
Abstract
Thymic epithelial cells could play an important role in lymphoid depletion during bovine viral diarrhea virus (BVDV) infection. To evaluate this hypothesis, we examined proliferation of lymphocytes, expression of cytokeratins by thymic epithelial cells, and ultrastructural features at sequential time points after experimental infection of colostrum-deprived calves with the noncytopathogenic BVDV1 strain 7443. Ten clinically healthy Friesian calves were used. Eight were inoculated with the virus, and 2 were used as uninfected controls. Calves were sedated and euthanized in batches between 3 and 14 days postinoculation. At necropsy, thymus samples were collected for structural, immunohistochemical, and ultrastructural study. Thymic lymphoid depletion was accompanied by a decrease in lymphocyte proliferation and immunohistochemical and ultrastructural changes in thymic epithelial cells. Immunohistochemical and ultrastructural results reflect a disturbance of the thymic epithelial cell network, which may explain the decrease in lymphocyte proliferation by defective thymocyte-epithelial cell interactions.
Bovine viral diarrhea virus (BVDV) 1 is an RNA virus classified in the family Flaviviridae, genus Pestivirus. 3 Acute infection with noncytopathogenic (ncp) BVDV1 strains of low virulence causes only minor signs of disease like a mild elevation of temperature and a decrease in circulating lymphocytes. 4,9,10 Thymic lymphoid depletion has been documented during acute BVDV2 infection. 5 –7 However, there is little information regarding thymic lesions during BVDV1 infection. 11 Although different mechanisms have been proposed, 1,2,7,10 the pathogenesis of this lymphoid depletion is not completely understood.
Because of this, in a work previously published in this journal, 10 development of thymic lymphoid depletion during BVDV1 infection was investigated, together with the role of thymic macrophages in the pathogenesis of lesions. This work is a continuation of that experiment. The aim of this study was to investigate the role of thymic epithelial cells (TECs) in the pathogenesis of thymic lymphoid depletion of calves during acute experimental infection with BVDV1.
Materials and Methods
Experimental design was described by Raya et al 10 in 2012. Briefly, 10 Friesian calves, colostrum deprived, 8 to 12 weeks old, and clinically healthy, were obtained immediately after birth from a herd free of tuberculosis, brucellosis, bovine leucosis virus, and bovine herpesvirus-1. Antigen and antibody enzyme-linked immunosorbent assay (ELISA) was performed to confirm that they were free of BVDV. 10 Eight animals were inoculated (day 0) by intranasal instillation with 10 ml (5 ml per nostril) containing 10 5 median tissue culture infective dose (TCID50)/ml of ncp BVDV1 7443 (courtesy of the Institute für Virologie, TIHO, Hannover, Germany). Two control calves were housed in another barn with the same conditions, without contact with infected animals. They were inoculated by intranasal instillation of tissue culture medium and used as uninfected controls. 10 Animals were sedated with xylazine (Rompun 2% solution; Bayer Healthcare, Kiel, Germany) and euthanized in batches of 2 at 3, 6, 9, and 14 days postinoculation (dpi) by overdosing with thiopental-sodium (Thiovet; Vet Limited, Leyland, Lancashire, UK). Control calves were euthanized at 14 dpi. The experimental procedure was carried out in accordance with the Code of Practice for Housing and Care of Animals Used in Scientific Procedures, approved by the European Economic Community in 1986 (86/609/EEC amended by the directive 2003/65/EC). 10
At necropsy, 1 sample from a standardized region of the thoracic thymus was collected from each animal, fixed in 10% neutral buffered formalin, embedded in paraffin, sectioned, and processed for immunohistochemistry by the avidin-biotin-peroxidase complex method as described. 10 Details of primary antibodies used in this study, including dilutions and pretreatments, are summarized in Table 1. Mouse nonimmune serum was used in place of specific primary antibodies for negative controls. 10
Details of Immunohistochemical Methodology.

aDako, Glostrup, Denmark.
Positively labeled cells were counted in a blind manner in 20 areas of 0.2 mm2 of cortex and medulla, respectively. 10 Identification of positive cells was based on their morphologic features and size. Values were given as the mean ± standard deviation. These values were evaluated for approximate normality of distribution by the Kolmogorov-Smirnov test. Differences between the means of control and inoculated animals were assessed by the Kruskal-Wallis test followed by the Mann-Whitney U nonparametric test (GraphPad Instat 3.05; GraphPad Software, San Diego, CA. P values less than .05 represented a statistically significant difference between inoculated and control animals.
For ultrastructural study, samples were processed as described 10 and examined with a Philips CM-10 transmission electron microscope (Philips, Eindhoven, The Netherlands).
Results
In the previous study, lymphoid depletion was observed in the thymic cortex from 6 dpi onward, with morphologic changes suggestive of apoptosis, confirmed by the terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) technique and by ultrastructural study. Virus infection was detected, from 6 to 14 dpi, in TECs, macrophages, and small numbers of lymphocytes. Lesions were more severe at 9 dpi, when the widest distribution and highest numbers of cells containing BVDV antigen were seen. 10
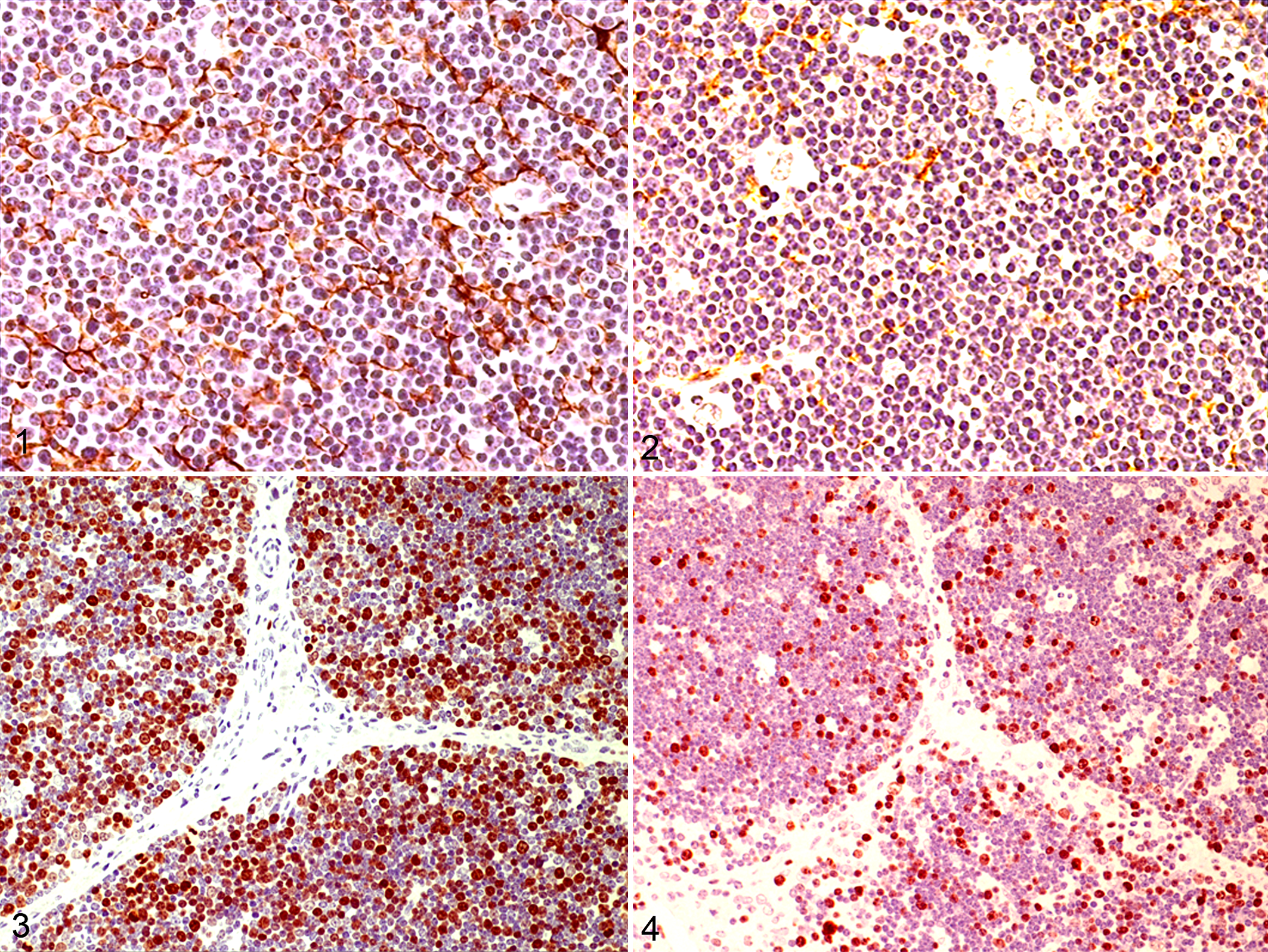
In this work, TECs were labeled with cytokeratin AE1/AE3 antibody (Figs. 1, 2). In control and inoculated animals at 3 dpi, cytokeratins showed strong expression in the cytoplasm of cells identified as TECs by their star shape, staining the stromal cell network. Strong staining was also observed in cells forming the Hassal’s corpuscles. From day 6, a significant decrease was noted in the number of TECs immunolabeled with this marker, both in the cortex and medulla, reaching the lowest number at 9 dpi. A slight increase in the number of marked cells was noted at 14 dpi (Fig. 5).
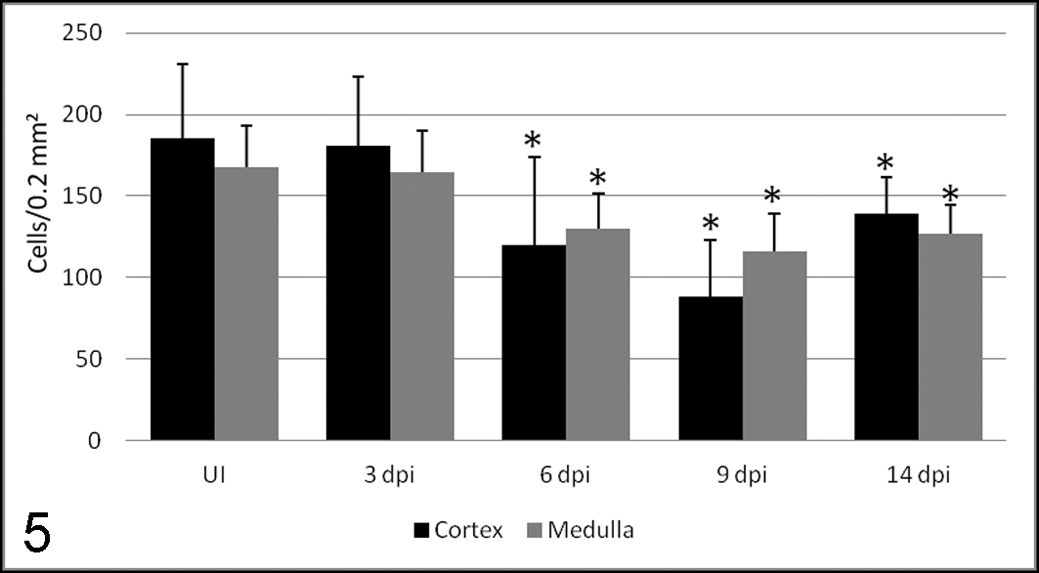
Number of cells (mean ± SD) immunolabeled with AE1/AE3 antibody in the thymic cortex and medulla of uninoculated control calves (UI) and calves inoculated with the noncytopathogenic bovine viral diarrhea virus type I strain 7443. dpi, days postinoculation. *P < .05 (between the inoculated group and controls).
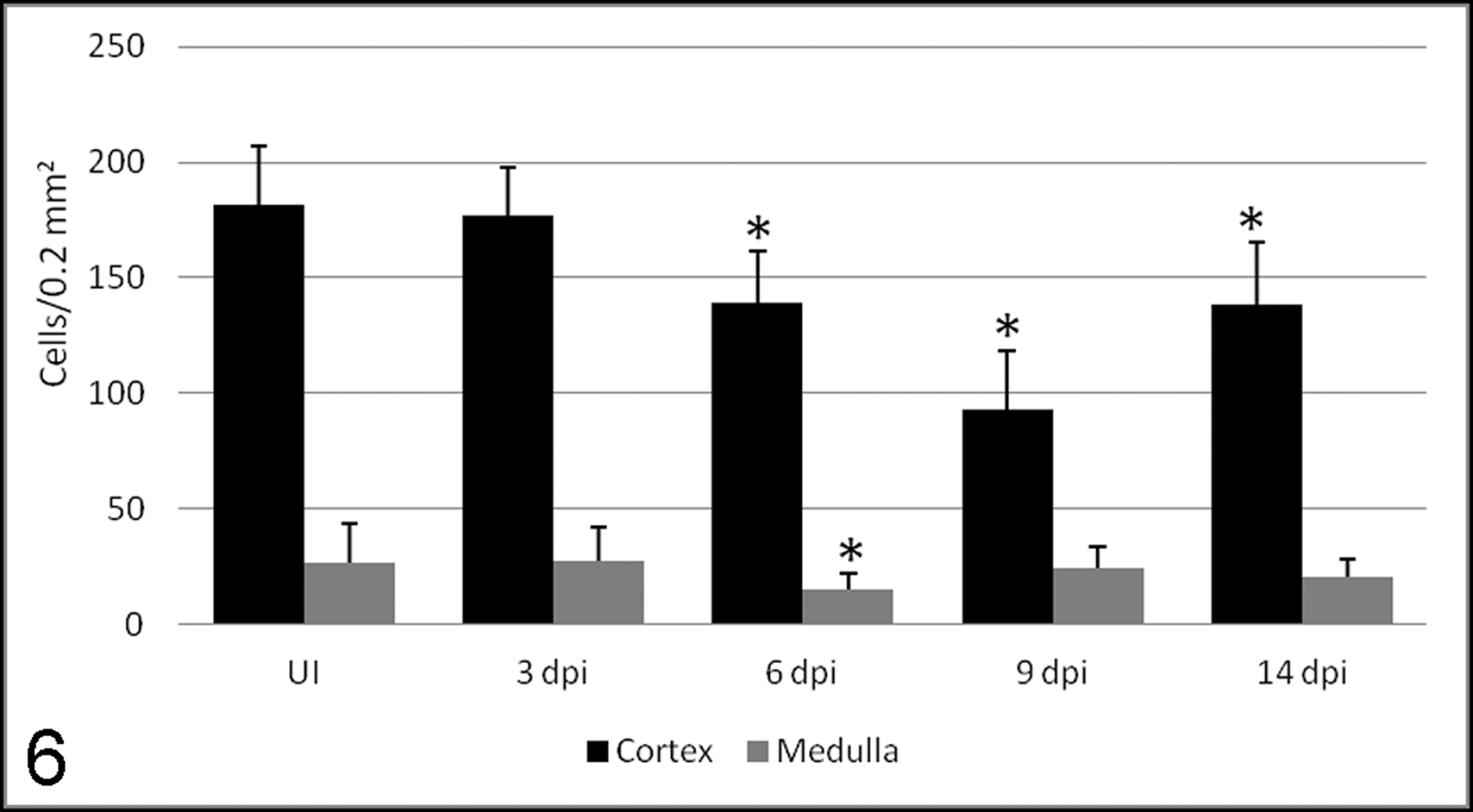
Number of cells (mean ± SD) immunolabeled with Ki-67 antibody in the thymic cortex and medulla of uninoculated control calves (UI) and calves inoculated with the noncytopathogenic bovine viral diarrhea virus type I strain 7443. dpi, days postinoculation. *P < .05 (between the inoculated group and controls).
Proliferation was detected by labeling for Ki-67 antigen both in control and inoculated animals (Figs. 3, 4). A moderate to strong nuclear staining reaction was present mainly in cells morphologically identified as lymphocytes, with a number of positive cells higher in the cortex than in the medulla. No statistically significant differences were observed among controls and animals euthanized at 3 dpi. A significant decrease in stained cells, mild at 6 and 14 dpi, was observed in the thymic cortex (Fig. 6). The decrease was severe at 9 dpi, coincident with the highest detection of viral antigen in TECs and macrophages 10 and the lowest cytokeratin staining in TECs.
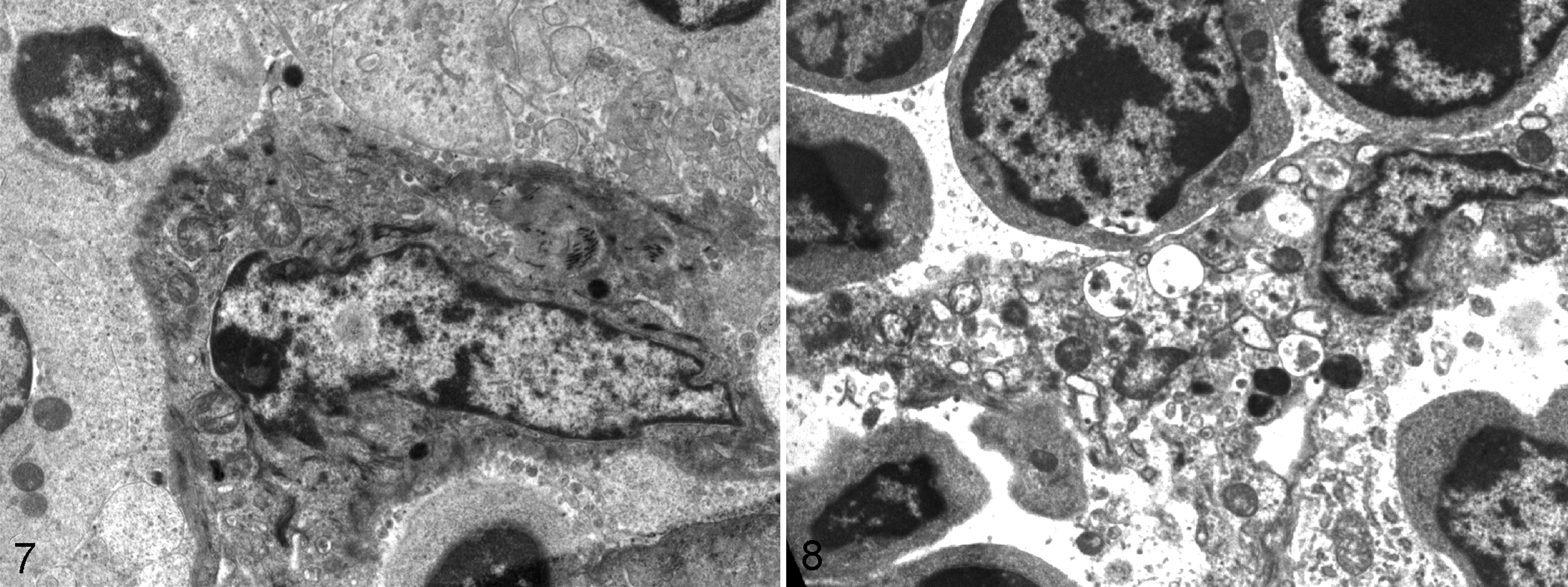
On electron microscopy, cortical epithelial cells had elongated cytoplasmic processes connected by numerous gap junctions to adjacent cells with cytoplasmic filaments (Fig. 7). From 6 dpi, degenerative changes were apparent in TECs, particularly in the cortical zone. Cisternae of rough endoplasmic reticulum were dilated and cytoplasmic vacuoles were abundant, both containing moderately electron-dense granular material and pleomorphic membranous structures (Fig. 8). An increase in primary lysosomes and autophagosomes was also observed in these cells, together with a decrease in cytoplasmic filaments. Also, a clear reduction of lymphocyte density was seen mainly affecting the cortex.
Discussion
Our results show that thymic lymphoid depletion during BVDV1 infection is accompanied by immunohistochemical and ultrastructural changes in TECs and a decrease in proliferating lymphocytes.
TECs are target cells of BVDV. 1,5 –7,10 Our study shows a decrease in the number of TECs expressing cytokeratins coincident with ultrastructural changes, such as cytoplasmic vacuolization, from 6 dpi. The network of TECs plays a crucial role in differentiation and selection processes from immature to mature T cells. 8 This damage in TECs could cause an interference in lymphocyte maturation by 2 mechanisms.
TECs stimulate lymphocyte proliferation mainly through cytokine production such as interleukin-7. 8 The presence of viral antigen in TECs and the reduction in cytokeratin staining were coincident with the decrease in lymphocyte proliferation observed at 6 dpi, which could be related to a lower stimulus of TECs. This decrease in proliferating lymphocytes has also been observed during infection with a BVDV2 strain, although the number of Ki-67–positive cells was not scored in that work. 5
Cortical TECs are responsible for the positive selection of maturing lymphocytes, mediated by major histocompatibility complex-restricted T-cell receptors, which allow T cells to receive a protective signal. So, T lymphocytes are programmed to undergo apoptosis if they are not “rescued” by cortical epithelial cells. 8 Degenerative changes observed in TECs by ultrastructural study could have produced an interference with these processes and explain the increase in apoptosis observed in the thymic cortex in the first part of this work. 10
In summary, thymic lymphoid depletion during ncp BVDV1 infection is accompanied by a disturbance in TECs, which could decrease lymphocyte proliferation and increase apoptosis because of an interference with positive selection processes. More studies regarding cytokine production by TECs are necessaries to clarify this hypothesis.
Footnotes
Acknowledgements
We thank the Institute für Virologie, TIHO (Hannover, Germany), which provided the BVDV.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Ministry of Education and Science (AGL 2006-01536). A. I. Raya was supported by a predoctoral grant from Spanish Ministry of Education and Science.