
Abstract
Nonsuppurative encephalitis was experimentally induced in 3-week-old piglets by a single intravenous inoculation of either of two strains (IB 2001 or AS-6) of Japanese encephalitis flavivirus (JEV) isolated from field pigs. The lesions, which consisted of neuronal necrosis, neuronophagia, glial nodules, and perivascular cuffing, were distributed in the cerebrum, midbrain, pons, medulla oblongata, and cerebellum, particularly in the gray matter of the frontal and temporal lobes and thalamus. The gray matter of the spinal cord of piglets that were given the AS-6 strain also was affected. JEV antigen was immunohistochemically detected in the cytoplasm of the nerve cells in the cortex of the frontal and temporal lobes and in the gray matter of the thalamus and midbrain. Two JEV strains isolated from field pigs exhibited neurovirulence, inducing nonsuppurative encephalitis in piglets.
Japanese encephalitis flavivirus (JEV) is one of four major encephalitic flaviviruses of public health importance, the other three being West Nile virus, St. Louis encephalitis virus in North America, and Murray Valley encephalitis virus in Australasia. 14 The disease is transmitted by mosquitoes of the genus Culex. 4,11,12 Many animals, such as swine, horses, dogs, chickens, ducks, and reptiles, are infected in nature. 4,11,12 Pigs are considered to be amplifier hosts with viremia that makes the virus available to mosquitoes. 4,11,12 Horses may develop fatal encephalitis, whereas infection in swine is generally unapparent, except for stillbirths and abortions when pregnant sows are infected and aspermia when boars are infected. 11,12 Among animals experimentally infected with the virus, only horses and donkeys develop encephalitis, whereas mice, rats, calves, 12 and monkeys 1 do not. There are few reports on encephalitis in piglets caused by postnatal infection with JEV. Moreover, there are limited reports on the immunohistochemical distribution of viral antigens of JEV in domestic animals. The neurotropism of JEV in pigs has not been well characterized.
In 2001, we examined four 40-day-old piglets that were exhibiting a wasting syndrome without neurologic signs. In these piglets, there was a nonsuppurative meningoencephalitis characterized by perivascular cuffing, focal gliosis, neuronal necrosis, and neuronopha-gia in the cerebrum, brainstem, and cerebellum (M. Yamada, unpublished). JEVs (IB 2001 strain) were isolated from a few piglet tonsils using an established African green monkey kidney (Vero) cell, but JEV was not isolated from the brains. Other pathogenic viruses, including Aujeszky's disease herpesvirus, hog cholera pestivirus, and porcine parvovirus, were not detected using direct immunofluorescence or the latex agglutination test. JEV was not detected in the brain using immunohistochemistry for JEV antigens. It was not evident whether the JEV isolate was neurovirulent and caused the encephalitis in the piglets. Three-week-old piglets were experimentally inoculated intravenously with JEV isolated from the wasting piglets and histo-logically examined to clarify the pathogenesis of this virus. This study describes the histologic lesions of JEV in 3-week-old piglets and the immunohistochem-ical distribution after experimental infection.
Materials and Methods
Virus
The JEV IB 2001 and AS-6 strains were used in this study. The IB 2001 strain was isolated from the tonsils of two piglets experiencing a wasting syndrome in Japan in 2001. This viral strain was passaged twice in IB-RS2 cells derived from porcine kidneys and then once in a Vero cell culture. The highly virulent AS-6 strain, isolated from porcine blood during the 1967 epizootic of Japanese encephalitis, 5 was obtained as a comparative control for JEV infection and was used after three passages in mouse brain. Infected Eagle's minimum essential medium culture fluid 18 (Nissui Corp., Tokyo, Japan) was used to inoculate the piglets.
Animals
Fourteen 3-week-old, specific-pathogen–free pigs with no serum antibodies against JEV, Aujeszky's disease herpesvi-rus, hog cholera pestivirus, or porcine reproductive and respiratory syndrome arterivirus were used. The pigs were divided into three groups for the experiment: six AS-6 strain infected pigs, six IB 2001 strain infected pigs, and two un-infected controls. The groups were housed separately to prevent cross-infection.
Experimental designs
The pigs were anesthetized with ketamine hydrochloride (30 mg/kg, intramuscularly) and xylazine (2.0 mg intramuscu-larly) before inoculation with JEV.
Pig Nos. 1–6 were inoculated intravenously with a syringe containing 5 ml of 106 tissue culture infection dose (50%) (TCID50)/ml of JEV IB 2001 strain. Pig Nos. 7–12 received 5 ml of 106 TCID50/ml of JEV AS-6 strain. Pig Nos. 13 and 14 were inoculated intravenously with 5 ml of uninfected culture medium as negative controls. Pig Nos. 1–3 (group 1), 7–9 (group 3), and 13 were observed clinically at post-inoculation day (PID) 3, and pig Nos. 4–6 (group 2), 10–12 (group 4), and 14 were observed clinically at PID 7. After observation, the piglets were euthanatized by an intravenous injection of sodium pentobarbital and necropsied.
Histopathologic and immunohistochemical examination
Tissue samples for histologic examination were collected from each piglet after gross examination at necropsy. These included liver, spleen, kidney, heart, lung, stomach, small and large intestine, lymph node, brain, spinal cord, trigeminal ganglia, tonsil, eye, and skeletal muscle. Tissues were fixed in 10% neutral-buffered formalin, embedded in paraf-fin wax, and processed by routine methods for staining with hematoxylin and eosin (HE).
The presence of JEV antigens in the formalin-fixed, par-affin-embedded tissues of the brain and spinal cord was demonstrated by the streptavidin–biotin complex immunoperox-idase (SAB-IP) method using an SAB kit (Nichirei Corp., Tokyo, Japan). Anti-JEV rabbit serum (Dr. Y. Miura, National Institute of Animal Health, Japan) was used as the primary antibody, at a dilution of 1 : 256. Samples were counterstained with hematoxylin. Tissue sections from the uninfected controls (pig Nos. 13 and 14) and sera from a nonimmunized rabbit were used as controls.
Results
Fever (40–41 C) was detected in pig Nos. 1–3 (group 1), which were necropsied 3 days after inoculation with the IB 2001 strain. No macroscopic lesions were observed at necropsy. Nonsuppurative encephalitis characterized by perivascular cuffing of mononu-clear cells and multifocal gliosis was histologically observed in the gray matter of the cerebrum. The lesions tended to surround blood vessels located at the junction of the gray and white matter. Neutrophils were observed within the lumen of some blood vessels. Occasional neuronal necrosis and neuronophagia were observed in the cortex of the frontal lobe. No signifi-cant changes were observed in the white matter. The distribution of histologic lesions is summarized in Table 1. Lesions in the cerebrum were distributed primarily in the frontal and temporal lobes; no lesions were observed in the occipital lobe. There also were lesions in the thalamus. Lesions in the midbrain, pons, and medulla oblongata were less prominent than those in the cerebrum. No histologic changes were observed in the other organs, including the cerebellum, spinal cord, and trigeminal ganglia.
Distribution of histologic lesions and JEV antigen in the CNS of piglets experimentally infected with JEV
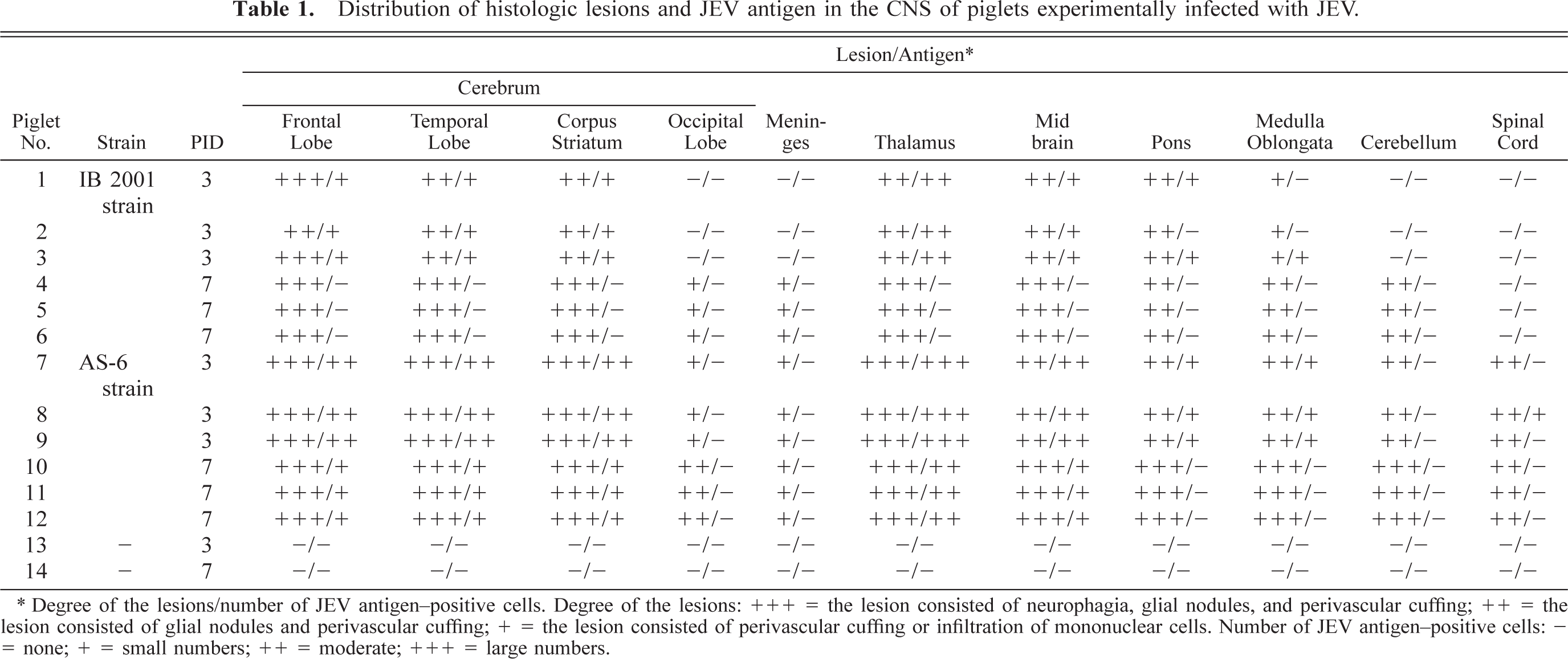
∗ Degree of the lesions/number of JEV antigen-positive cells. Degree of the lesions: +++ = the lesion consisted of neurophagia, glial nodules, and perivascular cuffing; ++ = the lesion consisted of glial nodules and perivascular cuffing; + = the lesion consisted of perivascular cuffing or infiltration of mononuclear cells. Number of JEV antigen–positive cells: = none; + = small numbers; ++ = moderate; +++ = large numbers.
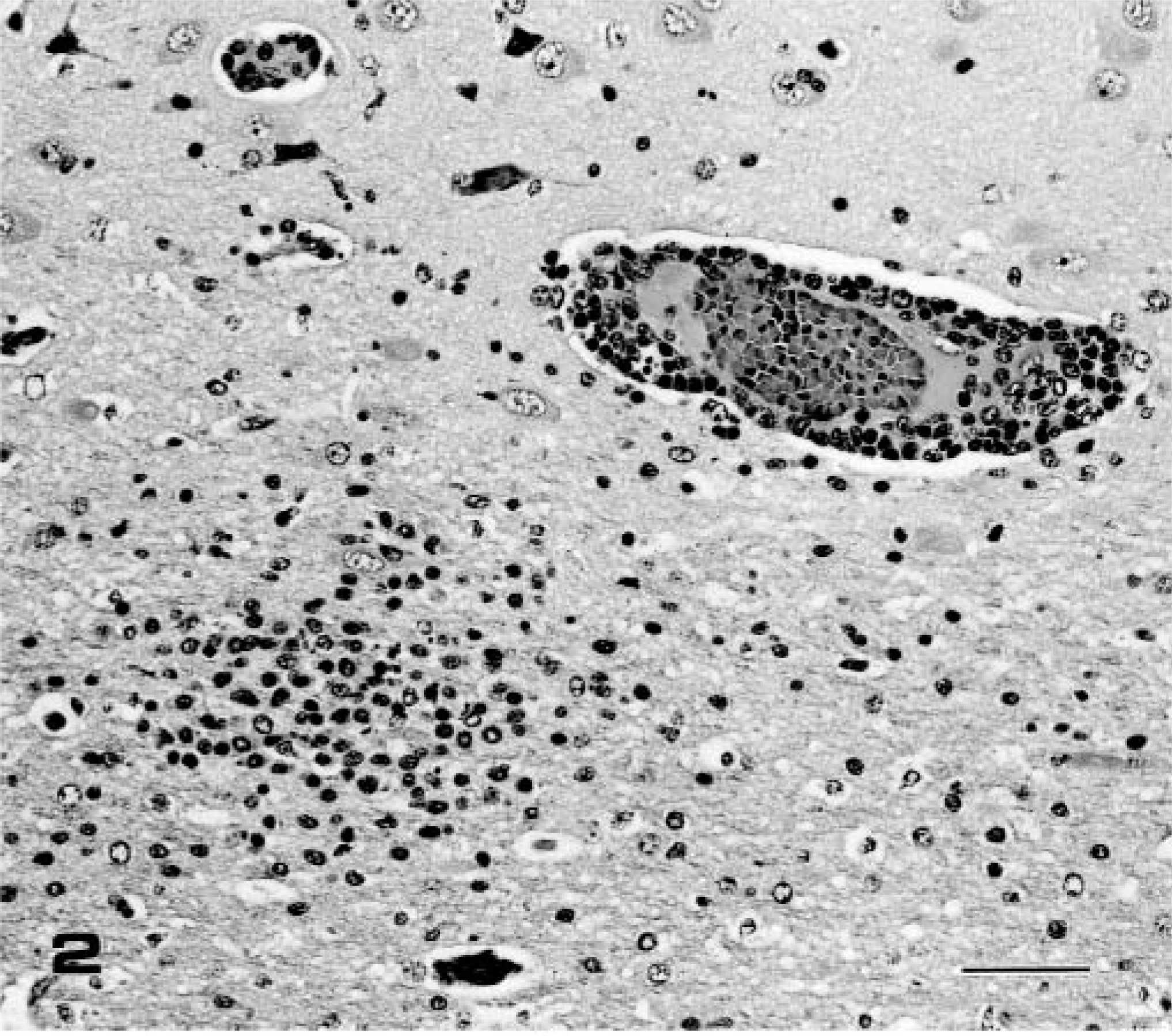
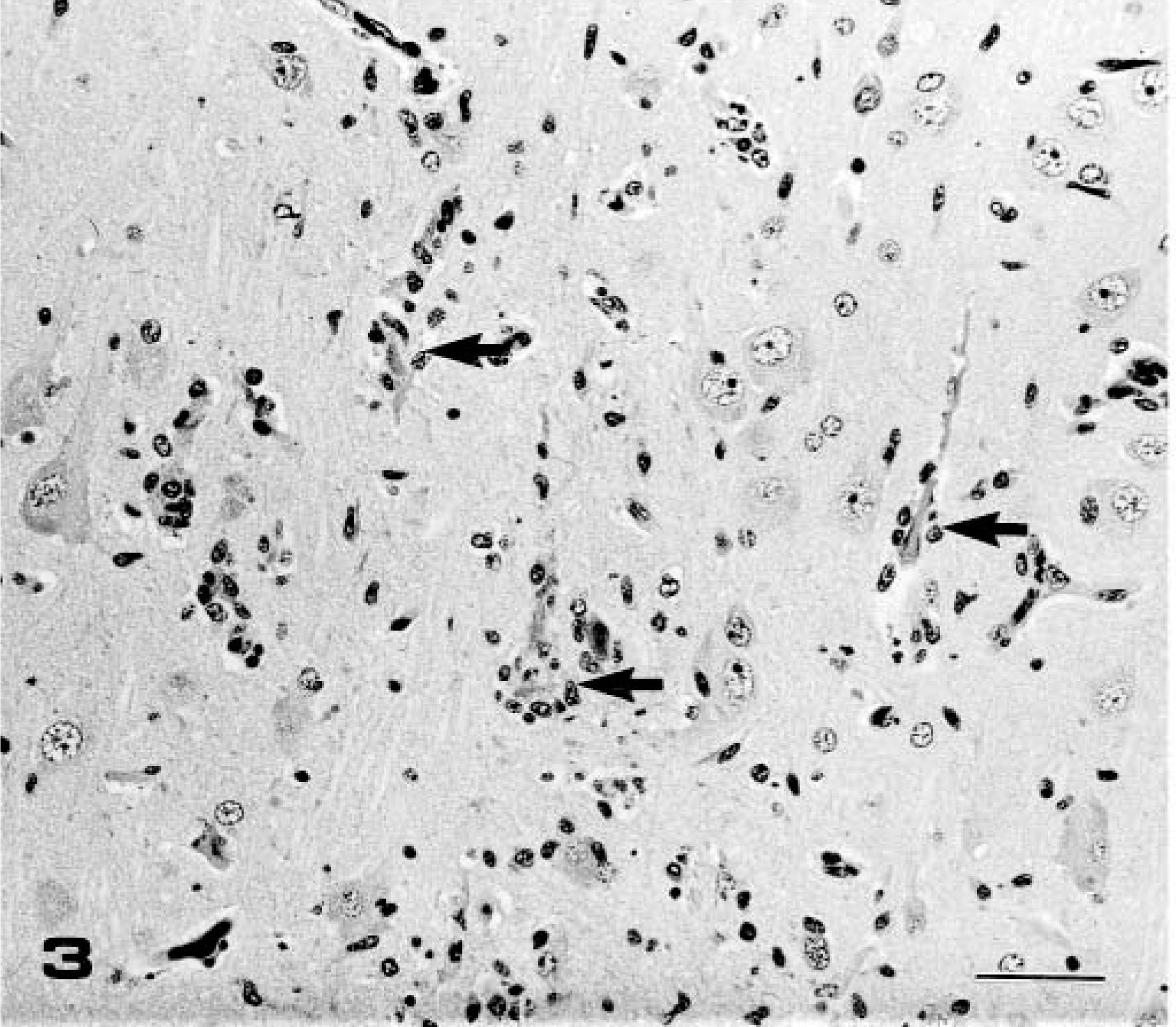
Pig Nos. 4–6 (group 2), which were necropsied on PID 7 with the IB 2001 strain, developed fever (40–41 C), depression, and slight tremor of the hindlimbs. Fever decreased in all piglets at PID 4. The brain was slightly swollen and edematous at necropsy in all pigs. Nonsuppurative encephalitis characterized by perivas-cular cuffing of the mononuclear cells, multifocal gli-osis (Figs. 1, 2), and severe neuronal necrosis and neu-ronophagia (Fig. 3) was observed in the gray and white matter. Mild meningitis also was observed in the cerebrum, midbrain, and cerebellum. The cerebral hemispheres and thalamus were significantly affected, and the lesions spread to the midbrain, pons, medulla oblongata, and cerebellum. No histologic changes were observed in the other organs, including the spinal cord and trigeminal ganglia.
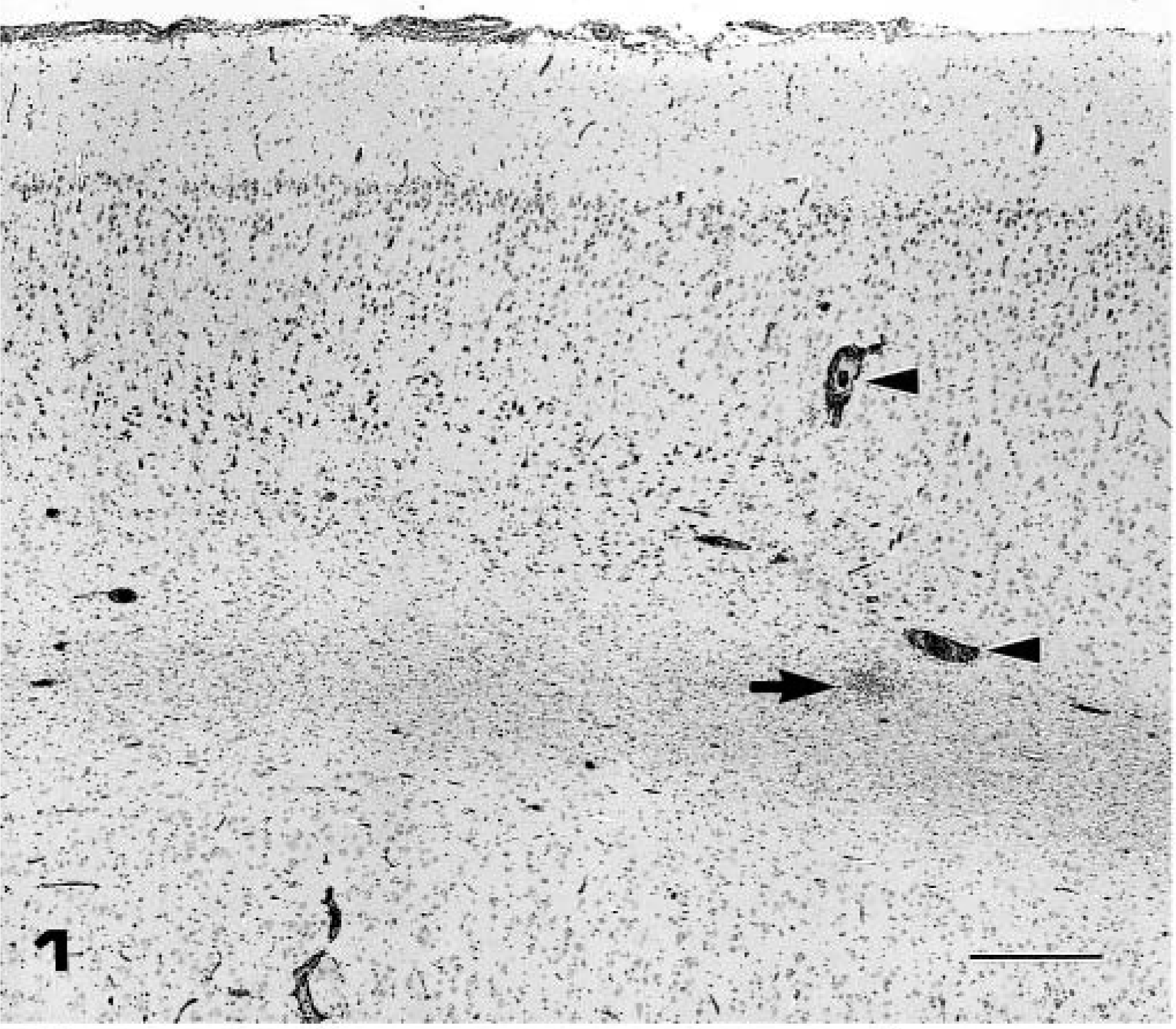
Frontal lobe of cerebrum; pig No. 5, infected with IB 2001 strain. Nonsuppurative encephalitis with perivascular cuffing (arrow heads), focal gliosis (arrow), and mild meningitis. HE. Bar = 400 µm.
Frontal lobe of cerebrum; pig No. 5, infected with IB 2001 strain. Higher magnification of Fig. 1. Lymphocytic perivascular cuffing and focal gliosis were observed. HE. Bar = 50 µm.
Frontal lobe of cerebrum; pig No. 5, infected with IB 2001 strain. Higher magnification of Fig. 1. Neuronal necrosis and neuronophagia (arrows) in the lower layer of the cerebral cortex were observed. HE. Bar = 50 µm.
Pig Nos. 7–12 (groups 3 and 4), inoculated with AS-6 strain, developed fever (41–42 C), depression, and severe tremors of the hindlimbs. The brain was edematous in both PID 3 and PID 7 pigs. Nonsuppu-rative meningoencephalomyelitis was histologically observed in the cerebrum (Fig. 4), cerebellum, thalamus, midbrain, pons, medulla oblongata, and spinal cord, both in PID 3 and PID 7 pigs. Although the histologic lesions in pigs given the AS-6 strain were more severe than those in pigs given the IB 2001 strain, the distribution and spread of the lesions in the brains of pigs given AS-6 strain were similar with those in pigs given IB 2001 strain.
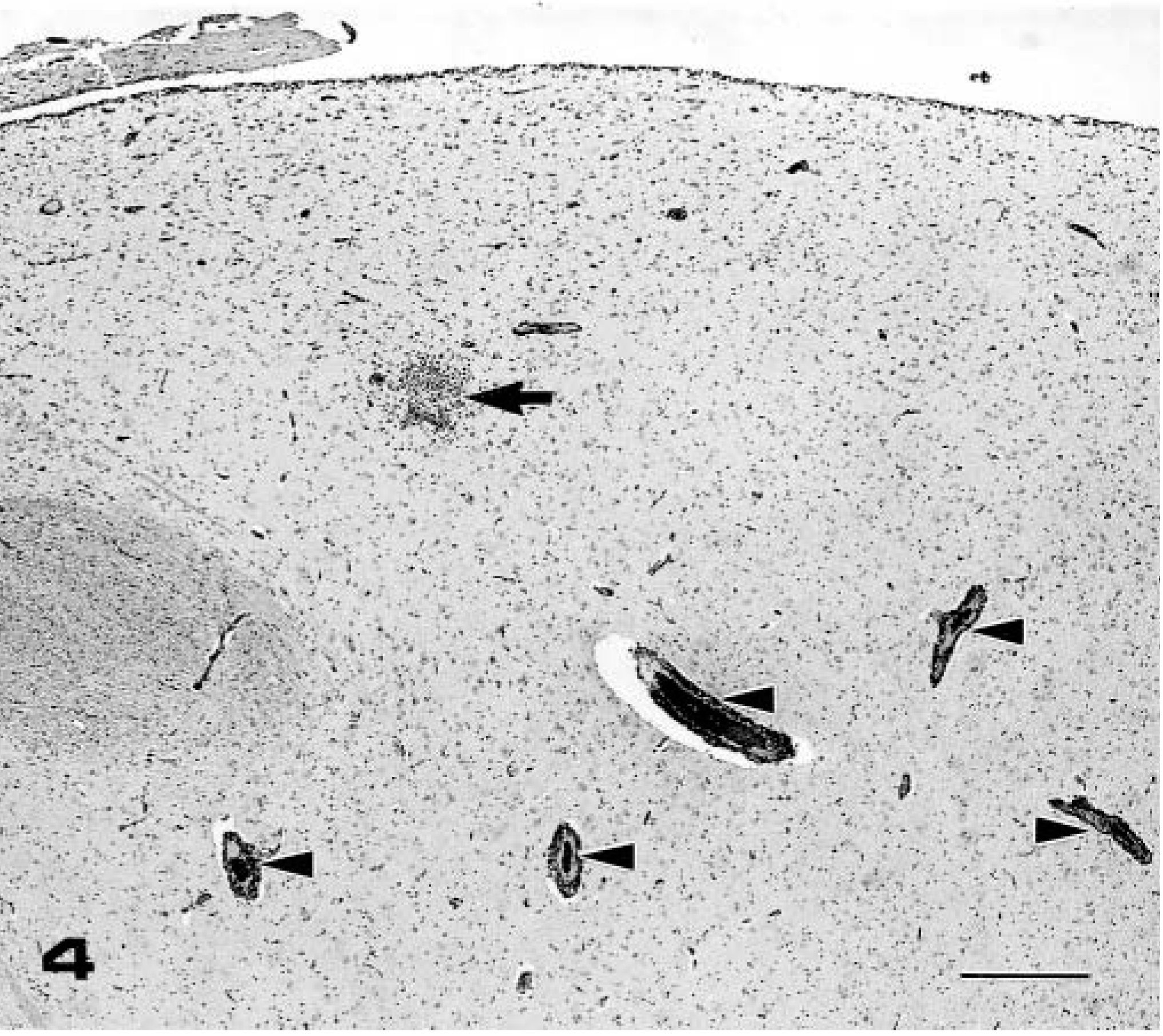
Thalamus; pig No. 7, infected with AS-6 strain. Perivascular cuffing (arrow heads) and focal gliosis (arrow) in the gray matter of the thalamus were observed. HE. Bar = 400 µm.
JEV antigens were immunohistochemically detected in the cytoplasm of nerve cells in the cortex of the frontal and temporal lobes and in the gray matter of the thalamus and midbrain in both PID 3 and PID 7 pigs given AS-6 strain (pig Nos. 7–12) (Table 1). A few immunolabeled, degenerating or necrotic neurons were surrounded by microglial cells. JEV antigen also was detected in the cortex of the frontal and temporal lobes and in the gray matter of the thalamus in PID 3 pigs given IB 2001 strain (Pig Nos. 1–3) (Table 1). The thalamus contained numerous JEV-positive neurons (Fig. 5). JEV-positive neurons were found in fourth, fifth, and sixth layers of the temporal cortex. In contrast, no JEV antigen was detected in the brains of PID 7 pigs given IB 2001 strain (pig Nos. 4–6) (Table 1). There was no JEV antigen in the white matter. The choroids plexus, ependyma, and glial cells were also negative in all examined cases. JEV antigen was detected in the cytoplasm of large neurons in the spinal cord of only one pig given AS-6 strain (pig No. 8). No viral antigen was detected in two noninfected control pigs (pig Nos. 13 and 14).
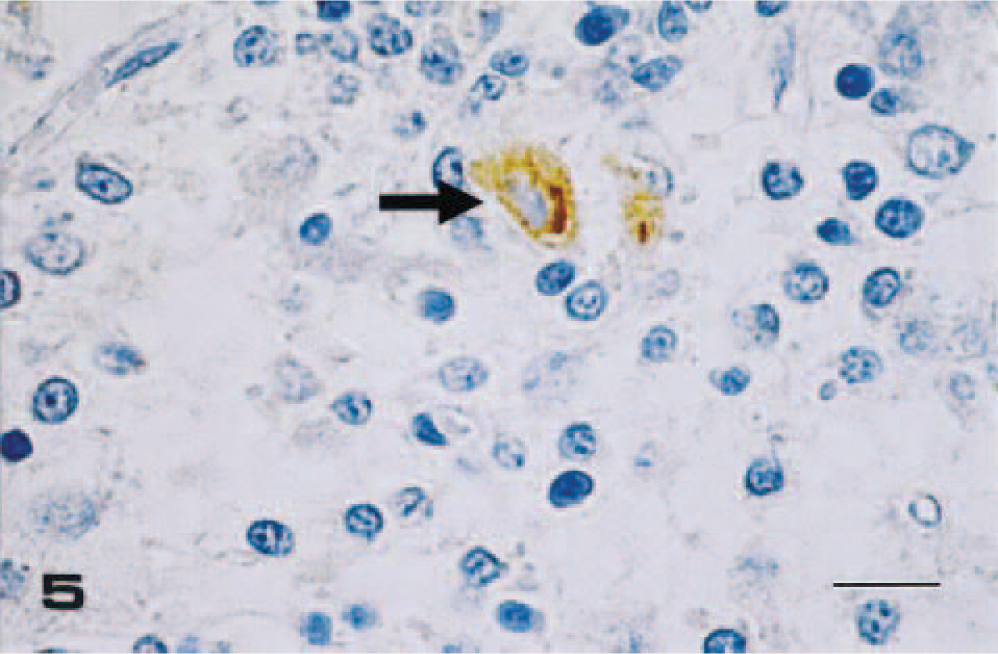
Thalamus; pig No. 3, infected with IB 2001 strain. JEV antigen is detected in the cytoplasm of a neuron (arrow). SAB-IP method, hematoxylin counterstain. Bar = 15 µm.
Discussion
Encephalitis with neuronal necrosis, neurophagia, glial nodules, and perivascular cuffing was experimentally induced in 3-week-old piglets by intravenous inoculation of JEV. JEV infection in swine is associated with stillbirths and abortions in sows and aspermia in boars; there have been few report of encephalitis in piglets. 12 We demonstrated that both JEV IB 2001 and JEV AS-6 strains are neurovirulent and able to induce nonsuppurative encephalitis in piglets.
The distribution of JEV antigen as detected by im-munohistochemistry was consistent with the lesions in the brains of piglets inoculated with AS-6 strain on PID 3 and PID 7 and with IB 2001 strain on PID 3. The immunohistochemical distribution of viral antigens of JEV has been studied in humans 3,9,10,13 and in experimentally infected rats. 16,17 In humans and rats, JEV infects and destroys neurons selectively, mostly in the brainstem, thalamus, basal ganglion, and the lower layers of the cortex. 3,9,10,13 However, there are limited reports on the distribution of viral antigens of JEV in domestic animals, and the neurotropism of JEV in pigs has not been well characterized. The viral antigens of the JEVs of this study were immunohisto-chemically detected in the cytoplasm of nerve cells located mainly in the cerebral cortex and thalamus. These results were nearly consistent with those of human cases. 3,9,10,13 We confirmed that JEV selectively infects the neurons in the brains of experimentally inoculated pigs in the same manner as it does in humans and in other experimental animals.
The earliest evidence of central nervous system (CNS) infection in pigs was observed mainly in the gray matter on PID 3. CNS lesions, consisting of glial nodules and perivascular cuffing, initially tended to start around blood vessels located at the junction of gray and white matter. The lesions had spread to the white matter on PID 7. We speculate that the earliest lesions in the brain of piglets infected with JEV localize in the gray matter of the brain and then spread to the white matter. JEV reportedly enters the CNS via the cerebral spinal fluid either by endothelial cell, macrophage, or lymphocyte infection or by a hematoge-nous route. 11 There were no viral antigens in the cho-roid plexus and ependyma in this study, suggesting that dissemination of the infection was not by a cere-brospinal fluid pathway. JEV likely spreads to the brain by a hematogenous route.
The result of this study that an intravenous, single inoculation of 106 TCID50 of two strains of JEV induced encephalitis in 3-week-old piglets suggests that both IB 2001 and AS-6 strains can easily pass the blood-brain barrier (BBB) and invade the brain when severe viremia is induced, even in postnatally infected pigs. The BBB of animals and humans generally prevents viral invasion into CNS, and thus JEV gains entrance through some type of breach that is created in the barrier at the time of the viral transmission. 7 It is reported that a JEV-induced cytokine alters the integrity of the BBB during JEV infection, and this breakdown of BBB aids virus entry into the brain resulting in the development of encephalitis. 8,15 We considered in this study that the development of viremia may be important for the passage of JEV through the BBB, resulting in neural infection in these piglets. No major lesions have previously been found in pigs postnatally infected with JEV, 11 but the younger the animal, the more vulnerable it is to JEV infection. 2,6 The age susceptibility of rats to JEV infection is related to the degree of neuronal maturity, 16 suggesting that the relationship between the degree of neuronal maturity and neural infection in piglets should also be examined to clarify the pathogenesis of the neural infection. It also is currently unclear how JEV travels from the periphery to the CNS in pigs. Further studies are necessary to elucidate the mechanisms of the neural infections of JEV that cause encephalitis in piglets.
Footnotes
Acknowledgements
We thank M. Kobayashi and M. Shimada for preparing the pathologic sections and Dr. Y. Ando and T. Fujisawa for preparing the photographs.