
Abstract
The objectives of this study were to investigate the interactions between Mycoplasma hyopneumoniae and porcine circovirus type 2 (PCV2) and to establish a model for studying the pathogenesis of and testing intervention strategies for the control of PCV2-associated porcine respiratory disease complex (PRDC). Sixty-seven pigs were randomly assigned to four groups. Group 1 (n = 17) pigs served as controls, group 2 (n = 17) pigs were inoculated with M. hyopneumoniae, group 3 (n = 17) pigs were dual infected with M. hyopneumoniae and PCV2, and group 4 (n = 16) pigs were inoculated with PCV2. Pigs were inoculated intratracheally with M. hyopneumoniae at 4 weeks of age followed by intranasal inoculation with PCV2 at 6 weeks of age. Dual-infected pigs had moderate dyspnea, lethargy, and reduced weight gain. The overall severity of macroscopic lung lesions, PCV2-associated microscopic lesions in lung and lymphoid tissues, and the amount of PCV2-antigen associated with these lesions were significantly (P < 0.05) higher in dual-infected pigs compared with all other groups. Four of 17 (23.5%) dual-infected pigs had decreased growth rate and severe lymphoid depletion and granulomatous lymphadenitis associated with high amounts of PCV2-antigen consistent with postweaning multisystemic wasting syndrome (PMWS). PCV2-antigen in lung tissue was most often associated with M. hyopneumoniae-induced peribronchial lymphoid hyperplasia, suggesting that this is an important site for PCV2 replication in the lung. This study indicates that M. hyopneumoniae potentiates the severity of PCV2-associated lung and lymphoid lesions, increases the amount and prolongs the presence of PCV2-antigen, and increases the incidence of PMWS in pigs.
Keywords
Porcine circovirus type 2 (PCV2) is associated with several disease manifestations in swine including postweaning multisystemic wasting syndrome (PMWS). 1,20,21,33 PMWS is characterized clinically by wasting, decreased weight gain, enlarged lymph nodes, and dyspnea. 20,49 The hallmark microscopic lesions of PMWS are lymphoid depletion or granulomatous lymph-adenitis (or both) and the presence of PCV2 antigen or nucleic acids associated with the lymphoid lesions. 49
PCV2 alone is limited in its ability to induce the full spectrum of disease and lesions associated with PMWS in pigs 3,14,30,41,42,47 and is considered by some to be a ubiquitous opportunist. 49 Porcine reproductive and respiratory syndrome virus (PRRSV)–induced potentiation of PCV2-associated PMWS has been experimentally confirmed. 3,24,47 Porcine parvovirus (PPV) has also been found to potentiate the progression of PCV2 infection to clinical PMWS. 2,25,26,30,41 The authors are not aware of previous reports of bacterial potentiation of PCV2 infection.
Investigations carried out in the field further support the important role of coinfections in the pathogenesis of PCV2-associated disease. A retrospective analysis of PMWS cases in the Midwestern United States revealed that PCV2 alone was found in only 1.9% of all cases investigated. 44 Mycoplasma hyopneumoniae was found in combination with PCV2 in 35.5% of the cases, second only to PRRSV/PCV2 coinfection in 51.9% of the cases. 44
M. hyopneumoniae is associated with porcine enzootic pneumonia, 18,36 a disease characterized by high morbidity but low mortality in affected herds. In the early stages of infection, M. hyopneumoniae colonizes the luminal surface of bronchial and bronchiolar epithelial cells without invading epithelial cells. 5,31,48 Attachment to the cilia leads to progressive loss of cilia, exfoliation of epithelial cells, and accumulation of inflammatory cells in airway lumina and around airways. 9 The hallmark lesion associated with M. hyopneumoniae is hyperplasia of the bronchus-associated lymphoid tissue (BALT); however, M. hyopneumoniae antigen/nucleic acids are not detectable within the hyperplasic BALT. 31,48 Lymphoid hyperplasia of BALT may lead to obliteration of bronchioles and atelectasis of surrounding alveoli. 12 The authors have commonly observed the presence of abundant PCV2 antigen in the hyperplasic BALT of lungs from field cases confirmed to have M. hyopneumoniae/PCV2 coinfection by immunohistochemistry (IHC).
Porcine respiratory disease complex (PRDC) is a condition observed mainly in 8–26-week-old pigs and associated with multiple respiratory pathogens including PRRSV, swine influenza virus (SIV), and M. hyopneumoniae. PRDC is characterized by decreased rate of growth, decreased feed efficiency, anorexia, fever, cough, and dyspnea. Recent field investigations suggest that PCV2 may be playing an important role in some cases of PRDC. 23,28 Studies investigating the relationship of PCV2/M. hyopneumoniae coinfection are lacking in the literature. The objectives of this study were to investigate the interactions between M. hyopneumoniae and PCV2 and to establish a model for studying the pathogenesis of and testing intervention strategies for the control of PCV2-associated PRDC.
Materials and Methods
Animals
Sixty-seven colostrum-fed, crossbred piglets were purchased from a herd with no history of PCV2-associated diseases, free of M. hyopneumoniae and PRRSV based on monthly serologic testing of the breeding herd, and routinely using PPV and SIV vaccination of the breeding animals. The pigs were early-weaned at 12 days of age and transported to the livestock infectious disease isolation facility at Iowa State University, Ames, Iowa. The pigs were randomly assigned to four groups of 16–17 pigs each and separated into four rooms with three pens each and 5–6 pigs per pen.
Experimental design, M. hyopneumoniae, and PCV2 inocula
The experimental design is summarized in Table 1. At 4 weeks of age, a tissue homogenate containing strain 232, a derivative of M. hyopneumoniae strain 11 challenge inoculum (105 color changing units/ml), was administered intra-tracheally to each pig in groups 2 and 3 in a dilution of 1 : 100 in 10 ml Friis medium. 46 Before inoculation, the Friis medium and M. hyopneumoniae inoculum were tested and confirmed negative for the presence of PCV2-, PCV1-, and PPV-specific nucleic acids by polymerase chain reaction (PCR). 13,27
Experimental design.
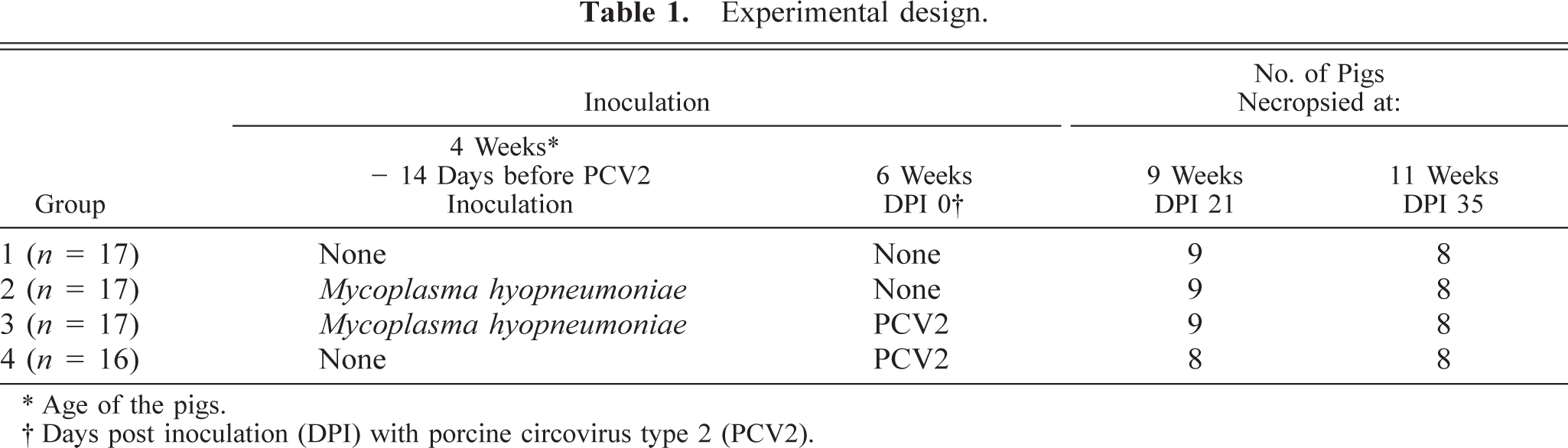
∗ Age of the pigs.
† Days post inoculation (DPI) with porcine circovirus type 2 (PCV2).
PCV2 isolate ISU-40895 was obtained through direct transfection of PK-15 cells with an infectious clone of PCV2. 14 Passage four of the virus at a titer of 104.8 50% tissue culture infective dose (TCID50) was used for inoculation of the pigs. Each pig in groups 3 and 4 received 6 ml of the PCV2 inoculum intranasally at 6 weeks of age. In addition, each pig in groups 1 and 2 was sham inoculated at 6 weeks of age by administration of 6 ml PK15 cell supernatant intranasally.
Clinical evaluation
After PCV2 challenge, the pigs were monitored daily and scored for severity of clinical respiratory disease ranging from 0 to 6 (0 = normal; 1 = mild dyspnea or tachypnea or both when stressed; 2 = mild dyspnea or tachypnea or both when at rest; 3 = moderate dyspnea or tachypnea or both when stressed; 4 = moderate dyspnea or tachypnea or both when at rest; 5 = severe dyspnea or tachypnea or both when stressed; 6 = severe dyspnea or tachypnea or both when at rest). 19 In addition, pigs were evaluated daily for clinical signs including sneezing (score range from 0 [no sneezing] to 3 [severe persistent sneezing]), coughing (score range from 0 [no coughing] to 3 [severe persistent coughing]), and jaundice (score range from 0 [normal] to 3 [severe icterus]). Rectal temperatures, wasting, and behavioral changes such as lethargy were recorded daily. The pigs were weighed on the day of arrival, −14 days before PCV2 inoculation, at the day of PCV2 inoculation, and at 7, 14, 21, and 35 days postinoculation (DPI).
Serology
Blood samples were collected at −24 and −14 days before, on the day of PCV2 inoculation, and at 7, 14, 21, 28, and 35 DPI. A PCV2 enzyme-linked immunosorbent assay (ELISA) based on the recombinant ORF2 capsid protein of PCV2 was performed on −24, −14, 0, 21, and 35 DPI sera samples. 40 Samples were considered positive if the calculated sample-to-positive (S/P) ratio was 0.2 or greater.
M. hyopneumoniae titers were determined by ELISA on −14, 21, and 35 DPI sera. 8 Known positive and negative sera were included on every plate. Readings more than two standard deviations above the mean value of the negative control were considered positive.
In addition, −14, 0, and 35 DPI sera from eight pigs in each group were tested for the presence of antibodies to PRRSV by PRRSV-ELISA (IDEXX Laboratories, Inc. Westbrook, MA), antibodies to SIV subtypes H1N1 and H3N2 by hemagglutination inhibition (HI) assay (protocol used at the Iowa State University Veterinary Diagnostic Laboratory), and for the presence of antibodies to PPV by HI assay. 38
PCV2 quantification
DNA extraction on sera collected on the day of PCV2 inoculation and at 7, 14, 21, 28, and 35 DPI was performed using the QIAamp® DNA Mini Kit (Qiagen, Valencia, CA). DNA extracts were used for quantification of the amount of PCV2 genomic DNA by real-time PCR. 42
Necropsy and histopathology
Necropsies were performed on half of the pigs in each group at 21 DPI and the remainder at 35 DPI. Lung lesions consistent with mycoplasmal pneumonia (well demarcated, dark red-to-purple consolidated areas in the cranioventral regions) were sketched on a standard diagram and assessed for the proportion of lung surface with lesions using a Zeiss SEM-IPS image analyzing system. 46 In addition, the total amount of macroscopic lung lesions (0–100% of the lung affected by grossly visible pneumonia) was estimated and recorded for each pig. 19 The scoring system is based on the approximate volume that each lung lobe contributes to the entire lung: the right cranial lobe, right middle lobe, cranial part of the left cranial lobe, and the caudal part of the left cranial lobe contribute 10% each to the total lung volume, the accessory lobe contributes 5%, and the right and left caudal lobes contribute 27.5% each. 19 The mean size of several lymph nodes (superficial inguinal, external iliac, mediastinal, tracheobronchial, mesenteric) was estimated (score range from 0 [normal] to 3 [three times the normal size]) and recorded for each pig. 42
Tracheobronchiolar swabs and bronchoalveolar lavage for bacterial and virologic examinations were collected at 21 and 35 DPI. 37 Fifty milliliters of sterile phosphate-buffered saline was used to lavage each lung. Lungs collected at 21 DPI were insufflated with fixative until the natural contour of the lung was reestablished, and then the right bronchus was clamped off and the right lung was submerged in fixative. 19 Sections of lung from each lobe were collected at 35 DPI and submerged in 10% neutral buffered formalin. Sections of lymph nodes (superficial inguinal, mediastinal, tracheobronchial, mesenteric), tonsil, thymus, ileum, kidney, colon, spleen, and liver were collected at necropsy and fixed in 10% neutral buffered formalin and routinely processed for histologic examination. Microscopic lesions were evaluated in a blinded fashion by two veterinary pathologists (T. Opriessnig and P. G. Halbur). Lung sections were scored for presence and severity of type 2 pneumocyte hypertrophy and hyperplasia, alveolar septal infiltration with inflammatory cells, peribronchial lymphoid hyperplasia, amount of alveolar exudate, and amount of inflammation in the lamina propria of bronchi and bronchioles ranging from 0 to 6 (0, normal; 1, mild multifocal; 2, mild diffuse; 3, moderate multi-focal; 4, moderate diffuse; 5, severe multifocal; 6, severe diffuse). Sections of heart, liver, kidney, ileum, and colon were evaluated for the presence of lymphohistiocytic inflammation and scored from 0 (none) to 3 (severe). Lymphoid tissues including lymph nodes, tonsil, and spleen were evaluated for the presence of lymphoid depletion ranging from 0 to 3 (0, normal; 1, mild lymphoid depletion with loss of overall cellularity; 2, moderate lymphoid depletion; 3, severe lymphoid depletion with loss of lymphoid follicle structure) and presence of inflammation ranging from 0 to 3 (0, normal; 1, mild histiocytic-to-granulomatous inflammation; 2, moderate histiocytic-to-granulomatous inflammation; 3, severe histiocytic-to-granulomatous inflammation with replacement of follicles).
In addition, Warthin-Starry (WS) silver staining for the identification of cilia-associated respiratory bacillus was done on lung tissues (three sections for each pig) on all pigs necropsied at 21 DPI (n = 35).
IHC for detection of PCV2 and image analysis for quantification of PCV2-specific staining
Immunohistochemical detection of PCV2-specific antigen was performed on sections of formalin-fixed, paraffin-embedded blocks of selected tissue samples (lungs [three sections], lymph nodes [one section each of superficial inguinal, external iliac, mediastinal, tracheobronchial, and mesenteric], tonsil [one section], thymus [one section] and spleen [one section]) using a streptavidin–biotin–peroxidase complex method (DAKO, Carpinteria, CA). The fixed sections were pretreated with 0.05% protease (Sigma, St. Louis, MO), and a rabbit polyclonal antiserum (identification number ISU-31) was used in a dilution of 1 : 1,000. 50 Assessment of staining for PCV2 antigen was done in a blinded fashion and scores ranged from 0 to 3 (0, negative; 1, less than 10% of the lymphoid follicles have cells with PCV2 antigen staining; 2, 10–50% of the lymphoid follicles contain cells with PCV2 antigen staining; 3, more than 50% of the lymphoid follicles contain cells with PCV2 antigen staining). 42 The mean group score was determined for each tissue and compared among groups.
Image analysis was done on 10 randomly selected fields of tracheobronchial lymph node (one section per pig) labeled with anti–PCV2 antibody by IHC from all singular PCV2–and dual-infected pigs. Images were captured with a Sony DXC-S500 color digital camera (Sony Electronics, Inc., Park Ridge, NJ) mounted on a Zeiss Axiophot microscope (Carl Zeiss, Inc., Thornwood, NY) and processed using KS400 image analysis software (Carl Zeiss). A 20× objective and a 1.25× optivar were used, resulting in a final magnification of 25×. The images were converted from RGB to HLS mode and the red and green color planes added together to isolate stained areas. The areas were then interactively discriminated from the surrounding tissue and measured. The software was calibrated to measure in microns. The total area for each field was 227,319.2 square microns, and the percentage of the mean PCV2-specific stained area compared with the total area was calculated for each pig.
IHC for detection of M. hyopneumoniae
Immunohistochemical detection of M. hyopneumoniae–specific antigen on selected lung tissues (three sections for each pig) was done using the heat-induced epitope retrieval technique. Paraffin-embedded tissue sections were dewaxed and rehydrated, covered with 1 : 10 ethylenediaminetetraacetic acid butter solution, pH 6.0 (Richard Allan Scientific, Kalamazoo, MI), placed in a microwave, and boiled for 5 minutes. After cooling for 20 minutes, the slides were rinsed, and M. hyopneumoniae monoclonal antibody (identification number D79DI–7; Dr. Richard Ross, Iowa State University) was applied in a 1 : 500 dilution for 2 hours at room temperature and further processed by using a labeled streptavidin–biotin detection kit (DAKO). Specificity of the M. hyopneumoniae IHC procedure was evaluated by testing sections from formalin-fixed paraffin-embedded blocks from pigs known positive for other pathogens (SIV, PRRSV, PCV2, Mycoplasma flocculare). Specificity was determined to be 100%. There was no evidence of cross-reaction with any of the pathogens tested. Sensitivity was evaluated by comparing the IHC results with those obtained with M. hyopneumoniae culture, which is considered the gold standard for M. hyopneumoniae detection. The sensitivity of the IHC procedure was determined to be 71% (24/34). Known M. hyopneumoniae–positive tissue sections as well as known M. hyopneumoniae–negative tissue sections were used as controls for each IHC run. Slides were scored ranging from 0 to 3 (0, no signal detectable; 1, weak labeling lining the ciliated epithelium of at least one airway; 2, weak-to-moderate labeling on the surface of a low number of airways; 3, intense labeling on the surface of several airways).
Isolation of M. hyopneumoniae and other bacteria from lungs
Isolation of M. hyopneumoniae was performed on tracheobronchial swabs that were inoculated into Friis medium. 46 Tracheobronchial and lung airway swabs were also used for routine bacterial culture on MacConkey and sheep blood agar plates.
Definition and diagnosis of PMWS in this study
Criteria required for the diagnosis of PMWS in this experiment include weight loss or decreased average daily weight gain compared with negative controls, severe lymphoid depletion and histiocytic inflammation in the majority of lymphoid tissues evaluated, and association of the lymphoid lesions with PCV2 antigen. To evaluate pigs in our experiment, we used a scoring system for each lymphoid tissue ranging from 0 to 9 (lymphoid depletion score 0–3; granulomatous inflammation score 0–3; PCV2-IHC score 0–3). The scores (lesions and PCV2-IHC) of the seven lymphoid tissues ([lymph node pool] × 4, tracheobronchial lymph node, spleen, and tonsil) were added together and divided by 7. The lymph node pool consisted of superficial inguinal, external iliac, mediastinal, and mesenteric lymph nodes. The mean treatment group lymphoid score was calculated and compared between groups. Pigs were grouped into four categories on the basis of overall microscopic lymphoid lesion scores: I (normal; score = 0), II (mild; score = 1–3), III (moderate; score = 4–6), and IV (severe; score = 7–9). A pig was diagnosed with PMWS if it had decreased weight gain or weight loss and if the mean lymphoid microscopic lesion severity score was in category IV.
Statistical analysis
Summary statistics were calculated for each group cross-sectionally to assess the overall quality of the data. Analysis of variance (ANOVA) was used for cross-sectional assessment of the average daily weight gain and nonrepeated continuous measures. The rejection level for the null hypothesis was 0.05 followed by pairwise testing using the Tukey-Kramer adjustment to identify the groups that were different. The chi-square test was used to analyze clinical scores, and daily rectal temperature data were analyzed with multivariate ANOVA. Nonrepeated measures of necropsy and histopathology data were assessed using nonparametric Kruskal-Wallis ANOVA. If a nonparametric ANOVA test was significant (P < 0.05), then Wilcoxon tests were used to assess the differences of pairs of groups. Differences in incidence were evaluated by using Fisher's exact test. Kendall's tau was used for nonparametric correlations.
Results
Clinical disease
Singular M. hyopneumoniae– and dual-infected pigs exhibited respiratory disease that was mild-to-moderate in the singular M. hyopneumoniae–infected pigs and mild-to-severe in the dual-infected pigs and was characterized mainly by increased respiratory rates, lethargy, coughing, and occasionally sneezing. Coughing in dual-infected pigs was significantly (P = 0.01) more severe compared with pigs infected singularly with M. hyopneumoniae. Coughing was not observed in control pigs or singular PCV2–infected pigs. Sneezing was occasionally observed in singular PCV2–infected pigs. Singular M. hyopneumoniae– and dual-infected pigs had significantly (P < 0.05) higher respiratory disease scores compared with all other groups. Dual-infected pigs had significantly (P < 0.05) higher respiratory scores compared with singular M. hyopneumoniae–infected pigs except in the 1st week after PCV2 inoculation, when the two groups were not different from each other. The majority of the dual-infected pigs had rough hair coats and appeared to be falling behind their cohorts in the other treatment groups.
Individual pigs within all groups had periodically elevated temperatures. None of the pigs developed persistent fever. Assessment of the mean rectal temperatures showed no significant (P = 0.06) differences between groups.
There was no difference in mean group weight on the day of M. hyopneumoniae inoculation (P = 0.318) or on the day of PCV2 inoculation (P = 0.207), and there was no difference (P = 0.162) in average daily weight gain in the 2 weeks before PCV2 inoculation. The average daily weight gain in the 1st 3 weeks after PCV2 inoculation was 721.7 ± 16.4 g for the control pigs, 662.2 ± 23.8 g for the singular M. hyopneumoniae–infected pigs, 576.6 ± 26.2 g for the dual-infected pigs, and 678.8 ± 32.5 g for the singular PCV2–infected pigs. During this time period, the average daily weight gain was significantly (P < 0.02) decreased in the dual-infected pigs compared with all other groups. The average daily weight gain from 21 to 35 DPI was not different (P = 0.605) among the groups.
Serology
All pigs were serologically negative for M. hyopneumoniae–specific antibodies before M. hyopneumoniae challenge. Singular M. hyopneumoniae– and dual-infected pigs seroconverted to M. hyopneumoniae between 21 and 35 DPI. At 21 DPI, 12/17 dual-infected pigs were positive, 1/17 dual-infected pigs were in the suspect positive category, and 4/17 dual-infected pigs were negative for M. hyopneumoniae–specific antibodies. There were 5/17 singular M. hyopneumoniae–infected pigs that were positive, 7/17 singular M. hyopneumoniae–infected pigs were suspect, and 5/17 singular M. hyopneumoniae–infected pigs were negative for M. hyopneumoniae–specific antibodies at 21 DPI. By 35 DPI, 8/8 dual-infected and 7/8 singular M. hyopneumoniae–infected pigs had seroconverted to M. hyopneumoniae. Dual-infected pigs had significantly (P < 0.05) higher M. hyopneumoniae S/P ratios compared with singular M. hyopneumoniae–infected pigs at 21 and at 35 DPI. Control and singular PCV2–infected pigs remained negative for M. hyopneumoniae–specific antibodies.
At arrival, the pigs had maternal antibodies to PCV2, which decayed below ELISA cutoff levels (S/P ratio < 0.2) before PCV2 inoculation. Control and singular M. hyopneumoniae–infected pigs remained PCV2 antibody negative. Singular PCV2– and dual-infected pigs seroconverted to PCV2 between 21 and 35 DPI. There were 11/17 dual-infected pigs and 6/16 singular PCV2–infected pigs positive for PCV2-specific antibodies by 21 DPI. By 35 DPI, all singular PCV2– and dual-infected pigs had seroconverted to PCV2. At 21 and at 35 DPI, S/P ratios were not different between the two PCV2-infected groups.
All pigs tested were negative for PRRSV as determined by ELISA at −14, 0, and 35 DPI. Some pigs had low maternal antibody titers to SIV and PPV, which decreased over time as expected in pigs from a vaccinated breeding herd.
PCV2 viremia length and genomic copy numbers
Control and singular M. hyopneumoniae–infected pigs were negative by PCR for the presence of PCV2 nucleic acids in sera throughout the study. At 7 DPI, PCV2 viremia was detected in 11/17 dual-infected pigs and in 4/16 singular PCV2–infected pigs. All dual- and singular PCV2–infected pigs were viremic by 14 DPI. The duration of the PCV2-viremia was based on results from the eight dual-infected and eight singular PCV2–infected pigs that remained in the study for the entire 35 days. PCV2 viremia was determined to be 4.6 ± 0.26 weeks in the dual-infected pigs compared with 3.9 ± 0.40 weeks in the singular PCV2–infected group (P = 0.138). The mean copy number of PCV2 genomic DNA in the dual-infected pigs was significantly higher at 14 (P < 0.01) and 21 (P < 0.05) DPI compared with the singular PCV2–infected pigs (Fig. 1). At 7, 28, and 35 DPI, there was no significant difference in the amount of PCV2 genomic DNA in the sera, although dual-infected pigs had higher values.
Mean group PCV2 genomic copy numbers obtained from singular PCV2 and dual Mycoplasma hyopneumoniae/PCV2–infected pigs. Error bars represent standard errors. ∗ = significant (P < 0.05) differences between groups.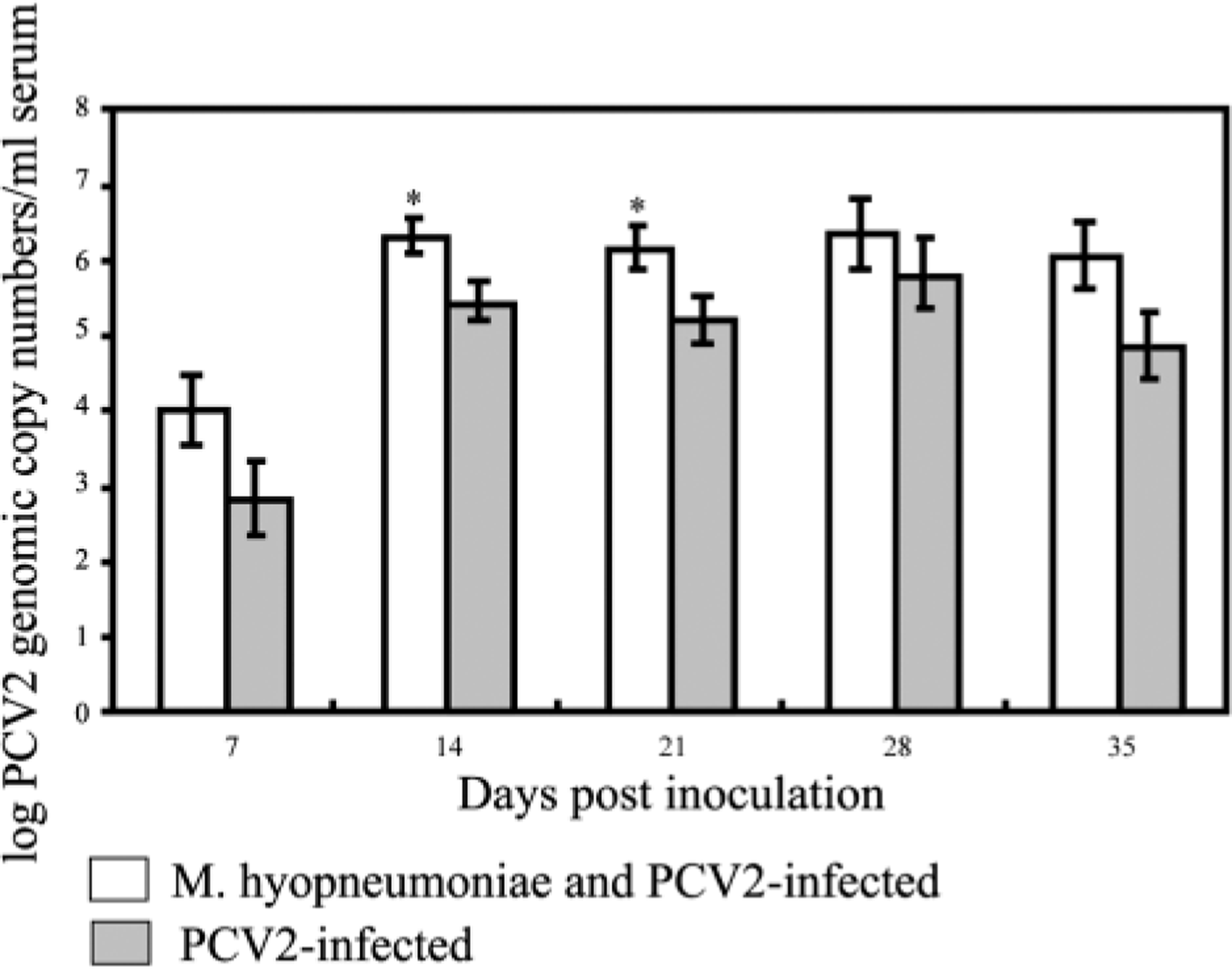
Macroscopic lesions
Macroscopic lesions are summarized in Table 2. Mean percentages of the lung tissue with grossly visible pneumonia in singular M. hyopneumoniae–infected pigs and dual-infected pigs were most severe at 21 DPI and generally appeared to be resolving by 35 DPI. The severity (%) of M. hyopneumoniae–associated lung lesions did not differ between singular M. hyopneumoniae– and dual-infected pigs, although the total gross lung lesion scores were significantly (P < 0.05) more severe in dual-infected pigs at 21 DPI compared with all other groups and at 35 DPI compared with singular PCV2–infected pigs and controls (Figs. 2–5). Lymph nodes in dual- and singular PCV2–infected pigs were two to three times normal size on both necropsy days, whereas control and singular M. hyopneumoniae–infected pigs had essentially normal-sized lymph nodes.
Macroscopic lesions at 21 and 35 days after PCV2 inoculation. Data presented as group mean ± SE.
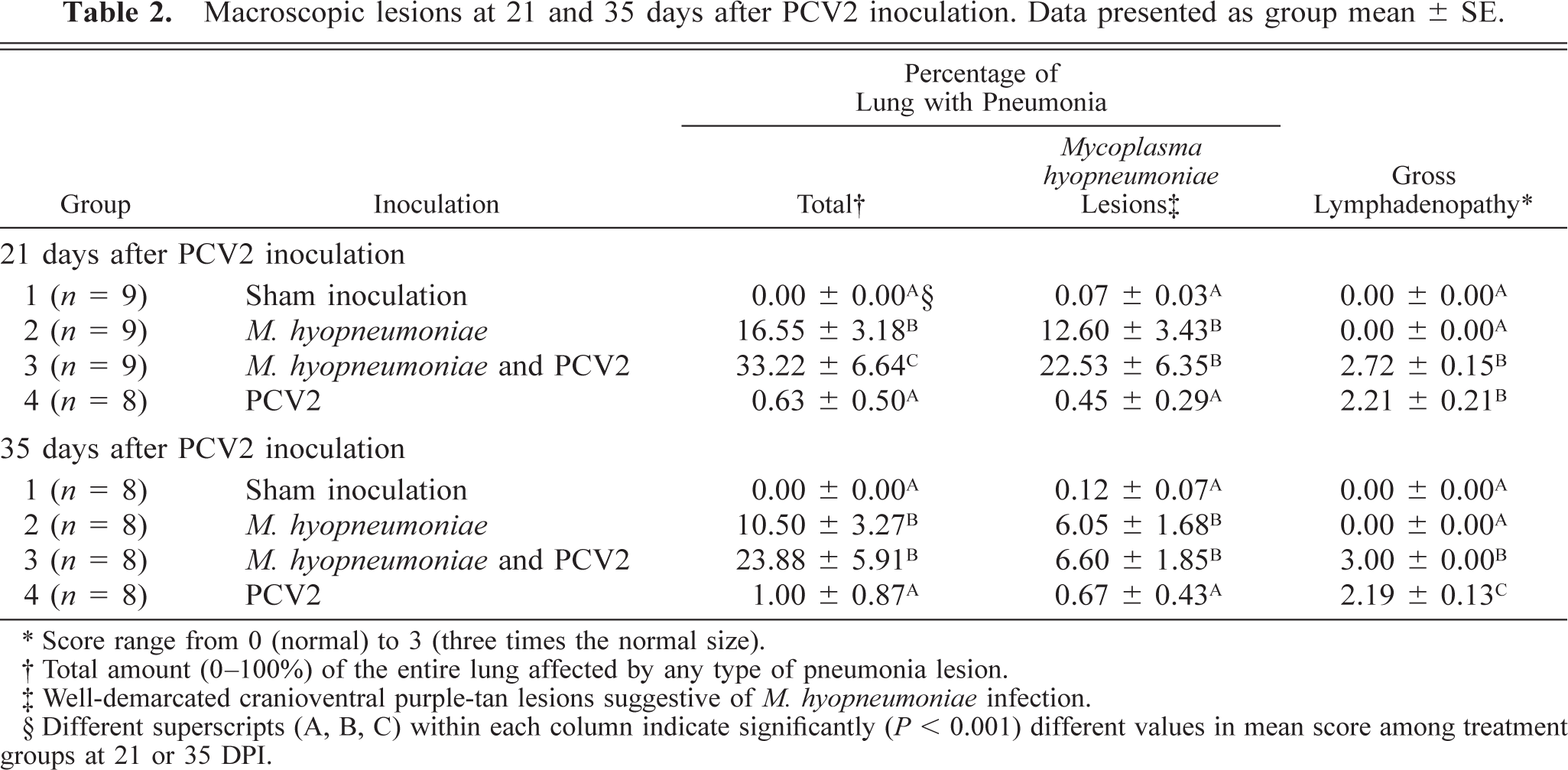
∗ Score range from 0 (normal) to 3 (three times the normal size).
† Total amount (0–100%) of the entire lung affected by any type of pneumonia lesion.
‡ Well-demarcated cranioventral purple-tan lesions suggestive of M. hyopneumoniae infection.
§ Different superscripts (A, B, C) within each column indicate significantly (P < 0.001) different values in mean score among treatment groups at 21 or 35 DPI.
Lung; pig, 35 days after inoculation with PCV2. No gross lesions were observed.
Lung; pig, 35 days after inoculation with Mycoplasma hyopneumoniae. The lung has well-demarcated, dark red areas of pneumonia in the cranioventral region.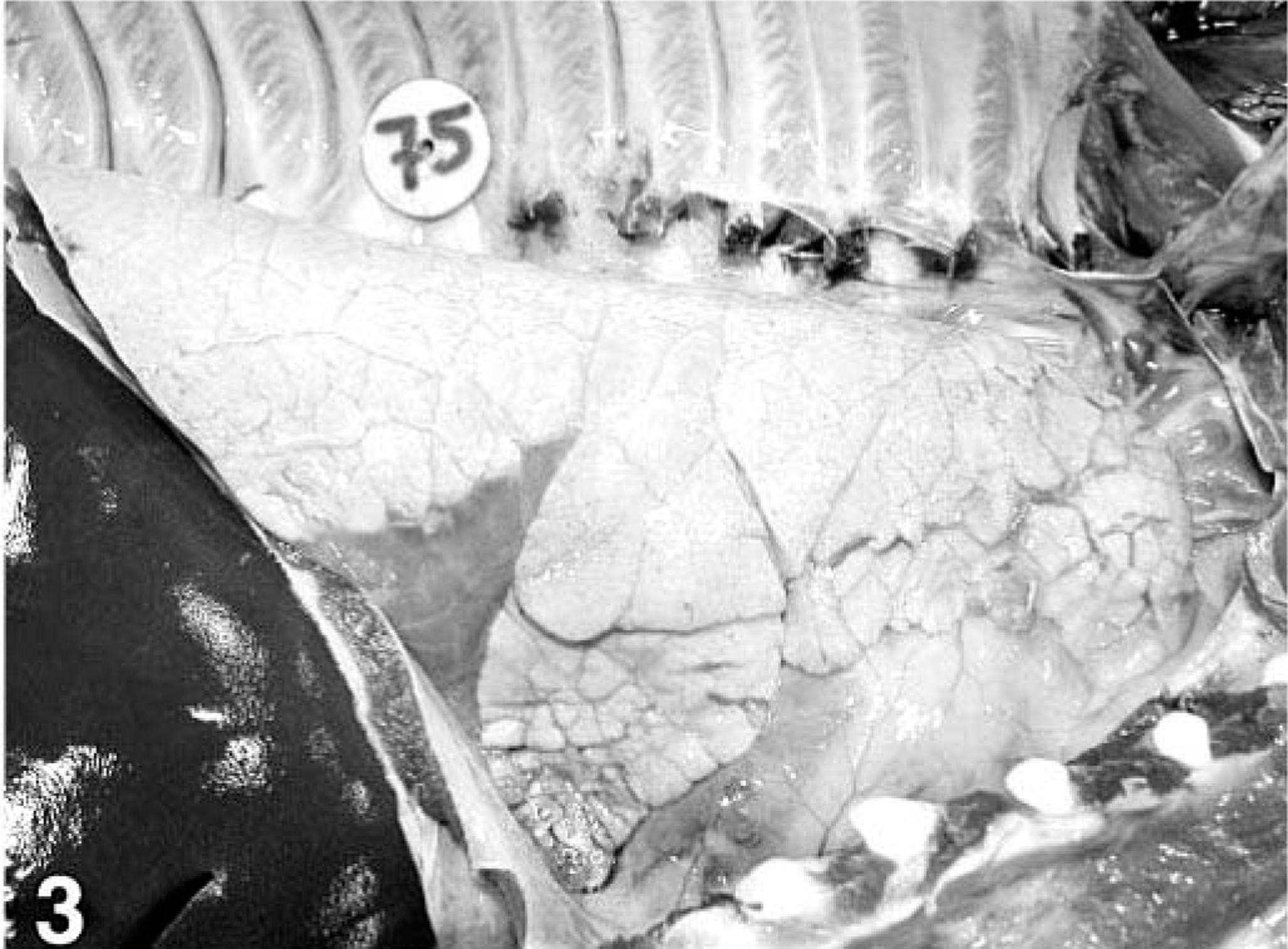
Lung; pig, 35 days after inoculation with Mycoplasma hyopneumoniae and 21 days after inoculation with PCV2. The lung failed to collapse and has a diffuse mottled-tan appearance and severe multifocal dark purple-to-tan consolidation.
Lung; pig, 49 days after inoculation with Mycoplasma hyopneumoniae and 35 days after inoculation with PCV2. The lung is diffusely mottled-tan and has well-demarcated, dark red tan areas of consolidation.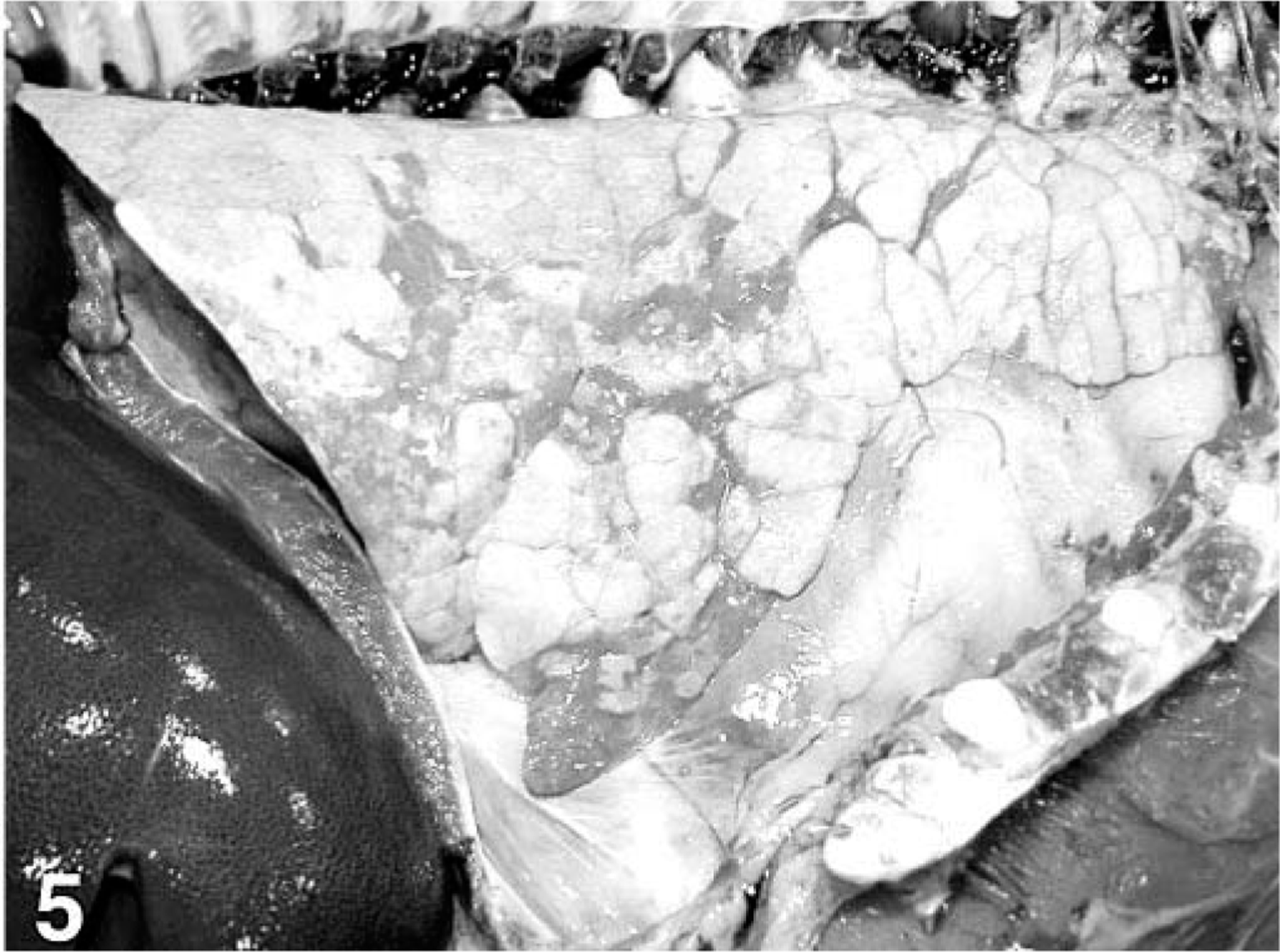
Microscopic lesions
Microscopic lesions are summarized in Tables 3–5. All treatment groups except controls (Fig. 6) had interstitial pneumonia characterized by type 2 pneumocyte hypertrophy and hyperplasia and alveolar wall thickening by macrophages and lymphocytes (Table 3). If present, these lesions were mild in the singular M. hyopneumoniae–infected pigs, mild-to-moderate in the singular PCV2–infected pigs and mild-to-severe in the dual-infected pigs (Fig. 7). Lungs from singular M. hyopneumoniae– and dual-infected pigs also had moderate-to-severe peribronchiolar and perivascular lymphohistiocytic cuffing and nodule formation, moderate-to-severe increase in alveolar exudate and eosinophilic fluid, lymphohistiocytic inflammation in the lamina propria of airways, and mixed inflammation in the lumina of the airways (Figs. 8, 9). There was clumping and loss of cilia, sloughing of epithelial cells into the airway lumina, mild necrosis and ulceration of epithelium over hyperplastic peribronchiolar lymphoid nodules, and infiltration of mononuclear cells in the lamina propria in the M. hyopneumoniae–infected groups. In addition, some PCV2- and dual-infected pigs had mild peribronchial fibroplasia. Multinucleated giant cells were infrequently observed in peribronchial areas of lymphoid hyperplasia.
Microscopic lung lesions in pigs in group 1 (sham inoculated), 2 (singular Mycoplasma hyopneumoniae infected), 3 (M. hyopneumoniae and PCV2 dual infected), and 4 (singular PCV2 infected) at 21 and 35 days after PCV2 inoculation. Data presented as incidence (group mean [mean severity score for affected animals]).
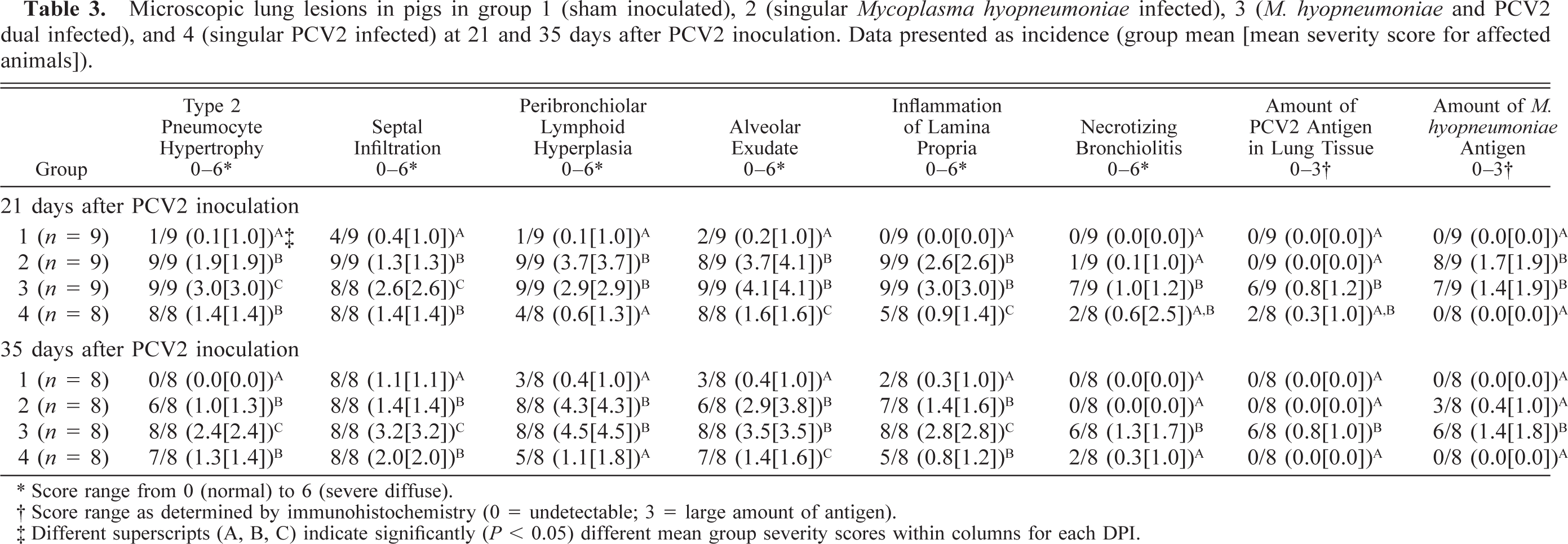
∗ Score range from 0 (normal) to ± (severe diffuse).
† Score range as determined by immunohistochemistry (0 = undetectable; 3 = large amount of antigen).
‡ Different superscripts (A, B, C) indicate significantly (P < 0.05) different mean group severity scores within columns for each DPI.
Lung; control pig. Normal. HE. Bar = 250 µm.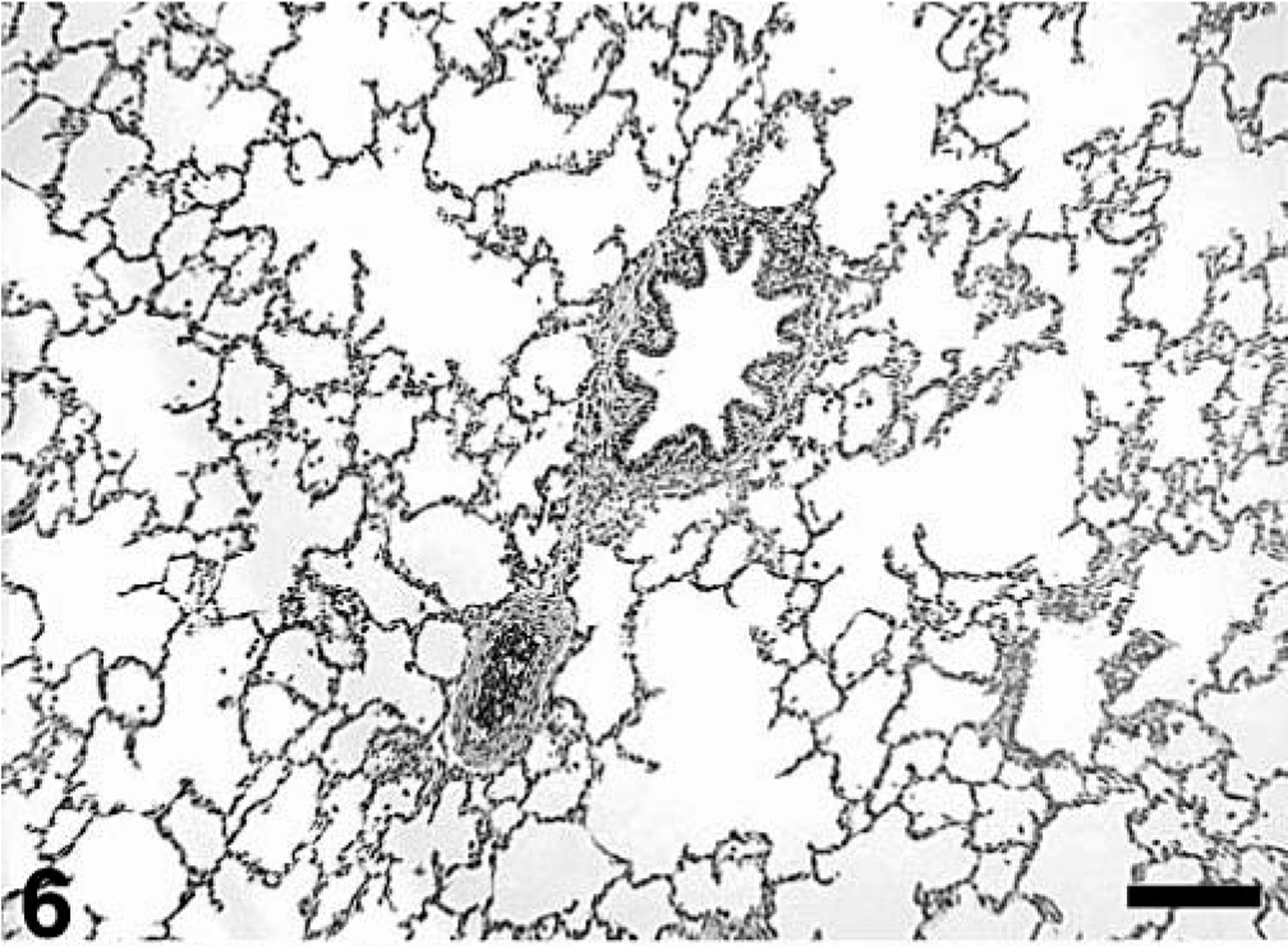
Lung; pig, 35 days after inoculation with Mycoplasma hyopneumoniae and 21 days after inoculation with PCV2. There is thickening of the alveolar septa by marked type 2 pneumocyte hypertrophy and hyperplasia and infiltration of alveolar space by moderate numbers of macrophages and low numbers of neutrophils. HE. Bar = 75 µm. Inset: Intense nuclear and cytoplasmic staining for PCV2 antigen in a macrophage-like cell. Immunohistochemical staining with anti–PCV2 polyclonal antibody. Streptavidin–biotin–peroxidase complex method counterstained with hematoxylin. Bar = 20 µm.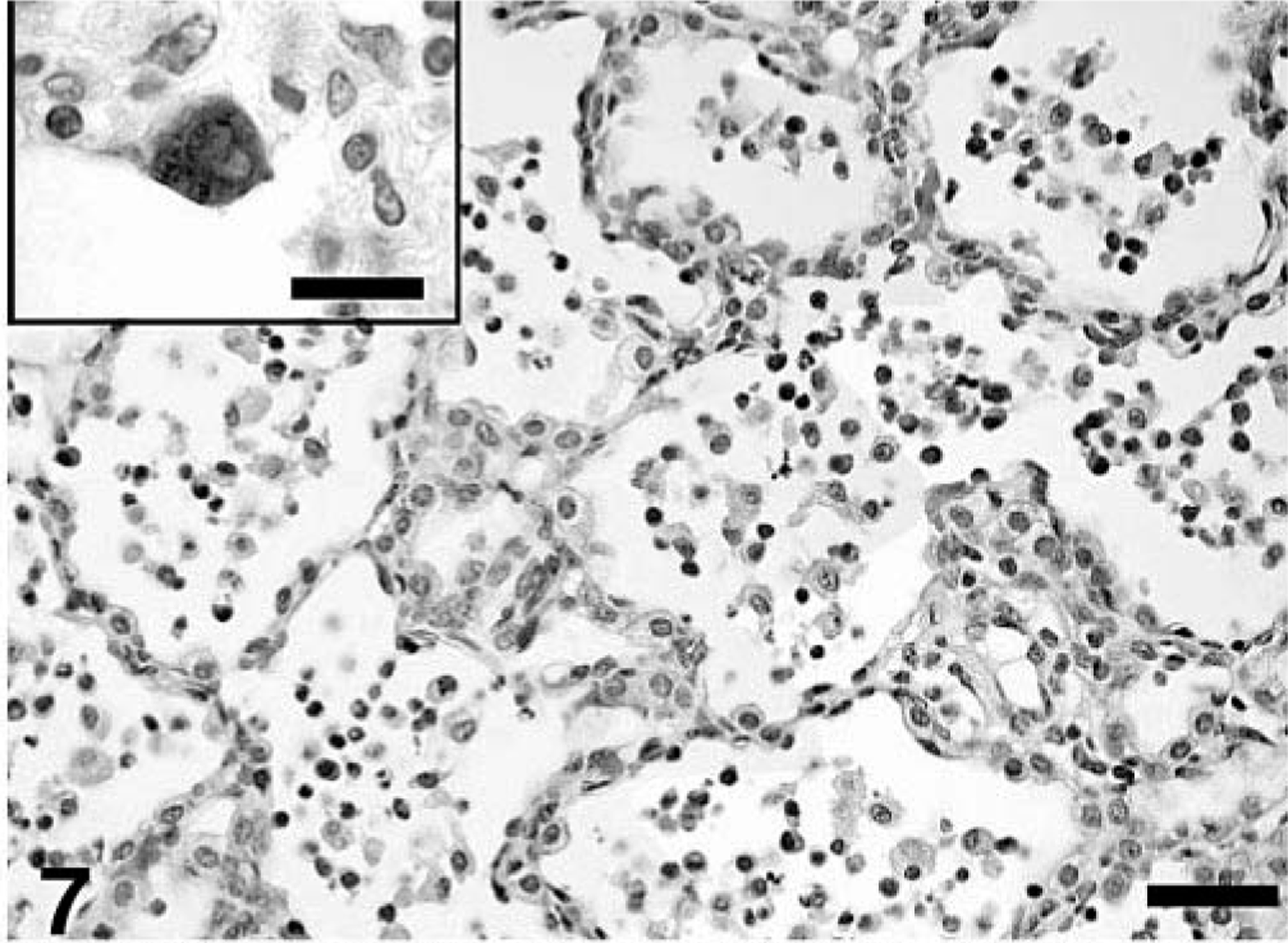
Lung; pig, 35 days after inoculation with Mycoplasma hyopneumoniae. There is peribronchiolar lymphoid hyperplasia typical of M. hyopneumoniae infection. HE. Bar = 250 µm.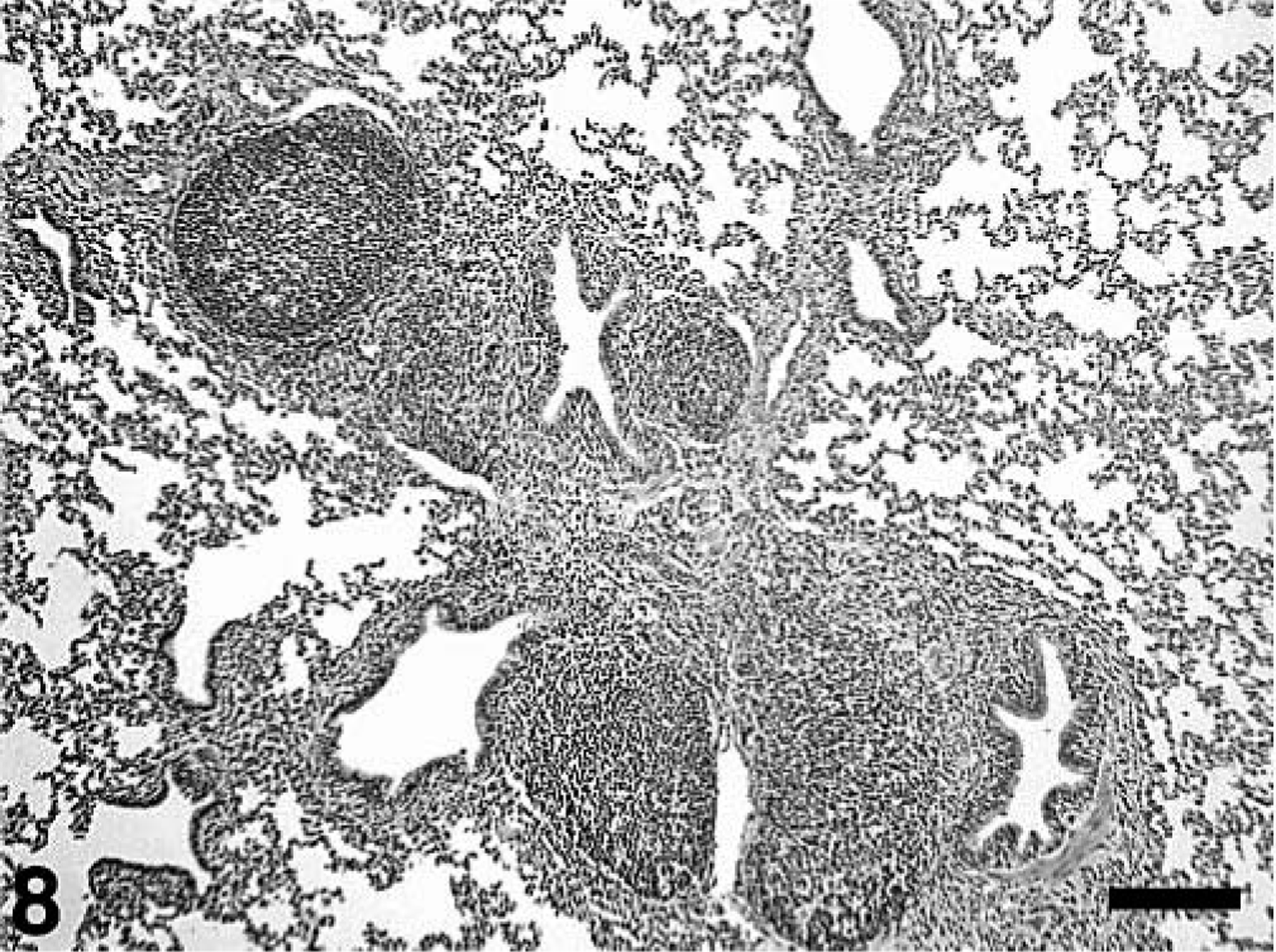
Lung; pig, 35 days after inoculation with Mycoplasma hyopneumoniae and 21 days after inoculation with PCV2. There is peribronchiolar lymphoid hyperplasia typical of M. hyopneumoniae infection. The interstitial pneumonia, peribronchial fibroplasia, and infiltration of the lamina propria with macrophages and lymphocytes (arrows) is typical of PCV2 infection. HE. Bar = 250 µm.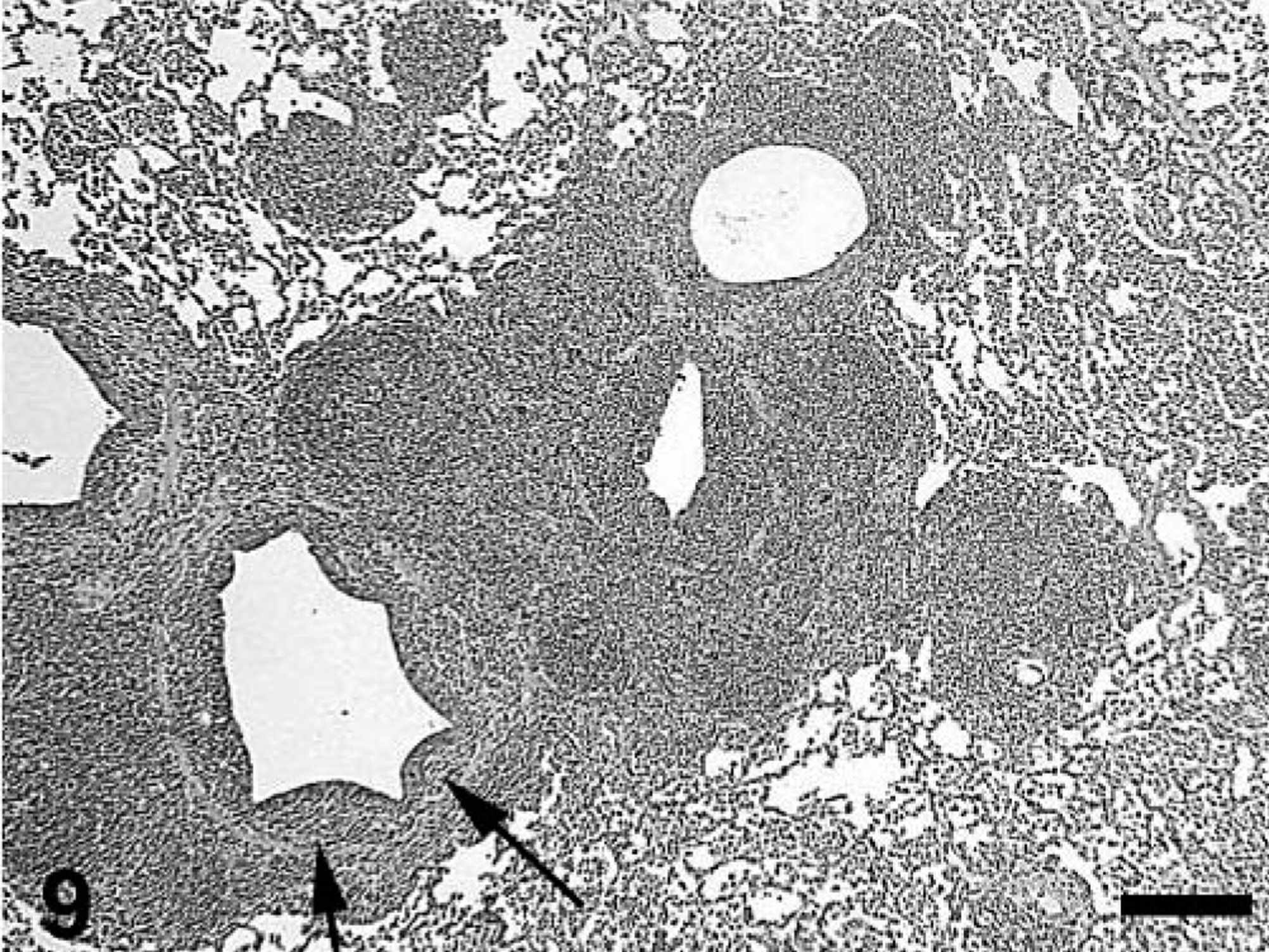
Lesions in the lymphoid tissues are summarized in Tables 4 and 5. Control and singular M. hyopneumoniae–infected pigs had essentially normal lymphoid tissues. PCV2-associated lesions in lymphoid tissues were characterized by mild-to-severe lymphoid depletion and mild-to-severe histiocytic-to-granulomatous inflammation with low-to-moderate numbers of multinucleated giant cells. Low-to-moderate numbers of macrophages and multinucleated giant cells contained several, variable-sized, spherical, basophilic intracytoplasmic inclusion bodies. If present (31/34 singular PCV2– and dual-infected pigs), the lymphoid lesions were restricted to one or two of the seven lymphoid tissues examined in 16/31 (51.8%) of the pigs. Two of the singular PCV2–infected pigs had severe lymphoid depletion and moderate histiocytic inflammation; however, this was restricted to individual lymph nodes, was not associated with high amounts of PCV2 antigen (PCV2 IHC score 0 and 1, respectively), and was not observed in tonsil or spleen. The individual scores for overall severity of microscopic lymphoid lesions associated with PCV2 antigen in dual and singular PCV2–infected pigs are summarized in Fig. 10. The mean group severity score for lymphoid lesions was 4.8 ± 0.8 for the dual-infected pigs and 2.1 ± 0.6 for the singular PCV2–infected pigs at 21 DPI, 3.1 ± 0.7 for the dual-infected pigs and 1.8 ± 0.5 for the singular PCV2–infected pigs at 35 DPI. Four dual-infected pigs were in category IV (three were necropsied at 21 DPI, and one was necropsied at 35 DPI) and had decreased rate of weight gain and moderate respiratory disease and fulfilled our definition for PMWS.
Microscopic lesions (lymphoid depletion and inflammation) and amount of PCV2-specific antigen demonstrated by immunohistochemistry (IHC) in lymph nodes at 21 and 35 days after PCV2 inoculation. Data presented as incidence (mean group severity score [mean severity for affected animals]).
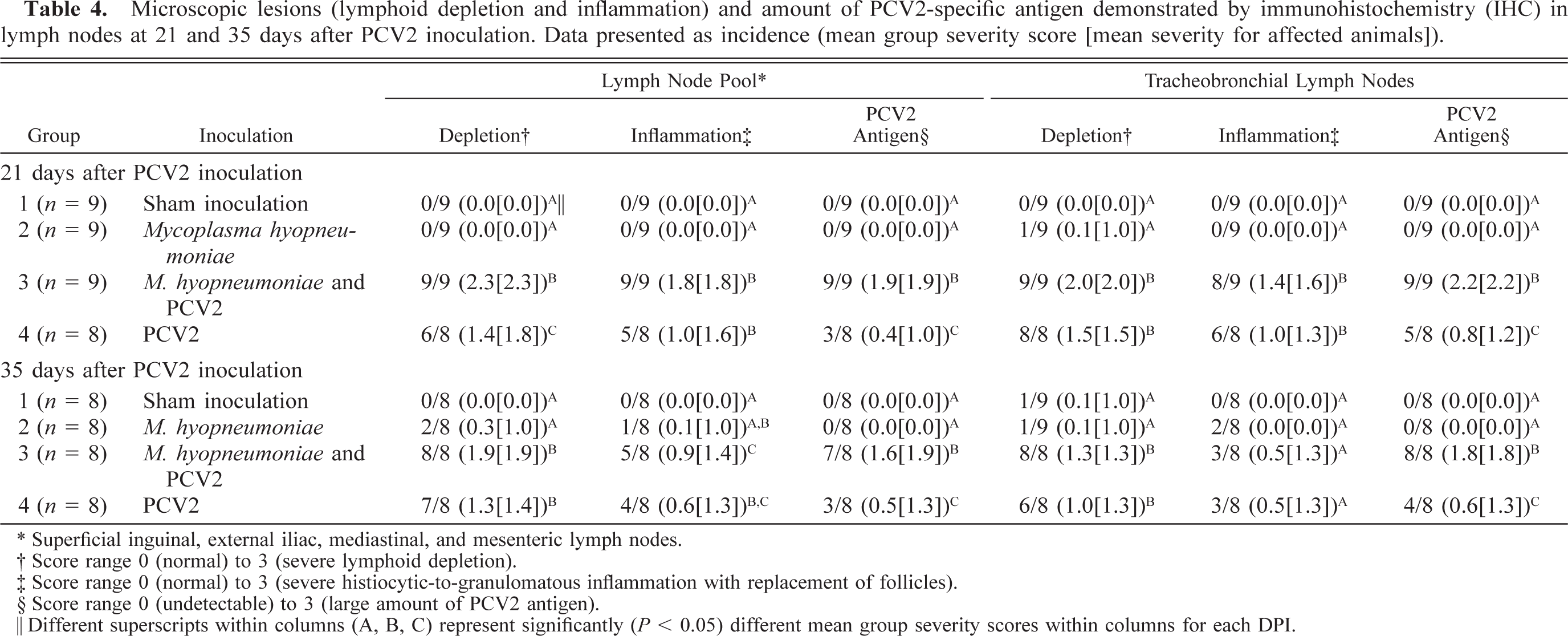
∗ Superficial inguinal, external iliac, mediastinal, and mesenteric lymph nodes.
† Score range 0 (normal) to 3 (severe lymphoid depletion).
‡ Score range 0 (normal) to 3 (severe histiocytic-to-granulomatous inflammation with replacement of follicles).
§ Score range 0 (undetectable) to 3 (large amount of PCV2 antigen).
‖ Different superscripts within columns (A, B, C) represent significantly (P < 0.05) different mean group severity scores within columns for each DPI.
Microscopic lesions (lymphoid depletion and inflammation) and amount of PCV2-specific antigen demonstrated by immunohistochemistry (IHC) in tonsil and spleen at 21 and 35 days after PCV2 inoculation. Data presented as incidence (mean group severity score [mean severity for affected animals]).
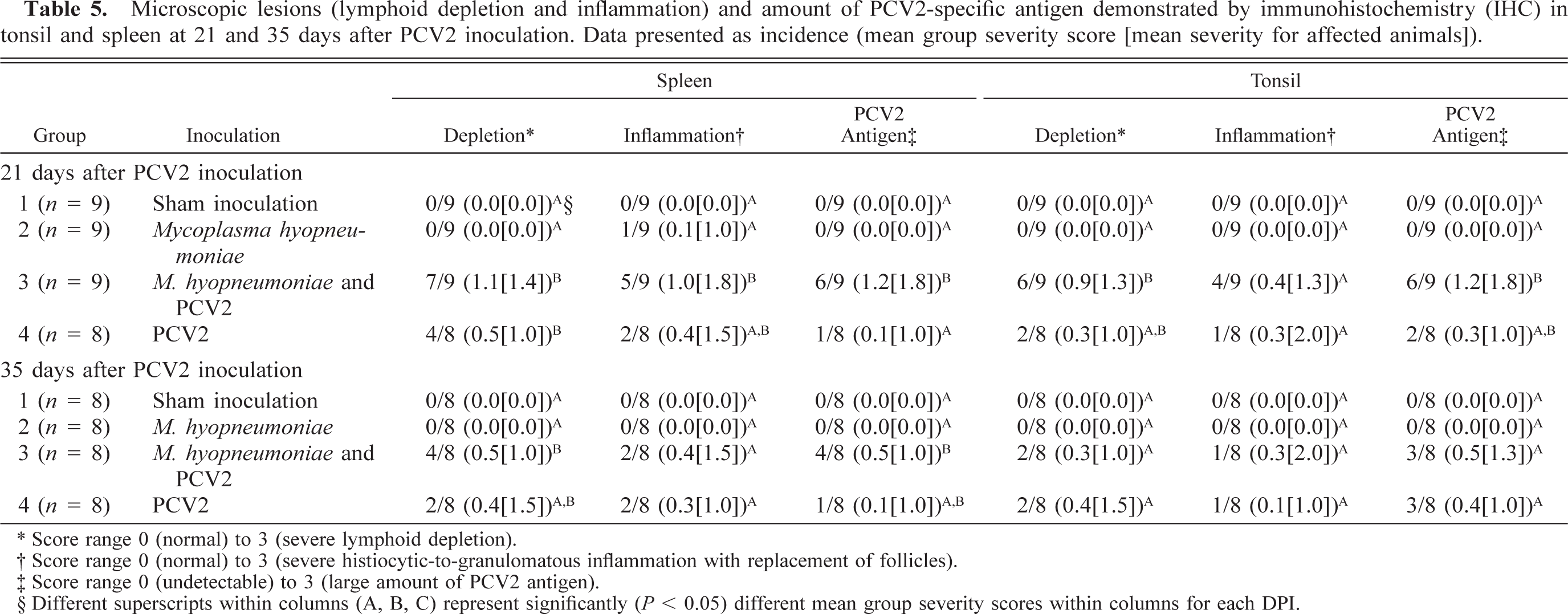
∗ Score range 0 (normal) to 3 (severe lymphoid depletion).
† Score range 0 (normal) to 3 (severe histiocytic-to-granulomatous inflammation with replacement of follicles).
‡ Score range 0 (undetectable) to 3 (large amount of PCV2 antigen).
§ Different superscripts within columns (A, B, C) represent significantly (P < 0.05) different mean group severity scores within columns for each DPI.
Distribution of individual pig lymphoid lesion (lymphoid depletion and inflammation and PCV2 antigen) scores in pigs in singular PCV2–infected (n = 16) and dual Mycoplasma hyopneumoniae/PCV2–infected (n = 17) groups. Score ranges from 0 to 9.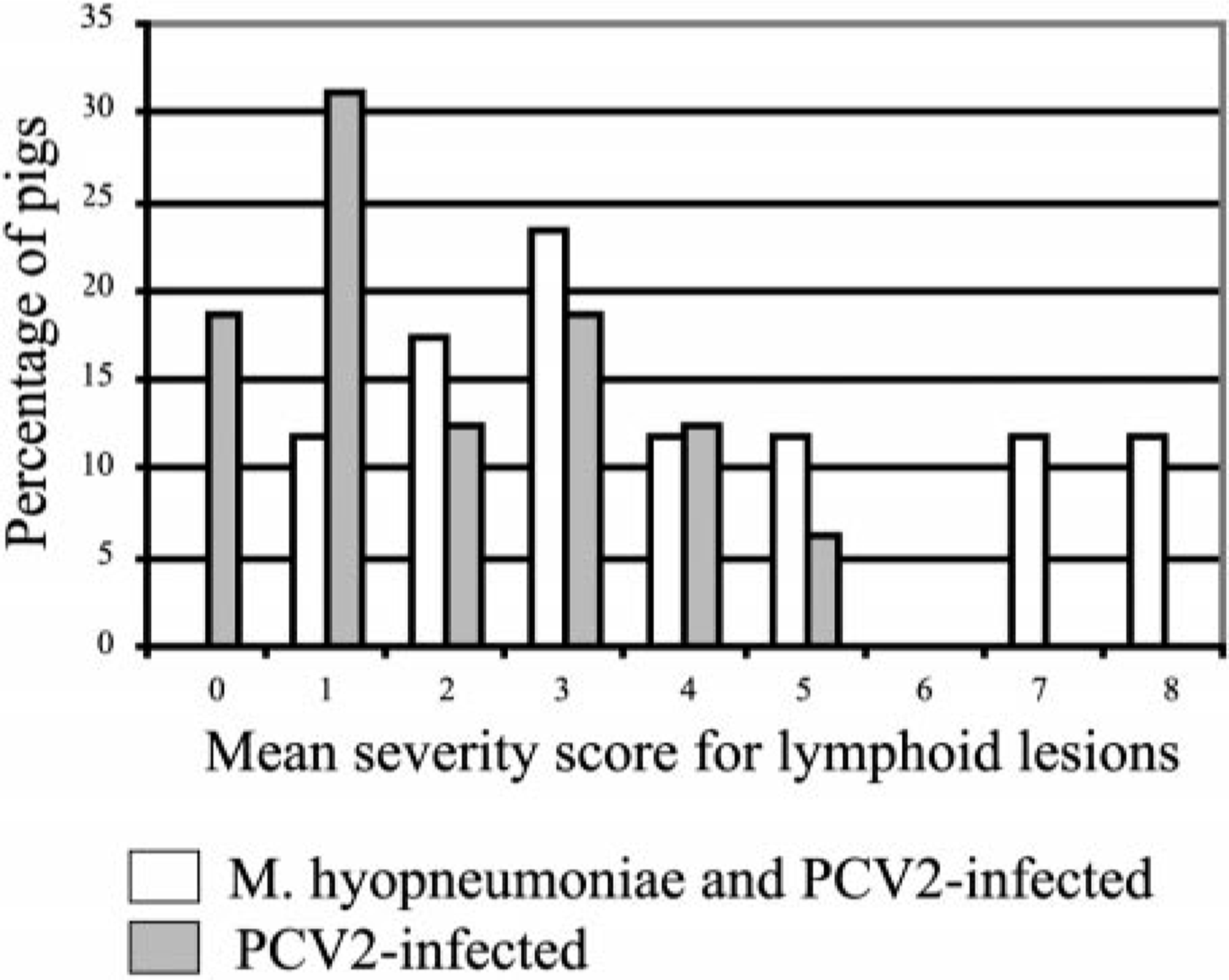
Individual pigs in all groups had mild-to-moderate lymphohistiocytic myocarditis, mild-to-moderate lymphohistiocytic hepatitis, mild-to-moderate lymphohistiocytic interstitial nephritis, and mild lymphohistiocytic enteritis. Dual-infected pigs had significantly (P < 0.05) more severe lymphohistiocytic myocarditis compared with control pigs and M. hyopneumoniae–infected pigs but not compared with the singular PCV2–infected group at DPI 21 and 35. The four pigs that were diagnosed with PMWS had moderate lymphohistiocytic myocarditis and hepatitis, moderate lymphohistiocytic-to-granulomatous nephritis, and mild-to-moderate histiocytic-to-granulomatous inflammation of Peyer's patches.
IHC, image analysis, and M. hyopneumoniae isolation
The estimated mean amount of PCV2 antigen in lung and lymphoid tissues by groups as determined by IHC is summarized in Tables 3–5. PCV2 antigen in lung tissue was most often associated with cells that had morphologic features consistent with macrophages or dendritic cells in areas of M. hyopneumoniae–induced peribronchial lymphoid hyperplasia (Fig. 11), within the expanded lamina propria beneath areas of M. hyopneumoniae colonization and damage to epithelial cells (Fig. 12), within alveolar macrophages, and in cells morphologically consistent with septal fibroblasts. In these cells, abundant staining was observed mainly in the cytoplasm but also in few nuclei (Fig. 7 inset). Compared with singular PCV2–infected pigs, dual-infected pigs had significantly (P < 0.05) more PCV2 antigen in lymph nodes at 21 (Fig. 13) and 35 DPI, in spleen at 21 DPI, and in the lung at 35 DPI. Two dual-infected pigs and one PCV2–infected pig had a few PCV2-positive cells in the thymus (data not shown).
Lung; pig, 35 days after inoculation with Mycoplasma hyopneumoniae and 21 days after inoculation with PCV2. PCV2 antigen–positive cells (arrows) are associated with the peribronchial lymphoid hyperplasia. Immunohistochemical staining with anti–PCV2 polyclonal antibody. Streptavidin–biotin–peroxidase complex method counter-stained with hematoxylin. Bar = 120 µm. Inset: Detail of the area of follicular lymphoid hyperplasia with several PCV2 antigen–labeled macrophage- and dendritic-like cells. Bar = 35 µm.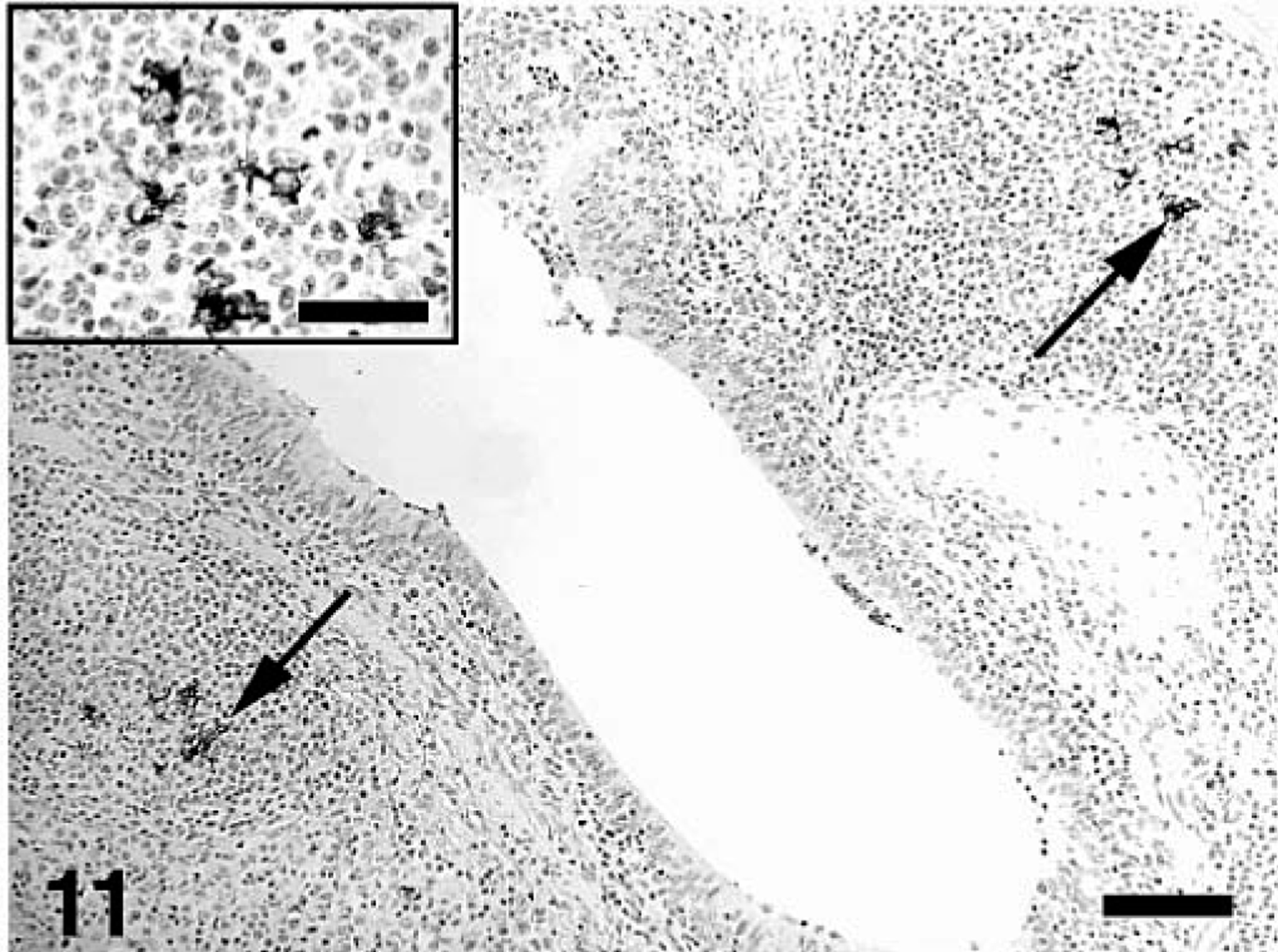
Lung; pig, 35 days after inoculation with Mycoplasma hyopneumoniae and 21 days after inoculation with PCV2. PCV2 antigen–positive cells (arrows) in the lamina propria and submucosa of a bronchus. Immunohistochemical staining with anti–PCV2 polyclonal antibody. Streptavidin–biotin–peroxidase complex method counterstained with hematoxylin. Bar = 80 µm. Inset: Detail of the area with macrophage-like cells with numerous variable-sized PCV2 antigen–positive intracytoplasmic spherical bodies as well as nuclear staining. Bar = 30 µm.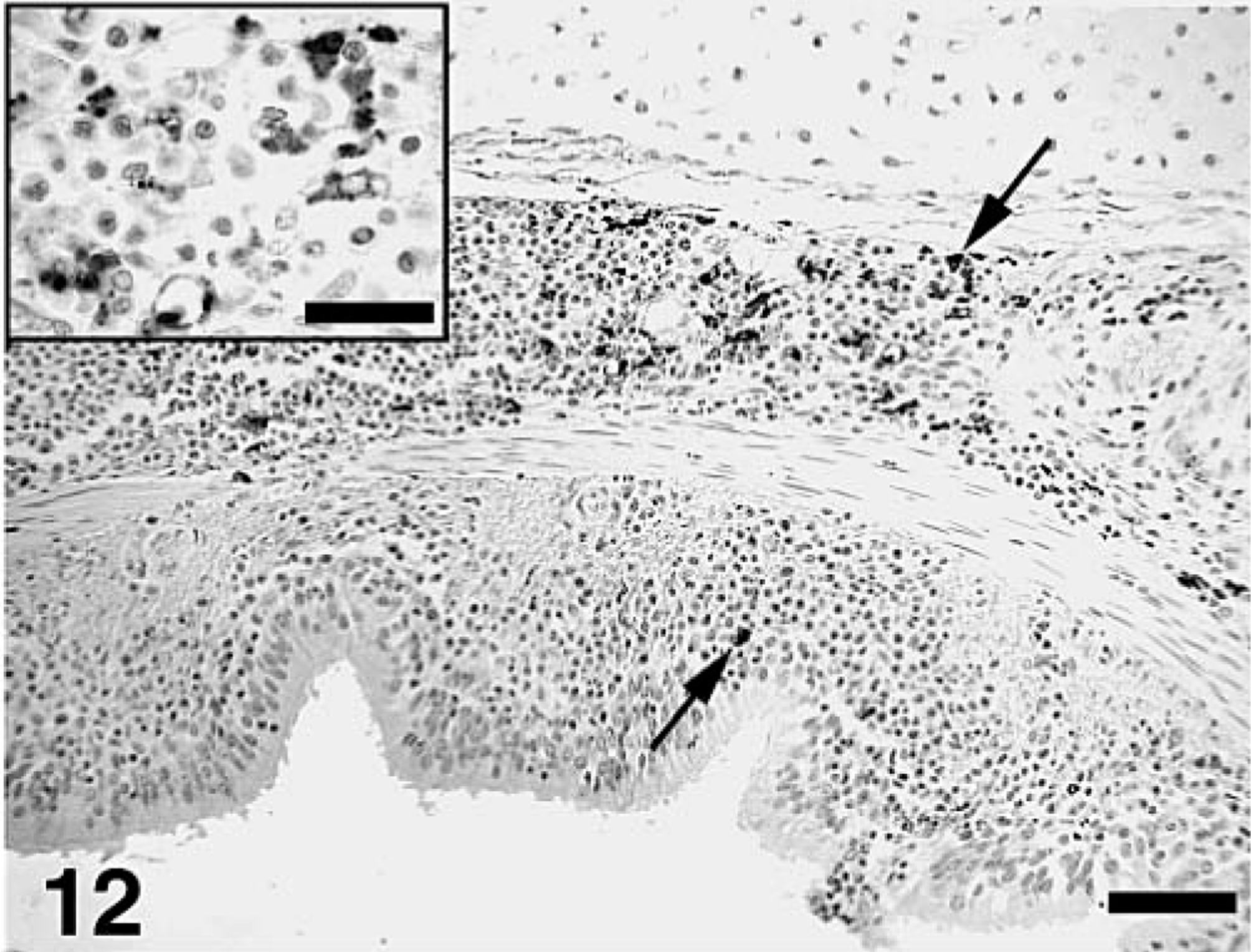
Tracheobronchial lymph node; pig. 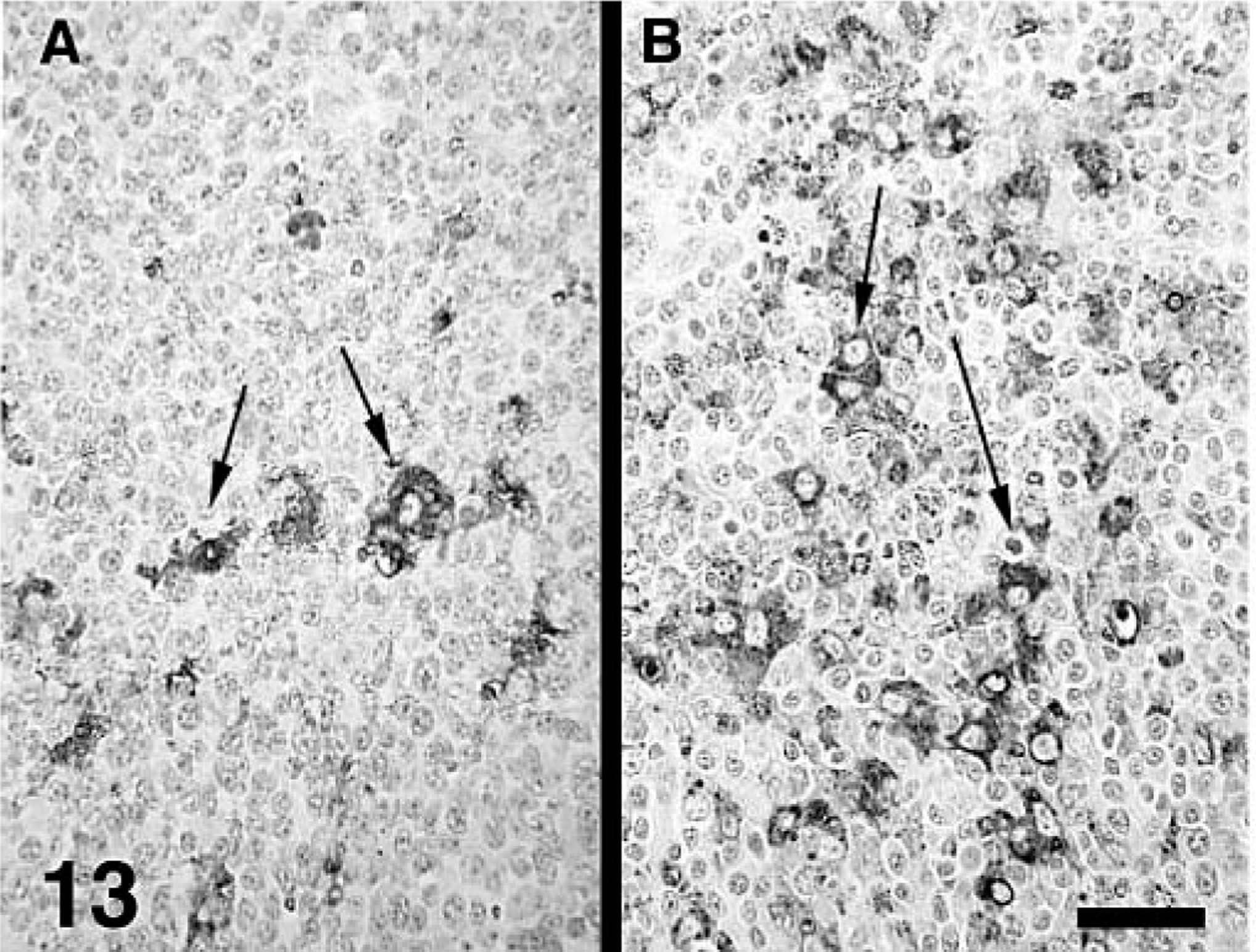
Image analysis on tracheobronchial lymph nodes demonstrated a significant (P < 0.05) difference between the singular PCV2– and the dual-infected pigs at both 21 and 35 DPI. Because of their distribution, the data are presented as medians with a 2.5% to 97.5% interval that contains 95% of the data. There was a median stained area of 1,173.1 (0.0; 15,883.2) square microns (0.516% of the total area) in the dual-infected pigs and 20.9 (0.0; 732.8) square microns (0.009% of the total area) in the singular PCV2–infected pigs at 21 DPI. There was a median stained area of 238.6 (2.1; 2,241.4) square microns (0.105% of the total area) in the dual-infected pigs and 5.5 (0.0; 174.2) square microns (0.002% of the total area) in the singular PCV2–infected pigs at 35 DPI. The correlation between IHC scoring and image analysis data was high (0.78) and statistically significant (P < 0.0001). Those pigs with IHC sores of 0 (n = 13), 1 (n = 10), 2 (n = 7), and 3 (n = 3) had mean stained areas of 11.8 ± 7.2 (0.01% of the total area), 122.0 ± 69.5 (0.05% of the total area), 1,207.3 ± 342.5 (0.53% of the total area), and 9,638.1 ± 3,368.3 (4.24% of the total area) square microns, respectively.
The IHC results for M. hyopneumoniae are summarized in Table 3. M. hyopneumoniae antigen was associated with the surface of epithelial cells of bronchi and bronchioli (Fig. 14). At 35 DPI, there was significantly (P < 0.05) more M. hyopneumoniae antigen in dual-infected pigs compared with singular M. hyopneumoniae–infected pigs, but there was no difference (P = 0.315) in incidence between the two groups. M. hyopneumoniae was isolated from 0/17 control pigs, from 17/17 singular M. hyopneumoniae–infected pigs, and from 17/17 dual-infected pigs. M. hyopneumoniae was also isolated from 1/16 singular PCV2–infected pigs. This pig was necropsied at 35 DPI and had no lesions characteristic of M. hyopneumoniae infection (peribronchial lymphoid hyperplasia score = 0). IHC staining for M. hyopneumoniae on this pig was negative, and the pig was seronegative for antibodies to M. hyopneumoniae at necropsy. Therefore, this likely is a case of contamination at some point in sample handling and processing.
Lung; pig, 35 days after inoculation with Mycoplasma hyopneumoniae and 21 days after inoculation with PCV2. M. hyopneumoniae–specific staining (arrow) at the luminal surface of the bronchus epithelial cells. Immunohistochemical staining with anti–M. hyopneumoniae monoclonal antibody. Streptavidin–biotin complex method counterstained with hematoxylin. Bar = 200 µm. Inset: The epithelial cell layer has intense staining for M. hyopneumoniae antigen on the surface. Bar = 30 µm.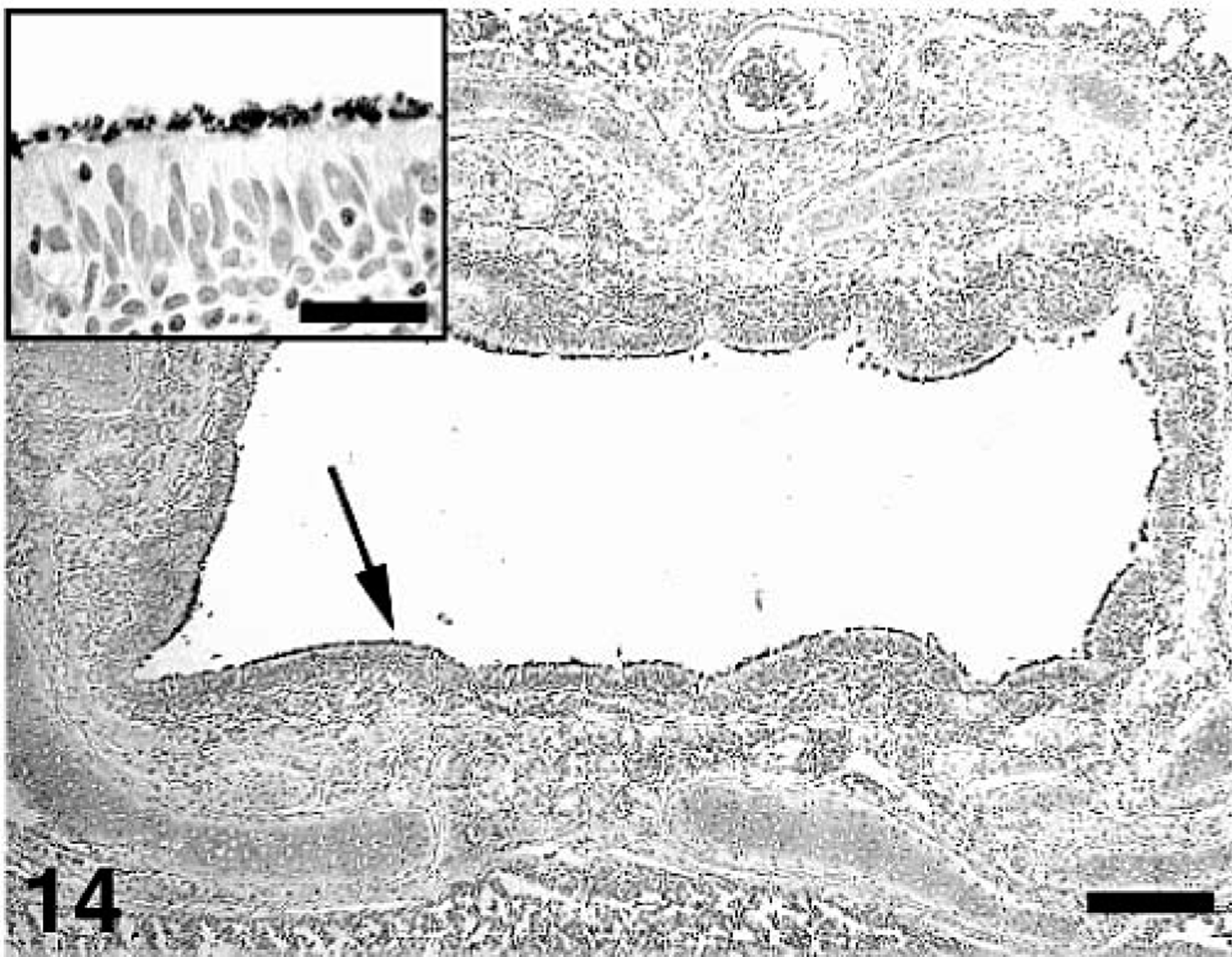
Bacteriology
Cultures of tracheobronchial and lung swabs of all pigs were negative for common swine respiratory pathogens including Pasteurella multocida, Bordetella bronchiseptica, Haemophilus parasuis, Streptococcus suis, Salmonella sp., Actinobacillus sp., and others.
Organisms morphologically consistent with cilia-associated respiratory bacillus were not detected in any of the lung sections of the 35 pigs tested as determined by WS silver staining.
Discussion
PCV2 is now considered an important contributor to the PRDC. 23,28,44 The overall goal of this study was to investigate the potential interaction between PCV2 and M. hyopneumoniae. We demonstrated that dual infection of pigs with M. hyopneumoniae and PCV2 results in increased severity of pneumonia and increased incidence of PMWS. Four of 17 (23.5%) of the dual-infected pigs had clinical signs and microscopic lesions consistent with PMWS, whereas none of the singular PCV2–infected pigs developed PMWS. This is to our knowledge the first report of bacterial potentiation of PCV2 infection resulting in PMWS in experimentally infected pigs. As in the results reported here, M. hyopneumoniae significantly prolonged and increased the severity of PRRSV-induced pneumonia; 52,55 however, M. hyopneumoniae had no effect on concurrent SIV infection. 53 Neither those studies nor the work described here demonstrated a virus-dependent enhancement of mycoplasmal pneumonia.
M. hyopneumoniae infection increased the amount of PCV2 genomic copy numbers in the sera, prolonged the presence of PCV2 antigen in lymphoid and lung tissues, and increased the amount of PCV2 antigen in lung tissues in dual-infected pigs. PCV2-associated lesions and the amount of PCV2 antigen associated with the lesions were enhanced by concurrent infection by viruses (PRRSV, PPV) that replicate in cells of the monocyte/macrophage lineage. 2,3,24,25,26,30,41,47 Two recent in vitro studies reported that monocytes, macrophages, and dendritic cells were successfully infected with PCV2; however, PCV2 was not found to replicate in vitro in these cell types under culture conditions. 17,58 PCV2 replication depends on the availability of host cell polymerase, which is active in regularly dividing cells. 57 Although tissue macrophages are capable of cell division, usually less than 5% of the resident macrophage population arise de novo. 34 Unlike PCV2, PPV and PRRSV both readily replicate in the monocyte lineage in vitro. 11,22 Because both PPV and PRRSV have been shown to enhance PCV2 replication in vivo, it is possible that these viruses activate the macrophages in a way that supports PCV2 replication.
M. hyopneumoniae infection induces an infiltration of inflammatory cells in BALT areas, 48 and PCV2 antigen in dual-infected pigs was demonstrated mainly in the cytoplasm and also in a few nuclei of cells with morphologic features consistent with macrophages, dentritic-like cells, and fibroblasts in areas associated with M. hyopneumoniae colonization. In situ hybridization and IHC have failed to demonstrate M. hyopneumoniae in these cells; 31,48 however, the immune response induced by M. hyopneumoniae is complex with reports of both immunostimulation and immunosuppression, 29,35,39 nonspecific mitogenic effects on swine lymphocytes, 39 immunogenic mycoplasmal proteins, 51 or cytokines secreted by activated macrophages. 6,7,54,56 Hence, the mitogenic activity of M. hyopneumoniae could upregulate the macrophage proliferation in the lung, thereby supporting PCV2 replication at the site of infection.
It has been shown that PCV2 is not degraded on uptake by bone marrow– or blood monocyte–derived dendritic cells in vitro, suggesting that these cells provide potentially important vehicle for transport of the virus throughout the body and tissues. 58 Macrophages and dendritic cells are upregulated by M. hyopneumoniae, 48 and thus M. hyopneumoniae coinfection might promote systemic distribution of PCV2. This could explain how localized M. hyopneumoniae infection could trigger systemic PCV2 infection to progress to PMWS as shown in this model.
It has been proposed that the diagnosis of PMWS requires a pig to exhibit characteristic clinical signs (wasting), histologic lesions (lymphoid depletion and granulomatous inflammation), and PCV2 antigen associated with these lesions. 49 Under field conditions, many other factors in addition to PCV2 infection can cause a pig to exhibit signs of wasting as described in the context of the current definition of PMWS. It is also not clear whether wasting actually means weight loss or decrease in rate of average daily gain (and the appearance of wasting) compared with healthy cohorts. Furthermore, the current PMWS definition 49 neither accounts for distribution or severity of lesions nor for relative amount of PCV2 antigen in association with the lesions. Lymphoid depletion and granulomatous inflammation can be present in various degrees and diffusely distributed throughout all lymphoid tissues or restricted to one or two lymph nodes in individual pigs. It is unknown what consequences these variable manifestations of lymphoid lesions have for individual pigs. It seems likely that even the pigs with mild lymphoid depletion may be immunologically compromised to some degree and perhaps more susceptible to secondary infections and more at risk for vaccination failures. To address this problem, and to properly distinguish between PCV2 infection (subclinically or associated with other diseases) and PMWS, an overall severity score for microscopic lymphoid lesions that accounted equally for distribution (lymph nodes, spleen, and tonsil), severity of lymphoid depletion, severity of inflammation, and amount of PCV2 antigen was calculated. Microscopic lesions (mild, moderate, and severe PCV2-associated lymphoid depletion and granulomatous inflammation) have been staged on cases of naturally occurring PMWS. 10,45 We believe that the mean group severity score for lymphoid lesions as developed and used in this model allows for a more uniform understanding and interpretation of PCV2-associated lesions.
Some dual-infected pigs in this study had severe dyspnea (persistent coughing and moderate-to-severe respiratory score) in combination with decreased weight gain and macroscopic (more than 40% of the lung affected by lesions) and microscopic lung lesions (moderate-to-severe bronchointerstitial pneumonia) consistent with PRDC. The macroscopic and microscopic lung lesions of these pigs were remarkably similar to those observed with PRRSV/PCV2 coinfection. 24 These pigs also had lymphoid depletion, histiocytic inflammation of lymph nodes, and PCV2 antigen associated with the lesions. However, we found that the lymphoid lesions lacked the overall severity and distribution to clearly classify them as cases of PMWS. We believe that PCV2-associated pneumonia and PCV2-associated lymphoid depletion are more descriptive terms in such cases.
It is of interest that at 21 and at 35 DPI, the dual-infected pigs had a significantly (P < 0.05) higher serum antibody response to M. hyopneumoniae compared with the singular M. hyopneumoniae–infected pigs. There was also a trend toward a higher antibody response to PCV2 in dual-infected pigs compared with the singular PCV2-infected pigs at 35 DPI (P = 0.055). Similar results have been reported with PPV antibody response in PCV2/PPV dual-infected pigs. 41 The three pigs that were diagnosed with PMWS in the current study did not seroconvert to M. hyopneumoniae by 21 DPI. The one PMWS pig that remained in the study through 35 DPI was negative at 21 DPI and had a weak positive antibody response (near ELISA cutoff value) to M. hyopneumoniae by 35 DPI. Three of the four PMWS pigs seroconverted to PCV2 at 21 DPI but had lower S/P ratios than the other pigs in this group. Similarly, the antibody response in PCV2-infected, immunostimulated, PMWS-affected pigs was markedly reduced. 33 The dual-infected pigs in this study certainly had more stimulus in the form of inflammation than did the singular-infected pigs. Thus, in the context of additional stimulation (coinfection or nonspecific immunostimulation), the B-cell response in PCV2-infected non-PMWS pigs appeared to be enhanced due to PCV2 infection, whereas it is severely diminished in pigs that progressed to PMWS. Upregulation of certain cytokines during the course of inflammation can lead to enhanced proliferation of T lymphocytes and B lymphocytes, inducing B cell– and plasma cell–dependent antibody production, which could explain the better response in the non-PMWS pigs. It appears that if the inflammatory stimulation reaches a certain level, it has more of a counterproductive effect subsequently leading to PMWS. However, the quality of the enhanced B-cell response, the presence of an enhanced T-cell response, and inflammatory aspects in the dual-infected pigs were not evaluated in this study, thus, making any conclusions about possible alterations of the immune system speculative. This coinfection model should allow us to further explore those questions.
The PCV2/M. hyopneumoniae coinfection model presented here demonstrated that, by themselves, PCV2 and M. hyopneumoniae induce mild transient respiratory disease and lesions but in concurrent infection induce severe respiratory disease and lesions consistent with PMWS as well as PRDC. The sequence of infection (M. hyopneumoniae followed 2 weeks later by PCV2) of this dual-infection model was designed to address our hypothesis that M. hyopneumoniae lesions are present first and create an ideal environment and cell-type accumulation in the lung for PCV2 replication. This may or may not be typical of the sequence of infection in the field. After weaning and mixing of pigs at 2–4 weeks of age, M. hyopneumoniae is commonly spread slowly throughout the population. PCV2 infection likely coincides with waning of maternal antibodies over a large window of time from 5–11 weeks of age. 43 The exact timing and sequence of infection of pigs with these two pathogens likely differs from pig to pig and from herd to herd, and these different scenarios should be further explored to see if there are similar effects as observed in this model.
This dual-infection model will be very useful for testing vaccine and antimicrobial intervention strategies. Experimental PCV2 vaccines have been shown to protect PCV2-infected pigs from developing the characteristic PCV2-associated lymphoid lesions. 15,16 M. hyopneumoniae vaccines are well established in the field and have been shown to be economically beneficial in reducing losses associated with respiratory disease. 35 However, recent field studies have indicated that M. hyopneumoniae vaccination in PMWS-affected herds actually increased the incidence of PMWS. 4,32 This information has led some practitioners to discontinue the use of M. hyopneumoniae vaccination programs in affected herds. Based on the PCV2/M. hyopneumoniae model, the detrimental effects from lack of M. hyopneumoniae control may be more costly than the detrimental effects of vaccine-induced enhancement of PCV2 replication. This risk and benefit associated with M. hyopneumoniae vaccination should be thoroughly assessed. It also needs to be determined what vaccine regimen (singular PCV2 or M. hyopneumoniae or a combination of both) is most efficient in reducing PRDC and PMWS in pigs from herds that are at a high risk of dual infection by M. hyopneumoniae and PCV2.
Footnotes
Acknowledgements
This study was funded by a grant from the Iowa Livestock Health Advisory Council and through an unrestricted gift from Merial. We thank Barb Erickson, Nancy Upchurch, and Kris Ruebling for technical assistance, Dave Cavanaugh for contributions to development of IHC, Jim Foss for photography, and Pete Thomas for assistance with the animal work. The Image Analysis Facility is supported by the Iowa State University Office of Biotechnology.