
Abstract
The objective of this study was to evaluate stomachs of 2-year-old Syrian hamsters that were naturally colonized by multiple Helicobacter species including Helicobacter aurati. A previous report on 7- to 12-month-old Syrian hamsters described chronic gastritis and intestinal metaplasia, a putative preneoplastic lesion in the stomach, without cancer. This report describes an invasive adenocarcinoma at the pyloric-duodenal junction in one of nine hamsters at a site of helicobacter-associated inflammation and marked intestinal metaplasia. Ceca of nine of nine animals were culture positive and polymerase chain reaction positive for Helicobacter spp. Furthermore, immunohistochemical analysis of the stomach using a H. pylori polyclonal antibody detected positive-staining bacteria within the pyloric region of three of nine hamsters including the neoplastic glands. However, argyrophilic bacteria were demonstrated only within the stomach of the hamster with gastric adenocarcinoma. This is a first report of gastric adenocarcinoma in helicobacter-infected hamsters. Syrian hamsters appear suitable as potential model for studying development of helicobacter-associated gastric adenocarcinomas.
Approximately 50% of the world's population is infected with Helicobacter pylori, with a greater prevalence in developing countries. 6, 14 Chronic H. pylori infection often leads to duodenal and gastric ulcers and is associated with an enhanced risk of developing gastric adenocarcinoma and lymphoma. 9, 14 This risk is markedly greater in patients who develop helicobacter-associated atrophic gastritis and intestinal metaplasia. 14, 23 Although gastric adenocarcinoma is associated with the presence of H. pylori in the stomach, only a small fraction (5%) of colonized individuals develop this common malignancy, thereby suggesting that H. pylori strain and host genotypes influence the risk of carcinogenesis. 14 Despite this low percentage of individuals with gastric cancer and concurrent helicobacter organisms, it is believed that the prevalence of helicobacter infection could be actually higher. This is especially so because at the time of gastric carcinoma diagnosis, colonization of the stomach by H. pylori is reduced or eliminated completely. 14 For these reasons, H. pylori has been classified by the World Health Organization as a grade-I carcinogen. 9, 14
Gastric adenocarcinomas have two histologically distinct variants, intestinal type and diffuse type, both of which are associated with chronic H. pylori infections. 10, 14 Both types of carcinoma have distinct epidemiological and pathophysiological features. 10, 14, 23 Diffuse-type gastric adenocarcinoma is less frequent and typically comprises individual neoplastic cells that do not form glandular structures and are not associated with atrophic gastritis and intestinal metaplasia. 10, 14 Intestinal-type adenocarcinoma is more common, well characterized, and typically follows a multistage process including chronic superficial gastritis, chronic atrophic gastritis, intestinal metaplasia, and dysplasia culminating in cancer. 12 This is similar to the multiple, distinct, histologically identifiable lesions observed during colorectal tumorigenesis. Despite this similarity, the consistent mutational events that correlate with the multiple stages in colorectal cancer have not been reported during the genesis of intestinal-type gastric adenocarcinomas. 8, 11, 23, 24
Modeling of Helicobacter infections in rodents have resulted in evidence that clarifies the critical role of several host- and environmental-dependent factors. These factors, to mention a few, include the host immune response (Th1 versus Th2), gastrin, transforming growth factor-α, Smad4, virulence of the bacterium, gastrointestinal parasitism, salt, carcinogens, and antioxidants in the diet. 2, 3, 19, 25 However, despite the usefulness of animal models in understanding the pathogenesis of human disease, most animal models do not recapitulate the human disease completely. For example, although subclinical to chronic progressive gastritis associated with different Helicobacter species has been demonstrated (experimentally or naturally) in several animal species including mice, rats, Mongolian gerbils, guinea pigs, ferrets, cats, dogs, pigs, macaques, and hamsters, most of these animals (except Mongolian gerbils, ferrets, C57BL/6, and INS-GAS mice) do not develop gastric cancer. 4, 12, 13, 19, 21, 25 Furthermore, only gerbils and hamsters infected with Helicobacter spp. develop intestinal metaplasia similar to humans. 7, 12, 13 Previous studies from our laboratory described chronic gastritis-associated preneoplastic lesions, without cancer in 7- to 12-month-old Syrian hamsters that were naturally infected with Helicobacter species. 12, 13 This report is an extension of a previous study wherein hamsters were housed in the same animal facility and were from the same commercial vendor as those in this report. In this study, a group of nine geriatric hamsters (aged 2 years) naturally infected with Helicobacter spp. were evaluated.
Nine retired breeder Syrian hamsters (Mesocricetus auratus) from a single vendor were housed in an Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC)–approved animal facility at Massachusetts Institute of Technology. The animals evaluated in this study included seven males and two females of approximately 2 years of age that were not experimentally manipulated. The female hamster presented in this case report had a history of weight loss but otherwise was clinically normal before necropsy. Complete necropsies were performed after euthanasia with carbon dioxide, and samples were collected for culture and polymerase chain reaction (PCR). All tissues were immersion fixed in 10% neutral-buffered formalin. Paraffin-embedded 5-μm sections were stained with hematoxylin and eosin. Gastric sections were also stained with Warthin-starry silver stain to demonstrate argyrophilic bacteria, whereas the Alcian blue/periodic acid–Schiff (PAS) stain, pH 2.5, was used to stain for mucins. Immunohistochemical (IHC) analyses of gastric tissues were performed using an anti–Helicobacter pylori rabbit polyclonal antibody (Novacastra, Newcastle upon Tyne, UK) following the manufacturer's directions. PCR-based analyses of cecal tissue and contents were performed in all the nine animals, using helicobacter-specific primers as described previously. 12, 13 The animals were considered positive or negative on the basis of IHC, silver stain, and PCR analyses. One-month-old Syrian hamsters without gastric lesions were used as negative controls for IHC. Gastric pathology was evaluated by a veterinary pathologist. Gastritis and intestinal metaplasia were graded according to criteria described previously. 12
One female hamster had a focal 0.3–cm-diameter erythematous mucosal nodule at the pyloric-duodenal junction and focally extensive thickening of the pyloric mucosa. This nodule was histologically consistent with an infiltrative gastric adenocarcinoma (Fig. 1). This infiltrative, transmural, unencapsulated tumor that partially effaced the gastric mucosa, extended from the luminal surface to the serosa, and emerged from an area of increased goblet cell differentiation (intestinal metaplasia). The mass comprised well-differentiated and moderately pleomorphic tubular to tortuous gastric glands supported by adequate fibrous connective tissue stroma. The neoplastic glands had anisokaryotic, vesicular, oval to elongate pseudostratified nuclei (Fig. 2). Mitotic figures were infrequent. There was marked infiltration of the stroma by mixed inflammatory infiltrates, especially foamy macrophages and occasional hemosiderophages, with extension of these infiltrates to the gastric serosa. There was no evidence of vascular invasion or metastasis in this animal. Also, within a few neoplastic glands, there were randomly scattered goblet cells (Fig. 2). The abutting mucosa also demonstrated increased numbers of Alcian Blue–positive goblet cells (intestinal metaplasia) producing predominantly sialomucins (Fig. 3). A few of the neoplastic gastric glands exclusively contained intraluminal, fusiform argyrophilic bacteria with intracellular circular bodies (< 1 μm diameter) (Fig. 4). The fusiform morphology of the bacteria is consistent with the earlier report describing H. aurati. 13 The circular bodies within the bacterial cell wall are most likely polyphosphate granules, which have been described previously in Helicobacter spp. 22 Furthermore, by IHC, we detected intraglandular gastric bacteria, which cross-reacted with H. pylori antibody in three of nine hamsters including the hamster with the focal gastric adenocarcinoma (Fig. 5).
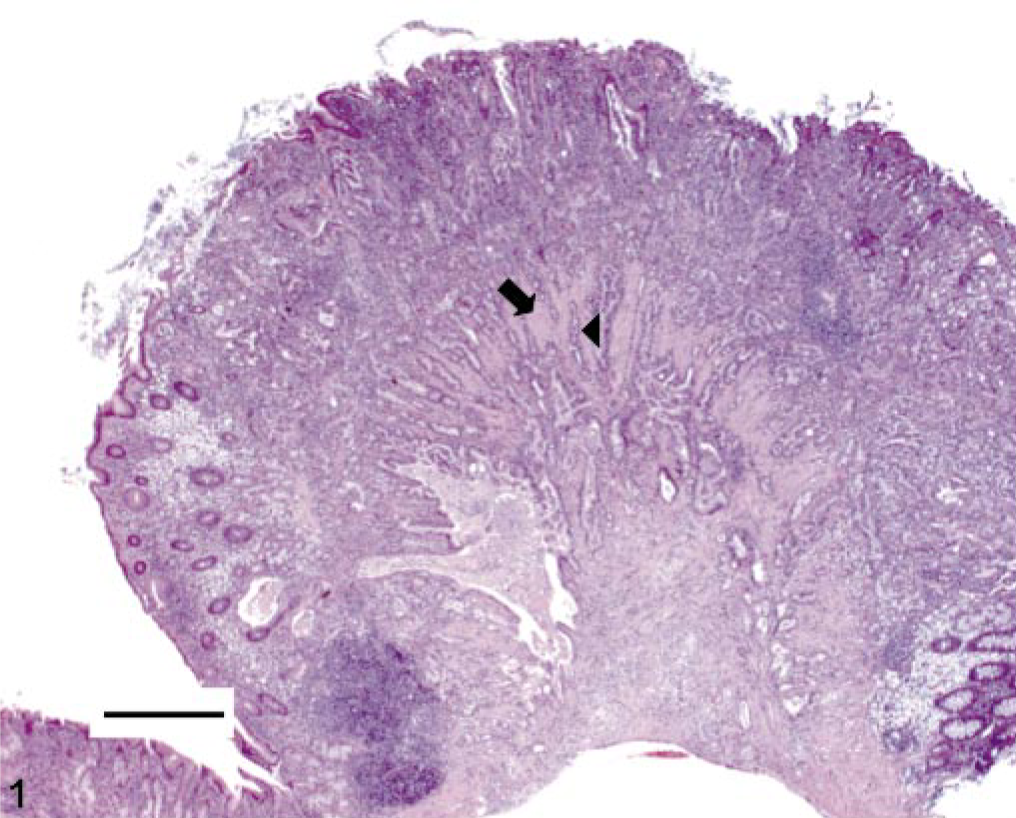
Stomach. Invasive gastric adenocarcinoma arising from the distal antrum-pyloric junction of a 2-year-old Syrian hamster. Note invasion of the muscular tunic (arrow) by neoplastic glands (arrow-head) HE stain. Bar = 500 μm.
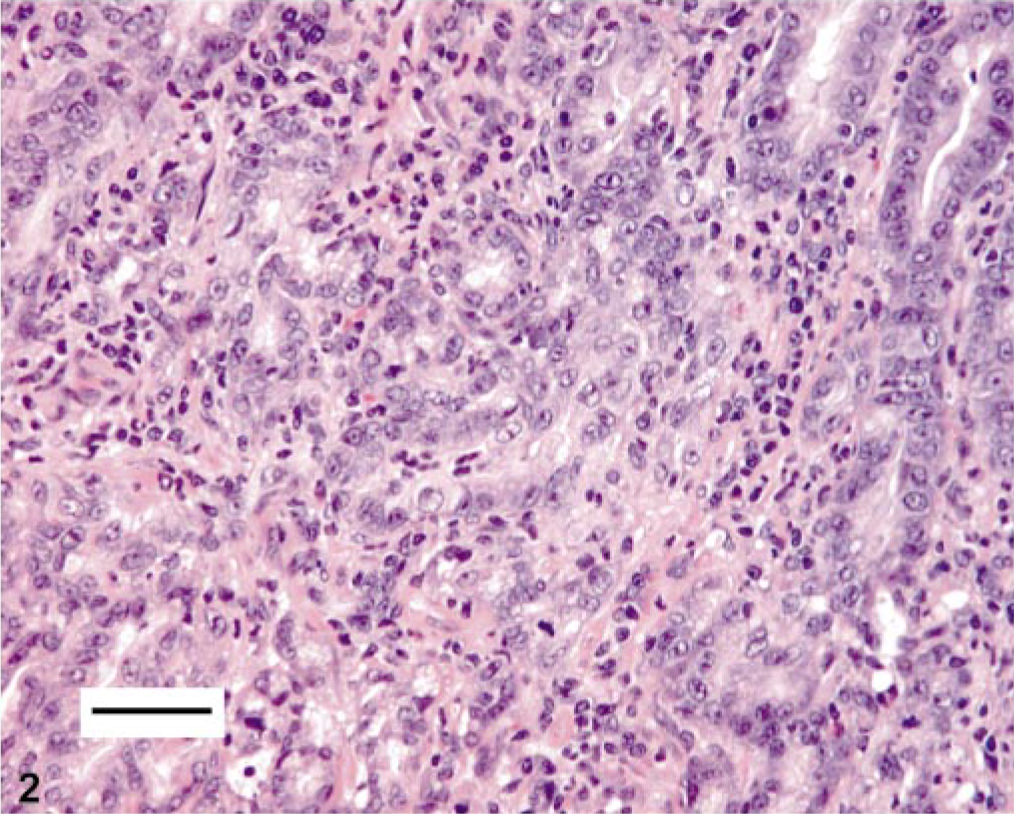
Stomach. Higher magnification of the invasive neoplastic glands within the muscular tunic of the stomach. The neoplastic glands are elongate and tortuous and lined by moderately pleomorphic glands with anisokaryotic and vesicular nuclei that were occasionally pseudostratified. The fibrous connective tissue stroma is infiltrated by moderate to high numbers of mononuclear inflammatory cells. HE stain. Bar = 50 μm.
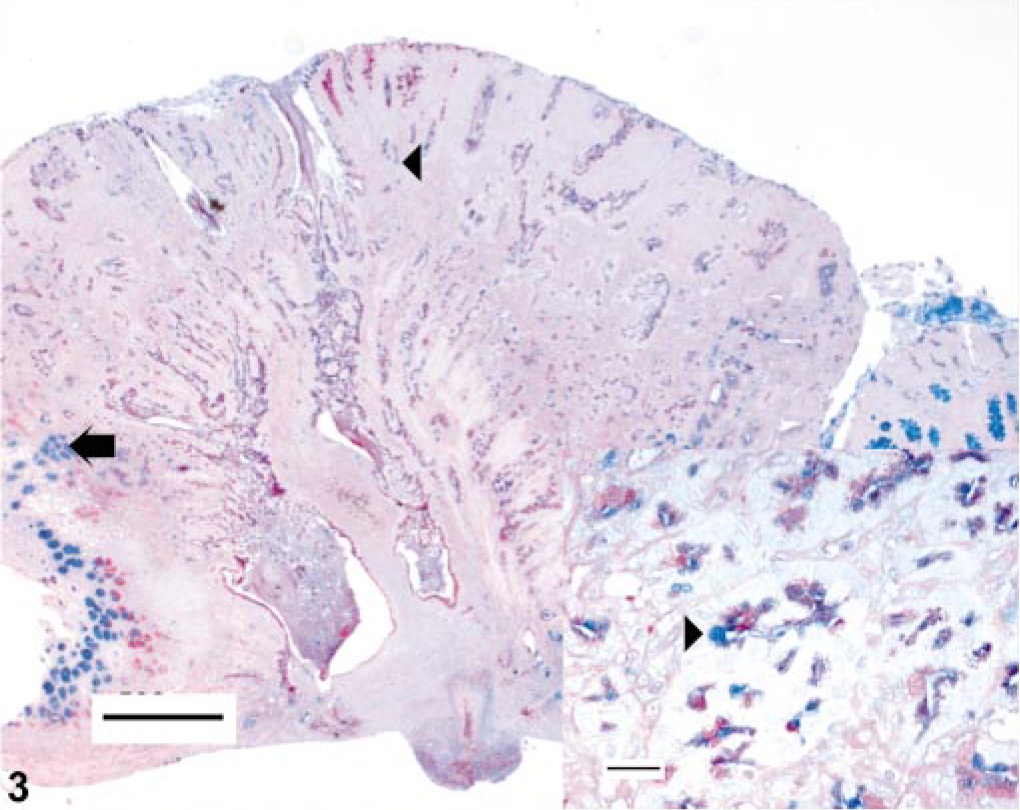
Stomach. Invasive gastric adenocarcinoma arising from the distal antrum-pyloric junction of a 2-year-old Syrian hamster. Goblet cells within the tumor (arrow-head) and within the abutting mucosa (arrow) are depicted. Bar = 500 μm. Inset: Higher magnification of the neoplastic glands with goblet cell differentiation. Alcian Blue, pH2.5, or PAS stain. Bar = 25 μm.
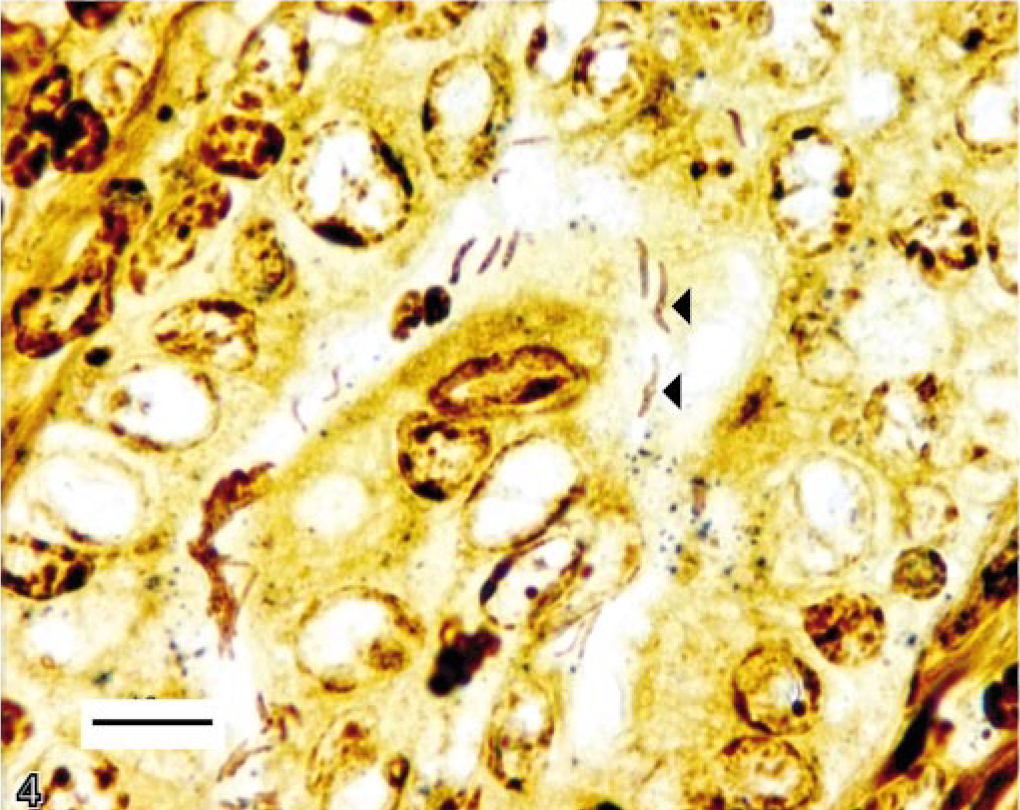
Stomach. Gastric gland within the inflamed gastric mucosa of a 2-year-old hamster with gastric adenocarcinoma. Argyrophilic bacteria are noted within gastric glands. Note intrabacterial circular bodies (arrow heads). Warthin-Starry stain. Bar = 10 μm.
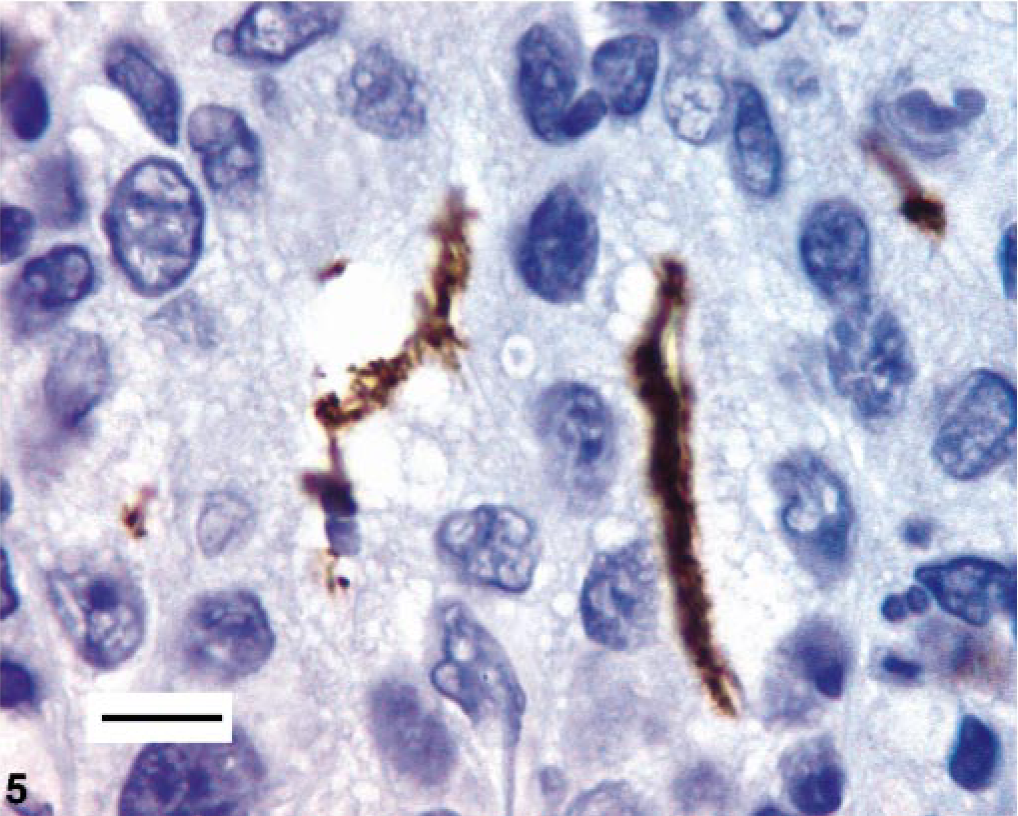
Stomach. Immunohistochemical analysis of the stomach using Helicobacter pylori antibody. Note positive-staining filamentous bacteria. Bar = 10 μm.
All hamsters were positive for Helicobacter species by isolation and PCR of the ceca (data not shown). Eight of nine hamsters including the animal with the gastric adenocarcinoma had locally extensive chronic gastritis with intestinal metaplasia, especially at the distal antrum near the pyloric-duodenal junction. However, only one hamster had grossly detectable, focally extensive thickening of the pyloric mucosa. Although, the overall inflammatory and metaplastic lesions within the stomach were qualitatively and quantitatively comparable with our earlier study reported in 1-year-old hamsters (Fig. 6), we found interesting differences in the inflammation scores between the two studies. Helicobacter-negative animals (by IHC) in the 2-year study had a higher but not significant mean inflammation score (1.83) compared with the helicobacter-negative animals (by silver stain) in the 1-year study (0.62). 12 A similar upward trend in inflammation scores were noted in helicobacter-positive animals in the 2-year study (Fig. 6). It is important to note that helicobacter positivity in the above comparisons for the 2-and 1-year study is based on the less “sensitive” IHC and silver stain, respectively. However, helicobacter was identified either in the ceca or stomachs of all animals involved in both studies by culture, PCR, or Southern blotting (data not shown). Thus, there was a progression of inflammation in this study relative to the 1-year study, albeit not statistically significant.
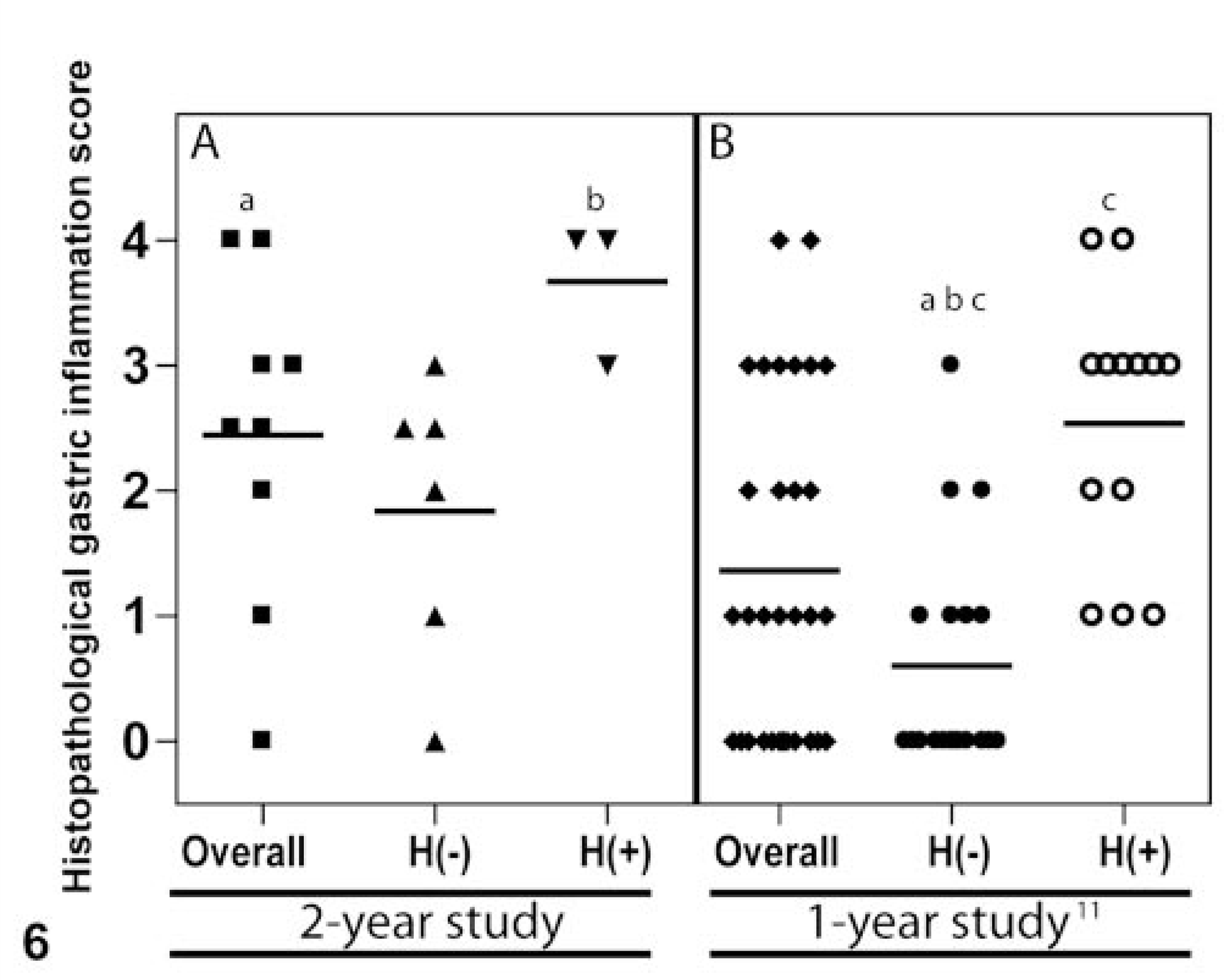
Histological scores for gastric inflammation in 2-year-old (panel A) and 1-year-old (panel B) helicobacter-infected Syrian hamsters (0–4; absent to severe). Horizontal bar depicts the mean inflammation score. Statistical analysis was performed by using Kruskall-Wallis nonparametric test with Dunn's multiple-pair comparison test. Similar letters indicates statistical significance (P < 0.05) between the compared groups across both panels. Helicobacter-negative and -positive stomachs are indicated by H(−) and H(+), respectively. Each shape represents a distinct Syrian hamster.
Chronic gastritis with intestinal metaplasia (a preneoplastic lesion for gastric cancer) associated with naturally occurring gastric colonization by Heliocbacter aurati and two species of unnamed microaerobic bacteria species was documented in 1-year-old Syrian hamsters in our earlier studies. 12, 13 Although, our investigations reported development of putative preneoplastic lesions in aging hamsters, there are no published reports on whether these lesions could progress to cancer. Our study suggests that chronic helicobacter-associated gastritis in hamsters can indeed develop into an infiltrative gastric adenocarcinoma and like humans progresses via multiple stages. Thus, our findings are similar to those observed in chronic H. pylori infections in humans.
The disparate demonstration of Helicobacter spp. within the stomach and ceca of three of nine and nine out of nine hamsters, respectively, is not an unusual finding. In fact, the low incidence of microscopically identifiable Helicobacter spp. in the stomachs of 2-year-old hamsters is consistent with the finding of Patterson et al., 2000, 12 wherein gastric glands with intestinal metaplasia had no argyrophilic Helicobacter spp. The lower gastrointestinal tract in hamsters is thought to be a more likely site of primary infection, with secondary colonization of the stomach during coprophagy. 13 Similar findings have been reported in infection of mice with H. muridarum, an inhabitant of the ileal and cecal mucosa, which can colonize the stomachs of mice and induce inflammation. 20, 22 It is also quite possible that the marked intestinal metaplasia within the pyloric mucosa makes the microenvironment conducive for Heliocbacter spp. to colonize the stomach.
In humans, parietal and chief cell loss (gastric atrophy) in the fundic mucosa results in elevation of gastric pH and subsequent shifts in H. pylori colonization. In mice, a similar shift (antrum to the corpus) in H. felis colonization has been reported with gastric atrophy and elevation of gastric pH. 1 However, in the 2-year-old hamsters, the inflammation and intestinal metaplasia were mostly localized to the antrum, with minimal effect on gastric glands within the fundus. Therefore, it is unlikely that the low detection rate of Helicobacter spp. within the antrum of these ageing hamsters is because of gastric atrophy. Alternatively, it could be speculated that helicobacter-associated intestinal metaplasia in the antrum causes an alteration in the gastric microbiota making it impossible to detect organisms by IHC or special stains. Although methods such as bacterial isolation or PCR (or both) are considered gold standards of helicobacter detection in tissues, whereas IHC and silver stains are considered insensitive, our previous data indicate that there was close correlation between animals that were positive by culture and by silver stains. 12 In addition, our previous study demonstrated a significant correlation between helicobacter infections and gastritis. 12
The lack of helicobacter-free hamsters makes it difficult to definitively establish the role of Helicobacter species in this case of chronic gastritis-associated adenocarcinoma. Antibiotic-mediated helicobacter eradication protocols are not recommended in hamsters because they are highly susceptible to fatal antibiotic-associated typhlocolitis due to Clostridium difficile. 15 Furthermore, in mice, although success in eradication of Helicobacter spp. has been reported with antibiotic therapy, long-term, large-scale studies evaluating the success of such eradication programs have not been reported. 27 In addition, the widespread distribution of Helicobacter spp. among commercial vendor animals and also the fact that hamster pups cannot be foster-raised to achieve germ-free status further complicates the ability to perform complicated experiments designed to test Koch's postulates.
Despite the inability to establish Koch's postulates, emergence of a tumor at a site (pyloric-duodenal junction) with chronic inflammation and prominent intestinal metaplasia is strongly supportive of the role of helicobacter-associated chronic gastritis in the development of this tumor. Furthermore, the significant correlation between the helicobacter-positive stomachs and histopathological inflammation scores is also indicative of the role of Helicobacter spp. in contributing to progressive gastritis in these geriatric hamsters. There is also a precedent for the findings in this study, as demonstrated by gastric helicobacter infection studies in Mongolian gerbils, C57Bl/6, and INS-GAS mice, wherein all infected animals develop chronic gastritis, and 37–80% of these animals subsequently develop adenocarcinomas. 4, 7, 26 Furthermore, studies have shown that H. muridarum, a normal inhabitant of the ileum and cecum of rodents, can colonize the stomach and cause gastritis in ageing rodents. 20, 22
The most common spontaneous gastric neoplasms reported in ageing hamsters (1.5–2.5 years of age) are papillomas in the forestomachs. 5, 16–18 In one carcinogenesis experiment, the control Syrian hamsters between 19 and 22 months of age did not develop gastric lesions. 28 This is the first report to our knowledge that describes progression of preneoplastic lesions to cancer in helicobacter-infected ageing hamsters. Syrian hamsters could therefore offer an attractive model for studying development of helicobacter-associated gastric adenocarcinomas. Because only small subsets of these animals develop adenocarcinomas with time, this model can be used to examine the various factors that can either exacerbate or arrest tumorigenesis.
Footnotes
Acknowledgements
We are grateful for the assistance provided by Dr. Elizabeth Theve and Kathleen Cormier. This work was supported in part by RR07036 (J. G. Fox) and R01A137750 (J. G. Fox).