
Abstract
Light and electron microscopic features and immunohistochemical features of Cryptosporidium andersoni (C. andersoni) and host reaction in the mucosa were studied. Although the affected cattle demonstrated no apparent clinical signs, a severe infection of C. andersoni was observed in the abomasum. C. andersoni were round in shape, measured 6-8 μm in size and were mainly observed to be freely located in the gastric pits, being attached in occasional cases to the surface of the abomasum epithelium. Frequent inflammatory cells had infiltrated the lamina propria of the affected mucosa, and frequent mitotic figures were observed in epithelial cells at the dilated isthmus. To access the cell kinetics, the number of epithelial cells infected with C. andersoni were counted and compared with noninfected cattle. The number of gastric pit cells in infected cattle was significantly higher than that in the controls. The number of proliferative cells determined by the Ki-67 antigen in C. andersoni infected cattle was also significantly higher than that in the controls. Transmission electron microscopy and scanning electron microscopy revealed that the morphology of the C. andersoni organism was common to those of other Cryptosporidium spp. immunohistochemically, several commercial antibodies against Cryptosporidium spp. showed positive reactions at the wall of these oocysts or parasitophorous vacuoles. This report is possibly the first to discuss the prominent hyperplasia of the abomasum mucosa, as well as morphologic features of C. andersoni in cattle.
Genus Cryptosporidium is an intracellular protozoan parasite and causes gastrointestinal disease in a wide variety of mammals and other vertebrate species worldwide. 1, 9, 11 Cryptosporidium spp. are transmitted by the ingestion of oocysts that are excreted in the feces of humans and animals. 9, 14, 17, 22 In cattle, three subspecies of Cryptosporidium have been reported: Cryptosporidium parvum, 9, 10, 20 Cryptosporidium felis, 3 and Cryptosporidium andersoni, recently renamed from Cryptosporidium muris. 2, 4, 8, 10, 12, 13, 18– 20
Cryptosporidium resides on the apical surface of intestinal epithelial cells. It is viewed as a minimally invasive, mucosal pathogen, which causes infection at the microvillus border of the gastrointestinal epithelium. However, infection elicits a cell-mediated response following both primary and secondary infections. 9 Some Cryptosporidium spp. cause acute diarrhea in immunocompetent animals and chronic life-threatening disease in immunocompromised animals. 15, 23
Many studies have been conducted on morphology, immunohistochemistry and epidemiology for some species of Cryptosporidium, particularly C. parvum. 5, 9 C. andersoni is a newly established member of Cryptosporidium spp., and there have been few reports on morphologic studies in cattle, including histopathological and ultrastructural features, as well as immunohistochemical reactivity. Furthermore, there has been no study on the histopathological alterations in the mucosa of host animals.
In the present study, we attempted to clarify the histopathological characteristics of host reactions in the mucosa, as well as morphological characteristics of C. andersoni, by using light and electron microscopy.
Eight Japanese black hair cattle with or without fecal excretion of Cryptosporidium spp. were examined. Four cattle (Nos. 1 and 3–5) were obtained from the farm of Tohoku University in Kawatabi, Miyagi, Japan. The other cattle (Nos. 2 and 6–8) were kept at dairy farms in Gifu Prefecture and were sent to Gifu University. In four cattle maintained at the farm of Tohoku University, oocysts of Cryptosporidium. spp. were collected from fecal samples using the sucrose flotation method and were identified as C. andersoni by sequencing the 18S ribosomal DNA, heat-shock protein 70 and oocyst wall protein genes. 18 Oocysts of C. andersoni were defecated for at least 14 months in cattle No. 1; 31 months in cattle Nos. 3 and 4; and 35 months in cattle No. 5 before euthanization, respectively (Table 1). The cattle were euthanatized, and complete necropsy was done. Organs and tissues, including the liver, spleen, kidneys, heart, lungs, rumen, reticulum, omasum, abomasum, ileum, cecum, colon, and adrenal glands, were collected and fixed in 10% buffered formalin and embedded in paraffin. For the abomasum, three samples were collected from the anterior, middle, and posterior parts. Specimens of abomasum from three Japanese black hair cattle with no lesions were used as a control (Nos. 6–8). Each of the sections was cut at a thickness of 4 µm, stained with hematoxylin and eosin (HE), and examined by light microscopy. In the three infected cattle (Nos. 3–5) and the three control cattle (Nos. 6–8), the number of cells was counted at 10 gastric pits per cow.
Summary of the clinical data of cattle.
∗ M = male; F = female; ND = not done.
Samples of the abomasums from three cattle (Nos. 3–5) were prepared for transmission electron microscopy (TEM). The specimens were rinsed in phosphate buffer, postfixed with 1% buffered osmium tetroxide, dehydrated in ethanol and propylene oxide, and embedded in epoxy resin. Individual samples were sectioned at 1 µm and stained with toluidine blue. Areas where C. andersoni were present were selected by light microscopy, and thin-cut sections, mounted on copper grids, were stained with lead citrate/uranyl acetate and examined with TEM (Hitachi, Tokyo, Japan, H-8100). Samples of formalin-fixed abomasum from one cow (No. 3) were washed in phosphate buffer and directly examined with scanning electron microscopy (SEM) (Hitachi, Tokyo, Japan, S-3000N) under low-vacuum conditions (150 Pa).
Immunohistochemical stain was determined in four of the infected cattle (Nos. 1, 3–5) and three of the control cattle at the anterior part of the abomasum. The stain was performed on prepared slides using the avidin-biotin-horseradish peroxidase (ABC) method. The primary antibodies used were mice monoclonal antibodies against Cryptosporidium parvum oocyst (Chemicon, Temecula, CA), Cryptosporidium parvum (Argene, Varilhes, France), and Cryptosporidium (VMRD, Pullman, WA). For analysis of cell kinetics at the affected sites, immunohistochemistry for Ki-67 antigen was conducted, and examination was made by light microscopy. In the three infected cattle (Nos. 3–5) and three control cattle (Nos. 6–8), the total number of epithelial cells and the number of Ki-67-positive epithelial cells were counted at 10 gastric pits per cow, and the percentage of Ki-67-positive epithelial cells in the three infected cattle was compared with the three noninfected controls. The number of gastric pit cells and the incidence of Ki-67-positive cells in the affected cattle and noninfected control cattle were expressed as averages ± SD and compared using the Wilcoxon signed rank test. Differences between groups were considered significant if the probability values evinced under 0.01.
Grossly, no abnormalities were observed in any organs or tissues, including the gastrointestinal tract, in any case examined.
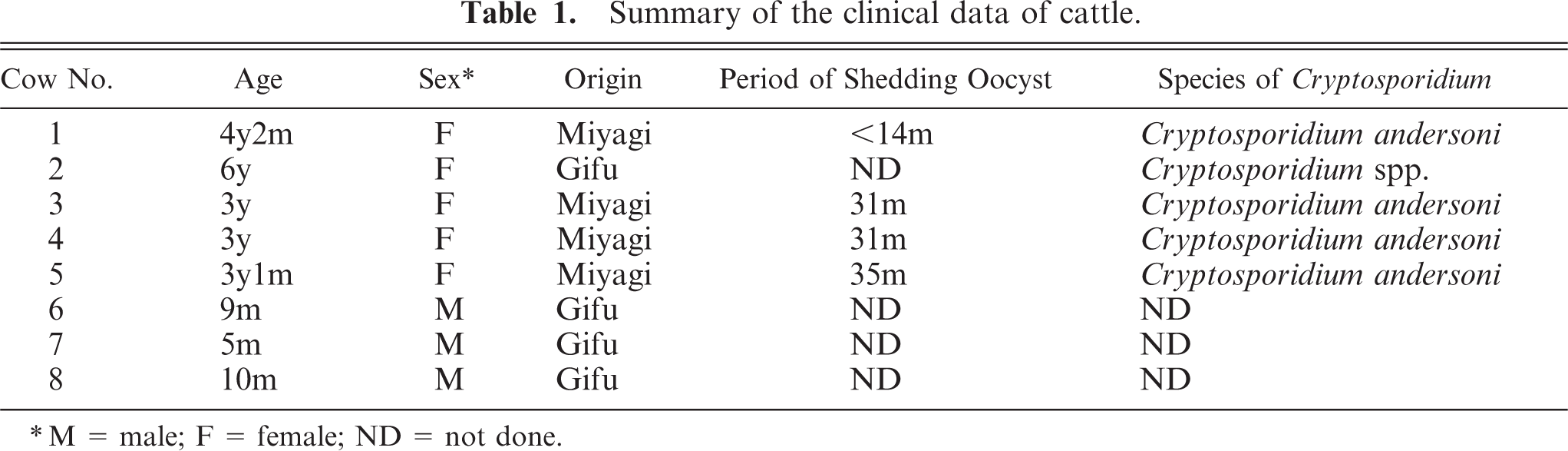
Histopathologically, no abnormalities were detected except in the abomasum. In the abomasum of all five positive cases, numerous weakly basophilic, round-to-oval organisms measuring 6–8 µm in size were observed on the apical surface of the epithelium in the gastric pits at the anterior and middle parts (Fig. 1). The length of the gastric pits was significantly extended, and the number of epithelial lining cells had increased in comparison with the noninfected control cattle (Fig. 2a, 3a). The average number of gastric pits cells for three C. andersoni infected cattle (291.9 ± 55.3 SD) was found to be significantly greater than that of the three control cattle (146.7 ± 29.2 SD). The level of significance was less than 0.01 in nonparametric testing. Many fragment nuclear epithelial cells were observed at the isthmus of the gastric pits (Fig. 1). In the lamina propria of the affected mucosa, mild-to-moderate diffuse infiltration of lymphoid cells, plasma cells, and eosinophils and mild edema were observed (Fig. 1). There were no lesions in the mucosal epithelium of the posterior part of the abomasum.
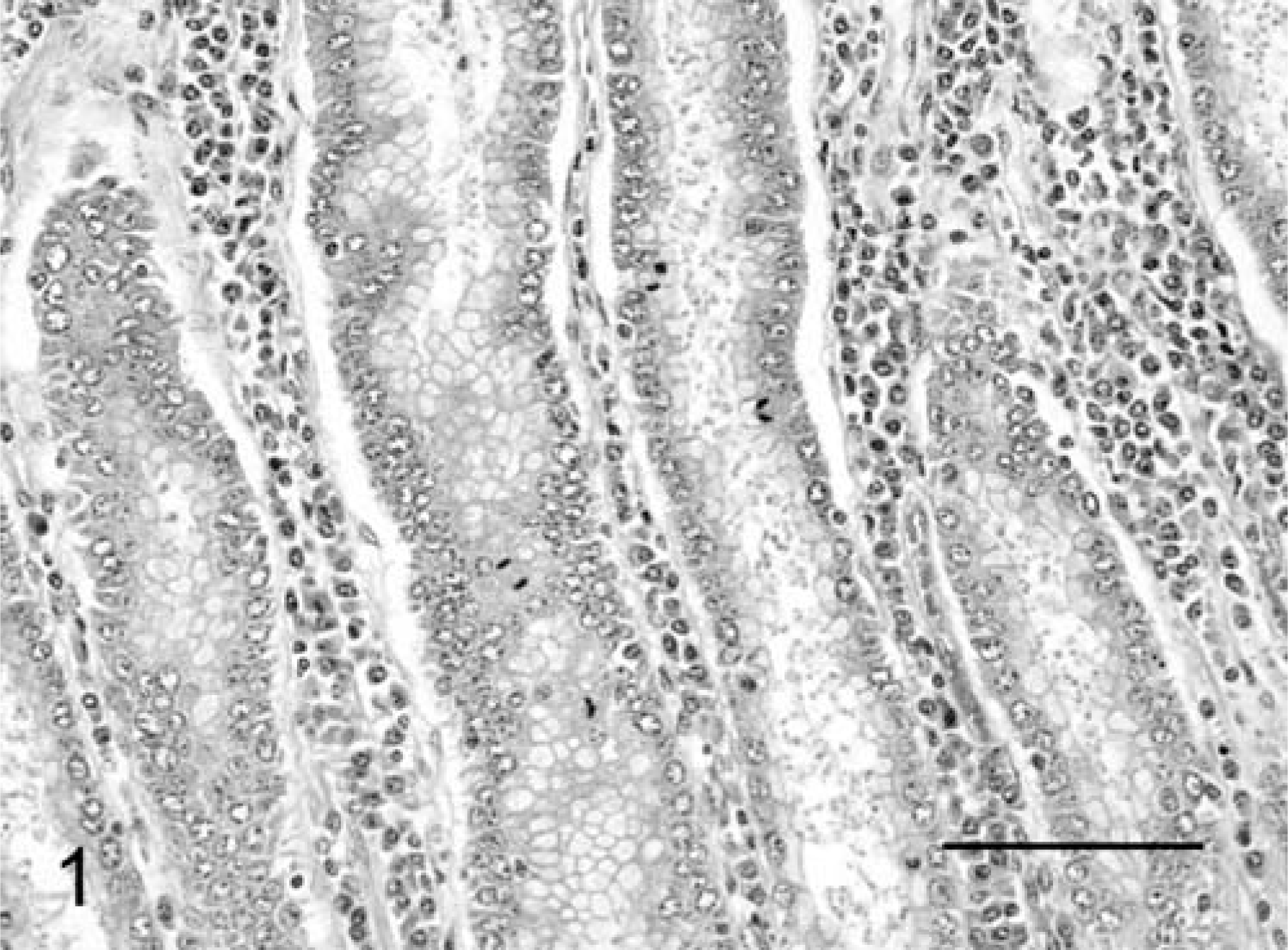
Abomasum; cow No. 3. Numerous C. andersoni in gastric pits with diffuse infiltration of inflammatory cells in the lamina propria, and mitotic figures of epithelial cells. HE, paraffin section. Bar = 100 µm.
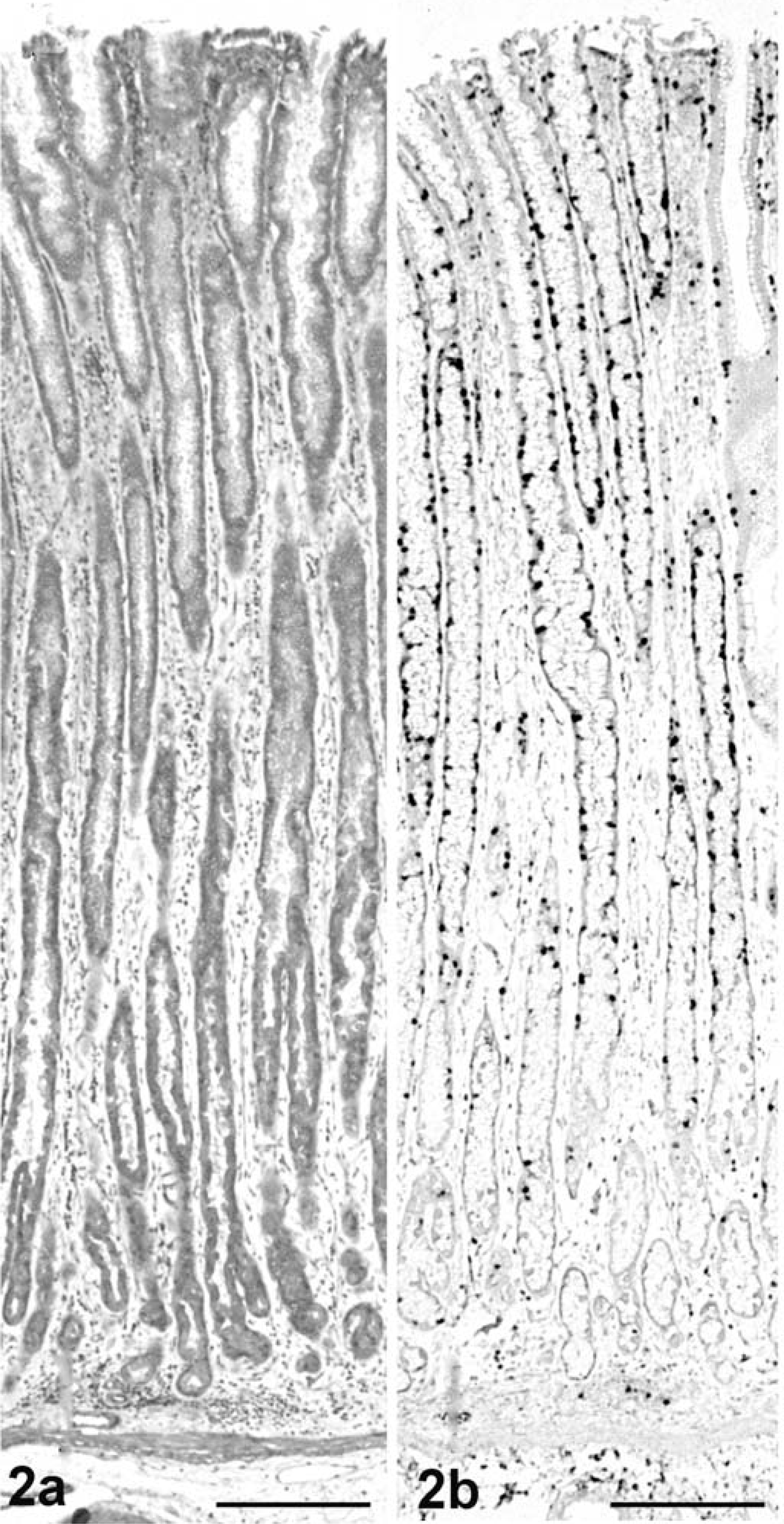
Abomasum; cow No. 5. The length of the gastric pits is greatly extended because of increase in the number of epithelial cells. HE, paraffin section. Bar = 200 µm.
Abomasum; cow No. 6. No lesions. HE, paraffin section. Bar = 200 µm.
TEM revealed various life-cycle stages of C. andersoni on the epithelial surface of the gastric pits. Trophozoites enveloped the microvilli lining on the surface of the mucosal epithelium in the gastric pits, and were located within a parasitophorous vacuole. They exhibited well-developed feeder organelle and a rough endoplasmic reticulum in the cytoplasm (Fig. 4). Generally, schizonts contained eight merozoites within a parasitophorous vacuole (Fig. 5).
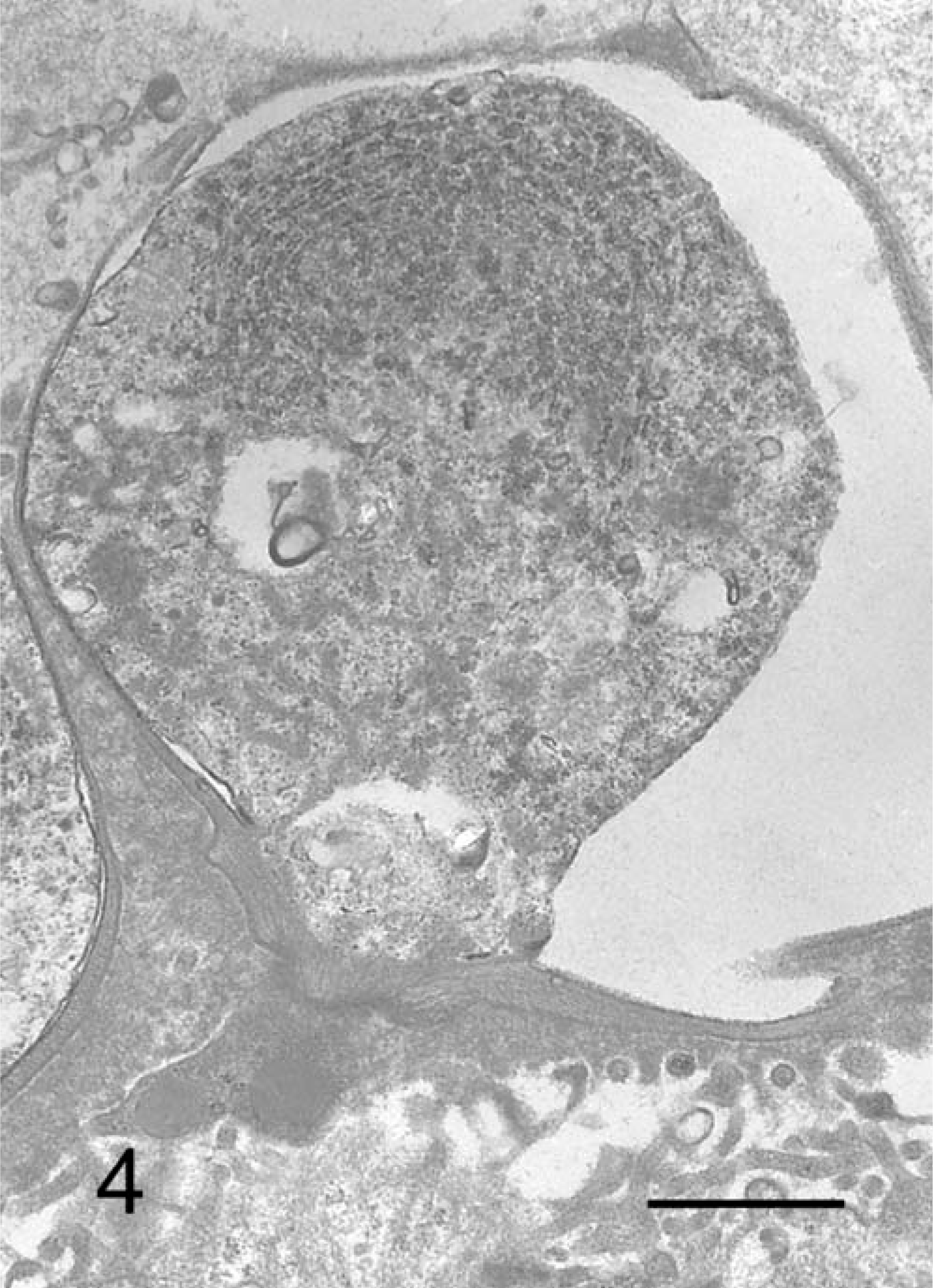
C. andersoni; cow No. 3. Trophozoite attaching to the epithelial cells of gastric pits with large parasitophorous vacuole. TEM. Bar = 1 µm.
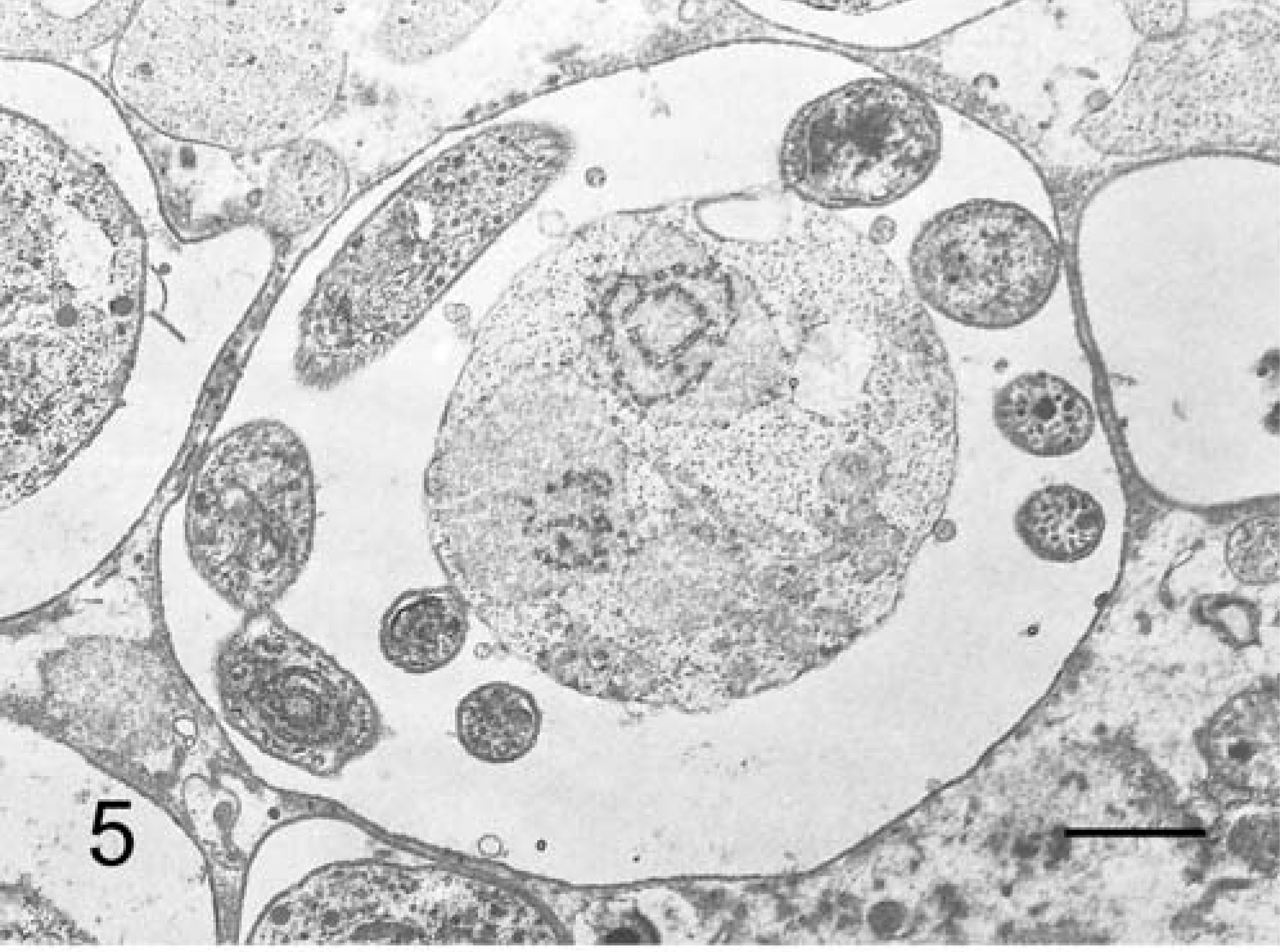
C. andersoni; cow No. 3. A mature shizont with eight merozoites in the parasitophorous vacuole. TEM. Bar = 1 µm.
Under SEM, numerous round-to-spherical organisms consistent with C. andersoni were observed on the surface of the abomasum mucosal epithelium (Fig. 6). Occasionally, hatching organisms of C. andersoni and the remaining shells were observed.
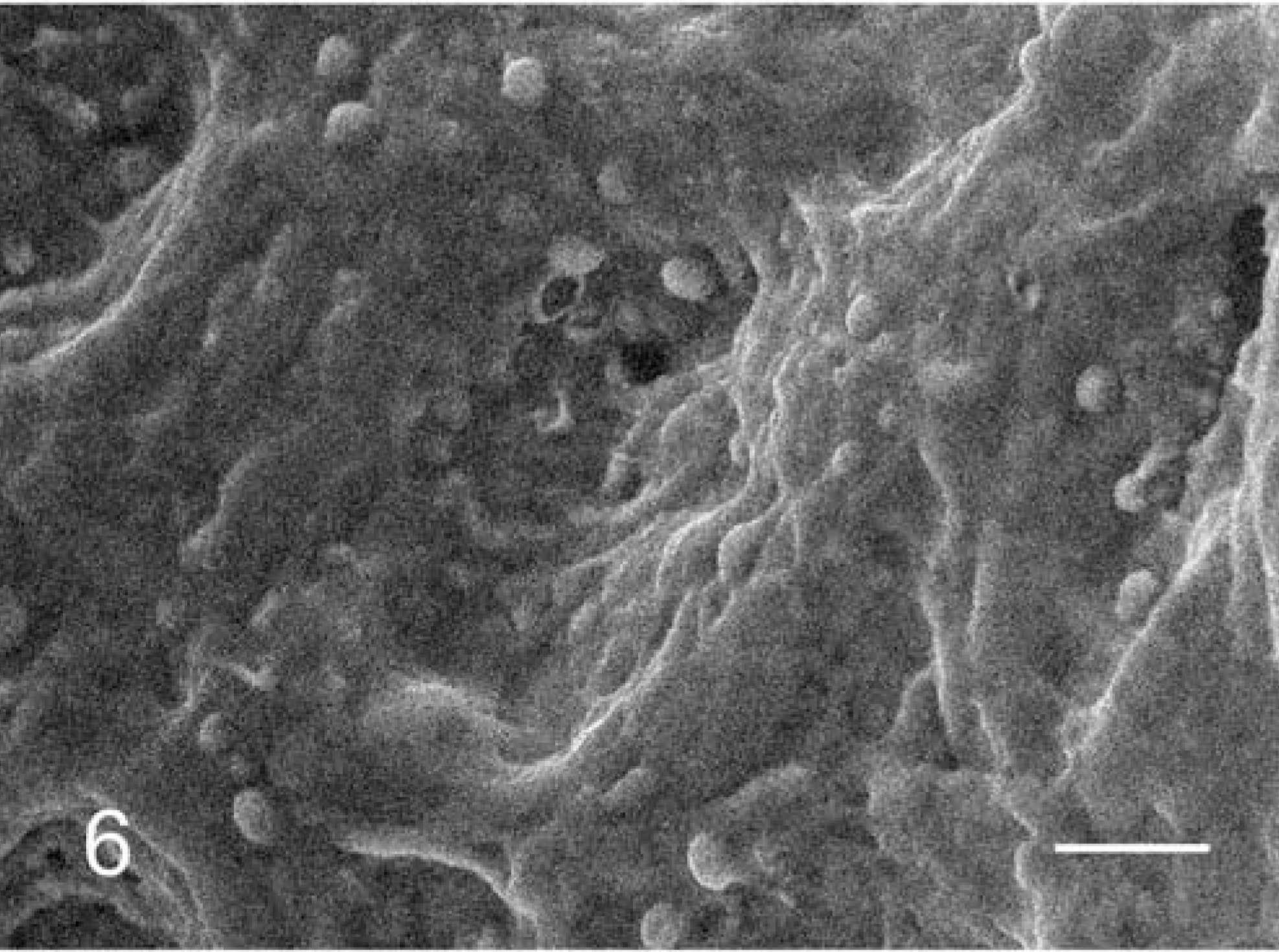
Abomasum; cow No. 3. Large numbers of C. andersoni on the epithelial cells. SEM. Bar = 20 µm.
Immunohistochemically, in the infected cattle that were examined (Nos. 1 and 3–5), most of the weakly basophilic, oval organisms in the gastric pits showed a positive reaction for primary antibodies against Cryptosporidium parvum oocyst (Chemicon, Temecula, CA), Cryptosporidium parvum (Argene, Varilhes, France) and Cryptosporidium (VMRD, Pullman, WA) (Fig. 7). Those organisms that were reactive to antibodies were larger in size than those observed with HE stain. The Ki-67 antigen–positive proliferation cells were clearly identifiable by their brown nuclei. A large number of epithelial cells showed positive reaction in the cervix portion of the gastric pits (Fig. 2b). The length of the cervix portion of the gastric pits was largely extended in comparison with the noninfected control cattle (Figs. 2b, 3b). The average number of Ki-67-positive index in the gastric pits in the three cattle with C. andersoni infection was 56.2% ± 11.1 SD, and the average number in the corresponding control cattle was 47.2% ± 16.9 SD. The average frequency of the proliferative cells in the gastric pits of infected cattle was significantly higher than that of the controls. The level of significance was less than 0.01 in nonparametric testing.
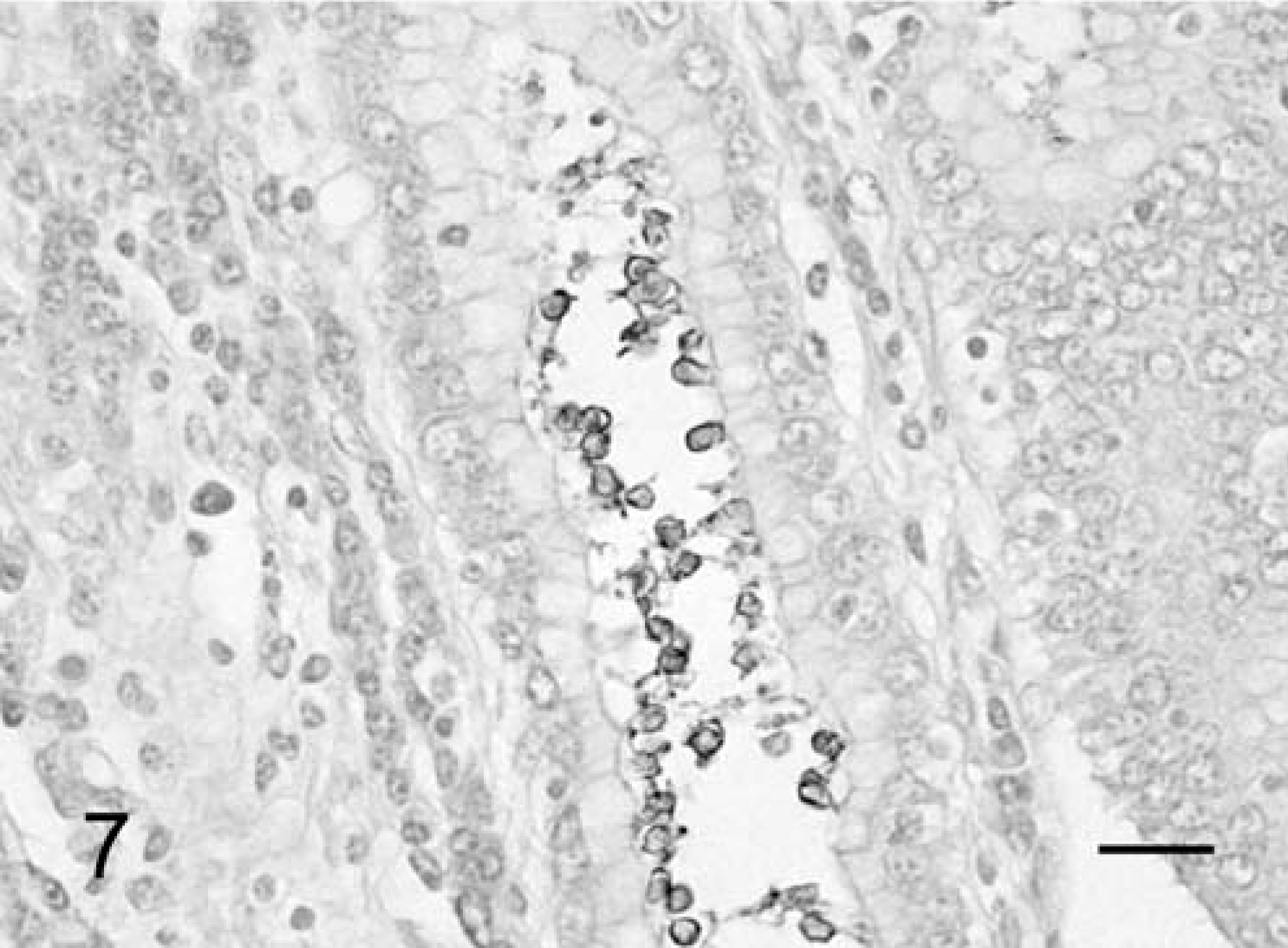
Abomasum; cow No. 3. Only the wall of parasitophorous vacuoles and oocysts of C. andersoni positive for primary antibody. Immunohistochemical stain for Cryptosporidium parvum oocyst (Chemicon, Temecula, CA), paraffin section. Bar = 20 µm.
HE sections revealed that C. andersoni infection was specifically limited to the mucosa of the abomasum in the present study. On the other hand, infection caused by other Cryptosporidium spp., such as C. parvum, showed up in the small intestine, particularly in the ileum. Unlike C. parvum, C. andersoni and C. muris were thought to have different infection sites. This is a characteristic feature of these species. Furthermore, on the same abomasum, there was a clear predisposition to infection of C. andersoni, which was located in the anterior and middle parts of the abomasum. These predisposed infection sites in the abomasum may be dependent on environmental factors, such as favored pH, ion balance, or the characteristics of the host cells, but detailed factors were not fully clarified. 4, 6
As with the in vitro examination of C. parvum, C. andersoni 4, 6, 9 showed increased nuclear fragmentation and enhanced apoptosis in the epithelial cells of the affected area in the present study. It was hypothesized that the cell division served as defense mechanisms or host response against attachment and invasion by C. andersoni organisms.
The length of the gastric pits was extended as a result of the accumulation of increased epithelial cells in the cervix portion, and the proliferation portion was not the principalis of the gastric pits, as it is in monkeys with Cryptosporidium muris-like infections. 7 The number of the gastric pit cells in cattle with C. andersoni had significantly increased (P < 0.01). This increase in the number of cells could conceivably correlate strongly with the larger frequency of Ki-67-positive cells. This result indicated that the extension of the isthmus of the gastric pits might be caused by the increasing number of cells as a consequence of the high frequency of cell divisions.
A moderate degree of infiltration of lymphocytes, plasmacytes, and eosinophils was found in the lamina propria in the affected abomasum. The severity of the inflammatory cell reaction in the lamina propria was more intense in infections caused by other Cryptosporidium spp., especially Cryptosporidium parvum, than in those caused by C. andersoni. 6, 8, 9, 16, 21 This relatively mild host reaction might relate to the long term colonic infection.
The ultrastructural features of C. andersoni were similar to those in other Cryptosporidium spp. for the most part, 7, 9 except for the size of the parasitophorous vacuole. The size of the C. andersoni parasitophorous vacuole was larger than that in other Cryptosporidium spp. This may be one of the characteristic features of C. andersoni on TEM observation. SEM revealed numerous C. andersoni organisms with a spherical shape attached to the surface of the mucosal epithelium of the abomasum. We concluded that C. andersoni mainly infected the apical surface of the pits of the abomasum and then grew and propagated in the abomasum. 5
Immunohistochemically, most C. andersoni organisms were reactive to all primary antibodies examined. However, C. andersoni with positive reactivity by immunohistochemistry were larger than those examined with HE stain. This discrepancy might be due to the presence of parasitophorous vacuole around the organisms. TEM observation showed that the size of the parasitophorous vacuole of C. andersoni was much larger than the size of the nucleus and cytoplasm, which might account for the discrepancy in size between the HE stain and immunohistochemical stain.
In conclusion, most histological and ultrastructural features of C. andersoni organisms and its mechanisms of attachment to epithelial cells might be common on the whole to those in other Cryptosporidium spp. However, the host response was different. Only in C. andersoni mucosal hyperplasia was observed.9a It is unknown whether the same phenomenon occurs in other animals. Further studies are needed to clarify the pathogenesis of C. andersoni in animals including humans.
Footnotes
Acknowledgements
We thank Dr. Oya at Gifu University, Faculty of Technology, for guidance provided in SEM techniques.