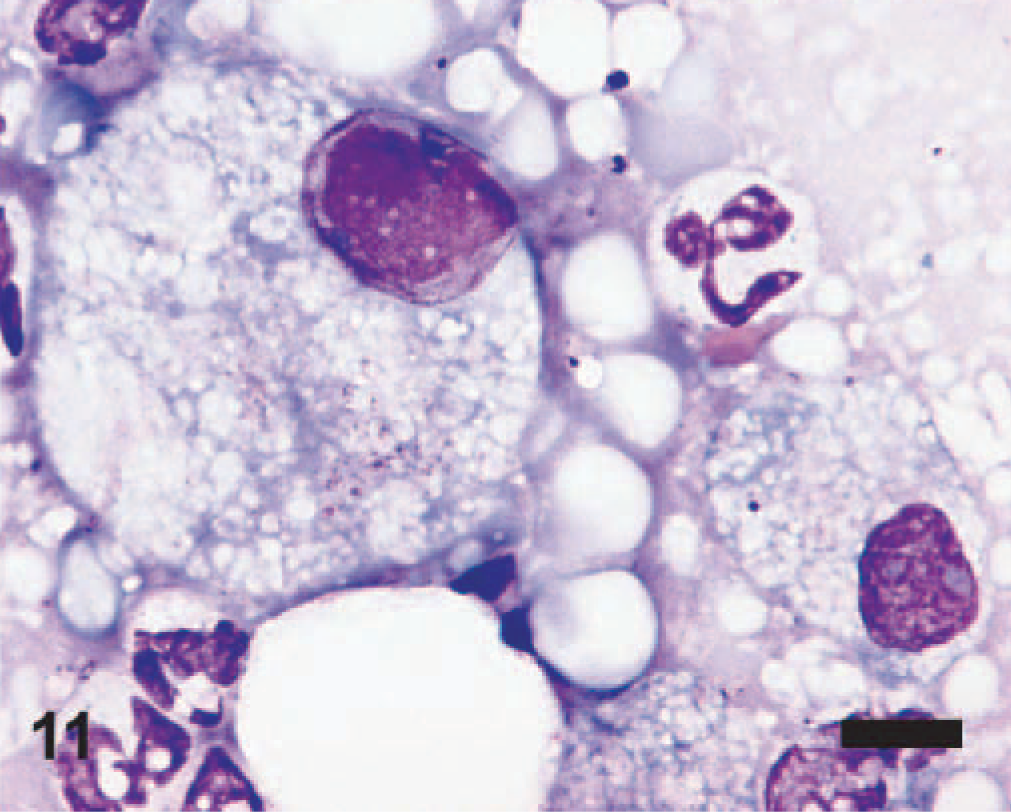
EMPF lung imprint cytology; horse No. 4. Vacuolated macrophages and neutrophils are intermixed. One macrophage (left) is markedly enlarged and has a single large eosinophilic intranuclear inclusion that marginates 2 nucleoli. Wright-Giemsa stain. Bar = 10 μm.
The histologic characteristics of the fibrosis of EMPF (described above) were distinct from the pattern of collagen deposition found in the control animals. Fibrosis was a significant histologic feature in 14/23 of the control horses. In the control animals, 5 of the animals had lung fibrosis as part of granulomatous pneumonia of unknown cause (control horses Nos. 1, 10, 11, 12, and 16), 5 had chronic interstitial pneumonia with fibrosis of unknown cause (control horses Nos. 13, 14, 15, 17, and 18), 2 had fibrosis in the lung secondary to chronic bacterial pneumonia (control horses Nos. 19 and 20), and fibrosis was a feature of a single case of chronic hypertension (control horse No. 22) and of metastatic adenocarcinoma to the lung (control horse No. 21). The diagnoses associated with the remaining control horses included lymphoma (control horse No. 9), chronic recurrent airway obstruction (control horses Nos. 2, 4, 5, 6, and 7), disseminated intravascular coagulation with pulmonary thrombosis (control horse No. 3), chronic Cushing disease (control horse No. 23), and congestion and edema associated with acute heart failure (control horse No. 8). All of the cases of EMPF were histologically distinguishable from control lungs with fibrosis based upon the nodular pattern of collagen deposition, presence of luminal neutrophils and macrophages within the “alveolar-like” architecture, and finding intranuclear viral inclusion bodies within the enlarged macrophages.
In Situ Hybridization
The digoxigenin label was localized only to the affected areas of the lungs of the EMPF horses; there was no specific labeling within the adjacent unaffected lung or within the lungs of the control horses. The label was found within the nuclei of scattered macrophages within the “alveolar-like” airspaces in the fibrotic lung (Fig. 12). There was no specific labeling with the nonsense probe or when the protocol excluded the virus-specific probe.
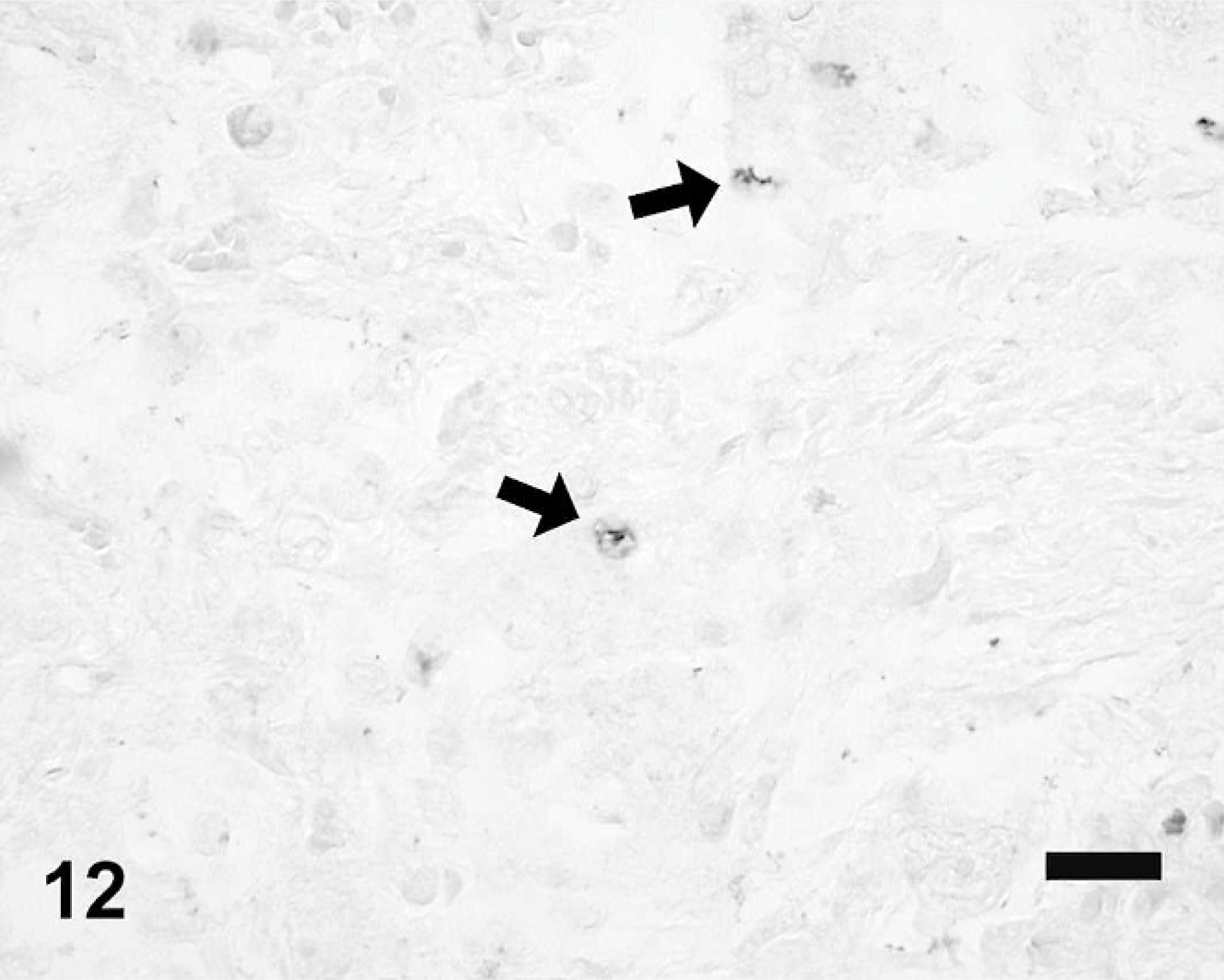
EMPF lung in situ hybridization; horse No. 6. EHV-5 DNA within nuclei of scattered alveolar macrophages (arrows). In situ hybridization with digoxigenin-labeled DNA oligonucleotide probe for the EHV-5 gH gene. Alkaline phosphatase/NBT/BCIP chromogen, no counterstain. Bar = 50 μm.
Transmission Electron Microscopy
Viral particles were identified only within the nuclei of macrophages within the lumen of the airspaces within the fibrotic lung. The chromatin of the affected cells was at the periphery of the nucleus, adjacent to the nuclear membrane (Fig. 13). Some nuclei contained numerous fully formed icosahedral capsids, with sizes (100 nm) and hexagonal shapes consistent with herpesvirus (Fig. 14).
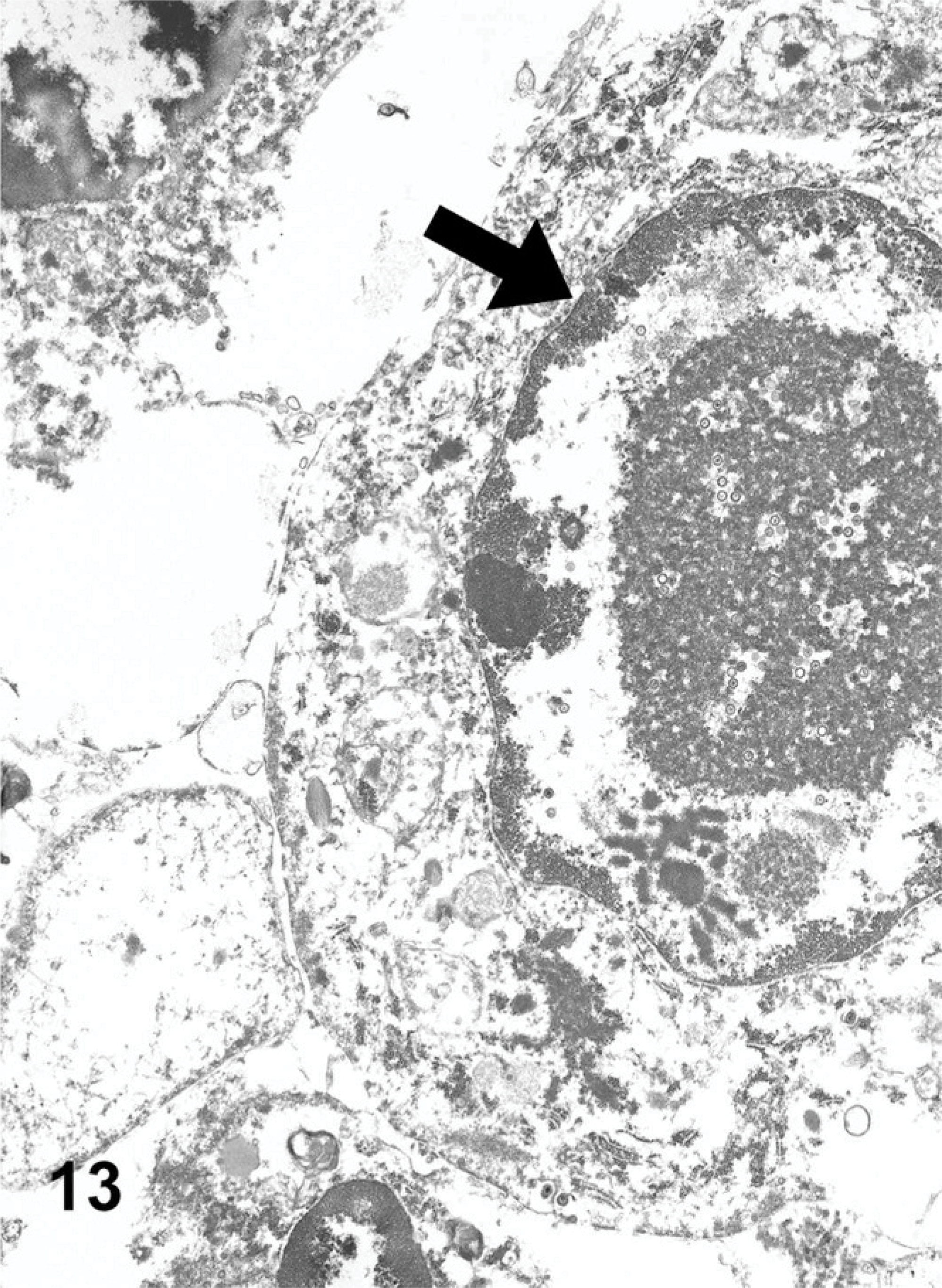
Lung ultrastructure EMPF; horse No. 4. Electron micrograph of an alveolar macrophage with an intranuclear inclusion; central viral capsids, and capsid proteins; note marginated chromatin (arrow).
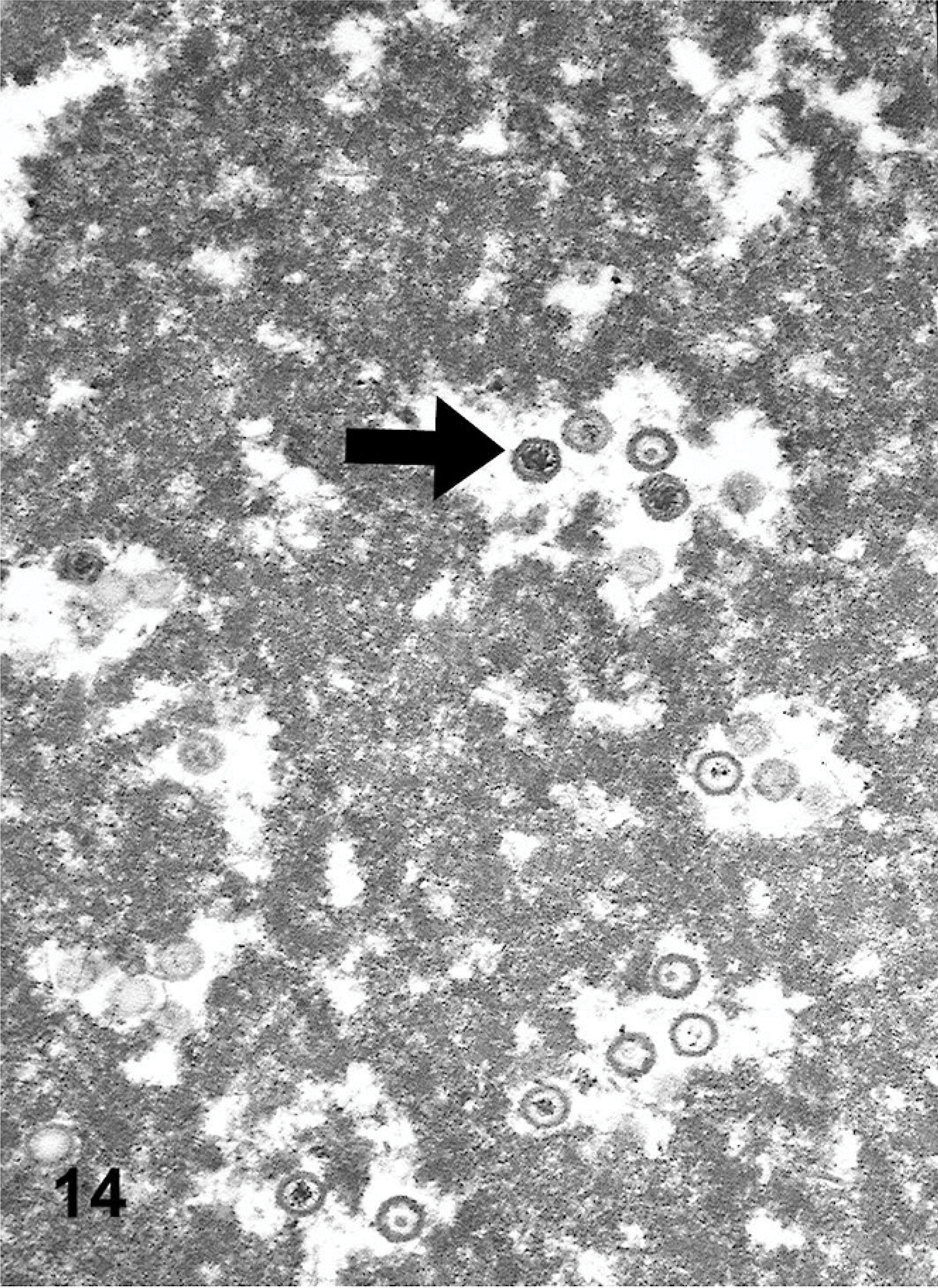
Lung ultrastructure EMPF; horse No. 4. Higher magnification electron micrograph of assembled herpesviral capsids and surrounding capsid proteins; note icosahedral shape of capsids (arrow).
PCR from Lung Tissue
All samples from control horses and EMPF horses were screened in PCR assays with primers specific for Herpesviridae family (viral DNA polymerase gene), γ-herpesvirus subfamily (viral DNA packaging protein gene), or virus genera (EHV-1, EHV-2, EHV-4, and EHV-5) (Table 5). PCR analysis of fresh lung tissue or of tissue derived from archived paraffin-embedded lung was used to detect the presence of herpesviral DNA polymerase gene in 19/24 (79%) affected horses, whereas 2/23 (8.7%) control horses were positive. Failure to detect the herpesvirus DNA polymerase gene in the affected horses was restricted to the formalin-fixed, paraffin-embedded tissues; this is due to the inefficiency of amplification of such large amplicons from DNA extracted from formalin-fixed, paraffin-embedded tissues. When virus genera–specific PCR was performed, all of the affected horses were positive for EHV-5 and 8/24 (33%) affected horses were positive for EHV-2. None of the horses in the control group that were positive for the herpesvirus DNA polymerase gene or for the γ-herpesvirus DNA packaging protein gene were positive for EHV-5. Tissue from 1 of the horses in the control group was positive for EHV-2. Nucleic acid sequencing (data not shown) of the amplicons generated by the PCR assays confirmed the specificity of the PCR products. Lung tissue from all affected and control horses were negative for EHV-1 and EHV-4 using the aforementioned PCR assays. Six control horses were positive for the γ-herpesvirus packaging gene. Of these 6, 1 horse was positive for EHV-2; the other 5 were negative for EHV-5 and EHV-2 using the aforementioned primer sets. Nucleic acid sequence analysis of the putative γ-herpesvirus detected in those 5 horses is ongoing.
Virus Isolation
EHV-5 was isolated from 2/7 EMPF horses (horses Nos. 2 and 6), and EHV-2 was isolated from 1/7 (horse No. 8). Cytopathic changes were first observed in the RK-13 cells on passage 3, and they developed slowly. The cytopathic effect for EHV-5 consisted of foci of infected cells, with some evidence of ballooning (Fig. 15); these foci became more numerous upon additional passages. The EHV-5–specific PCR was strongly positive for DNA extracted from passage 5 cells. A virus stock prepared with passage 5 material had a titer of 3.2 × 105 50% tissue culture infective dose per ml. EHV-5 was not isolated from any of the control horse lungs.

Rabbit kidney cells (RK-13) infected with EHV-5 (cytopathic effect). Cells surrounding a focus of lysis (plaque) are swollen (ballooning degeneration; arrow).
Discussion
Interstitial lung disease is an uncommon event in adult horses, and interstitial fibrosis as a major component of such lung disease is very poorly understood. 35 Interstitial lung fibrosis has been reported as part of the granulomatous process in an apparent silicate pneumoconiosis in horses from the Monterey-Carmel peninsula of California, 27 but there is little evidence of other factors associated with fibrotic lung disease in the horse. EHV-5 is a relatively recently described γ-herpesvirus that, thus far, has not been associated with clinical disease in equids. 1, 2, 30 The current study describes a newly recognized nodular fibrotic lung disease of horses that is associated with pulmonary infection with EHV-5. The virus was identified in all of the affected horses using virus genera–specific PCR, was isolated from 2/7 affected horses, and was visualized using transmission electron microscopy. In addition, in situ hybridization demonstrated viral genome within the pulmonary lesions but not in adjacent unaffected lung or within the lungs of control horses. Because of the characteristic gross and histologic findings and the identification of γ-herpesvirus EHV-5 in affected animals, the authors propose that this newly recognized disease be known as EMPF; further, we propose that this disease is associated with infection with EHV-5.
Idiopathic interstitial pneumonia with fibrosis, sometimes referred to as idiopathic pulmonary fibrosis, has been sporadically reported in individual horses, as well as in a retrospective study of 20 horses in Florida. 6, 8, 35, 36 In the report by Buergelt et al., 6 6 of the 20 horses were older adults, and interstitial fibrosis was described as a major feature of the disease in this subset of animals. The authors suggested that the fibrosis may be a sequela to previous diffuse alveolar damage, as noted in the series of foals from the same study. 6 With the description of EMPF in the current study and the ability to identify the presence of the virus in archived paraffin-embedded lung tissue, it will be interesting to survey additional animals for this newly recognized virus-associated pulmonary disease. Whereas this report is the first to thoroughly describe the pathology of EMPF and its association with a viral infection, it seems likely that the disease has been present for years; similar cases have been reported by other pathologists for at least 15 years (J. M. King, personal communication). The pattern of lung fibrosis, the paucity of viral inclusion bodies, and an incomplete general understanding of viral infections and tissue fibrosis may have contributed to this disease going unreported for some time.
Recently Kleiboeker et al. 13 reported on a series of 17 donkeys (Equus asinus) with asinine herpesvirus-associated interstitial pneumonia. The described gross and histopathologic lesions of the asinine disease differ from EMPF. EMPF is a distinctly nodular disease, with a relatively sharp border of demarcation between the fibrotic and unaffected lung, whereas asinine herpesvirus-associated interstitial pneumonia of donkeys is described as a coalescing inflammatory disease with syncytial cell formation. The authors described areas of interstitial fibrosis associated with the disease but emphasized the inflammatory component rather than fibrosis as the primary finding within the lungs of affected donkeys. 13 Further, viral inclusion bodies were not identified in the donkeys, whereas low numbers of intranuclear viral inclusions were regularly found in the affected horses. Because of these differences, we consider EMPF to be a distinct disease of the domestic horse (Equus caballus).
The identification of this virus as EHV-5 is the first published association of this virus with a disease. There is limited information on the prevalence of EHV-5 infection in horses. Most studies have identified the presence of the virus in nasal swabs or peripheral blood leukocytes using PCR, and there has been no clear association with disease in the studied animals. 3, 20, 23, 33 There is no information on the prevalence of EHV-5 infection within the lower respiratory tract of horses. Earlier studies found that EHV-5 was detected in peripheral blood leukocytes at a later age and much lower frequency than EHV-2, suggesting that infection occurs later in life in most horses and is of low prevalence in equine populations. 20 Recently a study by Bell et al. 3 detected EHV-5 in 64% of nasal samples of young racehorses, and Wang et al. 3, 33 detected EHV-5 in 48% of nasal swabs from 5- to 9-month-old Thoroughbred foals. This is an interesting finding, given the age of the horses with EMPF, and suggests that this disease, if related to the viral infection, may have a prolonged lag period between exposure to the virus and development of the disease. Our studies failed to identify EHV-5 in any of the control horses, but uniform infection in the horses with EMPF was found, suggesting that it may play a role in the development of the disease. Detection of the γ-herpesvirus packaging gene, but neither of the two known γ-herpesviruses of horses (EHV-2 or EHV-5), in 5 of the control horses is an interesting finding and suggests that there may be additional unidentified γ-herpesviruses in equine lung tissue.
In situ hybridization identified viral genome within the regions of fibrosis but not in the adjacent unaffected lung or in the control horse lungs. Virus was detected within luminal macrophages of the EMPF lungs. This pattern of cellular infection is unusual for a γ-herpesvirus, for which the primary target cells for infection are usually B and T lymphocytes. 2 Infection of alveolar epithelial cells has been reported to occur following experimental infection with the murine γ-herpesvirus MHV-68, and this virus has also been associated with experimental fibrotic lung disease in mice. 10, 16, 26 The appearance of the infected alveolar macrophages bearing the inclusion bodies is suggestive of cytomegalovirus (CMV) infection. CMV belongs to the β-herpesvirus subfamily of the Herpesvirinae family. 2 Inclusion bodies are not a common feature of γ-herpesviral infections, although there is a recent report of γ-herpesvirus infection and intranuclear inclusion bodies within the epithelium of genital carcinomas of California sea lions. 14 Interestingly, EHV-5 was initially classified within the β-herpesvirus subfamily based upon shared biologic characteristics and the structure of the genome; 5 it was only after analysis of nucleotide and amino acid sequences that the virus was placed within the γ-herpesviruses. 30 Our preliminary sequencing data from the amplicon from the DNA polymerase gene (general herpesvirus primers) indicates that the virus in question is a not a β-herpesvirus but a γ-herpesvirus. Work is underway in our laboratory to further the molecular phylogenetic classification of this virus to determine its relationship to the reported EHV-5.
This is the first report of an association between a spontaneous, naturally occurring viral infection and fibrotic lung disease in veterinary medicine. There is considerable interest and controversy as to what role, if any, viral infections play in human interstitial lung diseases and pulmonary fibrosis. 15, 32 In humans, a variety of viral infections have been implicated in the pathogenesis of idiopathic pulmonary fibrosis (IPF), including hepatitis C virus, adenovirus, CMV, and Epstein-Barr virus. 15 Of these, there is the greatest amount of data concerning herpesviruses, especially Epstein-Barr, and IPF. 9, 12, 29 Tang et al. 29 used PCR to identify herpesviral DNA in 97% of human IPF cases compared with only 36% of non-IPF cases. Of the viruses that were investigated, human CMV, Epstein-Barr, and human herpesvirus 8 (HHV-8) were most commonly detected within the IPF lung; 29 both Epstein-Barr and HHV-8 are γ-herpesviruses, whereas CMV is a β-herpesvirus. Similar to the study by Tang et al., 29 we used PCR analysis to survey both affected and control horses for 4 different equine herpesviruses (EHV-1, EHV-2, EHV-4, and EHV-5); we detected EHV-5 only in the EMPF cases, suggesting that EHV-5 is specifically associated with EMPF and is not an incidental finding in diseased equine lungs. EHV-2 was found as a co-infection in 8 of the EMPF horses; this is likely not associated with development of EMPF, as most affected horses did not carry this virus within the lung. The actual prevalence of EHV-2 in the control horses is likely higher than reported here, as the nested PCR used to detect the virus is not as effective with DNA extracted from formalin-fixed, paraffin-embedded tissues.
How γ-herpesviruses may drive pulmonary fibrosis is not known at this time. In murine models of pulmonary fibrosis, lung infection with the murine γ-herpesvirus MHV-68 can act synergistically with bleomycin to potentiate the degree of fibrosis, although the virus alone did not result in lung fibrosis in infected mice. 16 In Th-2–biased, interferon-γR−/− mice, MHV-68 infection can result in the development of progressive lung fibrosis, as well as increased expression of important markers of a profibrotic lung environment, including increases in transforming growth factor β, production of Th-2 cytokines, and myofibroblast transformation. 18 Interestingly, many γ-herpesviruses encode viral homologs of a variety of cytokines and chemokine receptors. 19 These proteins may assist the virus in evading detection by the immune system and push the immune response away from a Th-1 response and toward a Th-2 response. 19 This shift in cytokine and chemokine profile may predispose the lung to a progressively fibrotic phenotype. Work is ongoing to evaluate the γ-herpesvirus associated with EMPF for similar cytokine and chemokine receptor homologs as a beginning step toward understanding the pathogenesis of this disease.
Footnotes
Acknowledgements
We thank Dr. Elizabeth Howerth at the University of Georgia for submitting a case for the study and Dr. Perry Habecker at New Bolton Center for contributing gross and histologic evaluation of several of the cases from the University of Pennsylvania.