
Abstract
Fowl glioma-inducing virus (FGV), which belongs to subgroup A of avian leukosis virus (ALV), shows tumorigenicity and pathogenicity, mainly in the nervous system, and causes astrocytoma and perineurioma. Apart from these neoplasms, cerebellar anomaly was found in chickens infected with FGV in ovo. The study reported here describes the morphologic characteristics of the affected cerebellum. Specific-pathogen-free chickens (C/O) were inoculated with FGV through the yolk sac on the 7th day of incubation. The cerebellar anomaly included diffuse depletion of granular cells of the internal granular layer (IGL), remnants of the external granular layer (EGL), and disorganization of the Purkinje cell layer. These cerebellar changes were observed in all birds except one. in the infected embryos, the EGL was thicker and had an irregular arrangement with a thin molecular layer (ML) and IGL, compared with the control. The granular cells were immunohistochemically positive for ALV common antigen. immunohistochemical analysis for vimentin revealed disarrangement and decreased number of Bergmann's fibers. Use of the terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling method and electron microscopy indicated that apoptotic granular cells were frequently observed in the EGL and ML. These results suggested that the cerebellar anomaly was hypoplasia, principally resulting from the apoptosis of granular cells in the EGL and ML caused by FGV infection and that the cell loss induced obstruction of granular cell migration and disarrangement of Bergmann's fibers in the ML.
Keywords
Fowl glioma is histologically characterized by multiple astrocytomas associated with disseminated nonsuppurative encephalitis. 22 We previously reported that fowl glioma-inducing virus (FGV), which belongs to avian leukosis virus subgroup A (ALV-A), causes fowl glioma in Japanese bantam and specific-pathogen-free (SPF) chickens. 11, 18, 23, 24 The FGV shows tumorigenicity and pathogenicity mainly in the central and peripheral nervous systems, whereas ALV-A generally induces B-cell lymphomas. 8, 25 It is suggested that unique sequences of the long terminal repeat promoter and env gene of FGV are responsible for the nervous system-specific pathogenicity. 24 We have found that the cerebellar anomaly concurrently developed in chickens inoculated with FGV through the yolk sac. Nonsuppurative encephalitis in SPF chickens infected in ovo with Rous-associated virus 1 (RAV-1; a standard strain of ALV-A) or RAV-7 has been reported. 7, 28 However, to the author's knowledge, there is no report describing cerebellar hypoplasia associated with ALV.
Congenital cerebellar anomaly is one of the common congenital nervous system defects of domestic animals. 14 Cerebellar hypoplasia secondary to an in utero or perinatal viral infection most commonly is seen in cats, cattle, and pigs. 3, 5, 6 The actively dividing granular cells in developing cerebellum are especially sensitive to viral or chemical teratogenic agents. Selective necrosis of the granular cells has been regarded as an important cause of virus-induced cerebellar hypoplasia.
The objective of the study reported here was to describe the pathomorphologic changes associated with the cerebellar anomaly observed in chickens inoculated with FGV through the yolk sac.
Materials and Methods
Virus
Fertile eggs of the commercial SPF White Leghorn strain WL-M/O (C/O) were purchased from Nippon Institute for Biological Science (Yamanashi, Japan). This strain lacks chicken helper factor and group-specific antigen, and is susceptible to ALV subgroups A–E (International registry of poultry genetic stocks, Bulletin 476, March 1988, Ralph G. Somes Jr, PhD., University of Connecticut, Storrs, CT). The presence of endogenous viral genes other than the chicken helper factor and group-specific antigen in this strain is not clear. Chicken embryo fibroblasts (CEF) were prepared from the C/O chicken embryos on the 10th day of incubation. Cell culture and preparation of a viral suspension containing FGV were performed using described methods. 11 For inoculation, a 100-fold concentrated culture supernatant was prepared according to the method described by Bowles et al. 1 A resistance-inducing factor test and sequencing analysis of the proviral genome indicated that FGV belonged to ALV-A. 23, 24
Animals and experimental design
Fertile eggs from the C/O chickens used for the CEF were used in the animal infection experiment. All fertile eggs were incubated at 37.5°C, and embryos were used for inoculation on the 7th day of incubation. Seven C/O chicken embryos (group A) were inoculated through the yolk sac with 0.1 ml of the FGV suspension (Table 1). Six embryos (group B) were inoculated with 0.1 ml of the culture supernatant of uninfected CEF instead of the viral suspension used for group A. All embryos in both groups were administered sterile saline intravenously twice weekly for 18 weeks, from 2 days of age onward, because these two groups were prepared for a carcinogenic study of methylnitrosourea (MNU). Because the results of MNU carcinogenicity were negative, groups injected with MNU were not included here. The hatched chicks were reared in isolators in the animal facilities of our school, and euthanized at 70 and 140 days of age.
Experimental design and histologic findings in the cerebellum of chickens (70 or 140 days).∗


FGV = culture supernatant of chicken embryo fibroblasts (CEF) infected with fowl glioma-inducing virus; medium = culture supernatant of uninfected CEF.
Number of chickens with lesions (%).
In another experiment, chicken embryos were inoculated through the yolk sac on the 7th day of incubation with 0.1 ml of the viral suspension. The embryos on the 10th, 12th, 14th, 16th, 18th, and 20th day of incubation and chicks at 2 and 4 days of age were euthanized to examine the initial stage of the cerebellar anomaly. Three infected embryos or chicks and three control birds were used for each day.
All experiments were performed humanely according to the guidelines set by the Hokkaido University Veterinary Medicine Animal Care and Use Committee. Chickens did not receive any vaccinations or medication.
Histologic examination and immunohistochemical analysis
The brain and other organs, including the liver, spleen, kidneys, heart, lungs, bone marrow, gastrointestinal tract, pancreas, bursa of Fabricius, thymus, gonads, spinal cord, peripheral nerves, eyes, skeletal muscles, and skin were fixed in neutral-buffered 20% formalin, embedded in paraffin wax, sectioned at 4-µm thickness, and examined by light microscopy after staining with hematoxylin and eosin (HE).
For the immunohistochemical examination, selected sections were deparaffinized and sections for vimentin evaluation were pretreated with protease (0.1% trypsin in 0.1 M Tris buffer) for 30 minutes at 37°C. The sections were treated with 0.3% hydrogen peroxide to inhibit endogenous peroxidase. Indirect immunoperoxidase staining was performed using a commercial streptavidin-biotin kit (Nichirei Corp., Tokyo, Japan). After the blocking of nonspecific reactions, the sections were incubated with a specific primary antibody: antibody to either vimentin (clone Vim3B4, diluted 1 ° 100; Dako, Glostrup, Denmark) or ALV common antigen (1 ° 50,000). The polyclonal antibody directed against ALV was generated by immunization of a rabbit with RAV-2 using a described method. 26 The antibody recognizes mainly gp85 and p27 of ALV. The staining was carried out with a biotinylated secondary antibody and peroxidase-labeled streptavidin. The distribution of the antigen was demonstrated by incubation of the sections with 3,3′-diaminobenzidine-H2O2 (DAB-H2O2) solution. The sections were counterstained with hematoxylin.
Terminal Deoxynucleotidyl Transferase-mediated dUTP-biotin Nick End Labeling analysis
Selected sections were deparaffinized and pretreated by incubation with 20 µg of proteinase K/ml in 0.1 M Tris buffer for 20 minutes at room temperature, washed in deionized water twice for 2 minutes each, and returned to phosphate-buffered saline. Sections were then covered with terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL) reaction mixture (Boehringer Mannheim, Indianapolis, IN), which contained terminal deoxynucleotidyl transferase and biotin-16-dUTP, and incubated for 1 hour at 37°C. The reaction was terminated and developed in similar manner as that done for the immunohistochemical analysis.
Ultrastructural study
The cerebellum from embryos inoculated with FGV was diced into 1-mm3 cubes for electron microscopy, fixed in 2.5% glutaraldehyde buffered with 0.1 M phosphoric acid (pH 7.4), postfixed in 1% osmium tetroxide buffered with 0.1 M phosphoric acid (pH 7.4), dehydrated, and embedded in epoxy resin. Semithin sections (0.5 µm thick) were stained with toluidine blue, and appropriate areas were selected. Ultrathin sections were prepared using an ultramicrotone stained with lead citrate and uranyl acetate and examined by transmission electron microscopy (JEM-1210; JEOL, Tokyo, Japan).
Results
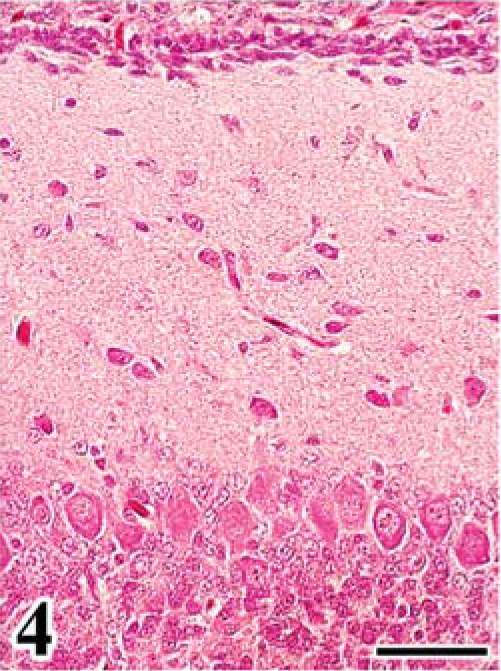
In group A, leg paralysis was noted in 2 chickens, and a large renal mass (4 cm in diameter) was evident in one of them. At necropsy, in all FGV-infected chickens except one, the cerebellum was mildly decreased in size, compared with that of chickens in group B. Common histologic findings observed in the brain of chickens in group A were a hypoplastic cerebellum and variably sized fowl glioma (Table 1). Fowl glioma was observed in the cerebrum of all birds of group A and in the cerebellum of 1 bird of group A. In severe cases, the structure of the cerebellum was almost destroyed (Fig. 1). The internal granular layer (IGL) in the cerebellum of FGV-infected chickens had thinned because of focal-to-diffuse depletion of granular cells (Fig. 1). A few granular cells accumulated directly beneath the pia mater, suggesting a remnant external granular layer (EGL) (Fig. 2). Purkinje cells (PCs) were severely disarranged and irregularly scattered in the molecular layer (ML) and IGL. These cerebellar lesions, including depletion of granular cells in the IGL, remnant of EGL, and disorganization of the Purkinje cell layer (PL), were observed in six (86%), five (71%), and six (86%) birds of group A (Table 1). One bird of group A did not have lesions in the cerebellum. Multifocal gliosis and vascularization with collagenous proliferation were occasionally observed in the affected cerebellum. Inflammation was not apparent in the central nervous system (CNS), except for a multifocal perivascular cuffing of lymphocytes and plasma cells. In the cerebellum of embryos on the 20th day of incubation, the EGL had thickened, with irregular cell alignment, and the ML was thin, compared with that of the controls (Figs. 3, 4).
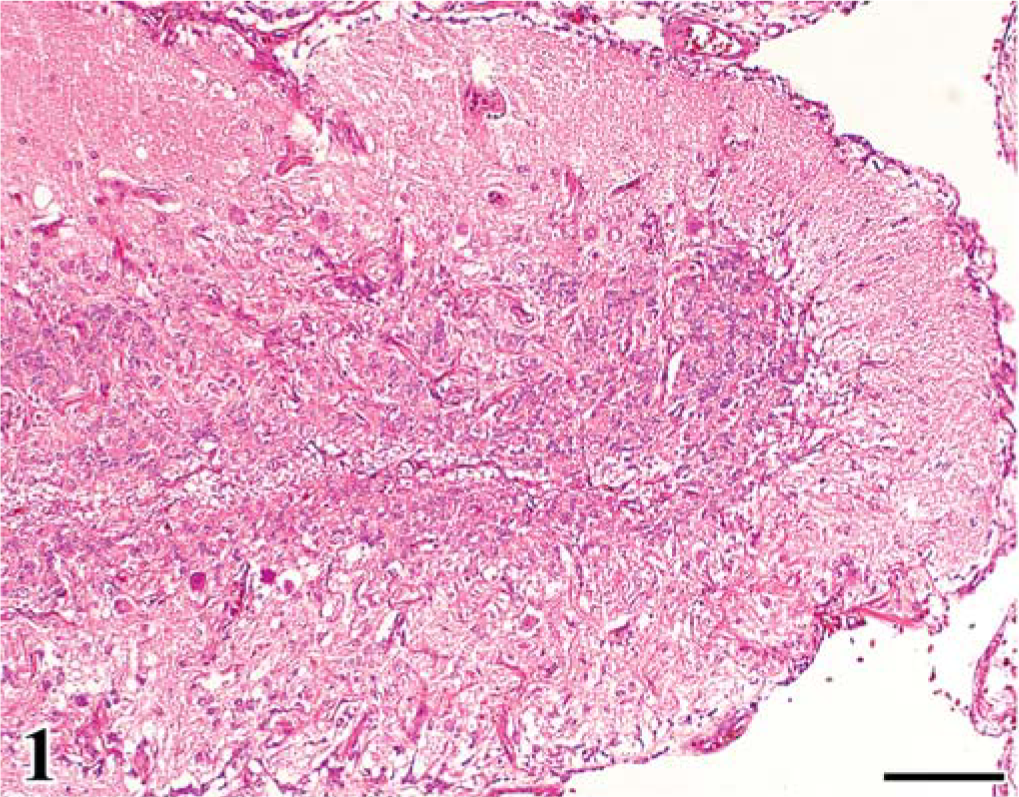
Cerebellum; chicken of group A, infected with FGV, 140 days of age. The cerebellar cortex shows diffuse absence of granular cells of the internal granular layer, irregular thickness of the molecular layer, and disorganization of the Purkinje cell layer. HE. Bar = 100 µm.
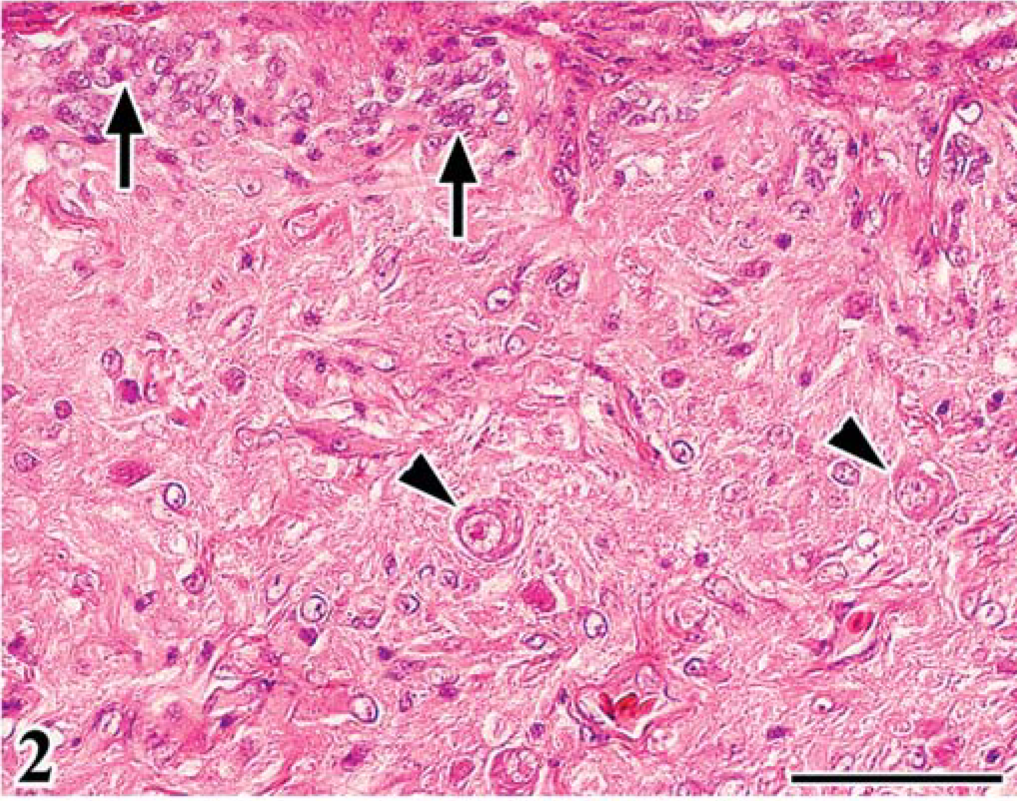
Cerebellum; chicken of group A, infected with FGV, 70 days of age. Notice remaining external granular layer (arrows) and displaced Purkinje cells (arrowheads) in the molecular layer. HE. Bar = 50 µm.
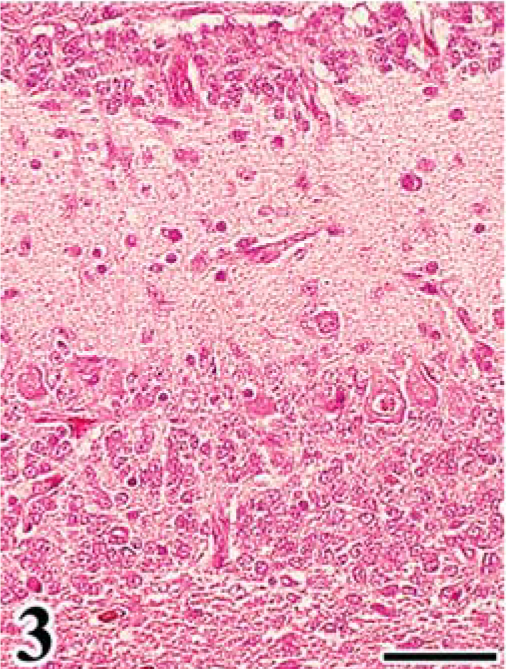
Initial stage of cerebellar anomaly in chick embryo. Cerebellum; chick embryo, infected with FGV, 20th day of incubation. The cerebellar cortex shows a thickened external granular layer with irregular cell alignment and thin molecular layer. HE. Bar = 50 µm.
Scattered astrocytes and granular cells were immunohistochemically positive for the ALV common antigen in the affected cerebellum, and atypical cells of astrocytoma were consistently positive. In the infected embryos, reactivity for the ALV common antigen was detected mainly in granular cells of the EGL and IGL after the 16th day of incubation (Figs. 5, 6, Table 2). In both experiments, PCs were negative for ALV common antigen. None of the control embryos had a positive reaction for the ALV common antigen in any organ. By use of immunohistochemical analysis for vimentin Vim3B4, a decreased number and disordered arrangement of Bergmann's fibers, which are glial processes radially extending toward the pial surface from Golgi epithelial cells (Bergmann's glia) in the PL, were seen in the infected embryos (Fig. 7), whereas the characteristic palisading structure of Bergmann's fibers was clearly detected in control embryos (Fig. 8).
Immunohistochemical analysis for ALV common antigen in cerebrum, cerebellum, heart, and kidney of embryos and chicks inoculated with FGV.∗
E = embryo days; PI = postinoculation (days); + = ALV common antigen-positive cells were observed; − = negative results.
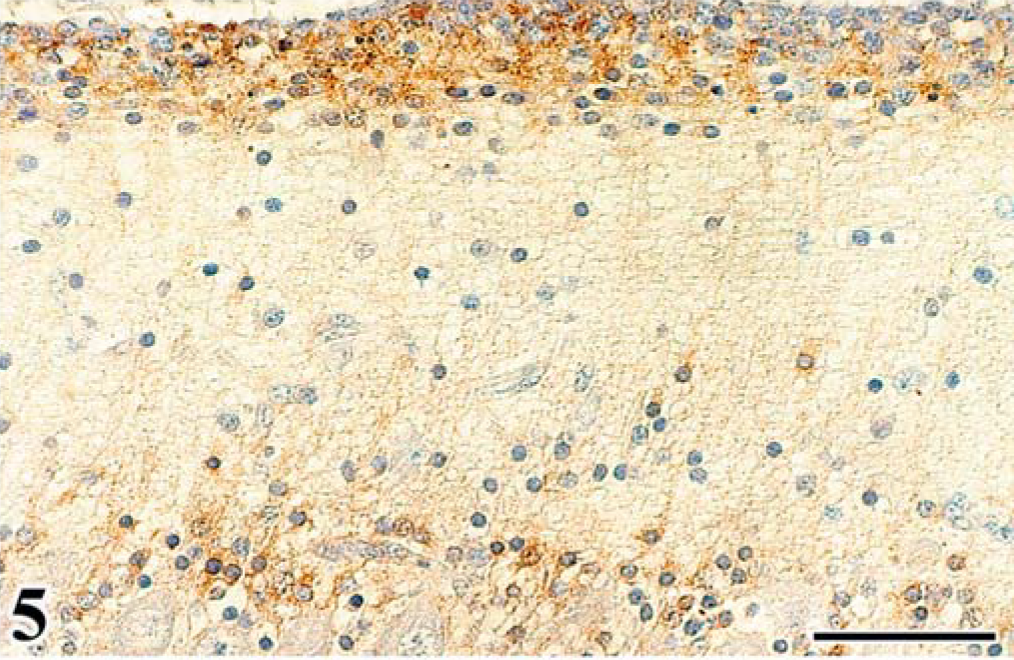
Immunohistochemistry for ALV common antigen. Cerebellum; chick embryo, infected with FGV, 18th day of incubation. Granular cells in the external granular layer are positive for ALV common antigen. Streptavidin-biotin-3,3′-diaminobenzidine-H2O2 (SAB-DAB-H2O2), hematoxylin counterstain. Bar = 50 µm.
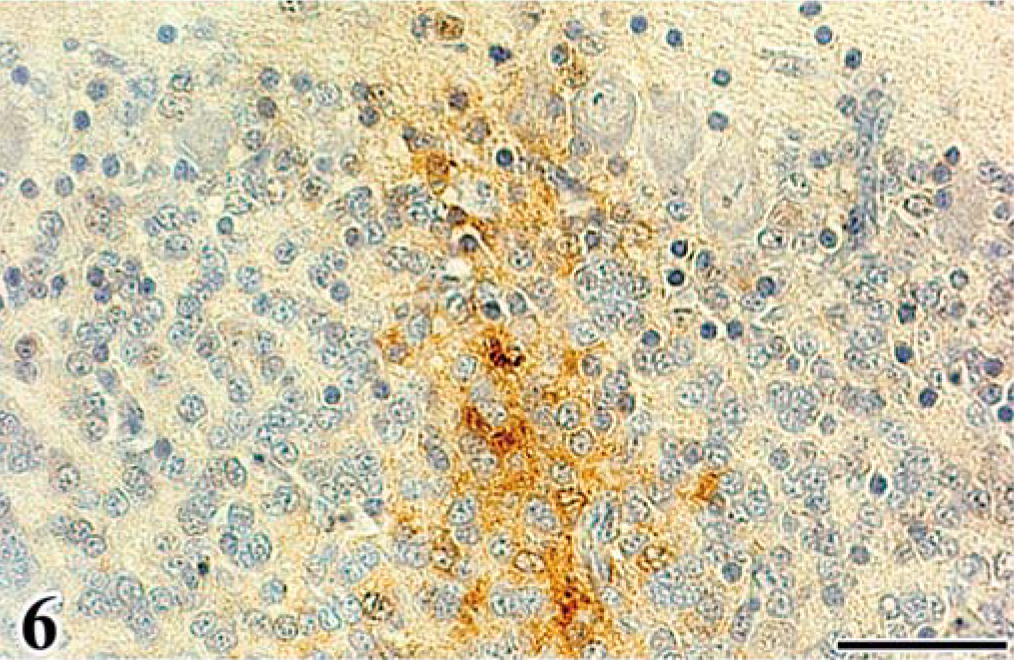
Immunohistochemistry for ALV common antigen. Cerebellum; chick embryo, infected with FGV, 18th day of incubation. Granular cells in the internal granular layer are positive for the antigen. SAB-DAB-H2O2, hematoxylin counterstain. Bar = 40 µm.
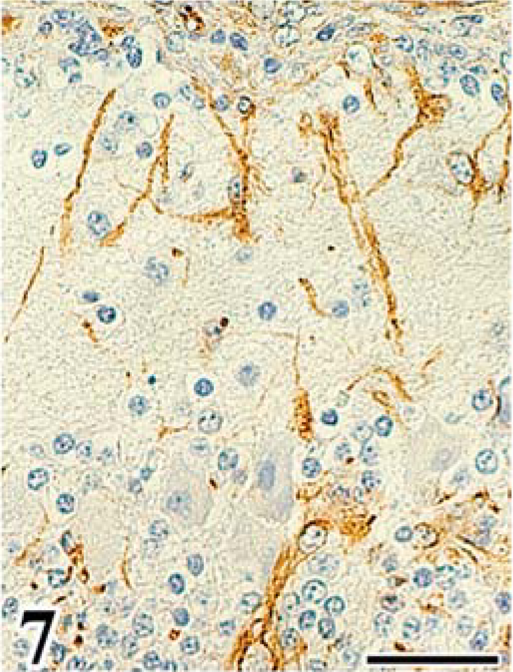
Immunohistochemistry for vimentin Vim3B4 in cerebellum of chick embryo. Cerebellum; chick embryo, infected with FGV, 20th day of incubation. Decreased numbers and irregular arrangement of Bergmann's fibers. SAB-DAB-H2O2, hematoxylin counterstain. Bar = 30 µm.
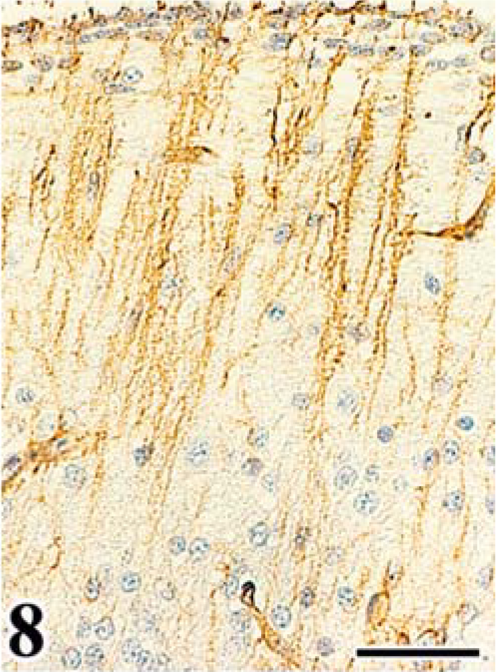
Immunohistochemistry for vimentin Vim3B4 in cerebellum of chick embryo. Cerebellum; chick embryo, uninfected control, 20th day of incubation. Bergmann's fibers showing a normal palisading structure. SAB-DAB-H2O2, hematoxylin counterstain. Bar = 40 µm.
FGV apoptosis was evaluated in the affected cerebellum by detection of DNA fragmentation using the TUNEL method and electron microscopy. Granular cells of the EGL and ML often had TUNEL-positive nuclei (Fig. 9), and these cells exhibited morphologic changes typical of apoptosis (apoptotic bodies), whereas TUNEL-positive cells were rare in the control embryos (Fig. 10). The TUNEL-positive cells were most frequently detected in the affected cerebellum on the 18th day of incubation.
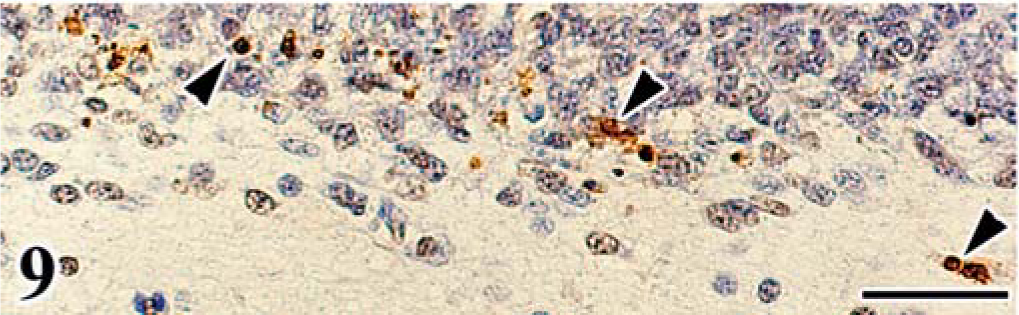
Apoptosis detected by the TUNEL method in cerebellum of chick embryo. Cerebellum; chick embryo, infected with FGV, 20th day of incubation. TUNEL-positive granular cells (arrowheads) are frequently detected in the external granular layer and molecular layer. Hematoxylin counterstain. Bar = 30 µm.
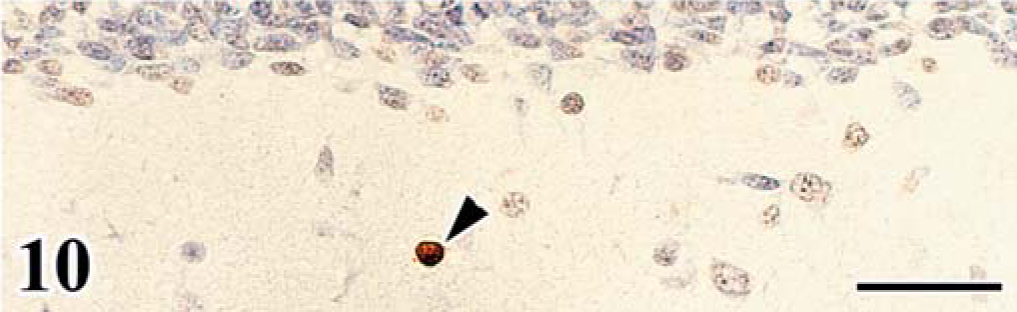
Apoptosis detected by the TUNEL method in cerebellum of chick embryo. Cerebellum; chick embryo, uninfected control, 20th day of incubation. TUNEL-positive cells (arrowhead) are rarely detected in the cerebellar cortex. Hematoxylin counterstain. Bar = 30 µm.
Ultrastructural changes typical of apoptosis were detected in the EGL and ML of the affected cerebellum. Apoptotic granular cells had condensation of chromatin, membranous degeneration, and rupture of nuclei.
Six (86%) chickens of group A had mild-to-moderate nonsuppurative myocarditis with matrix inclusions, which are intracytoplasmic inclusions in infected cardiac myocytes. 9, 12 Osteopetrosis was detected in 2 (29%) chickens of group A, and perineurioma and renal myxoid sarcoma were observed in 1 bird each (14%). There were no significant lesions in the other organs and tissues examined. Clinical signs of disease or microscopic lesions were not observed in chickens of group B.
Discussion
Most of the chickens inoculated with FGV in ovo developed mild-to-severe cerebellar anomaly, including diffuse depletion of granular cells of the IGL, a remnant of the EGL, and disorganization of the PL. In the infected embryos, the EGL was thick with a disturbed arrangement, and formation of the ML and IGL was delayed, compared with that of the control. After the 16th day of incubation of infected embryos, granular cells in the EGL and IGL were immunohistochemically positive for ALV common antigen. By use of immunohistochemical analysis for vimentin Vim3B4, decreased numbers and disordered arrangement of Bergmann's fibers were recognized. The TUNEL method and electron microscopy frequently revealed apoptosis of granular cells in the EGL and ML. These results suggested that the cerebellar changes were considered hypoplasia principally resulting from the apoptotic loss of granular cells infected with FGV in the embryonic stage.
The development of the cerebellar cortex proceeds in a similar order in various vertebrates. 13 The PCs and Golgi epithelial cells in chick embryos originate in the rhombic lip, and migrate to form an irregular layer from the 3rd to the 6th day of incubation. 29 The EGL is formed between the 6th and 15th day of incubation. 10, 27 The granular cells of the EGL migrate through the ML to form the IGL between the 18th day of incubation and 2 days of age when it peaks (Fig. 11). Bergmann's fibers act as guides for the migration of granular cells during this process. 17, 20 The granular cells pile up underneath the PCs, and parallel fibers, which are granular cell axons that have a trajectory parallel with the axis of the folium, form a taut barrier above them; then the PCs become mechanically sandwiched between these two growing layers and are forced to form a monocellular sheet. The results of immunohistochemical analysis, and TUNEL and electron microscopy in this study suggested that FGV infected the growing granular cells and induced apoptosis to inhibit cell migration. The delay of granular cell migration caused the thickening and irregular arrangement of the EGL. Furthermore, the formation of the ML that is based on accumulation of parallel fibers could become insufficient by the delay of migration. In the affected cerebellum, the palisading structure of Bergmann's fibers was collapsed and the retarded growth of the IGL and ML was considered to have caused the disorganization of the PL by inhibiting it from being single layered. 19
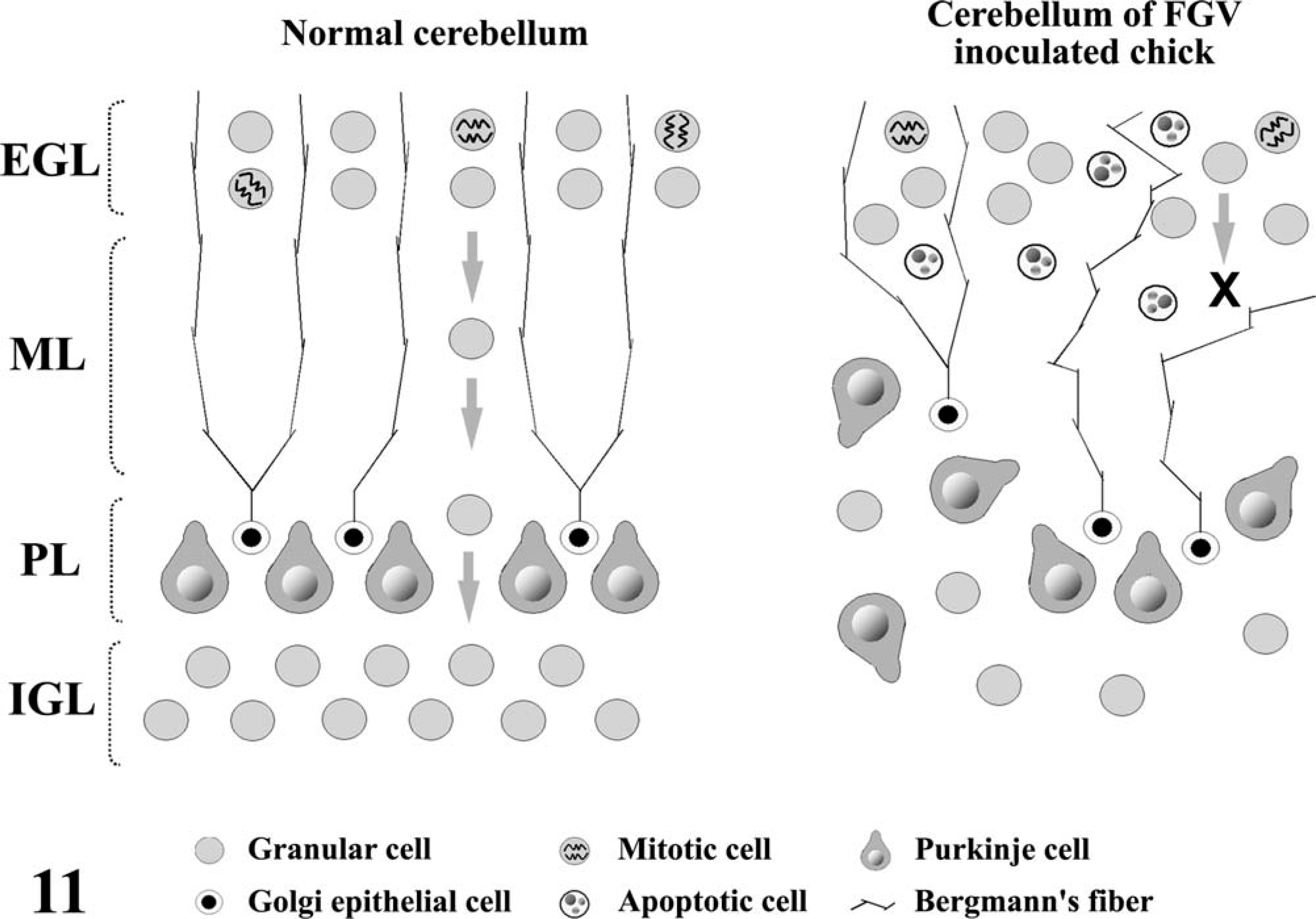
Schematic diagram illustrating the histogenesis of the cerebellar cortex in normal and FGV-infected chickens.
Although some ALVs showing pathogenicity in the CNS have been reported, these ALVs induce a nonsuppurative encephalomyelitis characterized by diffuse mononuclear cell infiltration of the brainstem parenchyma and perivascular cuffing. 7, 28 On infection in ovo, RAV-1 replicates in the granular layer of the cerebellum and choroid plexus. 7 In the study reported here, although mild perivascular cuffing of lymphocytes and plasma cells was detected in the cerebellum of chickens at 70 or 140 days of age, the frequency (33–50%) and severity of the lesions were low. Active inflammatory reaction was not observed in the cerebellum of embryos inoculated with FGV.
In veterinary medicine, feline panleukopenia virus (FPV), bovine viral diarrhea virus (BVDV), and hog cholera virus (HCV) are known as causative agents of cerebellar hypoplasia. 14 The FPV-induced cerebellar hypoplasia in kittens is characterized by depletion of granular cells and disorganization and loss of the PCs. 4, 5, 16 Granular cells of the EGL are most susceptible to FPV infection in the perinatal period, because FPV has a predilection for actively dividing cells. On the basis of these findings, the necrosis of the EGL infected with FPV is considered the main cause of the cerebellar hypoplasia. Furthermore, it is speculated that FPV also may have the ability to infect the PCs, which have already finished division at the time of the EGL formation, thus causing disorganization of the PL. 5
The BVDV-induced cerebellar lesion in newborn calves is characterized by loss of cerebellar cortex and cavitation in the white matter. 2, 3, 15, 21 The acute nonsuppurative inflammation predominately in the cerebellar cortex with granular cell necrosis has been thought to be the cause of the cerebellar lesions. 3 Vascular interruptions are suspected as the cause of the edema and hemorrhage in the cerebellum affected with BVDV. 3
In pigs, HCV infection results in cerebellar hypoplasia and degeneration as well as hypomyelinogenesis. The principal cause of the cerebellar lesions is considered hypoxia due to vasculitis. 6 The HCV also could infect endothelial cells of blood vessels and induce degeneration and necrosis of them.
In the study reported here, the negative reactivity of the PCs for ALV common antigen suggests that disorganization of the PL occurs secondary to the retarded growth of the ML and IGL, not to direct infection of the PCs with FGV. Although FGV could infect endothelial cells of blood vessels, it is suggested that FGV fails to cause the collapse of vessels because of the absence of edematous lesions in the cerebellum of embryos and chickens inoculated with FGV. 23
In conclusion, FGV is a unique avian leukosis virus that causes cerebellar hypoplasia in chickens infected in ovo, although more investigation about the mechanism of FGV-induced apoptosis of granular cells in the cerebellum is needed to clarify the neuropathogenicity of FGV.