
Abstract
A 12-year-old male harbor seal presented with progressive signs of neurologic dysfunction including head tremors, muzzle twitching, clonic spasms, and weakness. Lesions included polioenceph-alomyelitis with glial nodules, spheroids, neuronophagia, ring hemorrhages, and a few neutrophils. Neurons, fibers, and glial nodules were multifocally colonized with intracytoplasmic West Nile flavivirus antigens that were demonstrated using indirect immunohistochemical analysis. Flavivirus on cultured cells also was isolated and was identified by use of monoclonal antibodies and reverse transcriptase-polymerase chain reaction analysis. Clinical signs of disease and lesion morphology and distribution were similar to those of equine West Nile virus infection. Similar to horses, alpacas, humans, dogs, and reptiles, seals can be dead-end hosts of West Nile virus.
West Nile virus (WNV) is a single-stranded, RNA positive-strand virus, member of the Japanese encephalitis antigenic group, genus Flavivirus, family Flaviviridae. 2 All known members of this complex (Alfuy, Japanese encephalitis, Kokobera, Koutango, Kunjin, Murray Valley encephalitis, St. Louis encephalitis, Stratford, Usutu, and West Nile viruses) are transmittable by mosquitoes, and many of them can cause febrile, sometimes fatal illnesses in animals and humans. West Nile flavivirus is composed of a virion envelope from the host cell membrane, where integral E glycoprotein is localized, and an internal icosahedral nucleocapsid. The flavivirus genome is 11,000 to 12,000 nucleotides long. The 5′- and 3′- ends contain noncoding regions. The genome encodes 10 proteins; three of which are structural proteins (C, M, and E), and seven of which are nonstructural proteins (NS1, NS2a, NS2b, NS3, NS4a, NS4b, and NS5). The M protein is synthesized as a precursor (prM) protein. The prM protein is processed to pr + M protein late in virus maturation by the furin convertase enzyme. 7 West Nile virus was introduced into North America in 1999, when health officials in Queens, New York reported an outbreak of encephalitis accompanied by severe weakness and axonal neuropathy. The disease began an insidious spread across the continental United States. West Nile virus infection has now been recognized in almost all American states and in Canada; more than 250 species of animals have been affected. 14 We describe a fatal case of polioencephalomyelitis due to WNV infection in a harbor seal (Phoca vitulina). To our knowledge, this is the first report of WNV infection in marine mammals.
A 12-year-old male harbor seal at the New Jersey Aquarium experienced 9 days of anorexia, intermitted diarrhea and vomiting, grunting noises, and labored breathing. Progressive head tremors and muscular stiffness, involuntary spasms of the head and shoulder areas, swallowing difficulties, hind flipper weakness, involuntary blinking and mouth movement, and muscular twitching were followed by clonic spasm. The seal was anemic, leukopenic, and mildly hyponatremic, and its liver enzyme activities were increased. Total white blood cell count was mildly decreased (4.7 × 103/µl; normal, 7.0–19.4 × 103/µl); however, the differential cell distribution was within normal limits. Serum creatinine concentration was at the high end of normal (1.4 mg/µl). Serum alkaline phosphatase activity was decreased (151 IU/L; normal, 30–303 IU/L), and serum alanine transaminase activity was increased (338 IU/L; normal, 70–240 IU/L). Eventually, the disease led to natural death. Results of serologic testing for feline coronaviruses, sea lion calicivirus, morbillivirus, canine parvovirus, Leptospira interrogans serogroups pomona and hardjo, and tetanus toxin were negative. Aerobic blood culturing yielded no growth. Results of serologic testing for Borrelia sp. were positive (titer 1 : 320), using antidog fluorescein isothiocyanate-conjugated (FITC) antigen and were negative using antipig FITC antigen. Serum neutralization (SN) IgM titer for WNV was 1 : 40. As the facility was part of the WNV zoo and aquariums surveillance program, a WNV SN test had been performed four months before the onset of clinical signs of disease and results were negative.
Gross lesions included hyperemia of brainstem and spinal cord blood vessels and decreased blubber thickness. Fresh and buffered 10% formalin-fixed specimens were collected. Fresh tissues were submitted for rabies rhabdovirus detection by use of direct fluorescent antibody evaluation and WNV nested reverse transcriptase-polymerase chain reaction (RT-PCR) analysis. Fixed tissues were processed for histologic examination and indirect peroxidase immunohistochemical analysis for infective agents.
Histologic examination revealed moderate-to-severe nonsuppurative encephalomyelitis, mainly affecting the gray matter of the brainstem and spinal cord. Lesions in the spinal cord were characterized by severe, bilaterally symmetric, perivascular lymphocytic and lesser plasmacytic infiltrates with rod cells, a few neutrophils in the lateral and ventral horns, and occasional involvement of the dorsal horns (Fig. 1). Prominent gliosis, swollen axons, and ring hemorrhages also were present. Moderate inflammation, spheroids, and multifocal glial nodules were observed in the medulla oblongata, cerebellar medulla, pons, mesencephalon, thalamus, and basal nuclei, where chromatolysis and occasional neuronophagia also were identified (Fig. 2). Inflammatory changes were milder in the cerebral and cerebellar cortices, and mainly were characterized by scattered small glial nodules, proliferation of microglial cells, and a few perivascular infiltrates. Scattered microhemorrhage was observed in the cerebellar cortex. Mild lymphocytic infiltration involved the leptomeninges of the spinal cord and cerebellum. Occasional hyalinosis and mineralization of leptomeningeal vessel walls and moderate fibrosis of the choroid plexus were observed and interpreted as age-related changes. Relevant lesions were not observed in other organs.
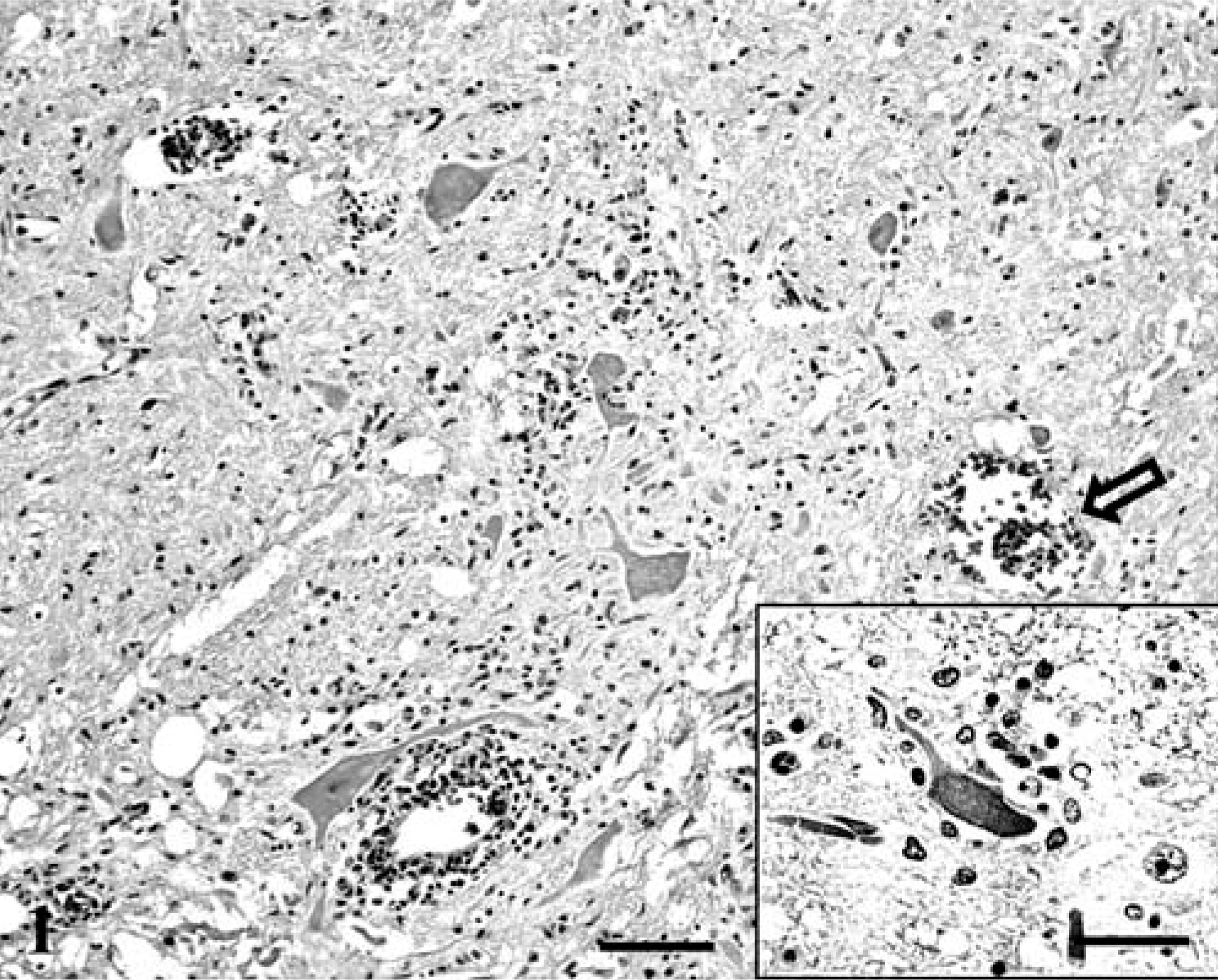
Spinal cord, ventral horn; seal with West Nile virus (WNV) infection. Poliomyelitis with perivascular moderate lymphocytic infiltrate, gliosis, and ring hemorrhage (open arrow). Bar = 300 µm.
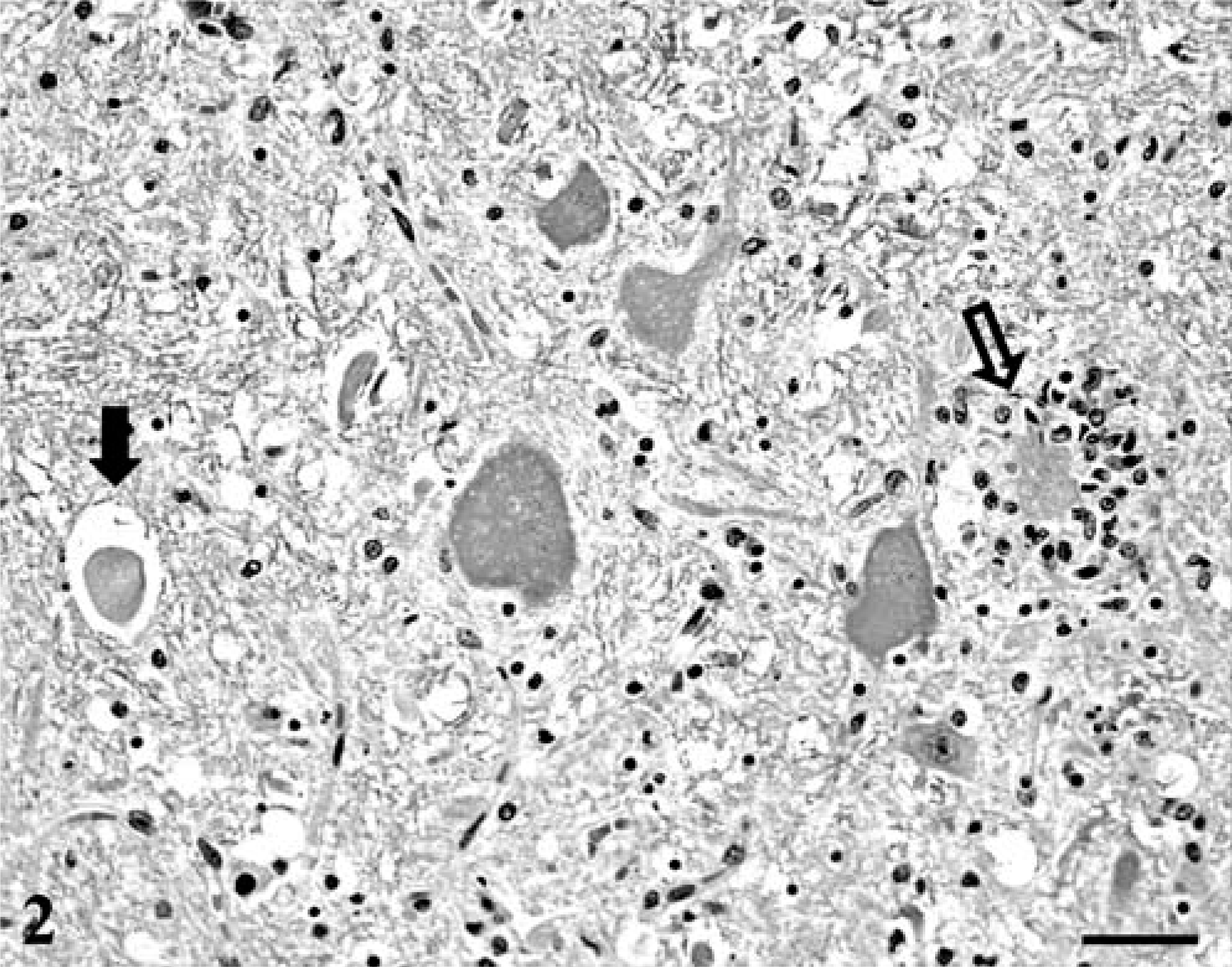
Pons; seal with WNV infection. Chromatolytic neurons with spheroid (solid arrow) and neuronophagia (open arrow). HE. Bar = 90 µm.
West Nile flavivirus was detected by use of indirect peroxidase immunohistochemical analysis 3 with a polyclonal antibody recognizing Japanese encephalitis flavivirus group epitopes (Bioreliance, Rockville, MD). The viral antigen was contained within the cytoplasm of neurons, fibers, and glial cells forming nodules (Fig. 3). In particular, the virus was localized within viable and necrotic neurons of the ventral horns. Small amounts of WNV also were identified in a few macrophages adjacent to the cerebellar cortex. Flavivirus antigen was not detected in extraneural tissues. The 248-bp WNV was identified from the brainstem of this seal by use of nested RT-PCR analysis 7 at the Pennsylvania Department of Health and the National Veterinary Services Laboratories. Brain tissue was negative for rabies rhabdovirus using direct fluorescent antibodies and for morbillivirus antigen using indirect peroxidase immunohistochemical analysis.
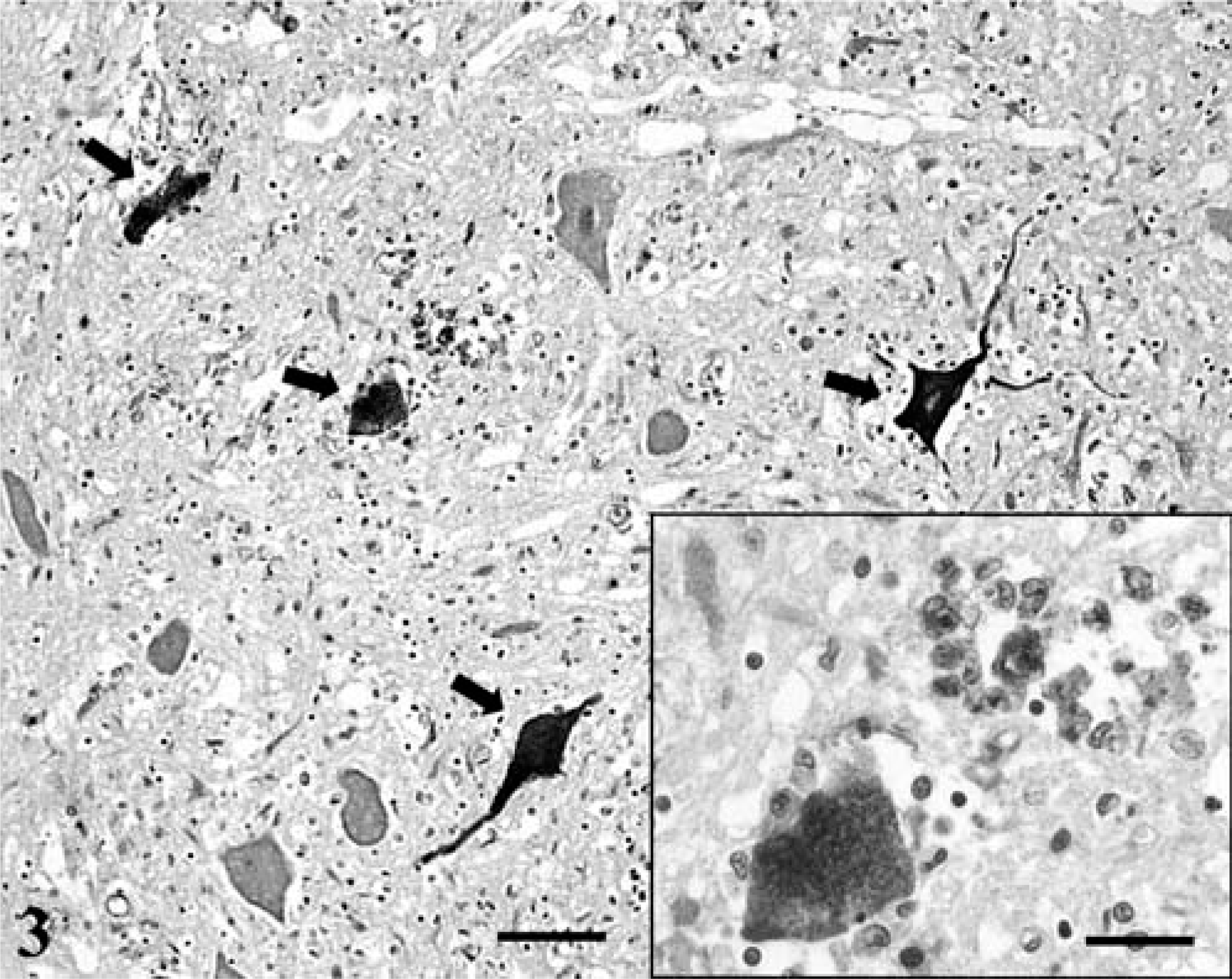
Spinal cord, ventral horn; seal with WNV infection. Neurons fibers (arrows) and glial cells contain intracytoplasmic WNV antigen. Bar = 90 µm.
A few viral, bacterial, and protozoal organisms are able to cause encephalomyelitis and/or meningoencephalitis in seals. Phocine morbillivirus is able to cause severe epizootic disease in seals with respiratory distress and signs of neurologic dysfunction. 6 Gross lesions include severe pneumonia. Bronchointerstitial pneumonia with intracytoplasmic and intranuclear acidophilic inclusion bodies is observed microscopically. In the brain, there is nonsuppurative demyelinating encephalitis with neuron necrosis, multifocal gliosis, and perivascular cuffing. Neurons and astrocytes contain intracytoplasmic and intranuclear inclusions. Morbillivirus antigen is detectable in the nucleus and cytoplasm of neurons, fibers and glial cells, lymphoid tissue dendritic cells, macrophages, and lung and urinary bladder epithelium. Influenza A 8 and B 11 orthomyxoviruses were isolated from neural and extraneural tissues of naturally infected harbor seals. 11 Toxoplasma gondii and Sarcocystis neurona were isolated from harbor seals with fatal nonsuppurative meningoencephalitis, with necrosis most severe in the cerebellar cortex. 9, 10 The most common isolates from suppurative meningoencephalitis, and sometimes ocular lesions in seals, including California sea lions (Zalophus californianus), harbor seals (Phoca vitulina), and northern elephant seals (Mirounga angustirostris), were Enterococcus spp., Escherichia coli, Klebsiella spp., Pseudomonas spp., and Salmonella spp. 13
Nonfatal WNV infection previously was serologically diagnosed in a seal at the Detroit zoo (personal communication from Dr. Ann Duncan) that exhibited signs of neurologic dysfunction of 4 days' duration, including similar tremors and twitching. Several of these signs, especially weakness, involuntary blinking, muzzle twitching, and body tremors, were similar to clinical signs of disease described in horses with WNV-induced encephalomyelitis. 4, 12 The clinicopathologic parameters were considered nonspecific and nondiagnostic for WNV and did not correlate with morphologic findings. Lesion morphology, topography, and WNV distribution also were similar to those described in horses 4 and alpacas, 5 in which polioencephalomyelitis with glial nodules, spheroids, ring hemorrhages, and mild-to-moderate immunoreactivity to WNV was restricted to neurons, fibers, glial cells, and macrophages in the central nervous system (CNS). Lesions and WNV distribution in this seal differed from the findings reported in a naturally infected dog where, in addition to polioencephalitis, myocarditis, and nephritis with tubular necrosis, WNV in renal tubular epithelium and intratubular casts were observed. 1
The severity of lesions when related to the paucity of WNV distribution and the sporadic occurrence seems to indicate that seals, like horses, may be incidental, dead-end hosts of this flavivirus. Consequently, seals, similar to horses, alpacas, humans, and dogs, most likely do not play an impoortant role in the epidemiology of WNV and may not pose a major risk to human health. However, special protection should be used during dissection and examination of the CNS of WNV-infected seals.
The WNV should be considered an important sporadic pathogen for seals, capable of inducing appreciable neurologic disease in regions where enzootic infection cycles may develop and be maintained. The paucity of WNV antigen in seal nervous tissues indicates that multiple sections of brain and spinal cord, labeled by immunohistochemical analysis, should be examined, and negative results should be interpreted with substantial caution. The brainstem appears to be the elective specimen for diagnostic evaluation. The combination of histologic examination and indirect immunohistochemical analysis, along with virus isolation and RT-PCR techniques, as previously documented in horses and alpacas, will increase the sensitivity for WNV identification, allowing further characterization of this increasingly important disease.