
Abstract
Leukodystrophies are inherited neurological disorders involving central nervous system white matter. They are uncommon in animals but a few, breed-specific entities have been described. In 2002, two young-adult, purebred Bullmastiff dogs from central New York State presented to their referring veterinarians displaying moderate to severe ataxia of all limbs, spastic tetraparesis that was worse in the pelvic limbs, and a diffuse, action-related, whole-body tremor. Clinical signs were insidious in onset and slowly progressive. Anatomic diagnoses considered were a C1–C5 lesion or, based on the whole-body tremor, a diffuse central nervous system disorder. No gross lesions were apparent in the brain or spinal cord. Histopathologically, numerous, multifocal, sharply demarcated, small, ovoid to angular areas of myelin pallor (plaques) were present throughout the major white matter tracts of the brainstem and spinal cord. These plaques, which often were traversed by axons, did not stain with luxol fast blue for myelin and were associated with minimal astrocytosis. Ultrastructural findings include occasional hypertrophic glia in white matter, rare unmyelinated segments of axons, and focal proliferation of tubule-containing cytoplasmic glial cell processes (oligodendroglial). The described clinical and morphological findings and age of onset are similar to the well-characterized, presumably hereditary, bovine syndrome known as Charolais ataxia or oligodendroglial dysplasia. This article presents the first description of a leukodystrophy in the Bullmastiff breed and the first report of oligodendroglial dysplasia in animals other than Charolais cattle.
Keywords
Leukodystrophies are a group of uncommon and heterogeneous disorders that can be broadly defined as hereditary abnormalities of central nervous system (CNS) white matter. In animals, the majority of leukodystrophies have been reported in the dog, often as novel, breed-specific entities, for example, globoid cell leukodystrophy in Cairn Terriers and West Highland White Terriers, and cavitating leukodystrophy in Dalmations. 1, 12 While hereditary leukodystrophies are also rare in cattle, a novel form occurs in the Charolais breed and is known clinically as Charolais ataxia. Ultrastructural studies have revealed abnormalities in oligodendrocytes that account for the characteristic dystrophic plaques in white matter of the brain and spinal cord of affected cattle and, for this reason, the disorder is also known as oligodendroglial dysplasia. 2, 3, 7, 8 In other species, hereditary leukodystrophies are almost unknown and acquired abnormalities of CNS white matter are much more common, such as equine leukoencephalomalacia secondary to ingestion of corn contaminated with the mycotoxin fumonisin B1 produced by Fusarium moniliforme.
The subject of this article is a condition morphologically resembling the Charolais disorder affecting two young-adult Bullmastiff dogs from north central New York State. To the authors' knowledge, this is the first report of a leukodystrophy in the Bullmastiff breed and the first report of oligodendrogial dysplasia in animals other than Charolais cattle.
Case Histories
Dog No. 1, a 2-year-old intact male Bullmastiff, was referred to Cornell University at 6 months of age for a progressive history of mild tetraparesis and ataxia in all four limbs since 5 weeks of age. At that time, an action-related head and neck tremor was identified. No ancillary studies were performed. The dog was presented to a board-certified veterinary surgeon at 2 years of age because of slowly progressive, severe neurological disease. He was alert and responsive and cranial nerves were within normal limits. The dog was unable to stand without assistance, but if supported, showed spastic tetraparesis and ataxia, more severe in the pelvic limbs. The dog's tone and spinal reflexes were normal to increased. A fine, rapid, whole-body tremor was identified that was very pronounced. Neuroanatomic localization was that of a diffuse CNS disorder.
Dog No. 2, an 8-month-old intact male Bullmastiff, presented to a board-certified veterinary surgeon at 5 months of age for a history of 2 months of progressive tetraparesis and ataxia in all four limbs. Neuroanatomic localization at that time was to cervical spinal cord segments C1–C5. Radiographs and myelogram disclosed no abnormalities. Lumbar cerebrospinal fluid (CSF) contained no cells and 40 mg/dl protein (normals: cell count <5 cells/µl, protein <25 mg/dl). The dog was discharged from the hospital with no specific diagnosis. One month later, he presented to Cornell because of worsening of the neurological signs. On examination, the dog was alert and responsive and cranial nerves were normal. The dog was unable to stand in the pelvic limbs without help; however, he could walk with assistance and exhibited a spastic paresis and ataxia in the pelvic limbs. The thoracic limbs had a stiff, long stride and a tendency to overreach during protraction. A fine, rapid tremor of the head, neck, and trunk was observed intermittently. The dog's tone and spinal reflexes were normal to increased. No vertebral column discomfort was elicited. Neuroanatomic localization was predominantly consistent with a C1–C5 lesion with possibly a more severe T3–L3 lesion. However, a more diffuse CNS disorder was suspected due to the intermittent whole-body tremor.
Both animals were humanely euthanatized and the brain from dog No. 1 and the brain and spinal cord from dog No. 2 were removed and immediately fixed in 10% neutral buffered formalin. After fixation, representative sections of all major anatomical regions of the CNS were routinely processed, sectioned at 5 microns, and stained with hematoxylin and eosin (HE) for histopathologic examination. Additional histochemical stains, including Bielschowsky silver stain and Luxol fast blue, were performed on selected sections in each case.
Immunohistochemical stains were performed on selected sections for glial fibrillary acidic protein (GFAP), neurofilament protein, and MAC387 using the streptavidin-biotin method with an Autostainer plus (Dakocytomation, Carpinteria, CA) according to the manufacturer's instructions. The primary antibodies were rabbit polyclonal for GFAP (DAKO) at a dilution of 1 : 3,000, mouse monoclonal for neurofilaments (DAKO) at a dilution of 1 : 300, and mouse monoclonal for MAC387 (DAKO) at a dilution of 1 : 300. No antigen retrieval pretreatments were performed for GFAP and neurofilaments. Antigen retrieval pretreatment for MAC387 consisted of pronase digestion for 20 minutes at room temperature. The chromagen was 3,3-diaminobenzidene-tetra hydrochloride and the slides were counterstained with hematoxylin (Dakocytomation, Carpinteria, CA).
For transmission electron microscopy, formalin-fixed samples of medulla (dog No. 1) and spinal cord (dog No. 2) were used; samples of medulla were further fixed in 2.5% gluteraldehyde in 0.1 M sodium cacodylate, pH 7.2, for 30 minutes at room temperature and 1.5 hours at 4°C. Tissues from both dogs were washed three times in 0.1 M sodium cacodylate, pH 7.2, for 10 minutes at 4°C, postfixed in 2% osmium tetroxide for 1 hour at room temperature, and then washed again three times in cacodylate buffer. The samples were successively dehydrated in graded ethanol solutions, incubated in 100% acetone and then embedded in epon araldite plastic. Thick (1-micron) sections stained with Azure B (0.5% in 0.1 M sodium phosphate, pH 7.8) were examined to identify white-matter plaques and such areas were trimmed, thin sectioned, stained with lead citrate and uranyl acetate, and examined in a Phillips 201 transmission electron microscope.
Results
Gross pathology
No abnormalities were observed on the external surface or the cut surface of the brain (both dogs) or spinal cord (dog No. 2).
Histopathology
The nature of the histologic findings was similar in both dogs, and both animals are described together. The distribution of the lesions varied between the two dogs, and that information follows. In HE-stained sections, the major white matter areas of the brainstem and spinal cord contained many small, round to ovoid, sharply defined, pale eosinophilic areas of pallor (plaques), which ranged in size from approximately 20 µm to 40 µm (Figs. 1, 2). The plaques were often surrounded by a thin peripheral ring of vacuolation and rarely by one or two compressed hyperchromatic nuclei (Figs. 2, 3). Typically, a single axon coursed through the center of most plaques, and these were easily demonstrated by Bielschowsky's silver stain (Fig. 4). The short segment of the axon traversing occasional plaques was often notably thinner than the adjacent segments. Axonal swellings—spheroids—were not observed. The plaques did not stain with luxol fast blue, contrasting with normal myelinated fibers adjacent to the plaques, which stained normally (Fig. 5). Plaques were confined to the white matter and were absent in gray matter areas, such as the cerebral or cerebellar cortices, basal nuclei, or the dorsal and ventral columns of the spinal cord. Multifocally, oligodendroglial cells were mildly hypertrophied, characterized by a plump nucleus with lacy chromatin and a mildly increased volume of pale basophilic cytoplasm with angular cell borders (Fig. 5). Such cells were particularly apparent in the short interfascicular chains of cells in longitudinal sections of the spinal cord. Occasional mildly hypertrophied astrocytes with eccentrically located, open-faced nuclei, small to moderate amounts of cytoplasm, and arborizing cellular processes were adjacent to occasional plaques (Fig. 6). In many affected areas, there was a slight hypercellularity, suggestive of minimal gliosis. Resting macrophages and foamy gitter cells were conspicuously lacking.
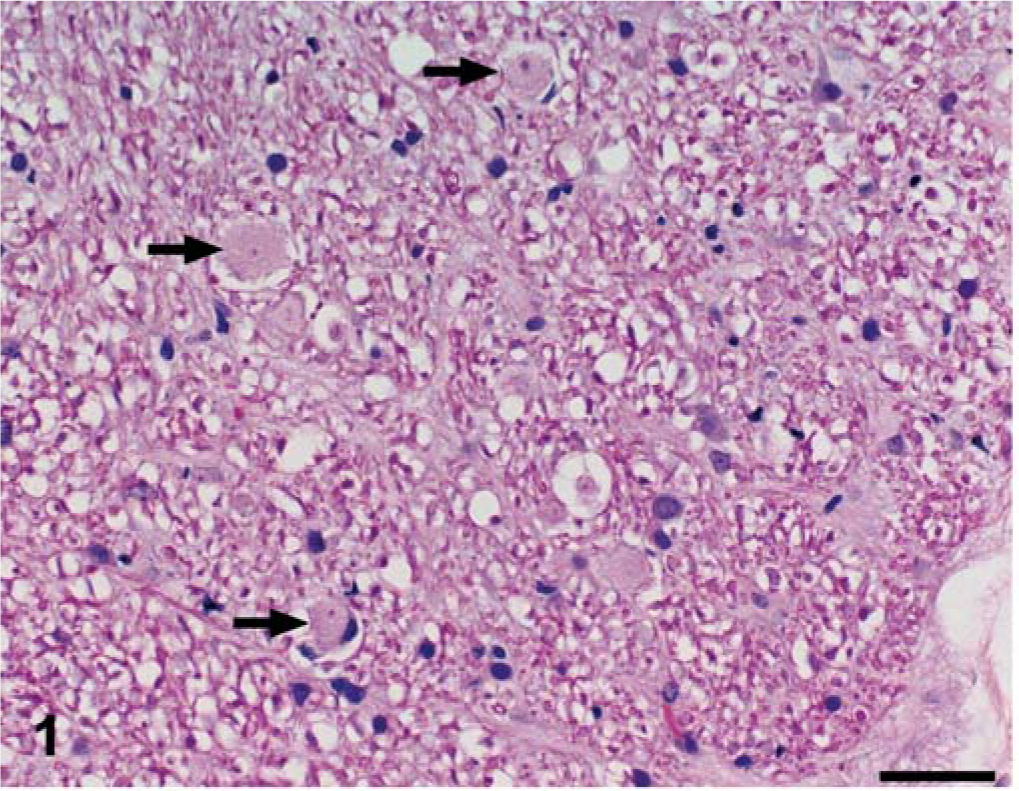
Brain; dog No. 1. The white matter of the brainstem contains a few small, round, pale eosinophilic areas of pallor (plaques); three are indicated by arrows. HE. Bar = 45 µm.
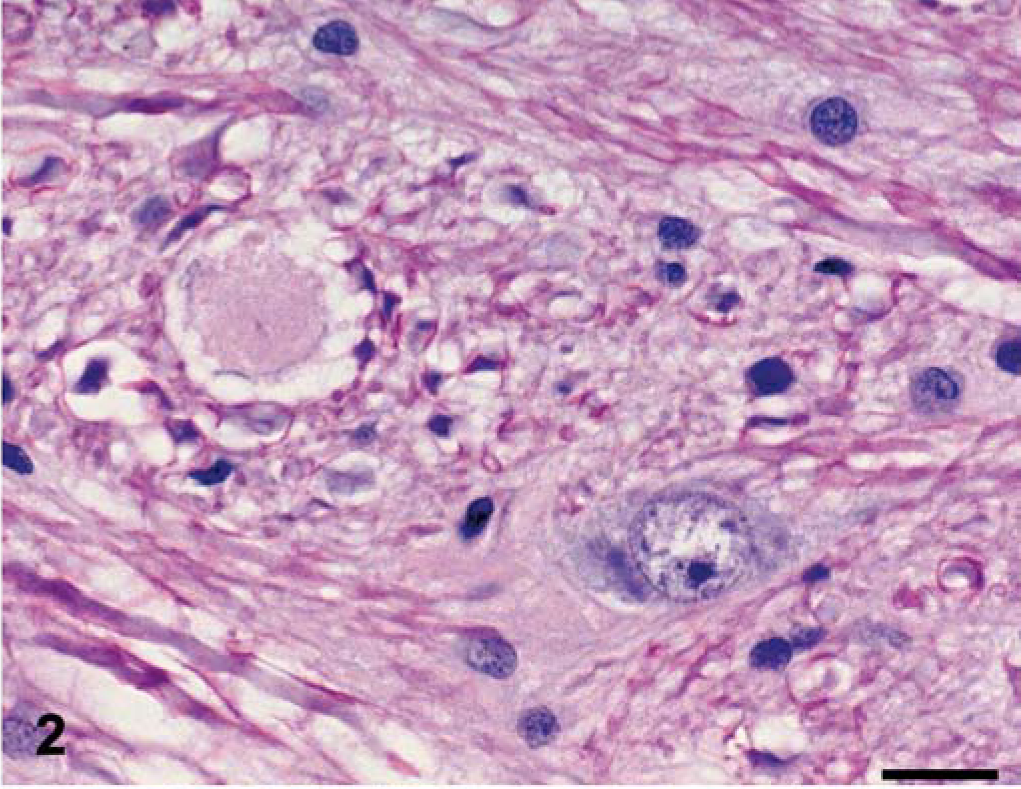
Brain; dog No. 1. The plaques were often surrounded by a thin ring of peripheral vacuolation and occasionally 1–2 compressed, hyperchromatic nuclei. HE. Bar = 20 µm.
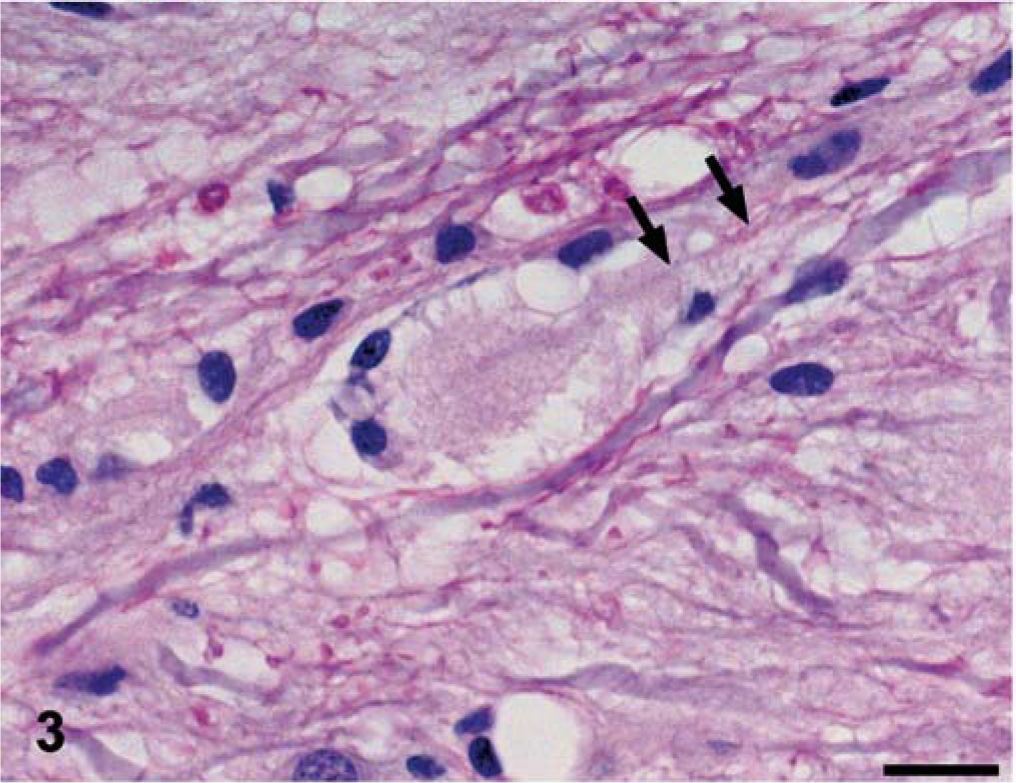
Spinal cord; dog No. 2. In longitudinal section, the plaques are ovoid and are traversed by a central axon (arrows). Note the displacement of the adjacent axons. HE. Bar = 20 µm.←
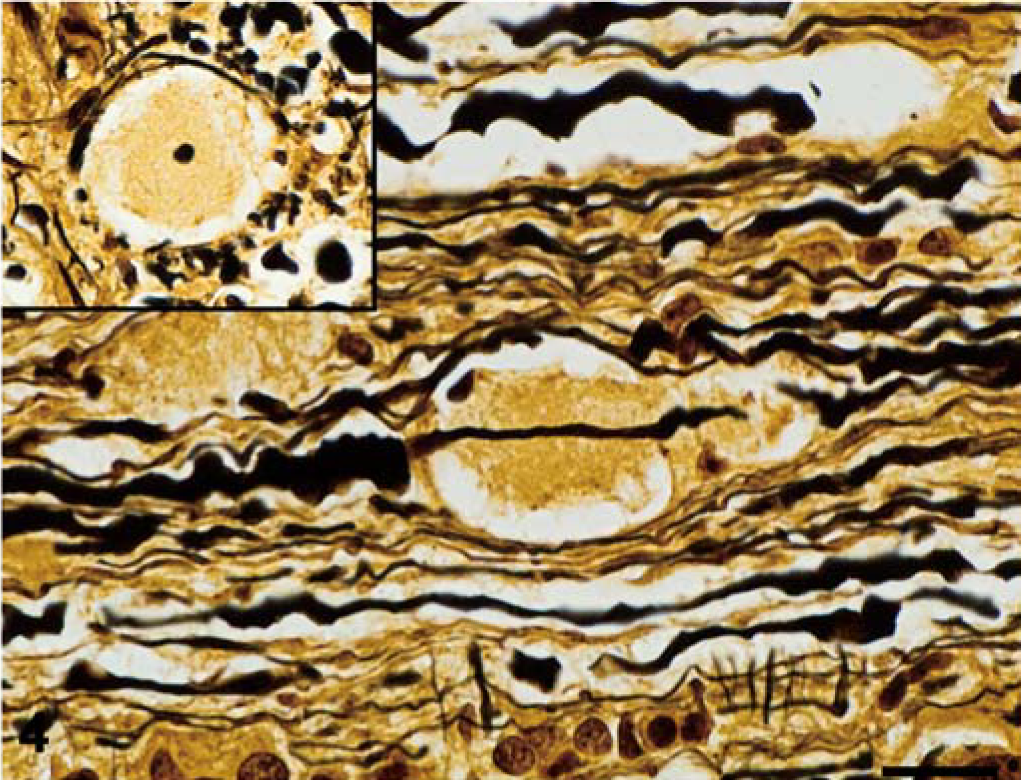
Spinal cord; dog No. 2. Typically, a single axon traverses the plaques, which is highlighted with a Bielschowsky silver stain. Often, the axon is notably thinner as it traverses the plaques than in the adjacent segments. The inset shows a plaque at the same magnification with a central, transected axon. Bielschowsky silver stain. Bar = 20 µm.
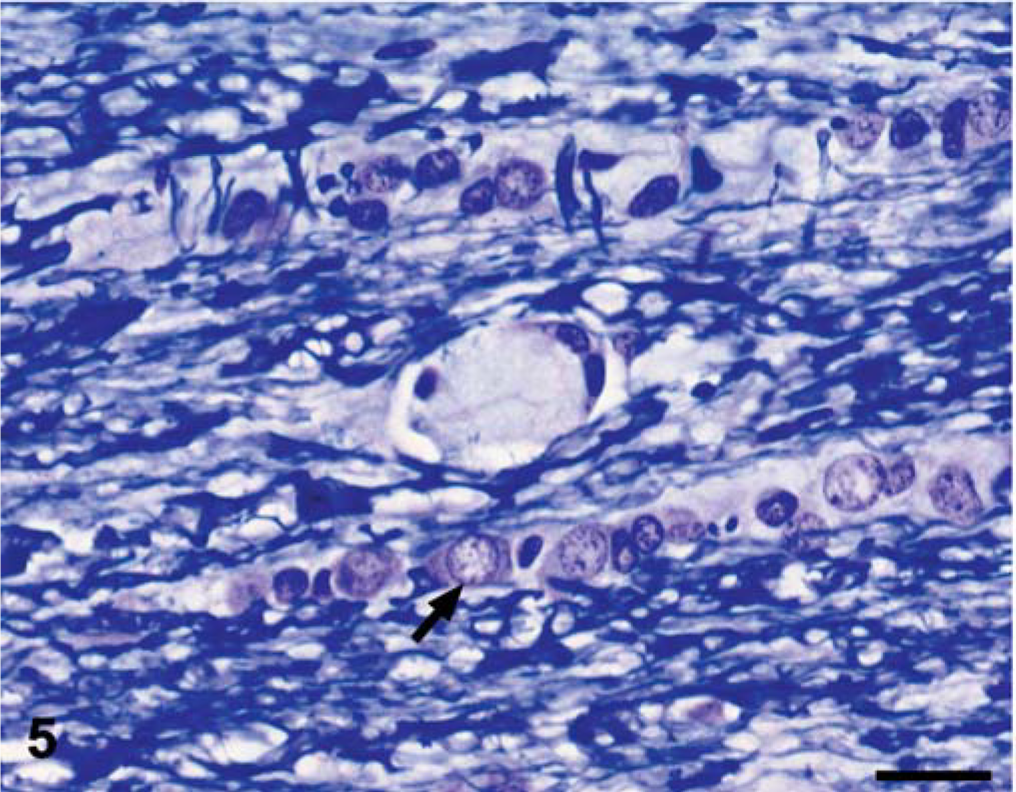
Spinal cord; dog No. 2. The plaques were unstained with Luxol fast blue, whereas the surrounding normal myelin stains deeply. Note the mildly hypertrophied oligodendroglial cell (arrow), which commonly occurred within the interfascicular chains. Luxol fast blue. Bar = 20 µm.
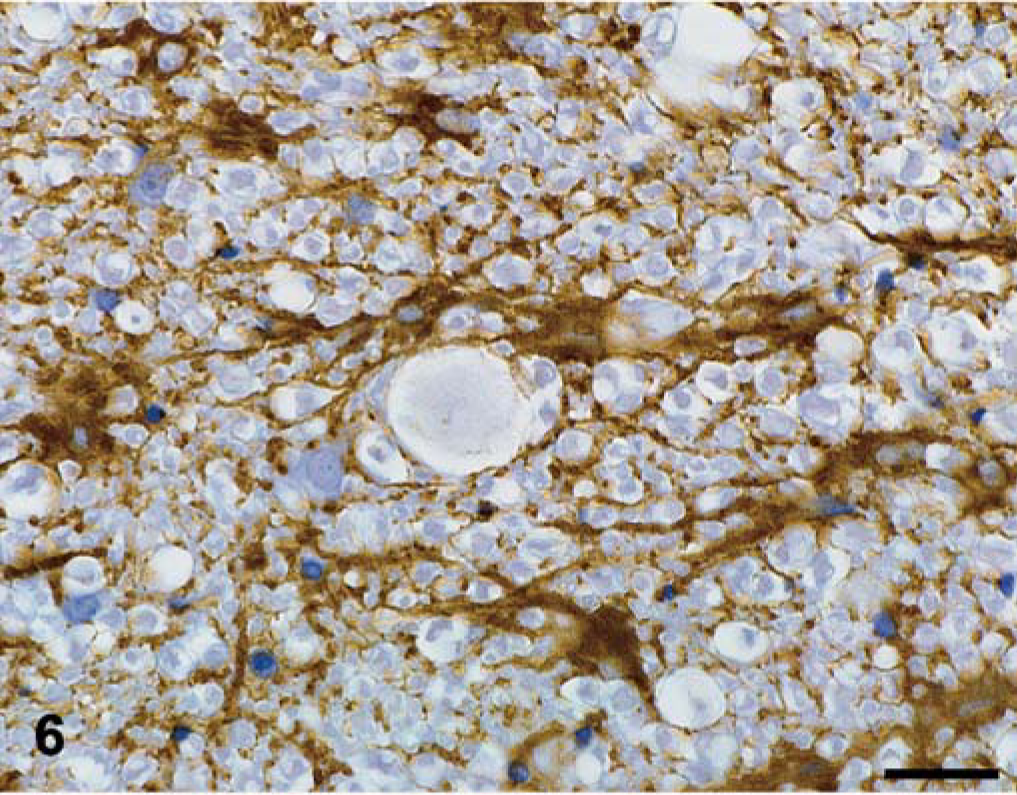
Spinal cord; dog No. 2. The plaques are devoid of astrocytic processes. Astrocytes in the surrounding white matter are mildly hypertrophied and contain GFAP-positive processes. GFAP immunohistochemistry counterstained with hematoxylin. Bar = 25 µm.
In dog No. 1, the plaques were most numerous in the cerebellar medulla, the optic tracts, and the dense white matter tracts of the brainstem (crus cerebri, pyramids, cerebellar peduncles, spinal tract of cranial nerve V, transverse fibers of the pons, and medial longitudinal fasciculus). Plaques in lesser numbers were present in the internal capsule and were rare in the centrum semiovale, corona radiata, and areas of mixed white and gray matter of the brain stem (reticular formation, thalamus). The spinal cord was not examined. In dog No. 2, the plaques were most numerous diffusely in the superficial lateral funiculi and to a lesser extent the ventral funiculi of the spinal cord. They were present in all segments and especially the cervical segments, with smaller numbers present in the remaining white matter spinal cord tracts and the white matter of the caudal brainstem similar to dog No. 1. In contrast with dog No. 1, few to no plaques were present in the cerebellar medulla or rostral to the mesencephalon.
Immunohistochemistry
The neurofilament stain identified single axons coursing through the center of the plaques, which ranged from thin to normal caliber. No significant axonal degeneration was appreciated in or immediately adjacent to the plaques. Occasional GFAP-positive hypertrophic astrocytes were observed adjacent to the plaques, but astrocytic processes within the plaques were rare (Fig. 6). Cells within and adjacent to the plaques were negative for MAC387, a marker of activated macrophages.
Electron microscopy
Electron microscopic studies focused on plaques identified in 1-µm-thick plastic sections. Tissue preservation was less than ideal, as only formalin-fixed tissue was available. Plaques were composed of an axon, which was abutted by numerous processes of glial cells (Figs. 7, 8). Where only short segments of an axon were observed, a myelin sheath was lacking, whereas when longer profiles of axons were seen, it was apparent that the myelin deficit was segmental and often asymmetric (Fig. 7). The areas where axons were unmyelinated appeared to involve expanded nodes of Ranvier, as loosened paranodal loops were present in the area of myelin loss (Fig. 9). Such axons appeared normal with a complement of filaments and tubules, mitochondria, and smooth vesicles. The numerous cell processes that encircled the axons were of light to medium electron density and varied from bundles of slender, filiform processes to broader expanses of cytoplasm (Fig. 8). Numerous myelin figures and myelin membranes were found between the cell processes (Fig. 7) and internalized within the glial cell cytoplasm. The slender processes contained microtubules, a feature of oligodendrocytes (Fig. 9). While occasional oligodendrocyte nuclei were encountered with a perinuclear zone of cytoplasm, processes of reactive astrocytes with glial filaments were rare and mononuclear macrophages with lysosomes and vesicles were absent.
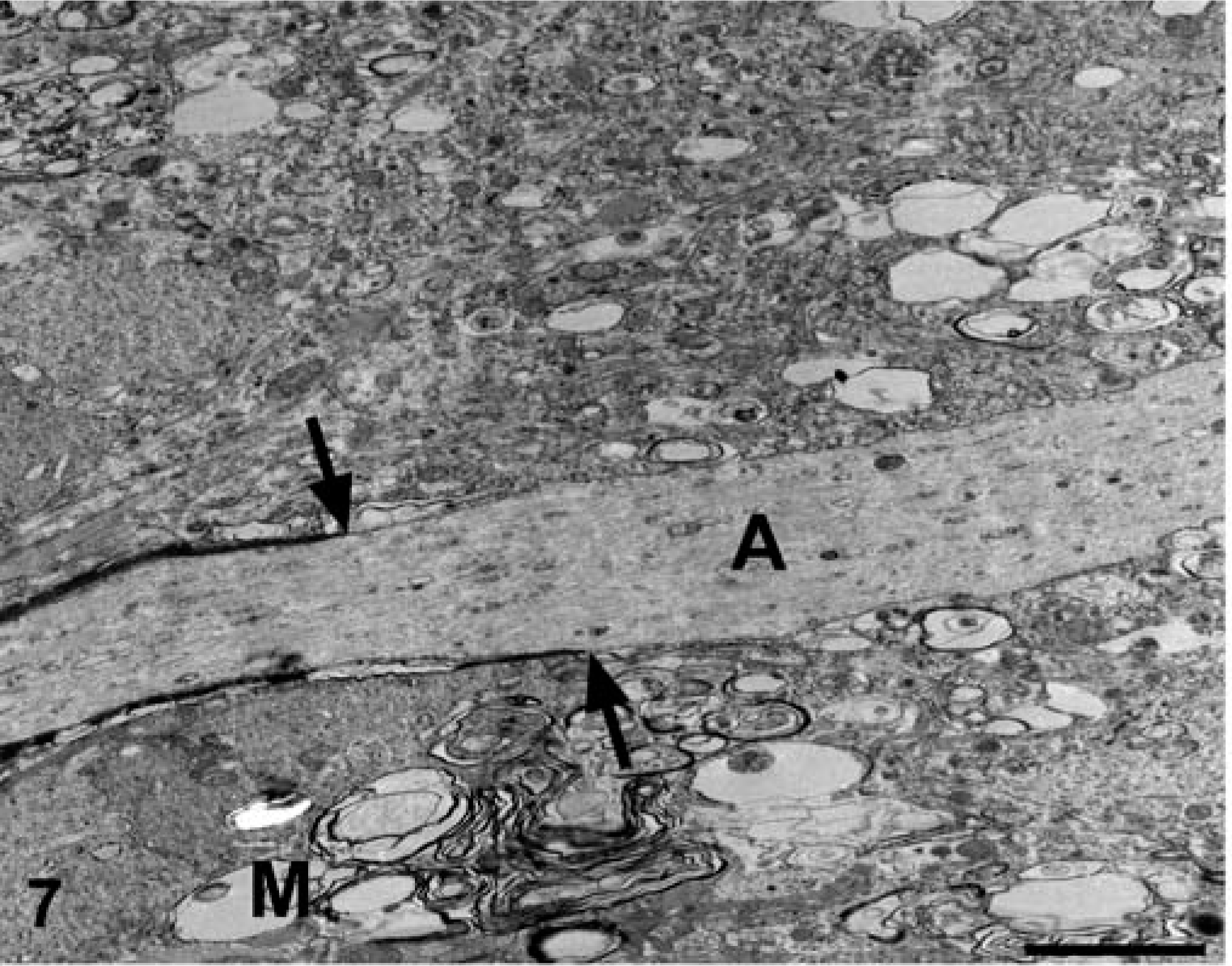
Brain; dog No. 1.The axon (A) is segmentally devoid of myelin and the narrowed myelin sheath ends asymmetrically (arrows). The adjacent white matter is egionally devoid of other axons and filled with irregular cellular processes and myelin debris (M). Electron micrograph. Bar = 2.5 μm.
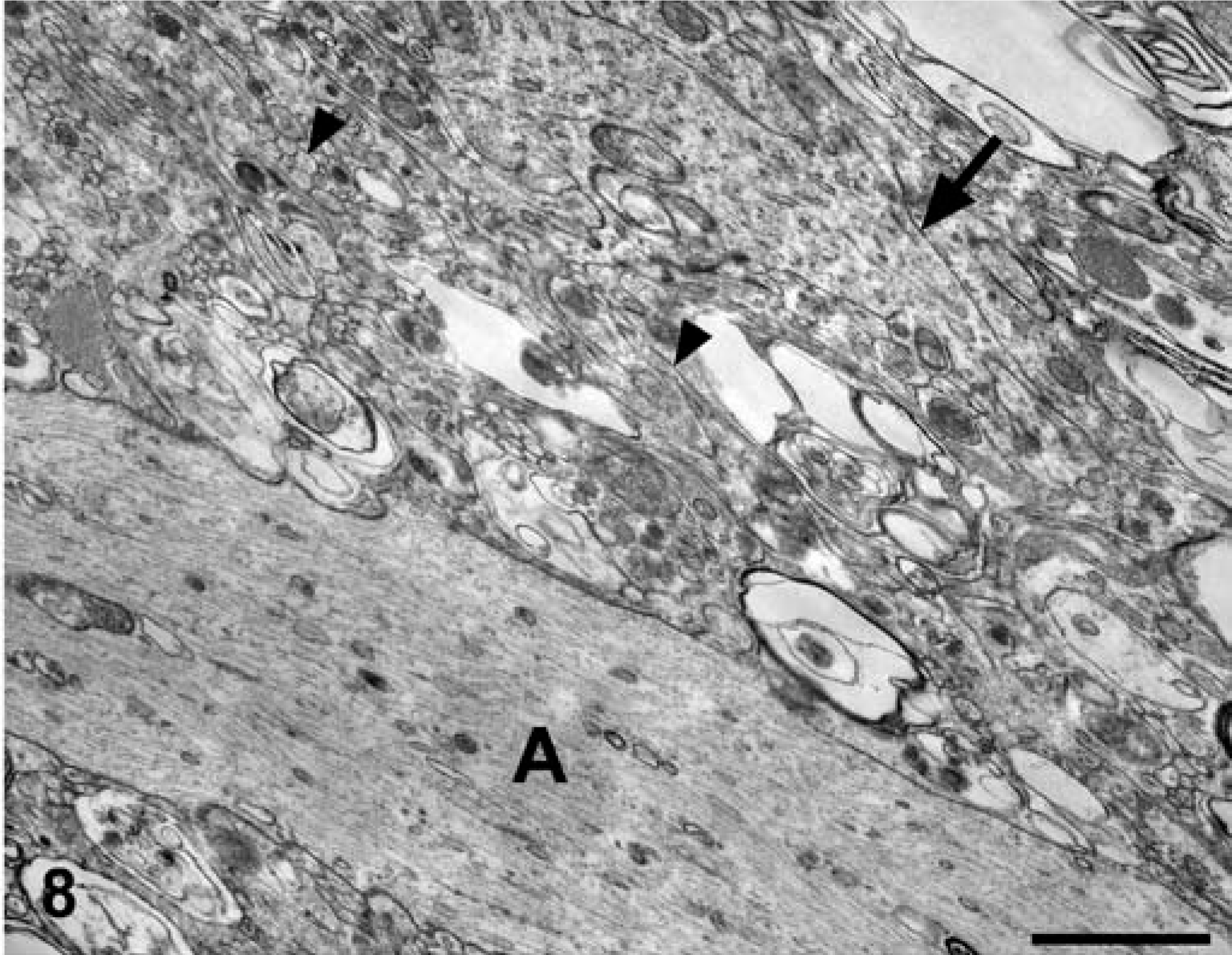
Brain; dog No. 1. The axon (A) is encircled by numerous, slender, filiform cell processes (arrowheads) and broader expanses of glial cell cytoplasm (arrow). Numerous myelin figures and myelin membranes were found between the cell processes and internalized within the glial cell cytoplasm. Electron micrograph. Bar = 1 μm.
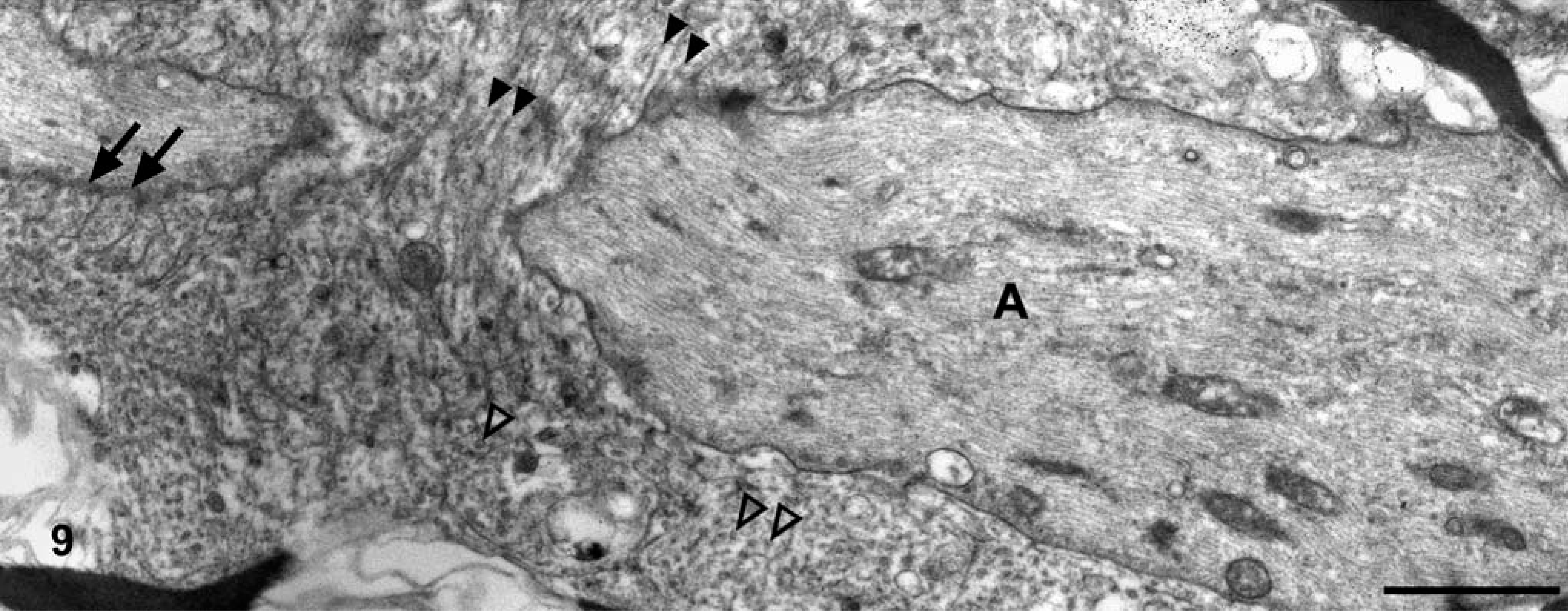
Brain; dog No. 1. Expanded nodal areas of an axon (A) are encircled by cellular processes, which often contain one or several microtubules, a marker of oligodendroglial cells. The microtubules are often captured in longitudinal (solid arrowheads) or transverse (open arrowheads) section. Note the remnants of lateral loops (arrows), indicating a paranodal region. Electron micrograph. Bar = 1 μm.
Discussion
Two male Bullmastiff dogs were evaluated for a diffuse neurologic disorder that presented early in life and progressively worsened over approximately 2 years in the first animal and approximately 3 months in the second. The differential diagnosis included lysosomal storage diseases; axonopathies, including neuraxonal dystrophy, metabolic encephalopathy, and diffuse white matter disorders, such as hypomyelinogenesis and leukodystrophy. 5, 10 Other diffuse CNS conditions, such as meningoencephalomyelitis due to canine distemper, Cryptococcus neoformans, Toxoplasma gondii, Neospora caninum, or granulomatous meningoencephalomyelitis (GME) were considered much less likely due to the slowly progressive and insidious nature of the clinical signs, the lack of systemic signs of illness, and the absence of pleocytosis on CSF examination performed in one dog. 5, 10
The microscopic lesions in these two dogs are novel in our experience of canine neuropathology. The critical findings consisted of pallid eosinophilic plaques that were largely confined to strictly white matter areas of the CNS or areas of mixed white and gray matter in the brainstem and the spinal cord, with extension rostrally into the internal capsule and the cerebral white matter in one animal. These plaques lacked normal myelin sheaths, but evidence of myelin breakdown and phagocytosis was not appreciated by light microscopic examination.
Electron microscopy revealed that the plaques consisted of irregular, hyperplastic tongues and processes of glial cells, interpreted as oligodendrocytes, centered on segments of axons lacking a myelin sheath. The lack of myelin was focal and, in one figure, the proliferative cell processes were adjacent to the lateral loops normally present at the paranode, consistent with an expanded node of Ranvier. That this process was simply primary demyelination, as may be seen in canine distemper, for example, in which myelin loss can be segmental, 6 was considered unlikely due to the lack of myelin phagocytosis by microglia or monocytes, the absence of reactive astrocytosis, and, in particular, the presence of the unusual proliferated oligodendroglial processes that abutted the axon.
These histologic and electron micrographic lesions are morphologically very similar to a well-characterized, presumably inherited leukodystrophy in the Charolais breed of cattle. To reflect the predominant clinical signs, this disorder is known as progressive ataxia, while ultrastructural studies showed an underlying oligodendroglial dysplasia. In cattle, nearly all major white matter areas of the CNS may be affected, with the highest density of plaques in the cerebellar medulla, the internal capsule, and the lateral and ventral funiculi of the spinal cord. 2 When we compared slides of the bovine and canine disorders, the plaques in the affected Charolais are much larger, more numerous, and more readily seen than in the Bullmastiff, and they are more strongly eosinophilic with HE staining. In addition, it has been reported that the plaques in cattle are luxol fast blue positive, 2 in contrast with the plaques in this canine disease, which are negative. This difference may simply represent variations in staining protocols, but the possibility that this represents a subtle difference in plaque composition cannot be excluded.
In affected Charolais cattle, it has been suggested that the development of the white matter plaques may represent a selective CNS oligodendrocyte abnormality that spares Schwann cells, which produce the peripheral nervous system myelin. 2 The regional demyelination of axons is believed to represent expanded nodes of Ranvier, where the formation of the paranode and lateral loops is abnormal. Accordingly, the term segmental dysmyelination has been invoked 2 for what was believed to be a unique abnormality in Charolais cattle but is now identified in Bullmastiff dogs. Associated with the failure of normal node of Ranvier formation, the hypertrophic oligodendrocytes produce voluminous cell processes that invest and encircle the axon. Presumably, normal myelin at the margins of the expanding node may degenerate, as was seen as occasional myelin figures in the Bullmastiffs and has been recorded in the Charolais. Some of these myelin droplets appeared to be within oligodendrocyte cytoplasm, which may represent either phagocytosis or (theoretically) aberrant myelin synthesis by these cells. Progressive widening of the nodes of Ranvier will disrupt and finally abolish saltatory conduction, resulting in greatly delayed and asynchronous propagation of nerve impulses in affected fiber tracts.
The distribution of plaques in the brains of both dogs was similar in the mesencephalon, cerebellum, pons, and medulla. Dog No. 1 had greater involvement of the cerebral white matter, internal capsule, diencephalon, and optic tracts. At the time of euthanasia, dog No. 1 was 18 months older than dog No. 2, which may account for the wider distribution of plaques. In dog No. 2, the lateral and ventral funiculi of the spinal cord were the most severely affected areas, which is broadly consistent with the clinical signs of upper motor neuron/general proprioceptive deficits. Because the first dog had also presented with tetraparesis and ataxia, it also probably had pronounced spinal cord involvement; unfortunately, the spinal cord was not available for study. As the clinical signs advanced, both animals developed a diffuse, whole-body tremor that was worse when the animals were excited. This is most consistent with a diffuse CNS disorder and in accord with the distribution of the plaques at the time of necropsy, especially as seen in the older animal. The lesion distribution in these two animals is similar to, but not quite as extensive as that reported in Charolais cattle.
As the more common leukodystrophies in animals (globoid cell leukodystrophy in Cairn Terriers and West Highland White Terriers and cavitating leukodystrophy in Dalmations 1, 12 ) are breed associated, a hereditary basis seems likely, as for the leukodystrophy in Charolais cattle. 3, 4, 13 Rare leukodystrophies are also described in cats, primates, and mice, 9, 11 for which a genetic basis is also probable. The pedigrees of these two Bullmastiff dogs are unknown and whether their residence in north central New York State is coincidence or reflects a common heritage remains to be determined. Recognition of additional cases and the determination of their pedigrees will allow a potential inherited basis for this syndrome to be proposed or discarded. This canine disorder is apparently unique to veterinary medicine as, to our knowledge, there is no comparable condition reported in humans.
Footnotes
Acknowledgements
We thank Mrs. Zipporah Fleischer for her support of canine neurology research at Cornell University's College of Veterinary Medicine, Alexis Wenski-Roberts for her assistance with the images, and Anita Aluisio for her assistance with the electron microscopy.