
Abstract
Expression of Mx protein and interferon-α (IFN-α) was examined by immunohistochemistry in pigs experimentally infected with swine influenza virus. In infected pigs euthanatized at 1 day postinoculation (dpi), the lumen of bronchioles were filled with large numbers of mononuclear cells, small numbers of neutrophils, sloughing epithelial cells, and proteinaceous fluid. Lesions at 3 and 5 dpi were similar but less severe. Alveolar spaces were filled with neutrophils. By 7 and 10 dpi, microscopic lesions were resolved. The immunohistochemical signals for Mx protein and IFN-α antigen were confined to cells in areas that had hybridization signal for swine influenza virus. In situ hybridization and immunohistochemistry of serial sections of lung indicated that areas containing numerous swine influenza virus RNA-positive cells also have numerous Mx and IFN-α antigen-positive cells. Mean immunohistochemical scores for Mx protein-positive cells were correlated with mean immunohistochemical scores for IFN-α antigen-positive cells (rs = 0.8799, p < 0.05). These results indicated that Mx protein and IFN-α antigen were expressed in the lung from pigs experimentally infected with swine influenza virus, but their biological functions remain to be examined.
The alpha/beta interferons (IFN-α/β) system is involved in host cell defense immediately after virus infection. Antiviral action of IFN-α is mediated by the induction of at least three major proteins, 2′,5′-oligoadenylate synthetase, protein kinase R, and Mx proteins. 29 The Mx protein, one of the IFN-inducible proteins, has been identified in a wide variety of vertebrates, including humans and mice, and is thought to play a critical role in the IFN-induced antivirus state. 6 Humans have two Mx proteins, MxA and MxB. 17 Human MxA protein accumulates in the cytoplasm, where it interferes with the multiplication of influenza virus, rhabdoviruses, and hantaviruses. 12, 25 Human MxB protein has been shown to be related to MxA protein by sequence analysis, and it has been shown to be found in both the nucleus and cytoplasm. 17, 19 A full-length cDNA clone encoding the porcine Mx1 was isolated and sequenced. 20 It contained an open reading frame of 663 amino acids that is very similar to human MxA. The porcine Mx1 was mapped on chromosome 13 with a somatic cell hybrid panel. 27
Influenza viruses are members of family Orthomyxoviridae and have segmented, negative-sense RNA genomes. Multiple subtypes within influenza A virus have been reported based on differences in hemagglutinin (H1–15) and neuraminidase (N1–9). 28 Among various subtypes of influenza A virus, H1N1, H1N2, and H3N2 have been commonly isolated from swine throughout the world. 23 Swine influenza is recognized as an acute febrile respiratory disease with high morbidity and low mortality. Clinical signs include anorexia and weight loss, dyspnea, pyrexia, coughing, and nasal discharges. 3, 9 The porcine Mx1 protein has antiviral properties against influenza virus in vitro. 7 The objectives of this study were to investigate the expression of Mx and IFN-α protein by immunohistochemistry in porcine lungs from pigs experimentally infected with swine influenza virus.
Materials and Methods
Swine influenza virus
In January 2003, a severe outbreak of influenza-like disease occurred in a pig farm with 200 breeding sows in Kyounggi Province, Korea. The typical clinical signs of influenza (sneezing, rough haircoat, and coughing) were noted in many pigs. The grower and finisher pigs were lethargic with dyspnea and high fever (up to 42°C). The A/SW/Kyounggi/0981/03 virus was isolated in Madin-Darby canine kidney (MDCK) cells from lung tissue from a pig (85 days old) that had influenza-like illness in January 2003. The subtype of A/SW/Kyounggi/0981/03 was confirmed as H1N2 on the basis of phylogenetic analysis. 10
Experimental design
Thirty colostrum-deprived, 3-week-old pigs were randomly allocated to infected or control groups. All pigs were confirmed seronegative for swine influenza virus infection by enzyme-linked immunosorbent assay (IDEXX Laboratories, Westbrook, ME) at the start of the experiment. They were maintained in a stainless-steel isolator (two pigs per isolator) and fed a commercial sterile milk substitute. The viral inoculum contained swine influenza virus at its second passage in MDCK cells. Fifteen pigs in the infected group were then inoculated intranasally with 3 ml of tissue culture fluid containing 2 × 106 tissue culture infective doses 50%/ml of swine influenza virus, while 15 pigs in the control group were exposed in the same manner to uninfected cell culture supernatants. Three pigs from each group were euthanatized at 1, 3, 5, 7, and 10 days postinoculation (dpi). Tissues were collected from each pig at necropsy. Tissues were fixed in 10% neutral buffered formalin for 1 day and routinely processed to paraffin block in an automatic tissue processor. In situ hybridization and immunohistochemistry were performed on four blocks of lung tissue, which included eight pieces (1 × 2 cm) of lung; two pieces from the right anterior lobe, two from the right middle lobe, one from the ventromedial part of the right caudal lobe, one from the dorsomedial part of the right caudal lobe, one from the midlateral part of the right caudal lobe, and one from the accessory lobe of each pig. The same eight lung tissues were also collected and frozen for reverse transcription-polymerase chain reaction (RT-PCR). All lung samples were taken from gross lesions. All the methods used were previously approved by the Seoul National University, Institutional Animal Care and Use Committee.
Semiquantitative reverse transcription-polymerase chain reaction
Semiquantitative RT-PCR was performed in the lung samples from all infected and uninfected negative control pigs. A 467-base pair (bp) cDNA fragment was used as probe for Mx (GenBank accession number M65087). The forward and reverse primers were 5′-TAAGCAGGAGACCATCAACT-3′ (nucleotide positions 724–743) and 5′-CGGAGCCATACTTCTGTAAC-3′ (nucleotide positions 1,171–1,190), respectively. A 420-bp cDNA fragment was used as a probe for IFN-α (GenBank accession number M28623). The forward and reverse primers were 5′-AGAATCTCTCCCTTCTCCTG-3′ (nucleotide positions 249–268) and 5′-GAGTCTGTCTTGCAGGTTTC-3′ (nucleotide positions 649–668), respectively. RT-PCR of IFN-α and Mx was performed as previously described. 2
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was also amplified as a quantitative control as previously described. 13 The forward and reverse primers for porcine GAPDH (GenBank accession number AF017079) were 5′-TGACCTCCACTACATGGTCT-3′ (nucleotides 449–468) and 5′-CATTTTGTGTCCTCTGACTT-3′ (nucleotides 1,173–1,192), respectively. The primer set resulted in amplified fragments of 744 bp.
For the semiquantitative analysis of RT-PCR, RT-PCR products were run on 2% agarose gel. The gels were stained with ethidium bromide, destained with distilled water, and then photographed with the EDAS 290 gel imaging system (Eastman Kodak Company, Rochester, NY), as previously described. 26 Net intensities of individual bands (same area) were measured with Kodak Digital Science 1D software (Eastman Kodak Company). Ratios of net intensity of target gene to that of the internal control bands (GAPDH) were calculated before statistical analysis. To minimize the between-assay error, samples from two groups were always processed in parallel. 22
In situ hybridization
A 411-bp segment of H1 HA genes was used as H1 probe. The forward and reverse primers were 5′-GGGACATGTTACCCAGGAGAT-3′ (nucleotides 345 to 365) and 5′-CTGCTTGACCTCTCACTTTGG-3′ (nucleotides 756–736), respectively. RT-PCR was carried out as previously described. 1 RT-PCR products of H1 HA were purified with a 30-kD cutoff membrane filter. The nucleotide sequences of the purified RT-PCR products were determined by means of BigDye chemistry with the ABI Prism Sequencer (Applied Biosystems, Foster City, CA). Sequencing was performed on the purified RT-PCR products before they were labeled by random priming with digoxigenin-dUTP (Boehringer Mannheim, Indianapolis, IN) according to the manufacturer's instructions. In situ hybridization was performed as previously described. 11 Briefly, after overnight hybridization at 45°C, sections were incubated for 90 minutes with antidigoxigenin conjugated to alkaline phosphatase (Boehringer Mannheim) diluted 1 : 200 in 0.1 M Tris-HCl (pH 7.4) and 0.15 M NaCl with 1% blocking reagent (Boehring Mannheim). Sections were colorized with nitroblue tetrazolium and 5-bromocresyl-3-indolylphosphate and then counterstained with 0.5% methyl green.
Immunohistochemistry
Two antibodies were used: 1) monoclonal mouse anti-human MxA protein antibody, 4 which labels porcine Mx1 protein (Dr. Haller, personal communication) and was kindly provided by Dr. Otto Haller from the University of Freiburg; and 2) monoclonal mouse anti-pig IFN-α antibody (Pierce Biotechnology Inc., Meridian, IL). 18 Monoclonal mouse anti-human Mx protein antibody was diluted 1 in 100 and monoclonal mouse anti-pig IFN-α antibody was diluted 1 in 50 in Tween 20 (phosphate-buffered saline [PBS] containing Tween 20, 0.1%).
Endogenous alkaline phosphatase was quenched with glacial acetic acid 20% for 2 minutes at 4°C. Slides were then subjected to optimal pressure cooking for 10 minutes for each antibody as previously described. 21 All slides were incubated with normal goat serum (Sigma Chemical, St. Louis, MO) in PBS for 30 minutes at room temperature to saturate nonspecific protein-binding sites. Sections were coated with monoclonal antibodies and incubated with antibody overnight at 4°C in a humid chamber.
After three washes with Tween 20, sections were flooded and incubated for 1 hour at 36°C with biotinylated goat anti-mouse IgG (Dako, Glostrup, Denmark) diluted 1 in 200 in Tween 20. The slides were then washed with Tween 20 before being flooded and incubated for 1 hour at 36°C with streptavidin-alkaline phosphatase conjugate (Roche Molecular Biochemicals, Mannheim, Germany). They were then equilibrated with Tris-buffer (pH 8.2) for 5 minutes at room temperature. The final reaction was produced by immersing the sections in a solution of red substrate (Boehringer Mannheim) for 10 minutes at room temperature. The sections were lightly counterstained with Mayer's hematoxylin.
Statistical analysis
Immunohistochemical signals of Mx protein and IFN-α antigen were counted in 10 random high-power fields (400×) and given a ranked score of 0–4 by subjective system: 0 = no immunohistochemical signal, 1 = very few cells stained (<1%), 2 = a few cells stained (1–10%), 3 = more cells stained (10–30%), and 4 = diffuse staining (>30%). The Mann–Whitney U-test was used to compare the infected and uninfected group at each time point. The Wilcoxon matched rank sum test was used to compare immunohistochemistry scores of Mx protein and IFN-α antigen at each time point to the previous time point. Mean immunohistochemical scores for Mx protein and IFN-α antigen were subjected to Pearson correlation analysis. P < 0.05 was considered to be statistically significant.
Results
Clinical signs
All pigs in the infected group showed respiratory signs. Anorexia was observed between 5 and 7 dpi and coughing and sneezing between 3 and 5 dpi. The control pigs remained clinically normal during the experiment.
Gross lesions
Gross lesions were observed at 1, 3, and 5 dpi, but occasionally at 7 and 10 dpi. The lesions were predominantly in the cranial, middle, and accessory lobes and the ventromedial portion of the caudal lobes. The lesions were characterized by multifocal, tan-mottled areas, with irregular and indistinct borders. No gross lesions were observed in uninfected negative control pigs.
Histopathology
In infected pigs euthanatized at 1 dpi, the lumens of bronchioles were filled with large numbers of mononuclear cells, small numbers of neutrophils, sloughing epithelial cells, and proteinaceous fluid. Interlobular septa were congested and thickened by proteinaceous fluid. Lesions at 3 and 5 dpi were similar but less severe. Alveolar spaces were filled with neutrophils. By 7 and 10 dpi, microscopic lesions were resolved. Interlobular septa were slightly distended by proteinaceous fluid and small numbers of neutrophils and mononuclear cells. Lesions were not seen in uninfected negative control pigs.
Semiquantitative RT-PCR
Amplification of template cDNA with primers of Mx protein and IFN-α resulted in amplified products corresponding to those of the predicted size, namely, 467 (Mx) and 420 (IFN-α) bp. RT-PCR products were sequenced, and their identity was confirmed as Mx and IFN-α (data not shown). Increased mRNA for both Mx protein and IFN-α were detected in lungs of the infected pigs at 1 and 3 dpi, but not in the uninfected negative control pigs (Fig. 1). The GAPDH gene was used as control gene because it is not involved in regulation of cytokine expression. The expression of lung-tissue GAPDH was similar for both infected and uninfected negative control pigs, indicating the observed cytokine mRNA level changes were likely related to the expression of the cytokines.
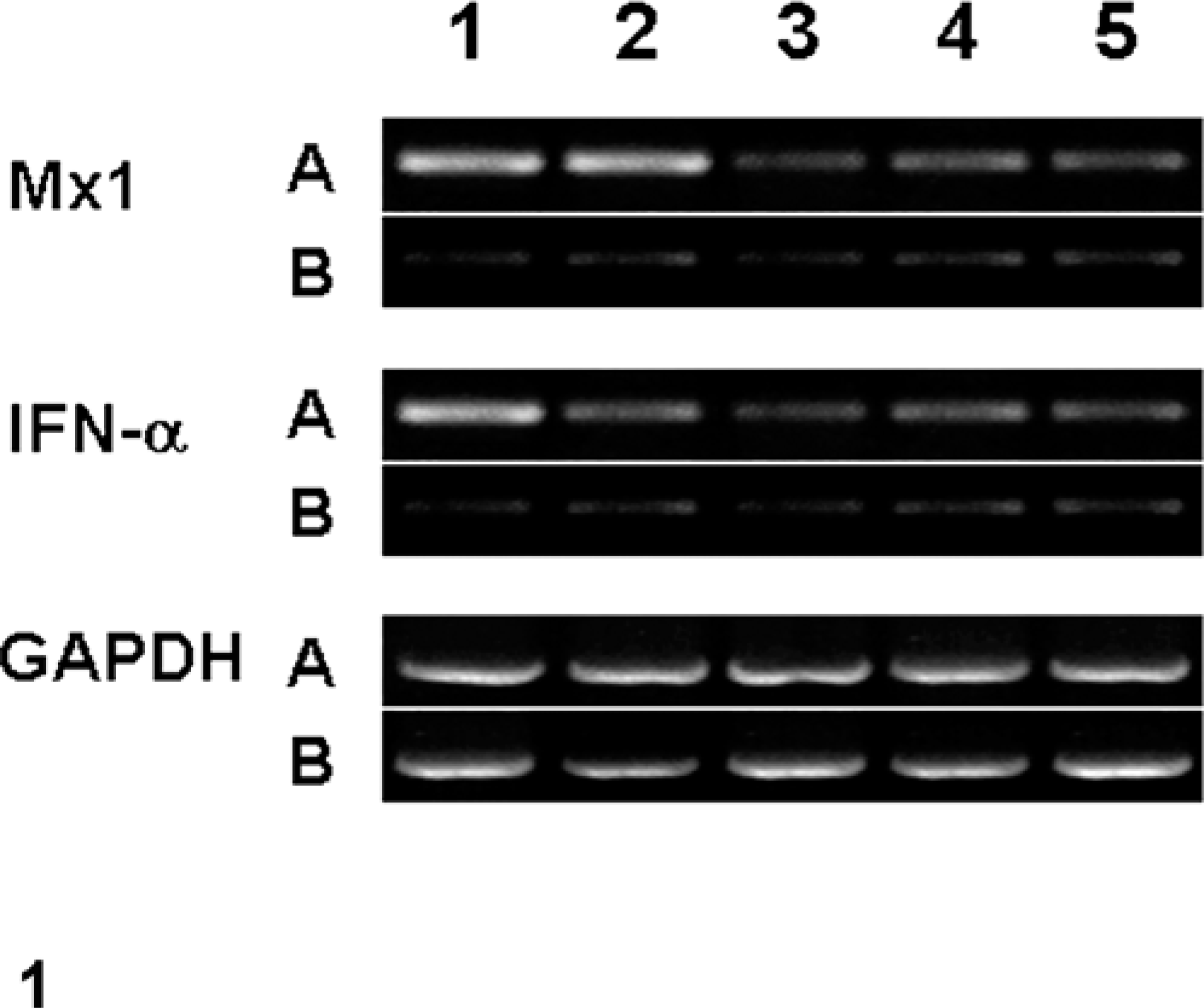
Agarose gel electrophoresis of semiquantitative RT-PCR of Mx, IFN-α, and GAPDH. From left to right: lanes 1–5 = Mx, IFN-α, and GAPDH in lung from pigs (A) experimentally inoculated with swine influenza virus at 1, 3, 5, 7, and 10 dpi and uninfected negative control pigs (B) that were similarly inoculated with sterile culture medium and euthanatized at corresponding dpi.
In situ hybridization
The 15 pigs experimentally infected with swine influenza virus had distinct positive hybridization signals for swine influenza virus RNA. Positive cells typically exhibited a dark-brown reaction product in the nucleus and cytoplasm without any background staining. Bronchial and bronchiolar epithelial cells had positive signals. Hybridization signals were also detected in inflammatory cells in the lumen of bronchioles (Fig. 2a). Pretreatment with RNase A eliminated hybridization signals for swine influenza virus from 15 experimentally infected pigs. Sections from uninfected negative control pigs showed no hybridization signal for swine influenza virus.
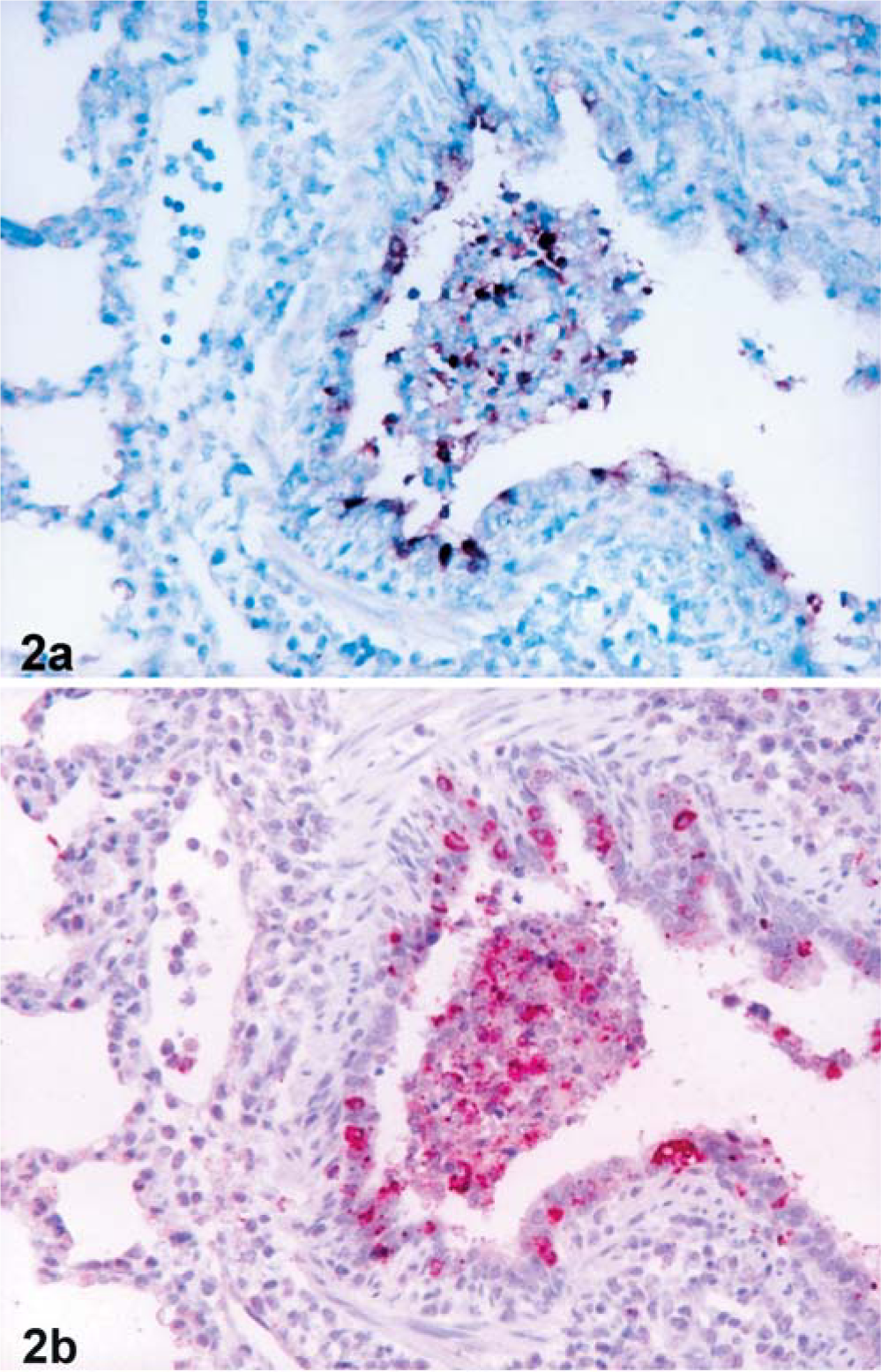
Lung tissues from pig experimentally infected with swine influenza virus at 1 dpi.
Immunohistochemistry
The 15 pigs experimentally infected with swine influenza virus had distinct positive immunohistochemical signals for Mx protein and IFN-α antigen. Positive cells typically exhibited a red reaction product in the cytoplasm without any background staining. Mx protein antigen was detected in the bronchial and bronchiolar epithelial cells, and macrophages in the lumen of bronchioles (Figs. 2b, 3). IFN-α antigen was also detected in macrophages in the lumen of bronchioles (Fig. 3) and in the bronchial and bronchiolar epithelial cells. Comparisons of immunohistochemistry with hematoxylin-and-eosin-stained sections from the same block indicated that many of the positive cells were macrophages, which had large oval nuclei and abundant cytoplasm.
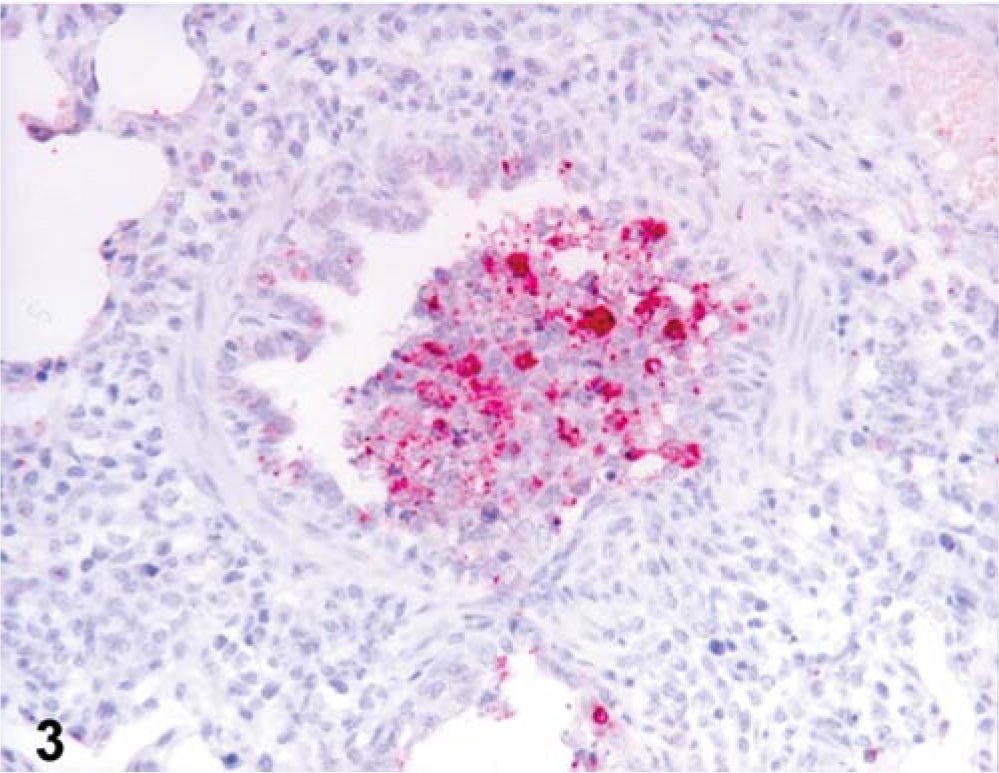
Lung tissues from pig experimentally infected with swine influenza virus at 1 dpi. IFN-α protein was detected in inflammatory cells in the lumen of bronchioles. Immunohistochemistry; alkaline phosphatase, red substrate, hematoxylin counterstain.
The immunohistochemical signals for Mx protein and IFN-α antigen were confined to cells in areas that had hybridization signal for swine influenza virus. In situ hybridization and immunohistochemistry of serial sections of lung indicated that areas containing numerous swine influenza virus RNA-positive cells also had numerous Mx protein (Fig. 2a, b) and IFN-α antigen-positive cells. In addition, a very close cell-to-cell correlation between Mx and IFN-α protein antigen in serial sections from lung was confirmed by immunohistochemistry. In the serial sections, cells positive for Mx protein were more numerous than cells positive for IFN-α antigen. Sections from uninfected control pigs showed no immunohistochemical signal for Mx protein and IFN-α antigen.
The score of Mx protein-positive cells were sustained between 1 and 7 dpi but declined markedly by 10 dpi (P < 0.05). The score of IFN-α antigen was significantly different at 3 dpi (P < 0.05) from that at 1 dpi, and the score of positive cells were decreased by 7 dpi (Fig. 4). The score of immunohistochemistry in Mx protein was correlated with the score of immunohistochemistry in IFN-α antigen (rs = 0.8799, P < 0.05).
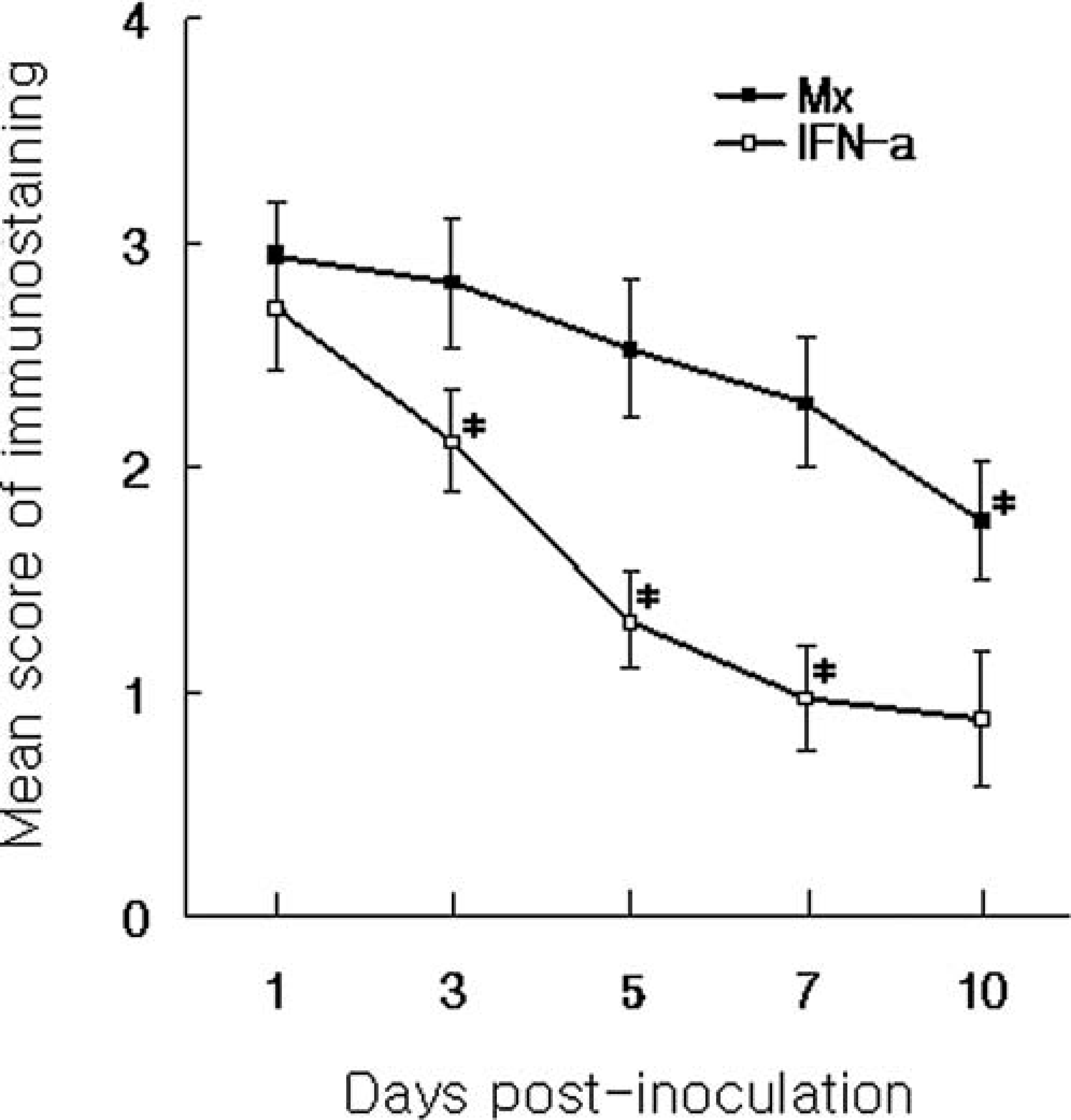
Mean score of immunohistochemistry of Mx and IFN-α in the lungs from pigs experimentally infected with swine influenza virus. ∗Significantly different at each time point compared with previous time (P < 0.05).
Discussion
Expression of IFN-α and Mx protein was markedly upregulated at the mRNA and protein levels in lung tissue from pigs experimentally infected with swine influenza virus. Simultaneous detection of mRNA and protein of Mx and IFN-α in the bronchial and bronchiolar epithelial cells and macrophages also indicates molecular evidence that these cells produce Mx and IFN-α protein. However, because mRNA was detected in lung homogenates and not via in situ hybridization or in situ RT-PCR, the identity of the cells producing these proteins has not been definitively shown in this study. Because the expression of Mx and IFN-α protein was confined to cells in areas that had hybridization signal for swine influenza virus, it would appear that induction of IFN-α and Mx protein is a direct response to swine influenza virus. Swine influenza virus infection of bronchiolar epithelial cells results in recruitment of macrophages; these macrophages within bronchiolar lumens appear to express IFN-α. Expression of IFN-α by macrophages appears to induce Mx protein expression in these cells and the adjacent bronchial epithelial cells.
Pigs are a useful experimental model for the study of human influenza virus. First, the clinical signs of influenza in pigs are similar to those in humans. 5 Second, pigs are also common hosts for influenza viruses because their tracheal epithelium contains receptors for both avian and mammalian influenza virus, 8 and therefore pigs can be infected with human influenza virus. And finally, as shown in the present study, Mx proteins are also induced in pigs experimentally infected with swine influenza virus, as in humans. It has been also reported that porcine Mx proteins have antiviral properties against influenza virus in vitro. 7
IFNs are essential for the survival of higher vertebrates because they provide an early line of defense against viral infection, hours to days before immune responses. This vital role has been demonstrated by the exquisite sensitivity to virus infections of mice lacking both IFN-α/β and γ receptors. 30 The maximal expression of IFN-α and Mx protein was observed at 1 dpi in the present study. Antiviral activity of IFN-α is mediated by induction of distinct intracellular proteins in the target cells, such as Mx proteins. Selective antiviral activities of IFN-α can be correlated with the synthesis of Mx protein in the case of influenza virus. 29 The present study confirms a close associated between Mx and IFN-α protein expression, reflected by close statistical correlations. In addition, cells positive for Mx protein were more numerous than cells positive for IFN-α in the same areas, indicating that, once produced by the cells, IFN-α binds to specific cellular receptors either on the producing cells themselves or on neighboring cells and then induces the expression of Mx protein.
The antiviral properties of Mx proteins are influenced by the intracellular localization of the individual proteins as well as by the viral pathogens. Human MxA proteins accumulate in the cytoplasm but not in the nucleus, 17 which is the site of influenza virus transcription and replication. 16 Therefore, MxA protein does not inhibit primary transcription of influenza virus RNA but does inhibit synthesis of viral proteins. 24 Although the molecular mechanisms of the antiviral action of MxA proteins against influenza virus are not understood clearly at present, human cytoplasmic MxA protein may inhibit replication of influenza virus at the posttranscriptional level. However, in a study of Thogoto virus, which is another orthomyxovirus, human MxA proteins interacted with the nucleocapsid proteins of virus that prevent the infecting particles from entering the cell nucleus. 14, 15 Therefore, the cytoplasmic protein Mx protein has a broader spectrum of antiviral activity.
In conclusion, these results indicated that Mx protein and IFN-α antigen were expressed in the lung from pigs experimentally infected with swine influenza virus, but their biological functions remain to be examined. Further studies are needed to determine the antiviral activities and mechanisms of porcine Mx protein against swine influenza virus in pigs.
Footnotes
Acknowledgements
The research reported here was supported by contract research funds of the Research Institute for Veterinary Science (RIVS) from the College of Veterinary Medicine, Seoul National University, and Brain Korea 21 Project, Republic of Korea.