
Abstract
The brain from a 15-month-old, black female Angus, with a 48-hour history of central nervous system disease, was submitted to the Oklahoma Animal Disease Diagnostic Laboratory. Microscopic findings consisted of acute, multifocal meningoencephalitis, with neuronal degeneration and necrosis and gliosis. Viral isolation yielded noncytopathic bovine viral diarrhea virus (BVDV). Virus genotyping classified the virus as BVDV type 2. Immunohistochemical labeling for BVDV antigens with BVD MAb 3.12F1 clone was prominent in the cytoplasm of neurons, glial cells, ependymal epithelium, perivascular macrophages and spindle cells, smooth muscle cells, and intravascular monocytes of the cerebrum and brain stem. Laboratory results support that tissue alterations occurred as a result of BVDV type 2 infection. In the absence of other clinical signs related to BVDV infection and using the microscopic and laboratory evidence presented, we propose that the BVDV type 2 isolated from this case may represent a neurovirulent strain of the virus. To the best of our knowledge, this is the first report of brain lesions and neuronal viral antigen localization in BVDV genotype 2 viral infection, acquired either congenitally or postnatally.
Keywords
A major pathogen, bovine viral diarrhea virus (BVDV), affects cattle worldwide. 18,19 As a member of the family Flaviviridae, genus Pestivirus, BVDV is a small, enveloped, positive-sense RNA virus. 24 Virulence, antigenic and genetic differences, along with various animal factors, including age, sex, pregnancy, stage of gestation, immunocompetence, and others, are responsible for the highly variable clinical manifestations or outcomes of BVDV infection. 2,4–6,8,9,19,27 This has led to the classification of BVDV into three genotypes: 1a, 1b, and 2. 20–22,26 Earlier work on experimental and natural infections of cattle with BVDV revealed that the viral antigen was localized mainly in the cytoplasm of neurons of the myenteric ganglion. 29
Although several reports on neuropathogenesis and distribution of BVDV antigen in the central nervous system of cattle have been published, very little is known about the neuropathology and neurovirulence of BVDV type 2 in cattle. 3,7,11,14–16 In this report, we describe the brain lesions and viral antigen localization in a case of BVDV genotype 2 infection.
The brain of a 15-month-old, black Angus heifer, with a 48-hour history of central nervous system disease, was submitted to the Oklahoma Animal Disease Diagnostic Laboratory (OADDL). The submitting veterinarian reported that the clinical signs progressed from slight gait abnormalities and lethargy to severe ataxia, recumbency, seizures, and bilateral bulging of the eyes or proptosis. During the postmortem examination, the veterinarian removed and submitted the entire, unfixed brain to OADDL.
Fresh samples from the posterior medulla, cerebellum, occipital cortex, and hippocampus were collected and submitted to the Oklahoma State Health Department for rabies testing by fluorescent antibody staining. Additional samples from the occipital cortex were stored at −20 C. The remaining brain tissue was fixed in neutral buffered, 30% formalin solution for 5 days. Representative sections of the fore-, mid-, and hind brain were trimmed, routinely processed, embedded in paraffin, sectioned at approximately 5 µm, stained with hematoxylin and eosin, and evaluated for microscopic lesions.
The occipital cortical tissue, which had been stored at −20 C after rabies testing, was thawed and used for virus isolation. 17 In brief, about 1 g of tissue was homogenized in nine parts of minimum essential medium (MEM). After centrifugation and filtration through a 0.45-µm pore size, 50 µl of inoculum was adsorbed onto 1-day-old monolayers of Madin-Darby bovine kidney (MDBK) cells in a 24-well plate for 2 hours at 37 C. The monolayers were refed with MEM containing 5% fetal bovine serum and incubated at 37 C. The plates were examined daily for evidence of viral cytopathic effects for 6 days, after which the cells were trypsinized and stained for the presence of noncytopathic (NCP) BVDV as described previously. 23 When positive BVDV staining was observed on the cell cultures, the supernatant was harvested and subjected to a reverse transcriptase–polymerase chain reaction procedure that simultaneously confirmed the presence of BVDV and determined the genotype (1 or 2) of the virus isolate. 10
Representative sections of the cerebrum and medulla were subjected to routine immunohistochemical detection of BVDV antigens. The primary antibody used was BVD MAb 3.12F1, an affinity-purified, murine origin, anti-BVDV IgG1 monoclonal antibody provided by J. T. Saliki (OADDL). As a negative control, an isotype-matched mouse IgG1 antibody of irrelevant specificity (Vector Laboratories, Burlingame, CA) was used. Antibody dilutions were carried out against positive controls to find the greatest dilution of the antibody, which yielded optimal staining. The optimal dilution of the BVD MAb 3.12F1 antibody was determined with a final concentration of 1 µg/ml. Positive tissue controls consisted of formalin-fixed, paraffin-embedded MDBK cell pellets that were infected with the NCP BVDV isolated from the suspect case and archival spleen tissue of a known NCP BVDV–positive case. Negative tissue controls consisted of formalin-fixed, paraffin-embedded, mock-inoculated, BVD-negative MDBK cell pellets and archival cerebral tissue from a BVDV–culture negative, 12-month-old bovine brain. As an assay control, the primary antibody was eliminated and substituted with phosphate-buffered saline containing 0.05% Tween 20. An affinity-purified, rabbit anti-mouse IgG (0.5 mg/ml, 61-0100; Zymed, San Francisco, CA) served as the secondary antibody. As the tertiary antibody, an affinity-purified, biotinylated goat anti-rabbit IgG (H + L)–specific antibody (0.75 mg/ml, 62-6140; Zymed) was used. The conjugate was streptavidin–horseradish peroxidase, and the chromogen was Nova RED substrate kit for peroxidase (SK-4800, Vector Laboratories). The tissue sections were counterstained with Mayer hematoxylin, dehydrated in alcohol, cleared in toluene, and mounted in a resinous medium.
The results from the state's health department were negative for rabies virus infection. Sections from the parietal, temporal, and frontal cerebral cortex, basal ganglia, thalamus, hippocampus, cerebellum, and medulla were evaluated microscopically. In all regions of the brain tissue, including the leptomeninges, there were multiple, discrete perivascular cuffs of lymphocytes, plasma cells, foamy, pigment-laden macrophages or gitter cells, and occasional neutrophils (Fig. 1). Widely scattered, individual neuronal degeneration and necrosis and occasional hypereosinophilic axonal spheroids existed in all regions as well (Fig. 2). The most prominent tissue alterations were observed in the cerebrum; there were multiple, discrete foci of cortical necrosis or infarction, with loss and rarefaction of the neuropil, increased prominence of blood vessels, gliosis, and vacuolation. Gliosis consisted predominantly of gemistocytic astrocytes and scattered microglia. Gemistocytic astrocytes had abundant eosinophilic cytoplasm, with occasional clear vacuoles; their nuclei were eccentrically located and hypochromatic with moderate pleomorphism. Some gemistocytic astrocytes were binucleated (Fig. 1). In some areas, perivascular spaces were filled with homogenous, eosinophilic proteinaceous material. In the thalamus, the tunica media of an arteriole was diffusely mineralized (Fig. 3). The cerebellum had no significant tissue alterations.
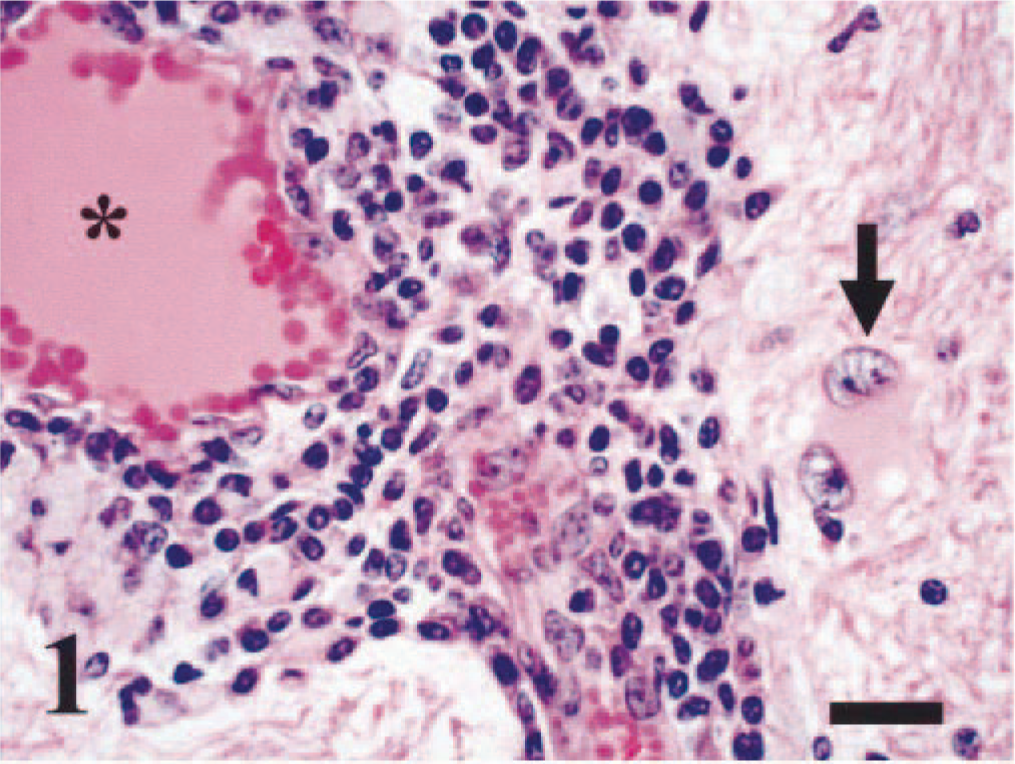
Brain, cerebrum; bovine. A dense collection of lymphocytes, plasma cells, and pigment-laden macrophages expand the perivascular spaces. The blood vessel (∗) is lined by hypertrophic endothelium. Within the white matter neuropil, there is a binucleated, gemistocytic astrocyte, with swollen, vacuolated eosinophilic cytoplasm and eccentrically located nuclei (arrow). HE. Bar = 20 µm.
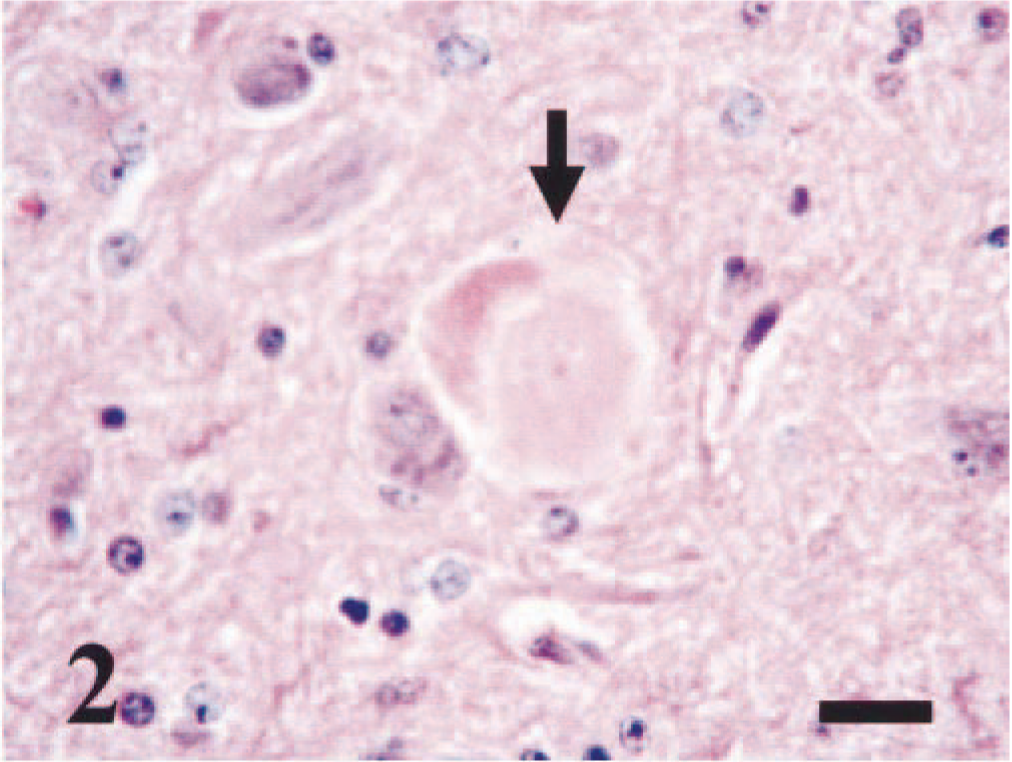
Brain, basal ganglia; bovine. There is discrete neuronal necrosis, with swollen, hypereosinophilic cytoplasm and karyolysis (arrow). HE. Bar = 25 µm.
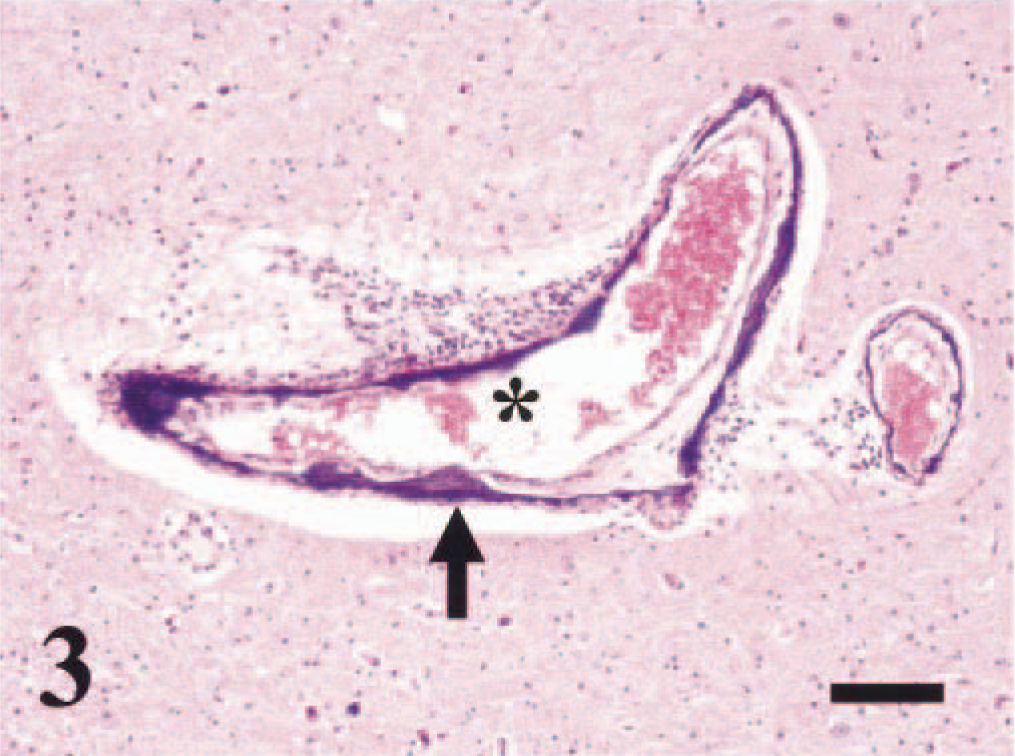
Brain, thalamus; bovine. Deposition of mineralized material (arrow) has obscured the tunica media of an arteriole (∗) and its branch. There are mild perivascular collections of lymphocytes, plasma cells, and pigment-laden macrophages. HE. Bar = 90 µm.
Brain tissue virus isolation yielded no cytopathic effects. However, fluorescent antibody staining of inoculated cells using a BVDV-specific monoclonal antibody indicated that the brain tissue was immunoreactive for NCP BVDV. Virus genotyping classified the virus as BVDV type 2.
The brain of the suspect case had specific staining of BVDV antigens throughout, both in areas associated with and without an inflammatory response. In the cerebrum and medulla, strong, specific cytoplasmic staining was observed of some, scattered normal and necrotic neurons (Fig. 4) in all glial cells, including gemistocytic astrocytes and other neuroglia (Fig. 5), and in some perivascular macrophages and perivascular spindle cells (Fig. 6). There was occasional cytoplasmic staining of the smooth muscle cells from the tunica media as well as of intravascular monocytes and ependymal epithelium.
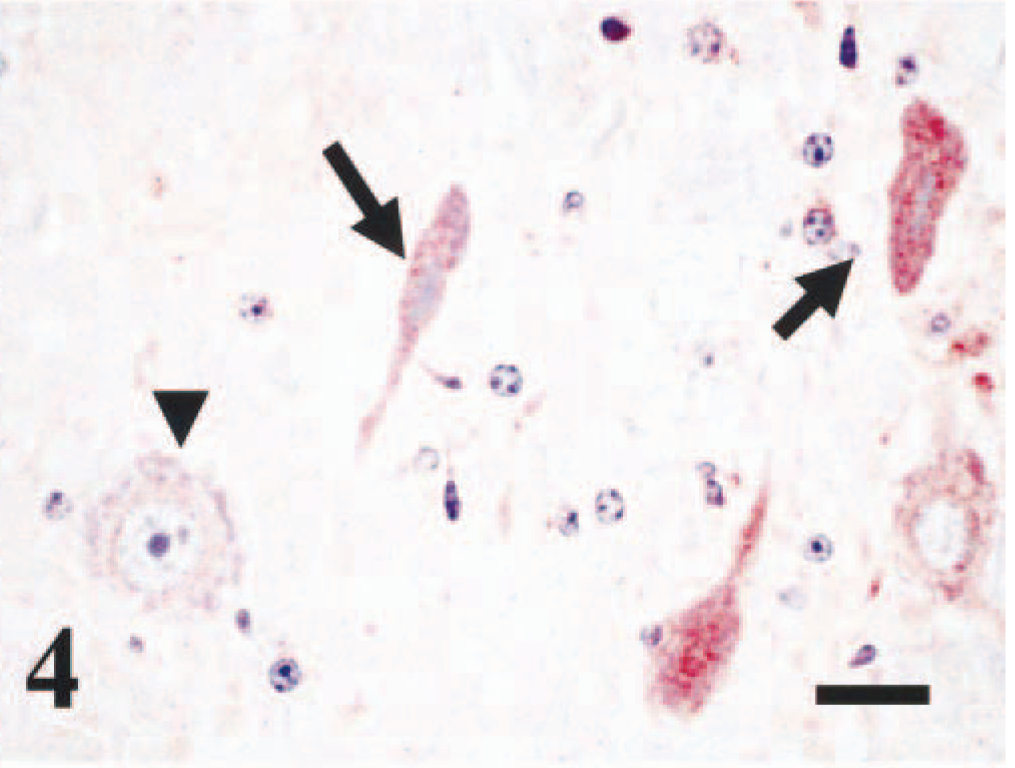
Brain, medulla; bovine. Several necrotic neurons, with shrunken cytoplasm and pycnotic nuclei, stain positive for BVDV antigens (arrows). Arrow head depicts a normal neuron, with negative immunostaining for BVDV antigens. Immunoperoxidase staining, BVD MAb 3.12F1 (1 µg/ml). Nova Red chromogen. Hematoxylin. Bar = 20 µm.
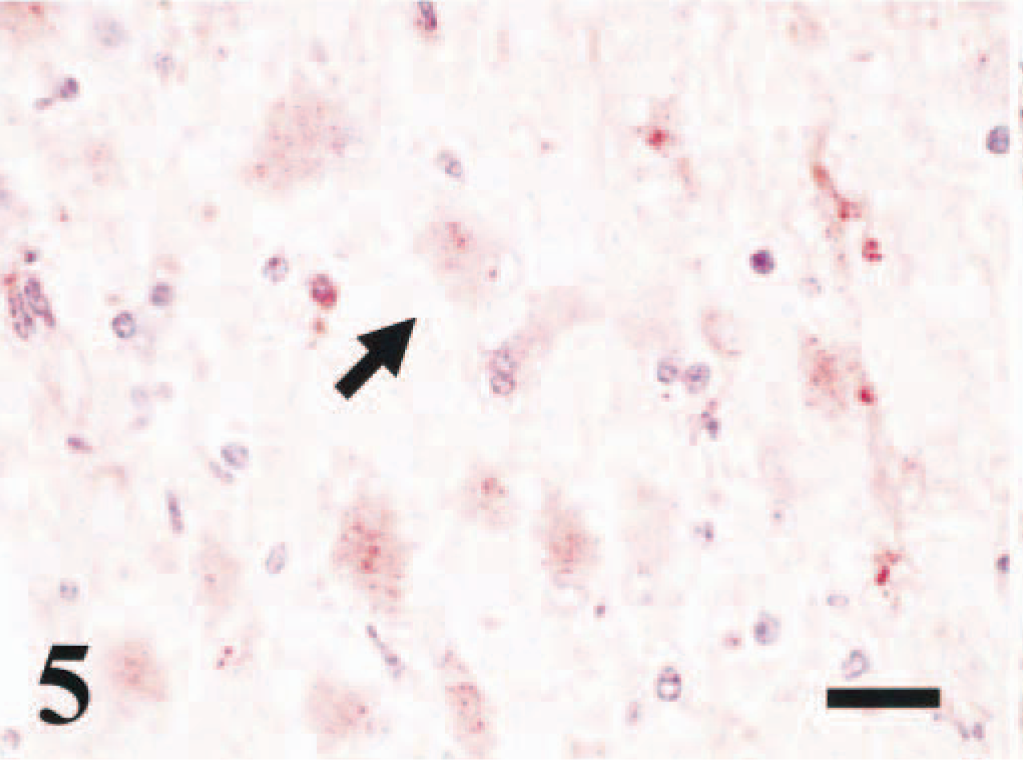
Brain, cerebrum; bovine. Within the white matter there are scattered gemistocytic astrocytes, with cytoplasmic immunostaining of BVDV antigens (arrow). Immunoperoxidase staining, BVD MAb 3.12F1 (1 µg/ml). Nova Red chromogen. Hematoxylin. Bar = 20 µm.
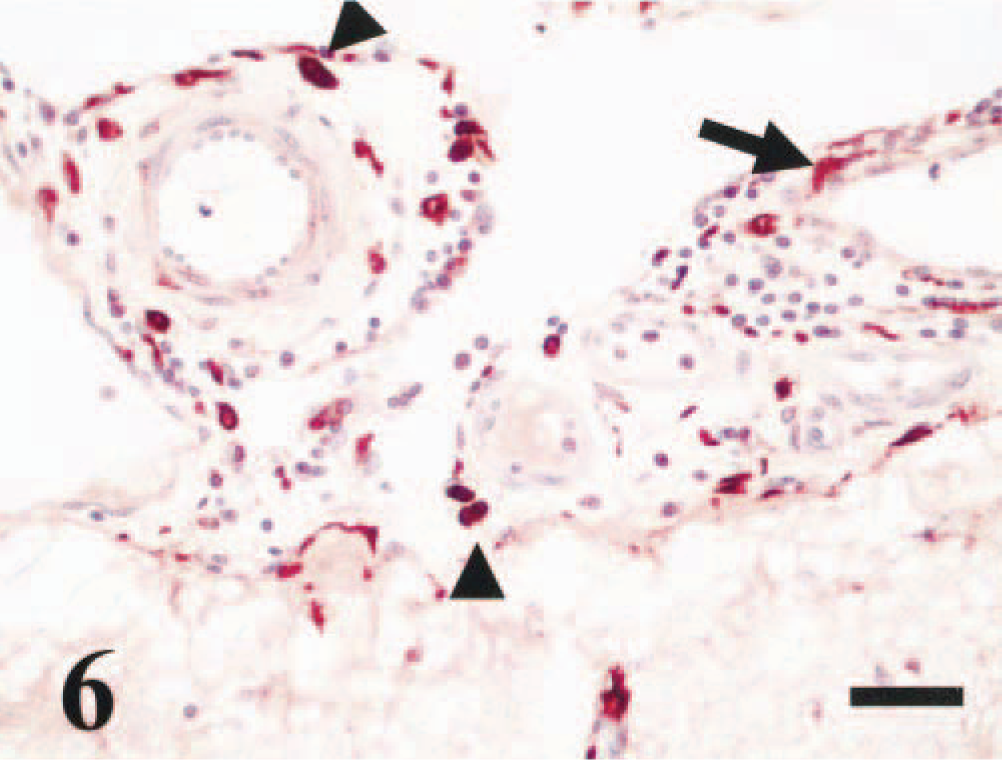
Brain, cerebrum, leptomeninges; bovine. There is mild expansion of perivascular spaces with lymphocytes, plasma cells, and BVDV-positive macrophages (arrowheads). Strong immunostaining for BVDV antigens exist within perivascular spindle cells (arrow). Immunoperoxidase staining, BVD MAb 3.12F1 (1 µg/ml). Nova Red chromogen. He matoxylin. Bar = 33 µm.
In general, little is known about the mechanisms of neuropathology and neurovirulence of BVDV in cattle. The most commonly reported brain lesion in calves with transplacental infection of BVDV (of undetermined genotype) is cerebellar hypoplasia often associated with hydranencephaly, internal hydrocephalus, microencephaly, or porencephaly. 15 The acute neurological signs observed in this heifer were consistent with the microscopic and laboratory findings described. The severity of the lesions along with the laboratory evidence presented indicates that this animal died from acute encephalitis associated with BVDV type 2 infection. In our laboratory, differentiation between a persistently infected (PI), prenatally acquired BVDV infection versus an acute (postnatally acquired) infection requires detection of the BVDV in two consecutive tests, 3 weeks apart. Because only brain tissue was received, differentiation between a PI and an acutely infected animal was not possible. Both persistent and acute infections with BVDV genotype 2 have been reported in the literature. 22 To the best of our knowledge, this article is the first report of brain lesions and neuronal viral antigen localization in BVD genotype 2 viral infection, acquired either congenitally or postnatally.
On the basis of our findings, we propose that the BVDV type 2 strain isolated from the brain in this case may represent a neurovirulent strain of the virus. Without experimental inoculation of animals, this proposal is unproven; however, the predominance of neurological disease in the absence of other remarkable signs generally attributed to BVDV infection, together with virus isolation and typing and demonstration of BVDV antigen in affected neurons, indicates that this viral isolate may be a neurovirulent strain of BVDV. In contrast, other BVDV type 2 virulent strains, the VS-260 and 1373 strains, have been reported to be of low neuropathogenic or neurovirulent potential to sheep and cattle, respectively. 24,26
The BVDV type 2 strain isolated from this case may potentially serve to study mechanisms of neuropathology of BVDV. Immunohistochemical detection of BVDV antigens within neurons, glial cells, ependymal epithelium, perivascular macrophages, vascular smooth muscle, and intravascular monocytes suggests that the virus crosses the blood-brain barrier and disseminates throughout the brain. Microscopic findings indicated that viral infection was associated with neuronal death. In light of this individual neuronal necrosis, we propose that this particular strain of NCP BVDV type 2 may induce apoptotic necrosis of neurons, with subsequent spheroid formation and loss of myelin. The association of BVDV with the induction and suppression of the apoptotic pathways has been described recently. 1,12,13,25,28 Additional studies are needed to corroborate such proposed mechanisms of BVDV type 2 neurovirulence and neuropathogenicity in cattle.