
Abstract
Neonatal diarrhea occurred at two swine breeding farms in Hokkaido. Ten piglets aged 2 to 4 days were examined. Grossly, significant changes were confined to the small intestine. The mucous membrane was muddy and thickened. The intraluminal contents from the jejunum to the colon were liquid and yellow. In the small intestine, numerous Gram-negative bacilli preferentially adhered to the apex of villi. The mucosa was erosive with villous atrophy. There were bacilli also in the lamina propria and in the cytoplasm of degenerated enterocytes. Nonhemolytic Escherichia coli strains, belonging to serogroup E. coli O18 and possessing K88 fimbriae, were isolated from the small intestine. They could not be classified into any of the diarrheagenic E. coli groups because of the absence of genes of LT, STh, STp, VT1, VT2, eae, invE, and ipaH. After inoculation of the isolates on HEp-2 cells, some bacilli were engulfed by cytoplasmic projections resembling membrane ruffles and subsequently were localized in cytoplasmic vacuoles or free in the cytoplasm. These findings support the view that the present E. coli O18 is a new invasive strain enteropathogenic to piglets.
In humans, diarrheagenic Escherichia coli are divided into at least six categories: enteropathogenic (EPEC), enterotoxigenic (ETEC), enteroinvasive (EIEC), enterohemorrhagic (EHEC), enteroaggregative (EAggEC), and diffusely adhering (DAEC). 6 ETEC was regarded as a primary etiologic agent in enteric colibacillosis of piglets. 3,12 In addition, attaching and effacing E. coli (AEEC) has been isolated from postweaning diarrhea. 12,14 Other diarrheagenic strains are poorly documented in this species. 3,7,8
We isolated E. coli strains from neonatal piglets with diarrhea in Hokkaido, Japan. Although the isolates could not be classified bacteriologically into any of the established E. coli groups, the bacilli were invasive into epithelial cells. The aim of this study is to record a new diarrheagenic E. coli that is enteroinvasive in nature.
Neonatal diarrhea occurred at swine breeding farms A and B in Hokkaido, Japan, but there was no epidemiologic relation between them. From October 1997 to February 1998, the first outbreak occurred at farm A, where 2 stock boars and 32 brood sows were kept. Two-day-old piglets developed signs of diarrhea, and death ensued within a few days. The mortality rate per litter varied from 20% to 50%, and the total number of animals lost was approximately 350. In farm B with 20 stock boars and 440 brood sows, diarrhea was detected in 2- to 5-day-old piglets in May 2001, and about 60 piglets died.
At necropsy, the stools were watery and yellowish or of light grayish hue but were bloody and mucoid in a few cases. No symptoms other than diarrhea and dehydration were seen, and affected piglets were all neonates.
Six piglets from farm A (pig Nos. 1–6) and four from farm B (pig Nos. 7–10), aged 2 to 4 days, were examined. Approximately 104 to 1010 colony-forming units per gram of intestinal tissue of nonhemolytic E. coli were isolated from the small intestines examined. Six E. coli strains were isolated from farm A, and the slide agglutination method (Denka Seiken Co., Tokyo, Japan) demonstrated that all belonged to serotype O18:H16. Among the four strains derived from farm B, one was O18:H21 (pig No. 10), but the other three were untypable. In all the 10 strains, K88 fimbriae were present. However, there were no strains producing either verotoxin (VT), heat-labile (LT), or heat-stable toxin (ST). 3,6 The detection tests for the former two were performed with a reversed passive latex agglutination kit, and the test for ST was done with an enzyme immunoassay kit (Denka Seiken Co.). In no strains was any product amplified by polymerase chain reaction (PCR) using primer sets for LT, STh, STp, VT1, VT2, eae, invE, and ipaH. 3,6,10,13 The oligonucleotide primers for eae were synthesized on the basis of the sequences reported, and the other primers were prepared commercially (Takara Shuzo Co., Kyoto, Japan). 5 Two strains from farm A were examined by the Serény test but were negative. 9 Thus, the isolated bacteria did not correspond to the five diarrheagenic E. coli groups (EPEC, ETEC, EIEC, EHEC, and AEEC). 6 Pathogenic organisms such as rotavirus, Clostridium, Salmonella, Cryptosporidium, and coccidium were not identified in the intestinal contents of any case.
At necropsy, similar findings were observed in all cases. Curds were present throughout the stomachs in all piglets, and the mucus membrane was normal. Significant changes were confined to the small intestine, and the mucous membrane was muddy yellowish-brown and moderately thickened. The intraluminal contents from the jejunum to the colon were liquid and yellow. Two pigs (pigs Nos. 9 and 10) from farm B had hemorrhagic contents in the small intestine, but erosive and ulcerative lesions were not detected in the mucous membrane. Gross changes were absent in other organs such as mesenteric lymph nodes.
Histologic changes were restricted to the jejunum and ileum, and their degree differed with the case. A great number of Gram-negative bacilli adhered to the apex of villi, in which the enterocytes became degenerative, squamous, and occasionally desquamative. In advanced lesions, there were villous atrophy, mucosal erosion, and hemorrhage (Fig. 1). The lamina propria revealed edema and a neutrophilic infiltration.
Jejunum; pig No. 2. Numerous bacilli (arrows) adhere to the apex of atrophic jejunal villi. HE. Bar = 50µm.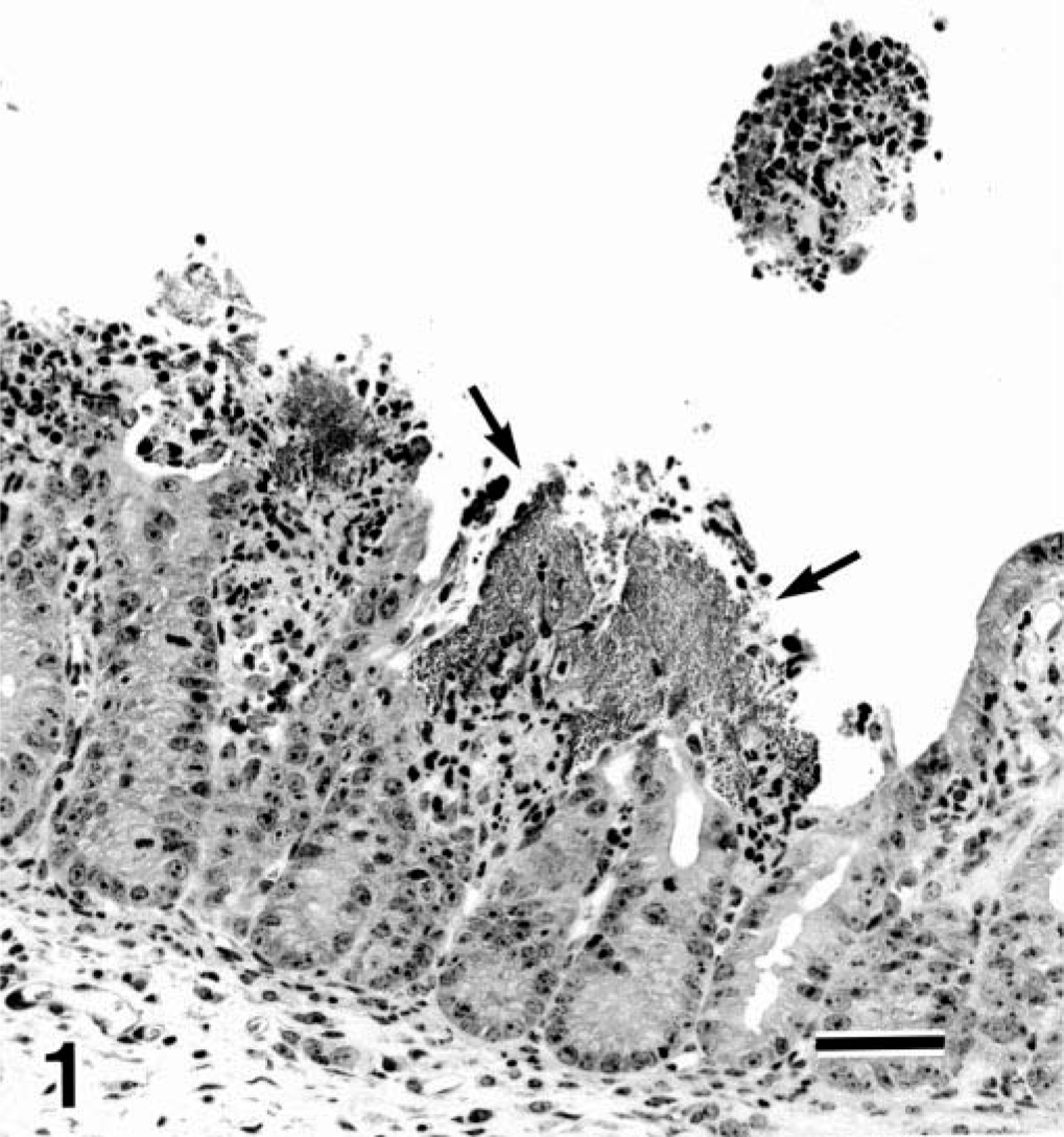
Immunohistochemically, the bacilli were stained positively by the streptavidin–biotin technique (Nichirei Co., Tokyo, Japan) using polyclonal antiserum against serogroup E. coli O18 (Denka Seiken Co.). They were present not only in the intestinal lumen but also in the lamina propria (Fig. 2) and in the cytoplasm of degenerated enterocytes.
Jejunum; pig No. 3. Bacilli, reacting positively with antiserum against E. coli O18, are present within the cytoplasm of enterocytes and in lamina propria (arrows) of the jejunum. Immunohistochemical technique with hematoxylin counterstain. Bar = 20µm.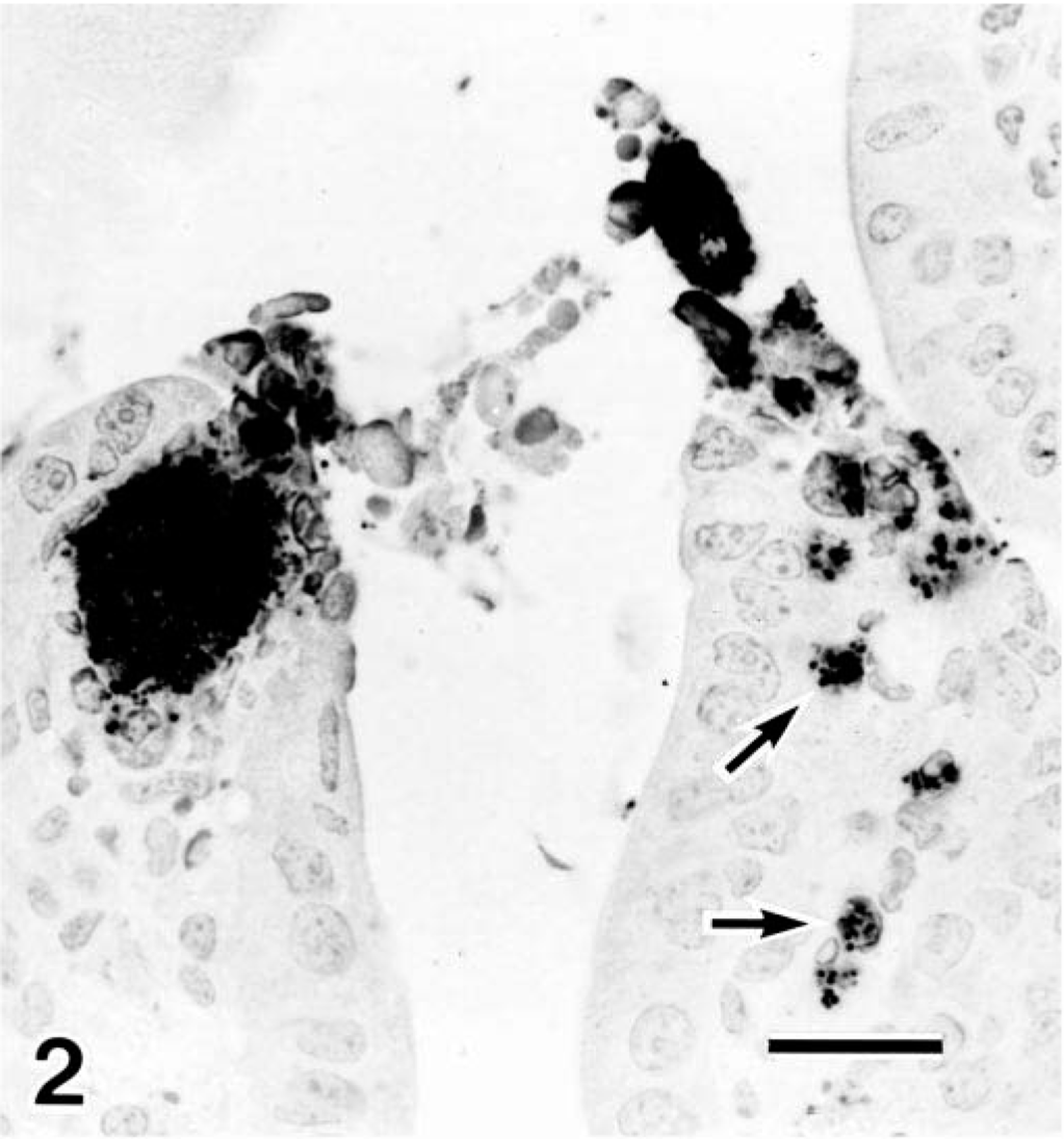
Transmission electron microscopy (TEM) disclosed the presence of many intracytoplasmic bacilli at the apex of villi, and the invaded epithelial cells were degenerated and lacking in microvilli (Fig. 3).
Jejunum; pig No. 3. Transmission electron micrograph of the jejunum. Intracytoplasmic bacilli are visible (arrow). Uranyl acetate and lead citrate. Bar = 4µm.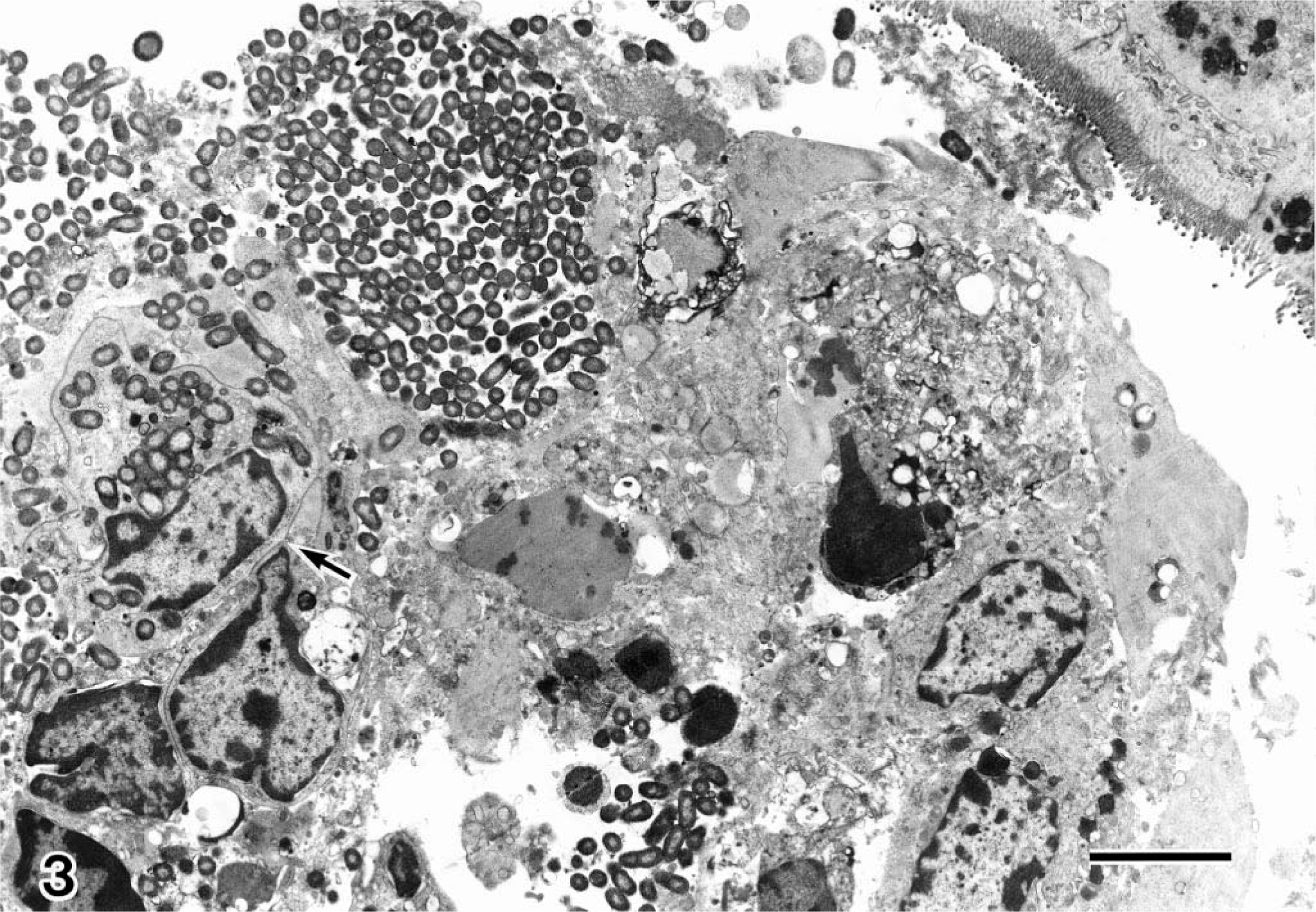
Adhesion assay to human cells (HEp-2 cells) with the E. coli O18:H21 strain was performed as described previously.
11
HEp-2 cells were grown for 48 hours at 37 C in 5% CO2 in Eagle minimum essential medium (MEM) supplemented with 10% fetal calf serum in 75 cm2 tissue culture flasks. After the cells were washed, Eagle MEM supplemented with 2% fetal calf serum and 1%
HEp-2 cells. Transmission electron micrograph of HEp-2 cells infected with E. coli O18:H21 strain. Cytoplasmic projections (arrows) of a HEp-2 cell surround bacilli. Uranyl acetate and lead citrate. Bar = 1 µm.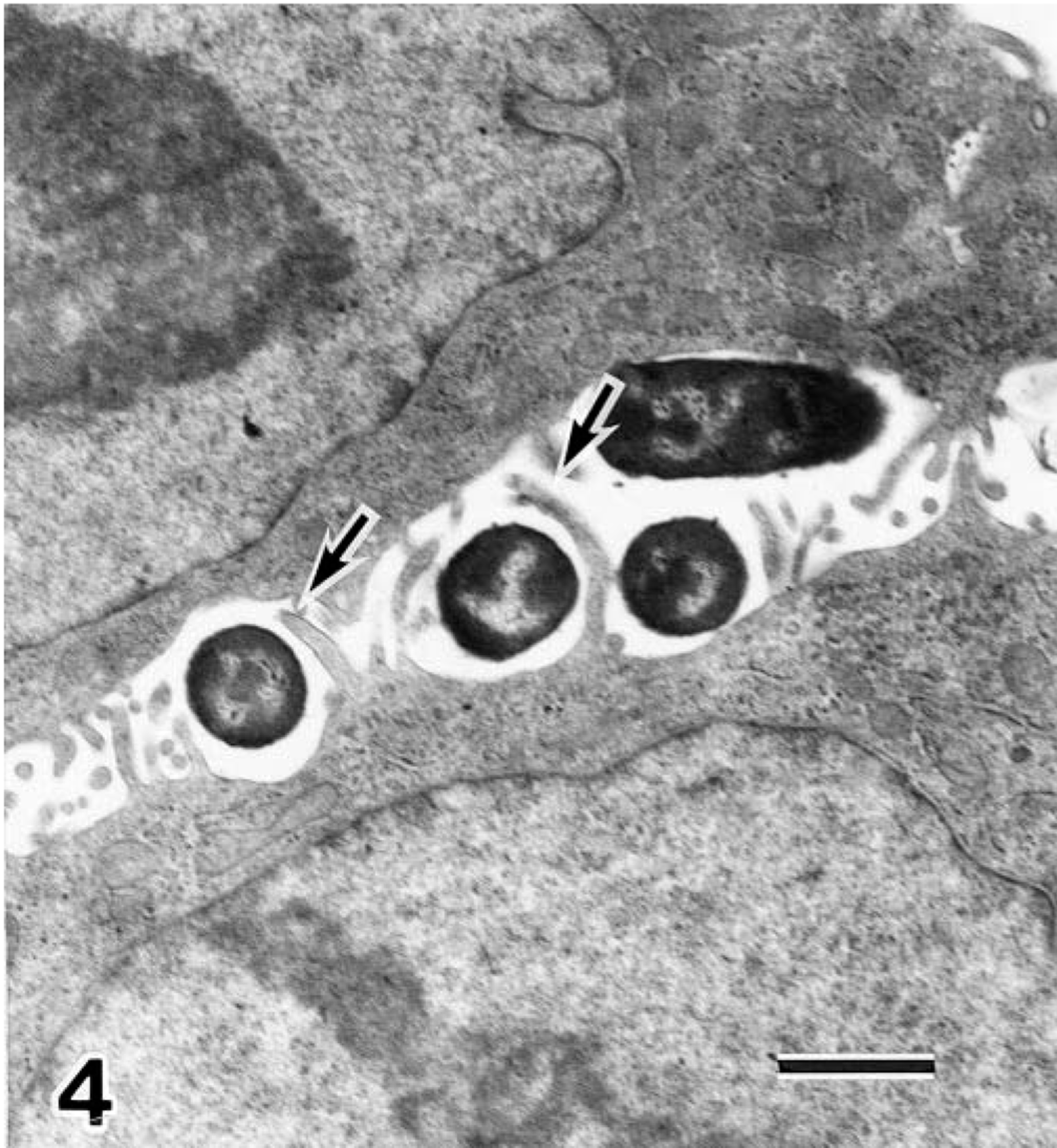
HEp-2 cells. Transmission electron micrograph of HEp-2 cells infected with E. coli O18:H21 strain. Numerous bacilli are located in vacuoles or free in the cytoplasm. Uranyl acetate and lead citrate. Bar = 2 µm.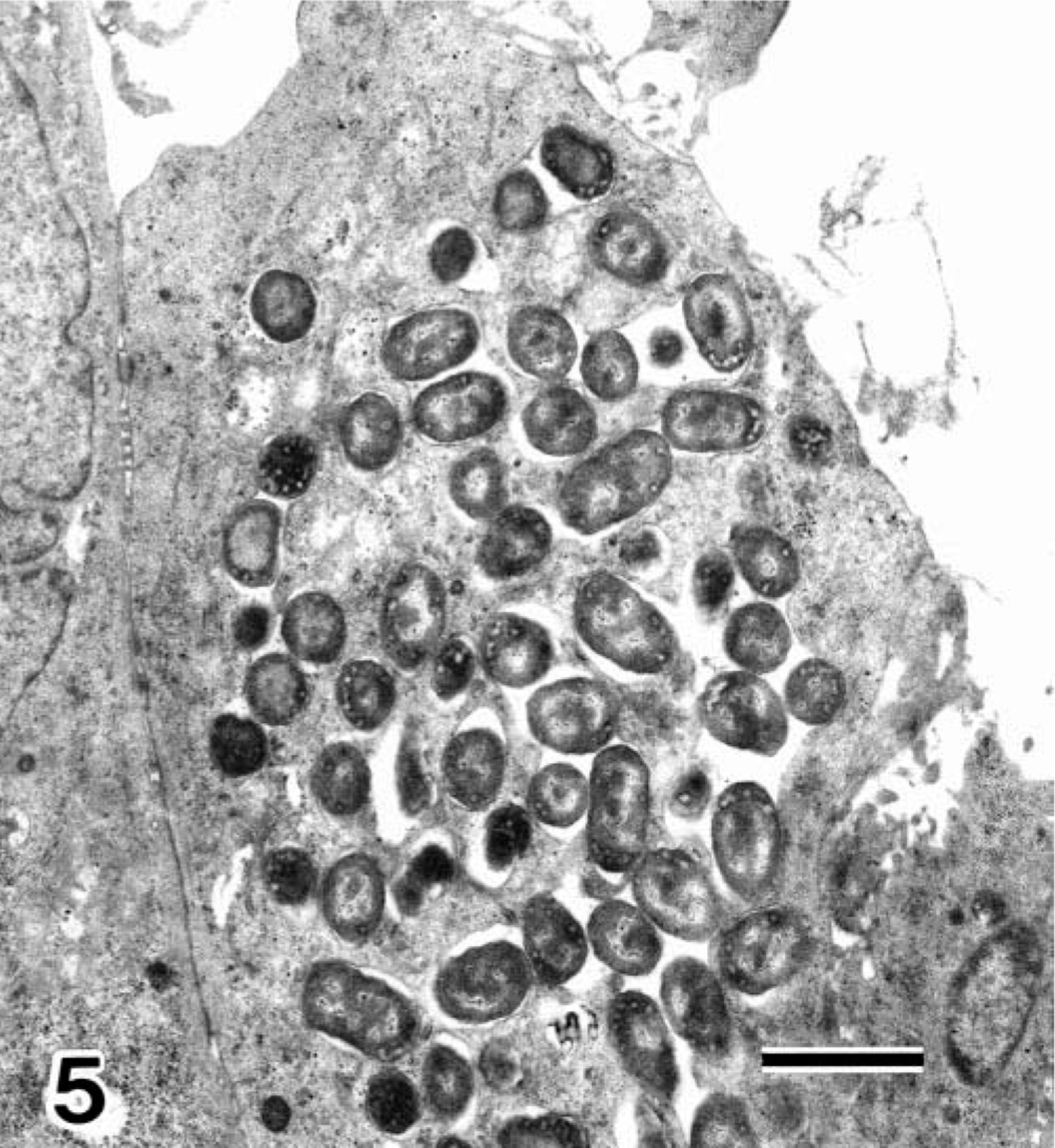
The bacteria described in this report had the ability to invade HEp-2 cells and could be distinguished from HEp-2–adherent E. coli (EAggEC and DAEC) and AEEC, capable of causing attaching and effacing lesions. 6,11,12
Epithelial cell invasion is a key virulence factor for EIEC but is not for other diarrheagenic colibacilli. Expression of the invasive phenotype is positively or negatively regulated by a plasmid or chromosomal gene, and invE enhances expression of some invasion-associated genes in shigellae and colibacilli. 13 The ipaH gene sequence is not an indicator of the invasive phenotype but is specific for shigellae and EIEC. 10 Neither invE nor ipaH were detected in the present bacteria. In addition, they were negative for the Serény test, which is a classical phenotypic assay for EIEC identification. 6 In contrast to O18 in the present bacteria, the serogroup of EIEC is O28ac, O29, O112ac, O124, O136, O143, O144, O152, O159, O164, and O167, and usually H antigens are nonmotile. 6 By accepted means of bacteriologic classification, the E. coli isolates described in this report could not be categorized as EIEC. Moreover, the serotypes of present strains (O18:H16 and O18:H21) has not been reported as those of enteropathogenic strains in human and animals. 6
In humans, a new invasive E. coli strain, LF82 has recently been isolated from a patient with Crohn's disease and was designated as adherent-invasive E. coli (AIEC). 2 PCR and hybridization experiments with LF82 did not reveal the presence of any of the genetic determinants encoding EIEC, EPEC, or ETEC proteins involved in invasiveness. 2 LF82 was engulfed by elongated microvilli of HEp-2 cells and were internalized within endocytotic vacuoles. Similar findings were observed in the present E. coli O18, but the serotype of LF82 was O83:H1. 2 Such elongation of microvillar extensions resembles membrane ruffles in Shigella flexneri or Salmonella typhimurium. 1,4 Like LF82, the present E. coli strains could not be classified bacteriologically into any of the established pathogenic E. coli groups and may be new invasive strains. 2 To determine whether the E. coli O18 strains are truly invasive or not, their invasion-associated genes should be identified.
Footnotes
Acknowledgements
The authors acknowledge Dr. Y. Ando and Mr. T. Fujisawa for preparation of photomicrographs.