
Abstract
Tyrosinase-related protein-2 (TRP-2) is a highly conserved melanogenic enzyme expressed in both pigmented and unpigmented melanomas of the mouse. To determine whether TRP-2 would be a good diagnostic marker for amelanotic melanomas of the dog, we performed immunohistochemistry for TRP-2, S-100, and Melan A on 21 canine tumors identified as amelanotic melanomas based on routine histopathologic examination. Thirteen of the tumors were TRP-2 positive, 10 were Melan A positive, and 19 were S-100 positive. TRP-2 was expressed in the cytoplasm of tumor cells in both primary and metastatic melanomas. S-100 staining was positive in all of three schwannomas and two of three gastrointestinal stromal tumors (one fibrosarcoma and one leiomyosarcoma) tested. Neither Melan A nor TRP-2 antibodies reacted with these tumors. Our findings indicate that staining for TRP-2 is a sensitive and specific method for confirming the diagnosis of amelanotic melanoma in dogs.
Loss of pigmentation in melanomas may be associated with malignancy; 2,7,8 thus, it is important to identify amelanotic melanomas and to distinguish them from other tumor types. Immunohistochemistry for S-100 is widely used to confirm the diagnosis of amelanotic melanoma in humans and animals. 10,12 Although staining for S-100 is a very robust and sensitive technique, the specificity of S-100 staining is low. S-100 antibodies react with a variety of normal tissues and tumors. 4,6,9,10,12 The sensitivity and specificity of other antibodies against melanocytic markers (HMB-45, NKI/C3, HMSA-1 and HMSA-5, and Melan A) have been tested in canine melanomas. Among them, Melan A has proven the most successful, although different investigators have reported varying success using this antibody. 5,10 Because of the low sensitivity of Melan A antibodies and the low specificity of S-100 immunostaining in amelanotic melanomas, it is generally recommended that more than a single immunohistochemical marker be used for diagnosis. 10,11
Several enzymes that participate in melanogenesis, including tyrosinase, tyrosinase-related protein-1 (TRP-1), tyrosinase-related protein-2 (TRP-2), and Pmel17/Silver, are expressed in pigmented melanomas. 7,8,13 Although many of these melanogenic enzymes are absent from amelanotic melanomas, TRP-2 (DOPAchrome tautomerase) appears to be retained in many unpigmented melanomas. 7,8 In fact, recent studies have suggested that TRP-2 may be a good candidate for immunotherapy of amelanotic melanoma. 7,8 In the mouse, TRP-2 is encoded by the slaty locus. The enzyme catalyzes the rearrangement of DOPAchrome to the carboxylated derivative DHICA (5,6-dihydroxyindole-2-carboxylic acid). DHICA, in turn, is used in the synthesis of eumelanin. 13 The purpose of this study was to determine whether immunohistochemistry to detect TRP-2 would be useful in confirming a diagnosis of amelanotic melanoma in the dog in formalin-fixed paraffin-embedded sections and in distinguishing between amelanotic melanomas and other tumor types in this species.
Twenty-one canine cases with a diagnosis of amelanotic melanoma based on examination of hematoxylin and eosin (HE)–stained sections were selected from the archives of the Department of Veterinary Biosciences of The Ohio State University. All cases were from the period 1986–1999. There were 20 primary tumors and 1 metastatic tumor. Tumor sites included oral cavity (15 cases), skin (3 cases), digit (2 cases), and lymph node (1 case). We also examined three schwannomas and three gastrointestinal stromal tumors (GISTs), including one fibrosarcoma, one leiomyoma, and one leiomyosarcoma. All tissues had been fixed in 10% neutral buffered formalin and processed routinely. For the present study, 4-µm sections were prepared from stored paraffin blocks and stained with HE and for S-100, Melan A, and TRP-2 using a standard avidin–biotin complex (ABC) technique.
The anti–TRP-2 antibody used was the generous gift of Dr. Vincent Hearing of the National Cancer Institute, National Institute of Health. 13,14 This polyclonal rabbit antibody was raised against a keyhole limpet hemocyanin–conjugated synthetic 15–amino acid peptide corresponding to the carboxyl terminus of the human TRP-2 protein. 14 Reactivity of the antibody with the immunizing peptide and specificity of the antibody for the TRP-2 protein were confirmed by enzyme-linked immunosorbent assay and western blotting, respectively. 13,14 After sections were deparaffinized and rehydrated, slides were steamed in target retrieval solution (Dako, Carpinteria, CA) for 20 minutes and then cooled in this solution for an additional 20 minutes. Endogenous peroxidase activity was quenched in 3% H2O2 peroxidase block solution (Dako), and a serum-free protein block solution (Dako) was applied to reduce nonspecific staining. Primary antibody (αPEP8h, rabbit anti–TRP-2 antiserum) at a dilution of 1 : 500 was applied to tissues, and they were incubated overnight at 4 C. After rinsing with Tris-buffered saline containing 0.05% Tween20 (TBST), biotinylated goat anti-rabbit IgG antibody (Vector Laboratories, Burlingame, CA) was applied for 30 minutes at room temperature. After TBST rinses, sections were incubated with ABC (Vector Laboratories) for 30 minutes at room temperature and then washed in TBST. Color development using 3,3′-diaminobenzidine (DAB) and counterstaining with hematoxylin were then carried out. For staining with anti–S-100 and anti-Melan A antibodies, antigen retrieval and peroxidase and protein blocking were performed as for anti–TRP-2 staining. Anti–S-100 rabbit polyclonal antibody (Code: Z0311, DAKO) and anti–Melan A mouse monoclonal antibody (Code: M7196, DAKO) were used as primary antibodies, at dilutions of 1 : 1,000 and 1 : 300, respectively. Each primary antibody was applied for a 30-minute incubation period at room temperature. Compatible biotinylated secondary antibodies, ABC, and DAB were applied sequentially. Immunostaining for S-100 and Melan A was performed using the DAKO Autostainer. Negative control sections were prepared using the appropriate normal sera instead of primary antibodies. The intensity of immunostaining was scored as none, weak (less than 30% positive tumor cells), moderate (30–70% positive tumor cells), and strong (70–100% positive tumor cells). Representative immunohistochemical results for two melanomas and one schwannoma are shown in Figs. 1–9.
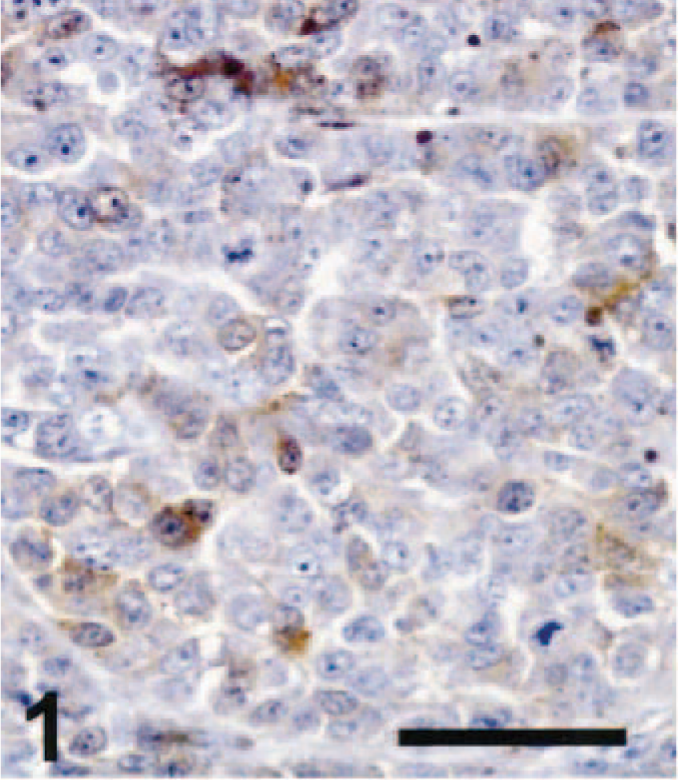
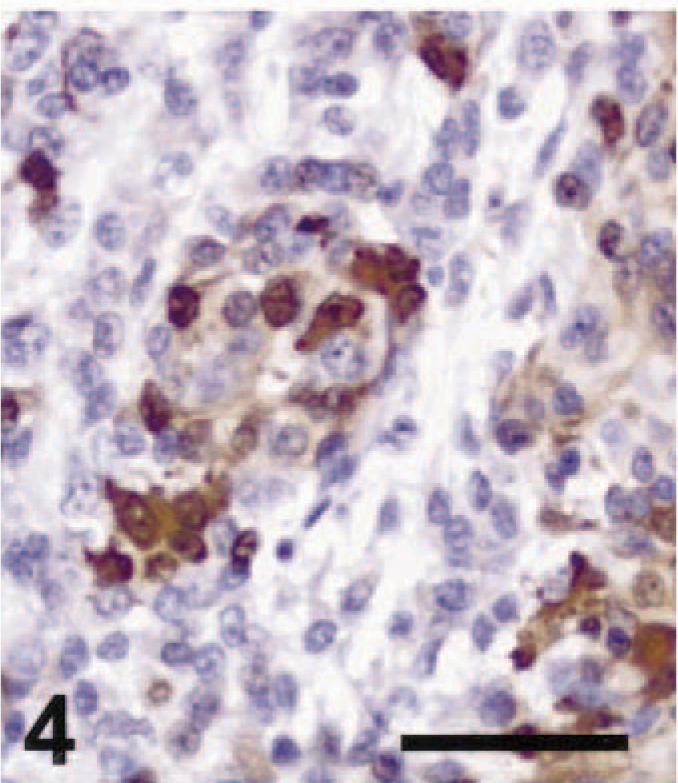
Cutaneous amelanotic melanoma; dog No. 18. Several tumor cells showed weak immunoreactivity for S-100 in this epithelioid amelanotic melanoma. ABC immunoperoxidase method and DAB as chromogen. Hematoxylin counterstain. Bar = 50 µm.
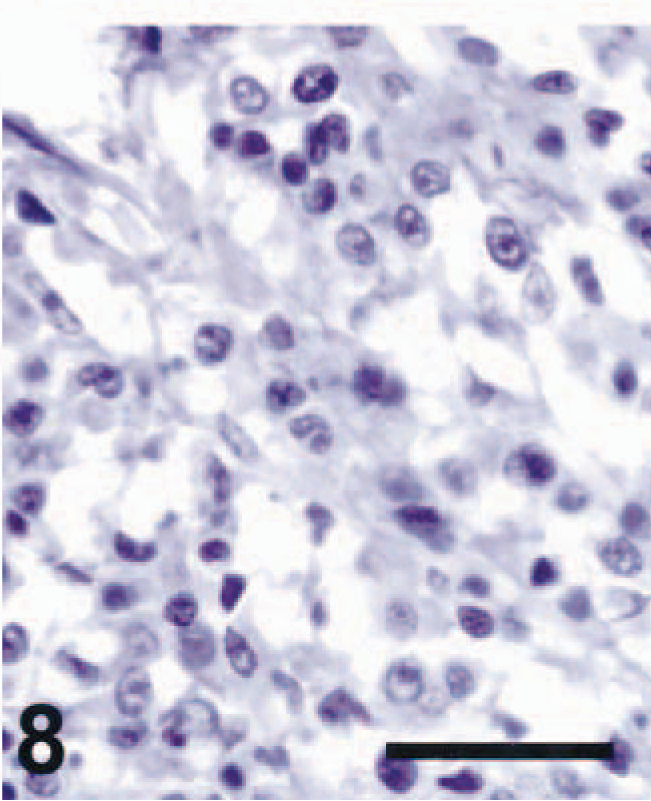
Spinal cord, schwannoma; dog No. 3. TRP-2 immunoreactivity was absent from the tumor. ABC immunoperoxidase method and DAB as chromogen. Hematoxylin counterstain. Bar = 50 µm.
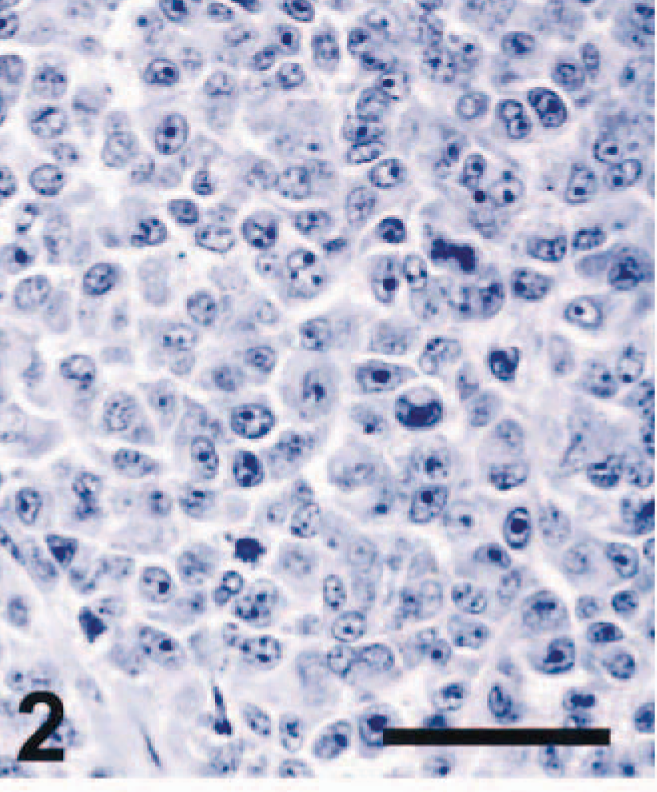
Nineteen of 21 amelanotic melanomas (90.5%) were immunohistochemically positive for S-100 (Table 1). S-100 immunoreactivity was evident mostly in the cytoplasm, but small to moderate numbers of nuclei were also S-100 positive. Four cases were strongly S-100 positive, seven cases were moderately positive (Fig. 4), eight cases were weakly positive (Fig. 1), and two cases were negative. These cases included 18 of 20 primary tumors and the single metastatic tumor examined. Immunohistochemistry for S-100 was positive in tissues from blocks up to 17 years old. Thus, S-100 staining was very sensitive. However, we also detected S-100 in several nonmelanoma tumor types that have been shown to be S-100 positive in other studies. 4,6,9 All three schwannomas we tested were S-100 positive (Fig. 7). The GISTs examined included one tumor classified as a leiomyoma, one as a leiomyosarcoma, and one as a fibrosarcoma. The leiomyosarcoma and the fibrosarcoma were S-100 positive, as indicated by nuclear and cytoplasmic staining of varying intensity in tumor cells. Our results for S-100 immunohistochemistry in melanomas, schwannomas, and GISTs were similar to those reported previously. 4,6,12 In one study, S-100 was detected immunohistochemically in 75% of pigmented melanomas and 90% of amelanotic melanomas of dogs. 5 All seven cell lines established from canine melanomas also expressed S-100 protein. 5 S-100 positivity has been reported in a variety of normal canine tissues (melanocytes and sweat glands of the skin, myoepithelial cells, chondrocytes, glial cells, neurons, Schwann cells, and Langerhans cells), in neural tumors (schwannoma, neurofibroma, and granular cell tumor), and in some GISTs, including fibrosarcomas, leiomyomas, and leiomyosarcomas. 4,6,9,12 A recent article, however, reported that S-100 immunoreactivity could be detected neither in leiomyomas of the gastrointestinal tract nor in a more restricted set of tumors believed to represent GISTs analogous to those in man. 3 Our study confirms previous findings indicating that many schwannomas and GISTs may be S-100 positive; 6,9 taken together, these studies indicate that S-100 positivity is a sensitive, but not specific, marker for amelanotic melanoma in the dog.
Immunostaining results for 21 cases of canine amelanotic melanomas. ∗
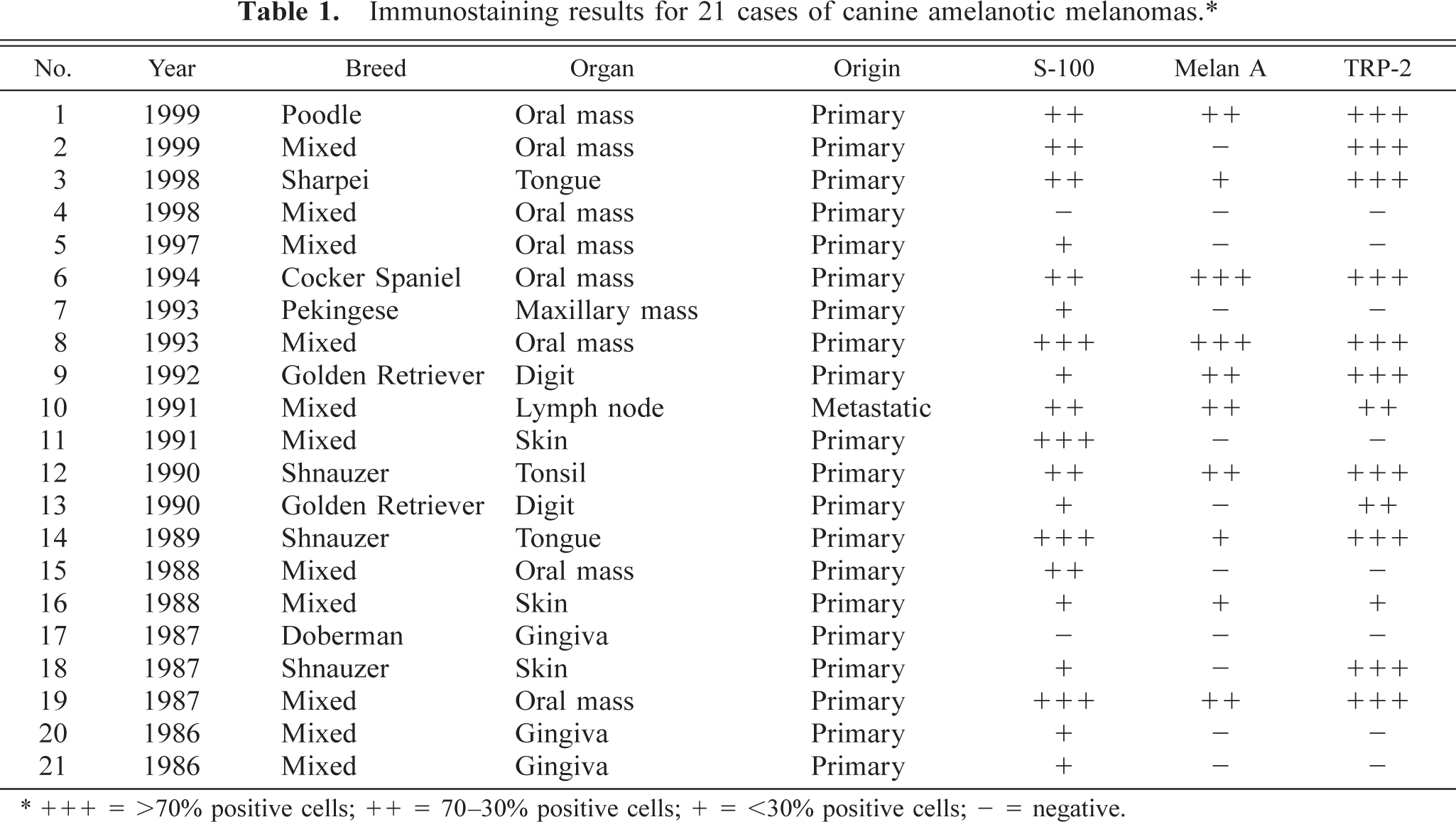
∗ + + + = >70% positive cells; ++ = 70–30% positive cells; + = <30% positive cells; — = negative.
Cutaneous amelanotic melanoma; dog No. 18. No Melan A immunoreactivity was detected. ABC immunoperoxidase method and DAB as chromogen. Hematoxylin counterstain. Bar = 50 µm.
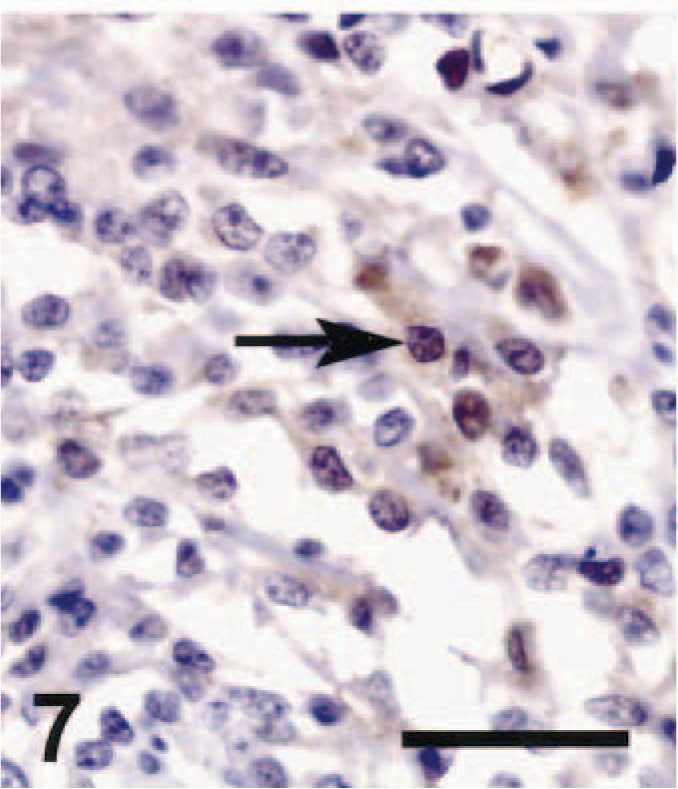
Spinal cord, schwannoma; dog No. 3. The tumor was strongly positive for S-100 immunostaining. The arrow indicates an S-100–positive nucleus. ABC immunoperoxidase method and DAB as chromogen. Hematoxylin counterstain. Bar = 50 µm.
Lymph node, metastatic amelanotic melanoma; dog No. 10. S-100 protein was detected at moderate intensity in the spindle-shaped cells of this amelanotic melanoma. ABC immunoperoxidase method and DAB as chromogen. Hematoxylin counterstain. Bar = 50 µm.
Ten of 21 amelanotic melanomas (47.6%) were positive for Melan A (Table 1). Staining for Melan A had a granular cytoplasmic pattern. Two cases were strongly positive, five cases were moderately positive (Fig. 5), and three cases were weakly positive. Nine of the 20 primary tumors and the single metastatic tumor were Melan A positive. Schwannomas (Fig. 8) and GISTs were not immunoreactive for Melan A (Table 2). Thus, Melan A immunoreactivity was specific for melanomas. However, Melan A was detected in less than half of the amelanotic melanomas, suggesting that it was not a sensitive marker for these tumors. Previous studies examining Melan A expression in canine melanomas demonstrated varying degrees of immunoreactivity for the protein. In one study, 92.6% of primary melanomas and 71.9% of metastatic melanomas in the dog, including amelanotic tumors, were Melan A positive. 10 In contrast, in another study, 90% of pigmented canine melanomas tested were positive for Melan A, but all nine amelanotic tumors tested were negative. 5 These investigators suggested that Melan A expression in canine melanomas was positively correlated with benign tumor behavior. 5 Furthermore, 75% of feline amelanotic melanomas tested were reported to be negative for Melan A, although 80% of pigmented feline melanomas were positive. 11 Amelanotic melanomas in the dog tend to be malignant, 10 thus the small number of Melan A–positive tumors in our study may have reflected the large proportion of malignant tumors in this population of unpigmented tumors.
Immunostaining results of schwannomas and GISTs of dogs. ∗
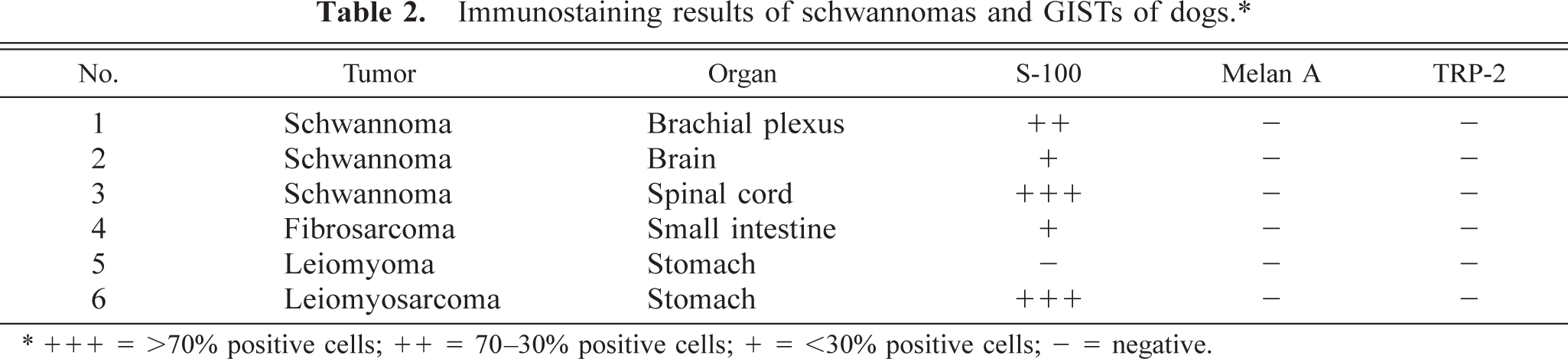
∗ + + + = >70% positive cells; ++ = 70–30% positive cells; + = <30% positive cells; — = negative.
Lymph node, metastatic amelanotic melanoma; dog No. 10. Moderate Melan A staining was apparent. ABC immunoperoxidase method and DAB as chromogen. Hematoxylin counterstain. Bar = 50 µm. Spinal cord, schwannoma; dog No. 3. No Melan A immunoreactivity was seen. ABC immunoperoxidase method and DAB as chromogen. Hematoxylin counterstain. Bar = 50 µm.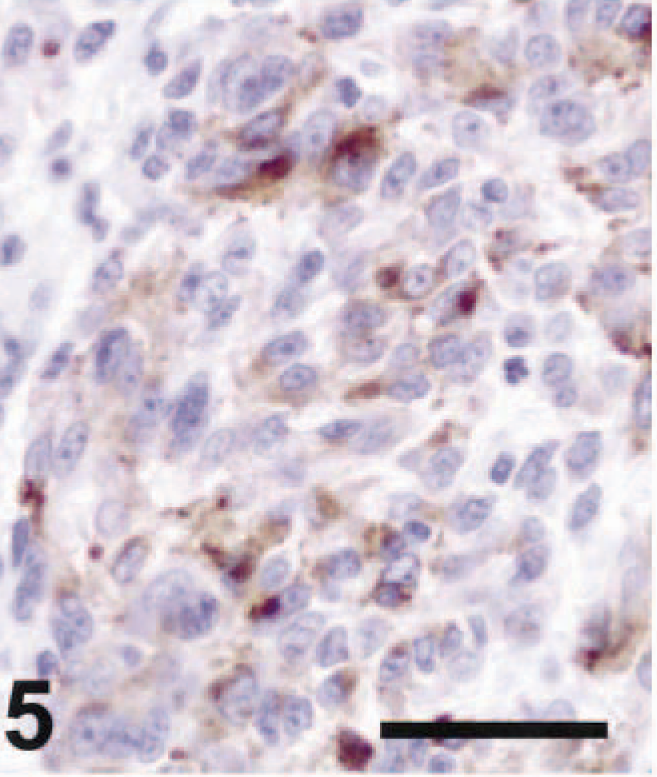
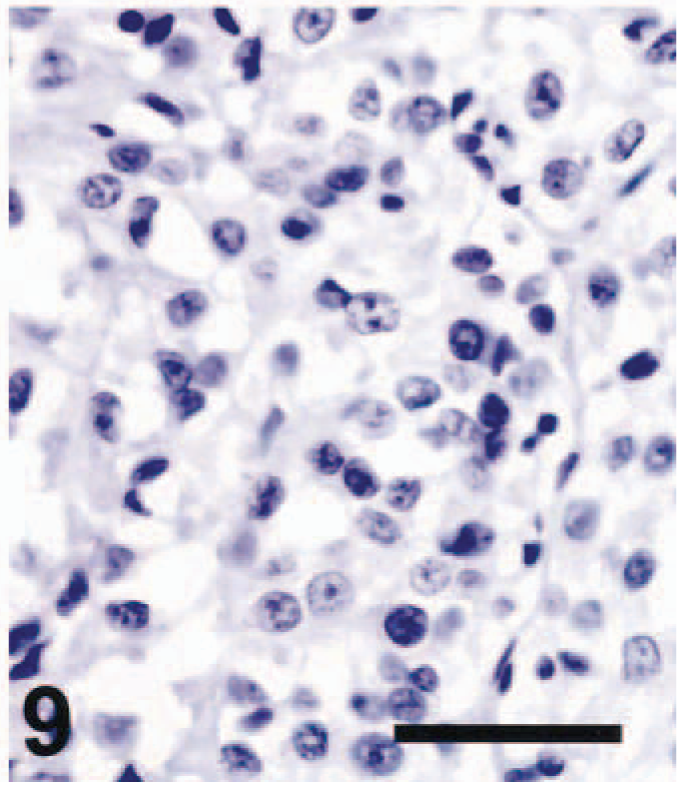
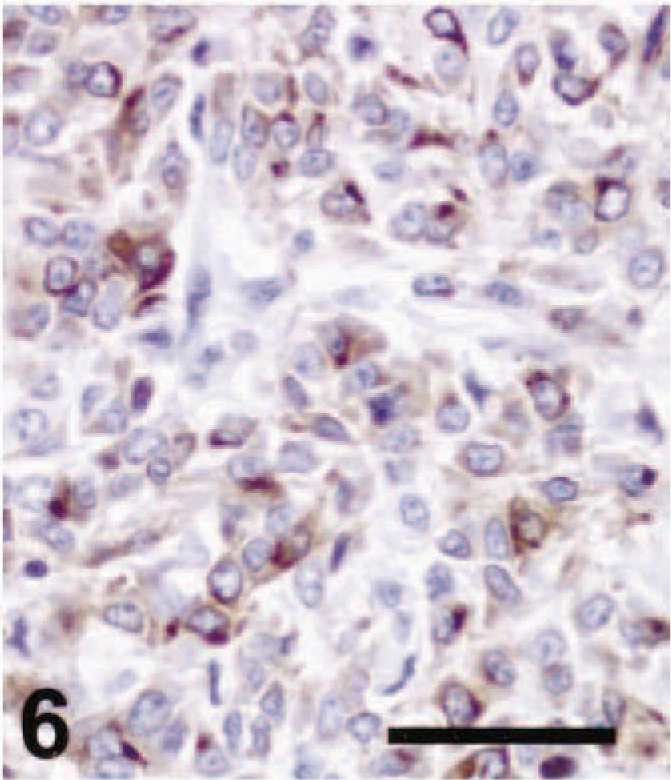
Thirteen of 21 amelanotic melanomas (61.9%) were positive for TRP-2 (Table 1). Staining for TRP-2 was restricted to the cytoplasm. Ten of 13 cases were strongly and diffusely positive for TRP-2 (Fig. 3), two cases were moderately positive (Fig. 6), and one case was weakly positive. Twelve primary tumors and one metastatic mass were TRP-2 positive. Diffuse TRP-2 immunoreactivity was detected in the cytoplasm of tumor cells. None of the nonmelanoma tumors tested stained for TRP-2 (Fig. 9). A recent study showed that pigmentation was partially or completely lost as mouse melanomas underwent malignant transformation. 2,7,8 Expression of the melanogenic enzymes tyrosinase, TRP-1, and Pmel17/silver diminished during melanoma progression, but TRP-2 protein was always detectable, regardless of the stage of malignant progression. 7,8 The presence of TRP-2 in most canine amelanotic melanomas in the present study is thus consistent with the findings of previous studies of mouse melanomas.
Cutaneous amelanotic melanoma; dog No. 18. There was strong diffuse TRP-2 staining in the tumor. ABC immunoperoxidase method and DAB as chromogen. Hematoxylin counterstain. Bar = 50 µm. Lymph node, metastatic amelanotic melanoma; dog No. 10. Moderate immunoreactivity for TRP-2 was present in the cytoplasm of the tumor cells. ABC immunoperoxidase method and DAB as chromogen. Hematoxylin counterstain. Bar = 50 µm.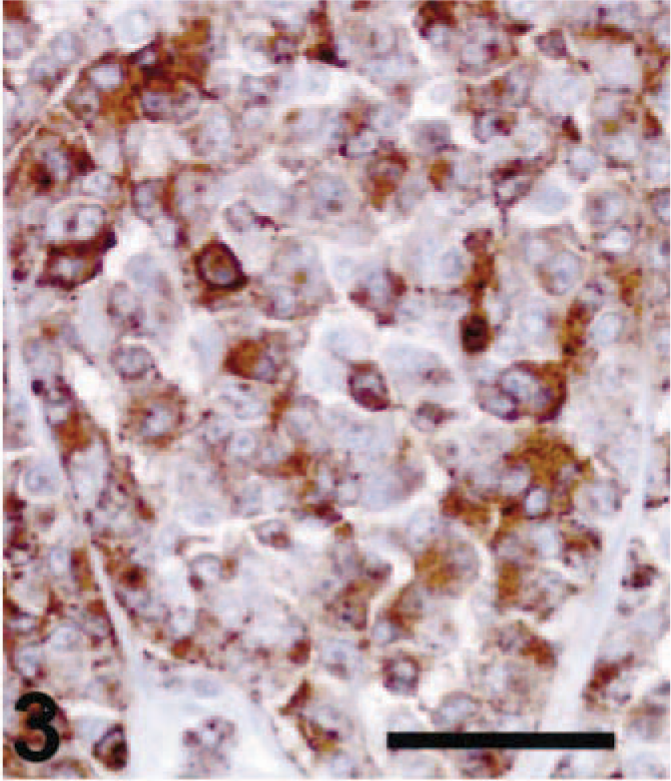
In 8 of 21 tumors classified as amelanotic melanomas (38.1%), we were unable to detect TRP-2. These tumors were also negative for Melan A, and their S-100 immunoreactivity was often weak (four tumors) or absent (two tumors). Most of these specimens were over 10 years old. It is possible that TRP-2 and Melan A antigenicity deteriorated more during storage than S-100 antigenicity, because of increased abundance or stability of S-100 protein compared with Melan A or TRP-2. It is also possible that some of the tumors we examined were not amelanotic melanomas. Two tumors were negative for all three melanoma markers. All Melan A–positive tumors were also TRP-2 positive.
Interestingly, it was recently reported that tyrosinase and Pmel17/silver could be identified immunohistochemically in melanin-containing schwannomas, although nonpigmented schwannomas did not exhibit either antigen. TRP-2 immunoreactivity has not been tested previously in schwannomas. 1 In the present study, we did not detect TRP-2 in three nonpigmented schwannomas (Table 2). Further study will be needed to determine whether TRP-2 is present in pigmented schwannomas.
We have shown that TRP-2 protein was expressed in most canine amelanotic melanomas but not in S-100–positive schwannomas or GISTs. This finding suggests that TRP-2 immunoreactivity may be more specific for melanomas in the dog than S-100 positivity. However, more extensive testing of the reactivity of this new antibody is required to validate its specificity. Based on our study, it appears that S-100 immunoreactivity is more sensitive than TRP-2 immunoreactivity (90.5% positive versus 61.9% positive) in identifying amelanotic melanomas but that TRP-2 immunoreactivity is more specific. TRP-2 staining appears to be somewhat more sensitive than Melan A staining in identifying amelanotic melanomas (61.9% positive versus 47.6% positive). Overall, our findings indicate that immunohistochemical detection of TRP-2 is useful in confirming a diagnosis of canine amelanotic melanoma.
Footnotes
Acknowledgements
This study was supported by the Korea Science and Engineering Foundation (KOSEF) and the Department of Veterinary Biosciences, The Ohio State University. We would like to thank Dr. Vincent Hearing for providing us with the rabbit anti–TRP-2 antiserum, αPEP8h, and Dr. Glenn Merlino for bringing this antibody to our attention.