
Abstract
Erythropoietin (EPO) is a cytokine primarily involved in the regulation of the erythropoiesis. Recently, it has been demonstrated that EPO and its receptor (EPOR) are expressed in several neoplastic cell lines and solid tumors. Furthermore, in vitro and in vivo studies have shown that EPO could promote human breast carcinoma growth by means of the binding with its receptor, although a clear function for EPO in this setting has not been yet established. While the human medical literature has been accumulating strong evidence on EPO's role in oncogenesis, to date, there are no veterinary reports focusing on such an issue. The aim of the present study was to investigate the immunohistochemical expression of EPOR in canine mammary gland dysplastic and neoplastic lesions. Our results show a weak to moderate EPOR expression in dysplastic glands, being immunoreactivity enhanced as the lesion shows an increasing malignant pattern. On the basis of these findings, this study describes, for the first time, the evidence for EPOR expression in canine mammary gland tumor and suggests a feasible EPO's role for canine mammary tumor progression.
Erythropoietin (EPO) is a glycoproteic hormone, production of which is induced by hypoxia inducible factor-1 (HIF-1) after hypoxic stimuli. 5 EPO belongs to the cytokine superfamily and acts through its receptor (EPOR), a member of the type-I cytokine receptor family. 12 The binding of EPO to its classical receptor results in proliferation, differentiation, and survival of erythroid progenitor cells by inhibition of apoptosis. 8
The hormone EPO was cloned in 1985 and, after that, was rapidly utilized in clinical practice. Over the past several years, its recombinant form (rHuEPO) has greatly enhanced the management of anemia resulting from chronic renal failure and neoplastic diseases, thus substantially improving the quality of life of human and animal beings. Canine EPO shows a strict molecular analogy with the human EPO, and rHuEPO is used in dogs for the treatment of renal-induced anemia and bone-marrow aplasia. 11 Although EPO has been long known for its erythropoietic action, recent in vitro and in vivo studies have demonstrated that EPO-mediated mitogenic and antiapoptotic effects involve all the cells expressing functional receptors for EPO such as endothelial cells, neurons, myoblasts, and kidney epithelial cells. EPO shows pleiotropic effects and acts as endogenous mediator of adaptive tissue response to metabolic stress protecting tissues from injuries. 4
Recently, the EPO/EPOR complex has been identified in several neoplastic cell lines and solid tumors. 14 In particular, EPOR detection in human mammary tumors has raised the suspicion that rHuEPO administration for the treatment of anemia may stimulate tumor proliferation. In this regard, in vitro evidence has shown that EPO/EPOR complex might be involved in the growth, viability, and angiogenesis in tumor. Although this effect has initially been shown to be quite modest in vitro, 1 recent studies suggest that this potential may be underestimated because most tumor cells produce EPO. It is worrisome that a large phase-III trial of rHuEPO treatment in human metastatic breast cancer was recently halted due to a significant increase in tumor growth. 9
The present study provides the first evidence for EPOR expression in canine mammary gland neoplastic lesions and suggests a feasible involvement of EPO in tumor progression.
Twenty canine mammary glands bearing various histotypes of lesions and five normal mammary glands were examined. According to the WHO animal tumor classification, the lesions included 10 hyperplastic/dysplastic lesions and 10 carcinomas. Samples were formalin fixed and paraffin embedded. Five-micrometer-thick sections were cut and deparaffinized in xylene and rehydrated in graded alcohols. Slides were steamed in 0.01 mol/L sodium citrate buffer, pH6, in a microwave oven for 15 minutes. Endogenous peroxidase activity was quenched by 0.3% hydrogen peroxide in methanol for 30 minutes while aspecific proteic reactions were blocked by incubation with horse and goat normal serum for 30 minutes. Slides were then incubated overnight at 4°C with anti-EPOR (rabbit polyclonal; clone C-20, Santa Cruz, 1 : 300 dilution) antibody. The specificity of the receptor immunostain was further supported by using a murine monoclonal anti-EPOR antibody (R&D Systems, 1 : 50 dilution). Slides were then incubated for 30 minutes at room temperature with horse anti-mouse and goat anti-rabbit biotynilated IgGs (Vector Laboratories), developed by the ABC method (Vectastain Elite ABC kit, Vector Laboratories), revealed with Vector Nova Red (Vector Laboratories), and counterstained with hematoxylin. For each sample, negative controls were also performed by omission of the primary antibody or substitution with normal mouse or rabbit IgGs. For each sample, 10 sections were randomly selected. Images were photographed through a microscope and evaluated by using image analysis software. Cells that contained red color with cytoplasmic and/or membrane immunoreactivity were identified as positive. 3 An investigator blinded to the experimental groups from which each sample was obtained conducted this assessment. The number of EPOR-positive cells was expressed as the mean labeled cell count ± standard deviation (SD). The numbers of EPOR-positive cells detected in normal tissue, hyperplastic/dysplastic lesions, and carcinomas were compared using the unpaired Student's t-test. Differences were considered significant at a P value of less than 0.05.
As far as the results are concerned, EPOR expression was almost undetectable in normal mammary gland (Fig. 1A). Conversely, EPOR immunoreactivity was found in epithelial cells of preneoplastic and neoplastic lesions showing an increase in intensity as the lesion became more severe. In hyperplasia/dysplasia, EPOR immunoreactivity was seen as a granular, not homogeneous, stain, localized in the whole cytoplasm or at one cellular pole (Fig. 1B). Intensity of the reaction was stronger in ducts and lobuli distended by a marked epithelial proliferation rather than in unaltered epithelium (Fig. 1C). In frankly malignant neoplastic lesions, immunoreactivity was granular and homogeneous with membranous or cytoplasmic localization. EPOR expression showed an increased intensity of stain in numerous epithelial neoplastic cells. EPOR was homogeneously localized in neoplastic lobuli and tubules, with a qualitatively stronger staining in areas showing more nuclear atypia (Fig. 1D). Furthermore, in areas of tumors with a solid pattern, clusters of EPOR-positive cells were mainly observed around tumor vessels (Fig. 1E).
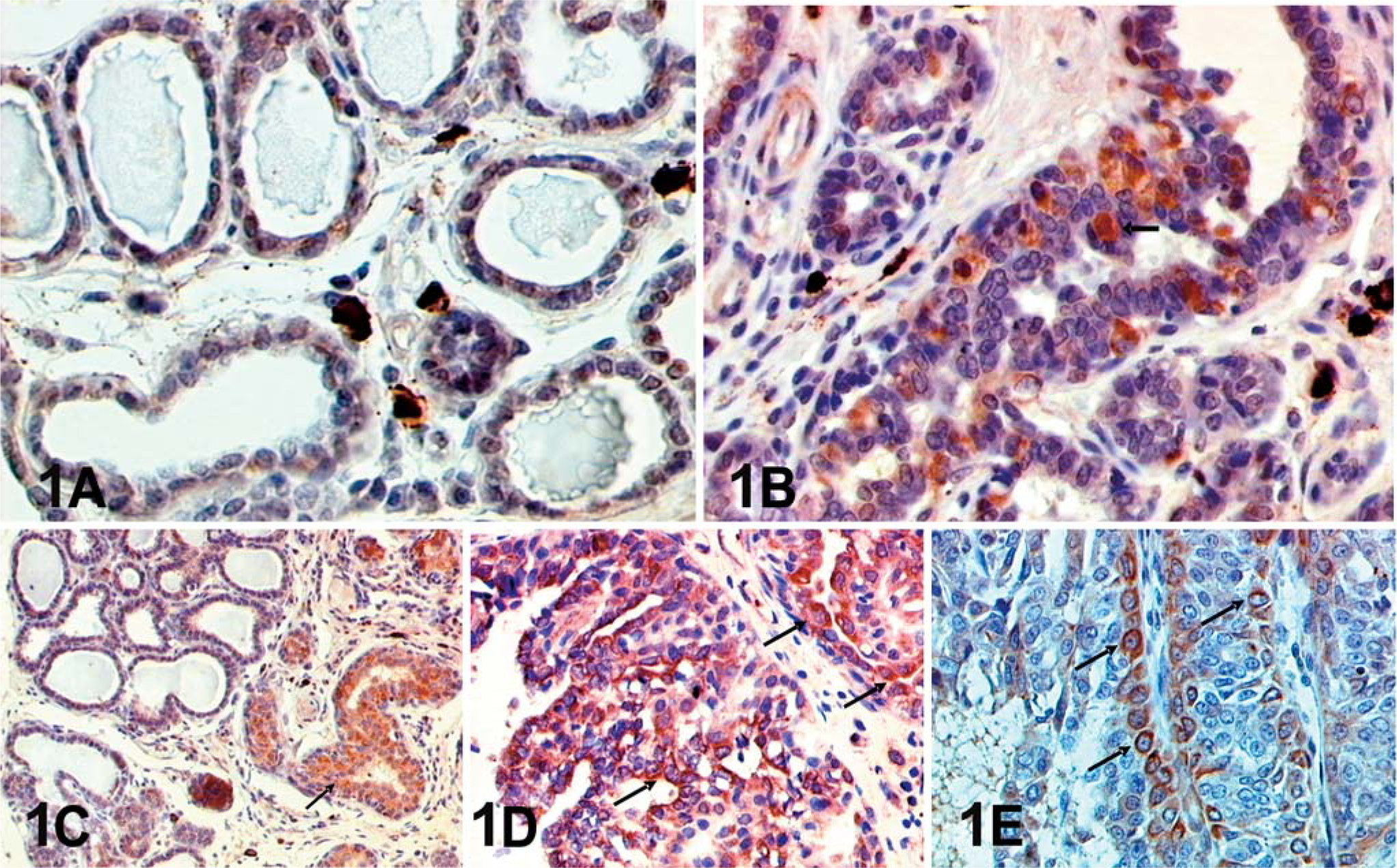
EPOR expression in mammary gland specimens.
Immunoreactive epithelial cells for EPOR were significantly higher in carcinoma compared with hyperplasia/dysplasia lesions (P < 0.05). Statistical differences were also observed between normal gland and preneoplastic lesions (P < 0.05). Data are summarized in Table 1.
Quantification of EPOR immunoreactive epithelial cells.

∗Significant statistical differences for EPOR-immunostained cells between normal mammary gland and hyperplasia/dysplasia lesions (P < 0.05).
∗∗Significant statistical differences for EPOR-immunostained cells between hyperplasia/dysplasia lesions and carcinoma (P < 0.05).
This study investigated, for the first time, the EPOR expression in canine mammary gland tumors by immunohistochemistry. Our findings demonstrated that EPOR is expressed both in preneoplastic and neoplastic canine mammary lesions, where EPOR immunoreactivity was found to increase in expression as long as the lesion showed an increasing malignant pattern.
These results, although based on few cases, support the hypothesis that EPOR is somewhat involved in the tumor growth process even if the mechanisms by which it acts are still to be clarified. More than an assumption, EPOR overexpression in severe neoplastic lesions seems to be in agreement with recent observations showing EPOR upregulation after experimental ischemic stroke, 2 spinal cord injury, 7 and demonstrating that EPO administration can rescue ischemic and hypoxic neurons following cerebral ischemia and subarachnoid hemorrhage. 2, 6 The increasing in expression of EPOR in solid tumors could similarly improve the hypoxic or ischemic survival of cancer cells by inducing the transcription of the EPO gene via activation of the HIF-1 transcription factor. 5 Overexpression of HIF-1 has been demonstrated in human cancers and its identification well correlated with increased cell proliferation, angiogenesis, and a malignant phenotype. 10 On the other hand, sustained activation of the EPOR by EPO is known to inhibit apoptosis in hematopoiesis. 8 Taken collectively, EPO signaling via functional EPOR may reduce apoptosis in cancer cells, thus providing a potential tumor growth pathway.
Although an increasing number of investigations are providing new insights about the EPO/EPOR role on human cancer, in veterinary medicine, to date, there are no data dealing with this important issue. Few studies have reported the existence of tumor-producing EPO in dogs without histopathologic evidence of EPOR expression and localization. 13 Because rHuEPO administration is the treatment of choice for cancer-related anemia both in humans and animals, a deep understanding of the EPOR role in cancer progression is necessary.
In conclusion, based on our results and according to the literature, the EPOR expression could provide significant implications for the treatment and prevention of cancer because exogenously administered EPO could conceivably promote proliferation and survival of cancer cells expressing functional EPOR.
Footnotes
Acknowledgements
This work has been partially supported by Fondo di Incentivazione alla Ricerca di Base (FIRB-cod. RBNE01TBTR) from the Ministero dell'Istruzione, Universita 'e Ricerca, Rome, Italy.