
Abstract
Telomerase has been studied extensively in human and murine tumors, but little is known about the role of telomerase in the tumor biology of other vertebrate species such as the chicken. We studied the telomerase activity of the lymphoblastoid cell line derived from lymphomas induced by Marek's disease virus (MDCC-MSB1) compared with another avian cell line (PA5) and peripheral blood lymphocytes (PBL) using the telomeric repeat amplification protocol (TRAP) Assay. Telomerase activity in MDCC-MSB1 was 4.5 times greater than in the PA5 cell line and normal avian lymphocytes. These results demonstrate for the first time that telomerase is more intense in one transformed cell line than in normal cells, suggesting a potential role for telomerase in carcinogenesis induced by an avian virus.
In a number of organisms, including humans, rodents, and small animals, telomerase elongates the 3′ ends of chromosomes (telomeres) with strings of TTAGGG repeats. 17 The repeat sequence is specified by a short template sequence within the RNA component of mammalian telomerase. 2,4 Maintenance of the telomeric repeat hexamer and its associated protein complex is essential for chromosome stability. Telomerase activity has been detected in some normal cells as well (basal epithelium of the skin, canine and feline and murine lens epithelium, and lymphocytes) and also in immortalized cell lines and malignant cells derived from tumors in mice and humans. 5 The regulation of telomerase in normal and tumor tissues appears to be different in the two systems. Rodents differ from humans in both telomere length and regulation of telomerase. In contrast to human cells, constitutive telomerase activity is found in several postnatal somatic tissues in the mouse and rat. Even the immortalization barrier in the three species is different. 3
Avian species may provide a good system to study the degree of conservation of the telomere clock and telomerase regulation in vertebrates. The longevity of avian species (they live for up to 30 years) suggests that bird cells may require control of cell lifespan comparable with that needed by human cells to avoid immortalization and carcinogenesis. This is supported by the extremely low rates of spontaneous immortalization exhibited by both human and avian cells in culture compared with rodents. 9 There has been no complete study in chickens to date linking telomerase activity and tumors, and there are no published reports on telomerase activity in avian lymphoblastoid cell lines. The aim of the present study was to measure telomerase activity in avian tumor cell lines.
Two Marek's disease lymphoblastoid cell lines were used, i.e., MSB1 cell line derived from splenic tumors and PA5 derived from testicular tumors. 1 Cells were seeded at 1 × 105 cells/ml and incubated at 40 C with Roswell Park Memorial Institute medium containing 10% fetal calf serum (Gibco BRL, France). Peripheral blood lymphocytes (PBL) were obtained from heparinized blood (50 IU/ml) by lymphocyte-separation medium LSM-1077 (Eurobio, Les Ulis, Cergy-Pontoise, France) centrifugation for 20 minutes at room temperature and used to measure telomerase activity.
For telomerase activity 1 ml (105 cells) was centrifuged at 1,400 × g for 20 seconds. Cell extracts were flash-frozen in liquid nitrogen and stored at −80 C. Protein concentrations were determined with BCA protein assay (Pierce, Brebieres, France) and adjusted to 1 µg/µl. Telomerase activity was determined with the polymerase chain reaction–based telomeric repeat amplification protocol (TRAP) assay described for human cell extracts. 6 The fluorescent TRAP assay can be performed to analyze telomerase activity in a semiquantitative manner. This assay was performed in two stages: 1) telomerase-mediated extension of an oligonucleotide TS primer, which serves as substrate of telomerase, and 2) amplification of the elongated products by PCR with reverse primer CX-ext and the TS primer. In this assay, an internal amplification standard (ITAS) was included to verify PCR amplification efficiency. We used tetramethylrhodamine (TAMRA)-labeled forward TS and CX-ext as reverse primers as originally described for human telomerase assay. 7 Fluorescent and nonfluorescent primers were synthesized by Eurogentec (Seraing, Belgium).
Quantitative TRAP assays require the inclusion of ITAS, and this was prepared as previously described. 6,16 The ITAS construct contains the 135-nucleotide RNase P element from Thermus thermophilus. It is flanked by sequences complementary to both the CX-ext and TS primers used in the TRAP, thus allowing its coamplification during PCR. The reaction mixture was incubated for telomerase extension at 30 C for 30 minutes, followed by 30 cycles at 94 C for 30 seconds, 50 C for 30 seconds, and 72 C for 1 minute. The samples were then analyzed by ABI prism 310 (Applied Biosystems, Paris, France). The relative telomerase activity (RTA) was calculated as follows: RTA = fluorescence intensity of 6 telomeric hexamer repeats (black peaks)/fluorescence intensity of ITAS.
We analyzed two dilution series of MSB1 cell extracts (100 and 1,000 cells). The 6-bp telomerase ladder of the avian cell line MSB1 is shown as black peaks in Fig. 1. We found that RTA was expressed in 100 cells (RTA = 3.2 ± 0.3) and 1,000 cells of the MSB1 cell line (RTA = 5.5 ± 1.1). Moderate levels of telomerase activity were detected in PBL from control chickens (RTA = 0.7 ± 0.1) and from the PA5 cell line derived from the testicular tumors (RTA = 0.8 ± 0.02) and compared with the MSB1 cell line derived from the splenic tumors (RTA = 3.2 ± 0.06) (Fig. 2). These results show that the MSB1 cell line has more telomerase activity than PBLs or nonlymphoblastoid cell lines (PA5).
Induction of telomerase activity in the MDCC-MSB1 cell line. 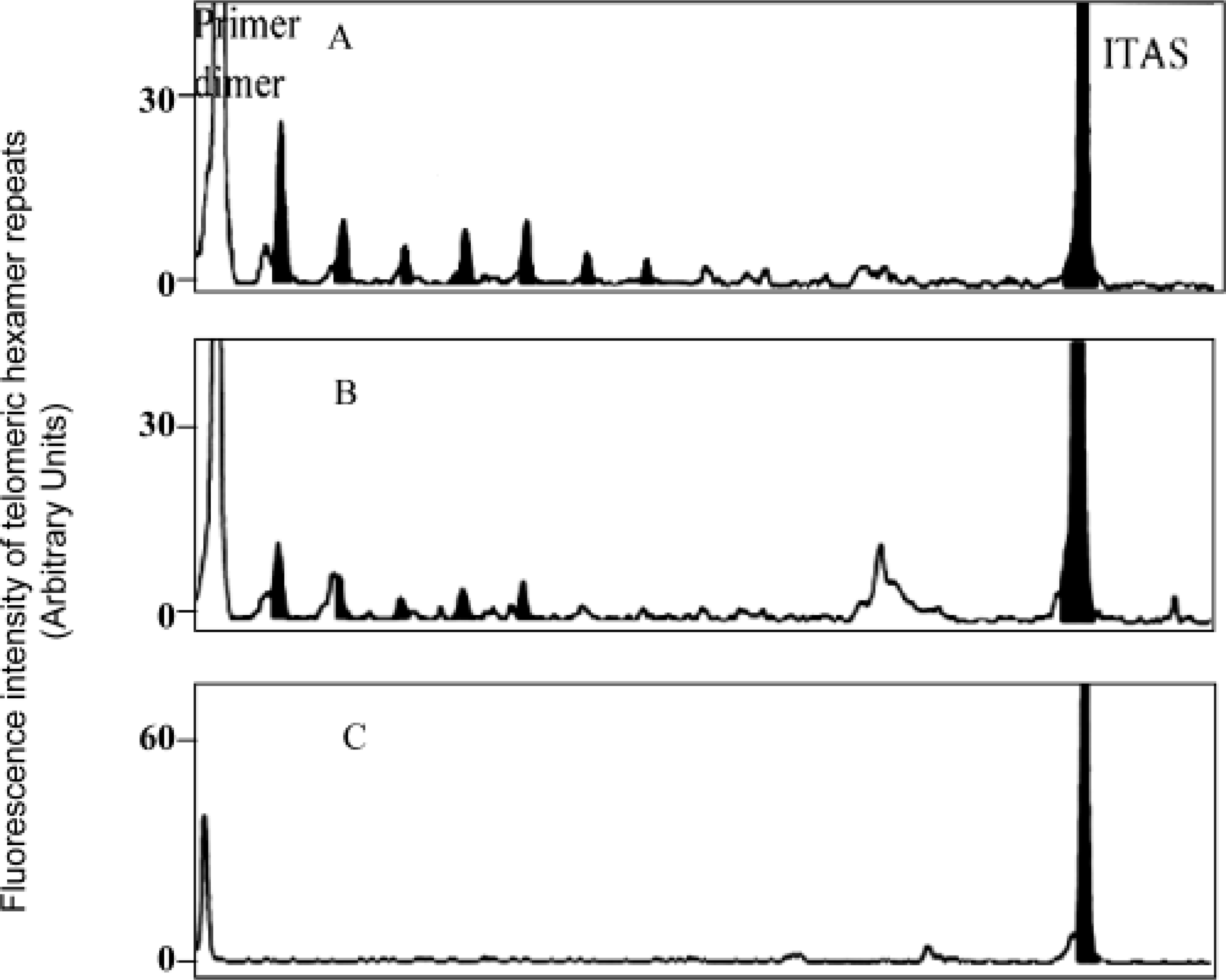
Relative telomerase activity in MDCC-MSB1 cell line in comparison with the untransformed lymphocyte (PBL) and PA5 cell line. Telomerase activity of 300 ng cellular protein extract was measured by TRAP assay as described previously. The value of relative telomerase activity was normalized by dividing fluorescence intensity values of six telomeric hexamer repeats by the value for ITAS. The data presented are the means of three independently performed experiments.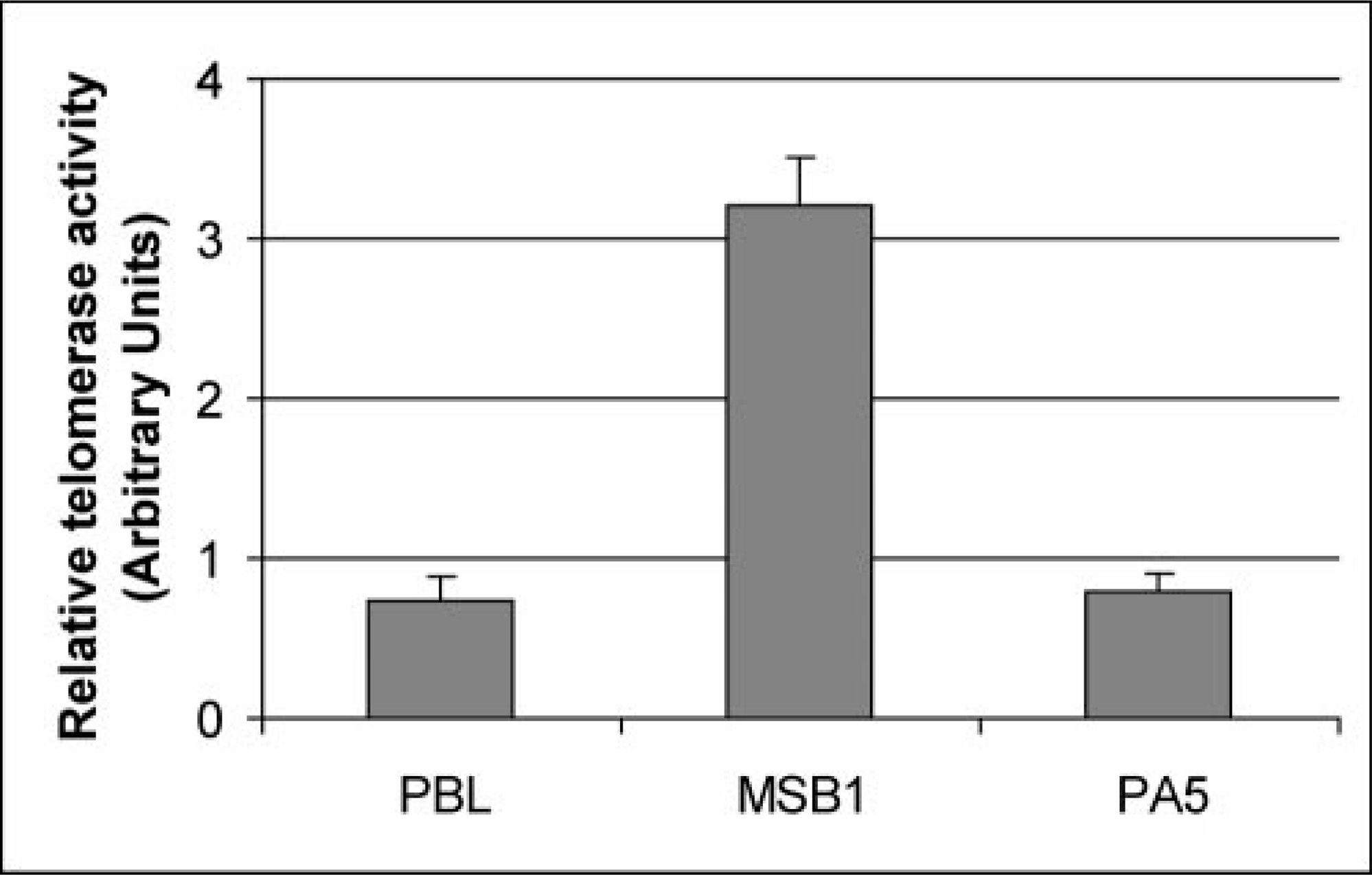
As shown in Fig. 3, the activity of the enzyme could be abolished by heating the MSB1 extract with RNase or heat before the TRAP assay. Omitting the TS primer from the telomerase reaction and adding it only for the PCR also abolished the activity. However, the absence of the CX-ext primer in the telomerase elongation step and addition to PCR amplification did not affect the activity.
Characterization of telomerase reaction in the MSB1 cell line. 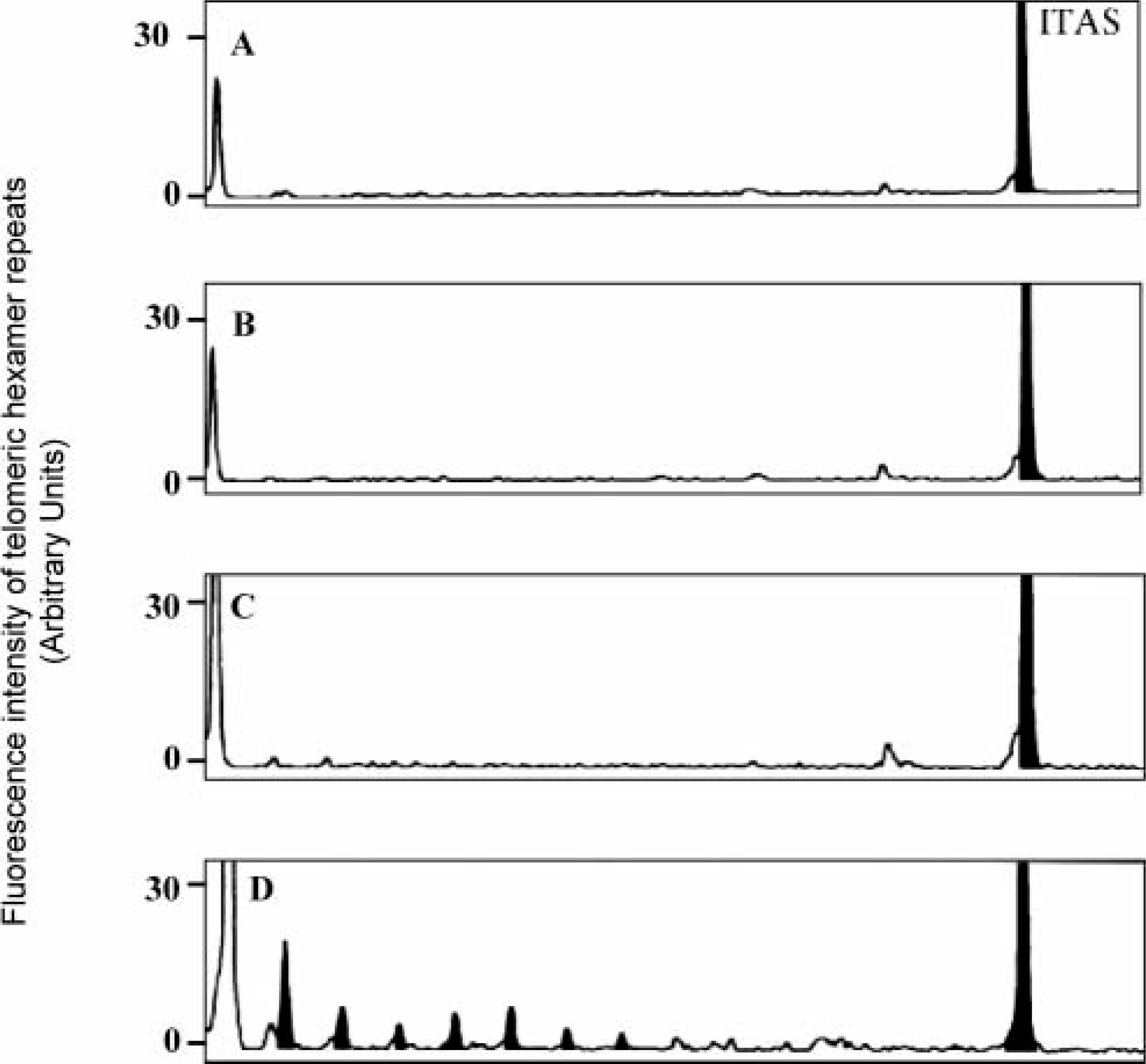
This study is the first to document the induction of telomerase activity in an avian transformed cell line MSB1. Our results indicate that telomerase activity in the MSB1 cell line was detected in 100 cell equivalents. This level is in agreement with previous reports noting high telomerase activity in tumor cell lines, 13 whereas it was found to be low in normal lymphocyte cells in chickens and another avian cell line, PA5. A very recent study of functional characteristics of chicken telomerase reveals the enzyme resembles the mouse and humans cases, in that its enzyme activity is related to the proliferation capability of the tissues. 15 So, we may speculate that the comparable telomerase activity between the PA5 cell line and PBL is probably related to their low proliferation capability. Further studies will be needed to address this issue.
There are two possible explanations for detecting telomerase activity in MSB1 cell lines. The first is deregulation of Bcl-2, which in human cancer cell lines such as HeLa cell lines is closely linked with increased levels of telomerase activity. 10 The finding that Bcl-2 enhances telomerase activity without elongation of telomeric terminal restriction fragment in HeLa cells is interesting because it raises the possibility that additional pathway(s) such as Bcl-2 deregulation may influence telomerase activity in cancer cells. Ohashi et al. reported deregulation of Bcl-2 expression and constitutive expression of Bcl-xL in the MSB1 cell line. 12 It remains to be seen whether the increase in the levels of telomerase activity observed in MSB1 is a phenomenon restricted to Bcl-2 deregulation or a general event associated with other gene products.
The second possible explanation for detecting telomerase activity in MSB1 may be related to the mutation of tumor suppressor protein p53. p53 inhibits tumor formation by inducing cell cycle arrest or apoptosis in response to a variety of types of cell damage. 8 Recent studies have shown that p53 mutations account for approximately 50% of human cancers and represent the most frequent genetic lesion in breast cancer. 11 The fact that telomerase activity is upregulated in most human cancers and highly proliferative somatic cells and downregulated with cell cycle exit and differentiation indicates that cell cycle regulators may also be involved in the regulation of telomerase. It is of interest that a short form of p53 transcript has been detected in MSB1 cell lines. 14 This short form is produced by deletion of the nucleotides encoding the c-terminal part of the core domain. The biologic significance of this short form of p53 transcript still remains to be established.
The telomerase activity in MSB1 is not regulated by only two possible mechanisms, Bcl-2 dysregulation or p53 mutation. Other possibilities such as c-myc upregulation, transcription factor–dependent regulation of expression of the gene for the catalytic subunit of telomerase, and posttranslational modification such as phosphorylation and numerous others. In summary, our results demonstrate for the first time that the avian lymphoblastoid cell line expresses high levels of telomerase, suggesting a potential role of telomerase during carcinogenesis induced by avian virus.