
Abstract
This report describes erythrocyte biochemical findings in an adult Spanish mustang mare that exhibited persistent methemoglobinemia, eccentrocytosis, and pyknocytosis that were not related to the consumption or administration of an exogenous oxidant. The methemoglobinemia was attributed to a deficiency in cytochrome-b5 reductase (Cb5R) activity, and the eccentrocytes and pyknocytes were attributed to a marked deficiency in reduced nicotinamide adenine dinucleotide phosphate—dependent glutathione reductase (GR) activity that resulted in decreased reduced glutathione concentration within erythrocytes. The GR activity increased to a near-normal value after addition of flavin adenine dinucleotide (FAD) to the enzyme assay, indicating a deficiency of FAD in erythrocytes. The methemoglobinemia, eccentrocytosis, and pyknocytosis were attributed to deficiency of FAD in erythrocytes because the GR and Cb5R enzymes use FAD as a cofactor. This deficiency in FAD results from a defect in erythrocyte riboflavin metabolism, which has not been documented previously in animals.
Keywords
Circulating erythrocytes are exposed to a variety of endogenously generated oxidants, including superoxide (O2 −), hydrogen peroxide, the hydroxyl radical (OH·), and various reactive nitrogen species. 18,28 Increased amounts of endogenous oxidants are generated in association with various disorders, including inflammation, 41,45,46 neoplasia, 7,10 and diabetes. 8 The damage that results from these oxidants may play an important role in natural aging and the removal of erythrocytes from the circulation by mononuclear phagocytes. 13
Oxidants damage erythrocytes in various ways. Methemoglobin is formed when the iron moiety of heme groups in hemoglobin is oxidized from the ferrous (Fe2+) to the ferric (Fe3+) state. Methemoglobin does not have the ability to bind oxygen, but this form of oxidant injury is reversible. Methemoglobin is reduced back to functional hemoglobin primarily by cytochrome-b5 reductase (Cb5R), also called reduced nicotinamide adenine dinucleotide (NADH)–dependent methemoglobin reductase, which requires flavin adenine dinucleotide (FAD) as a cofactor (Fig. 1). 18 This reduction of methemoglobin is corrective rather than protective.
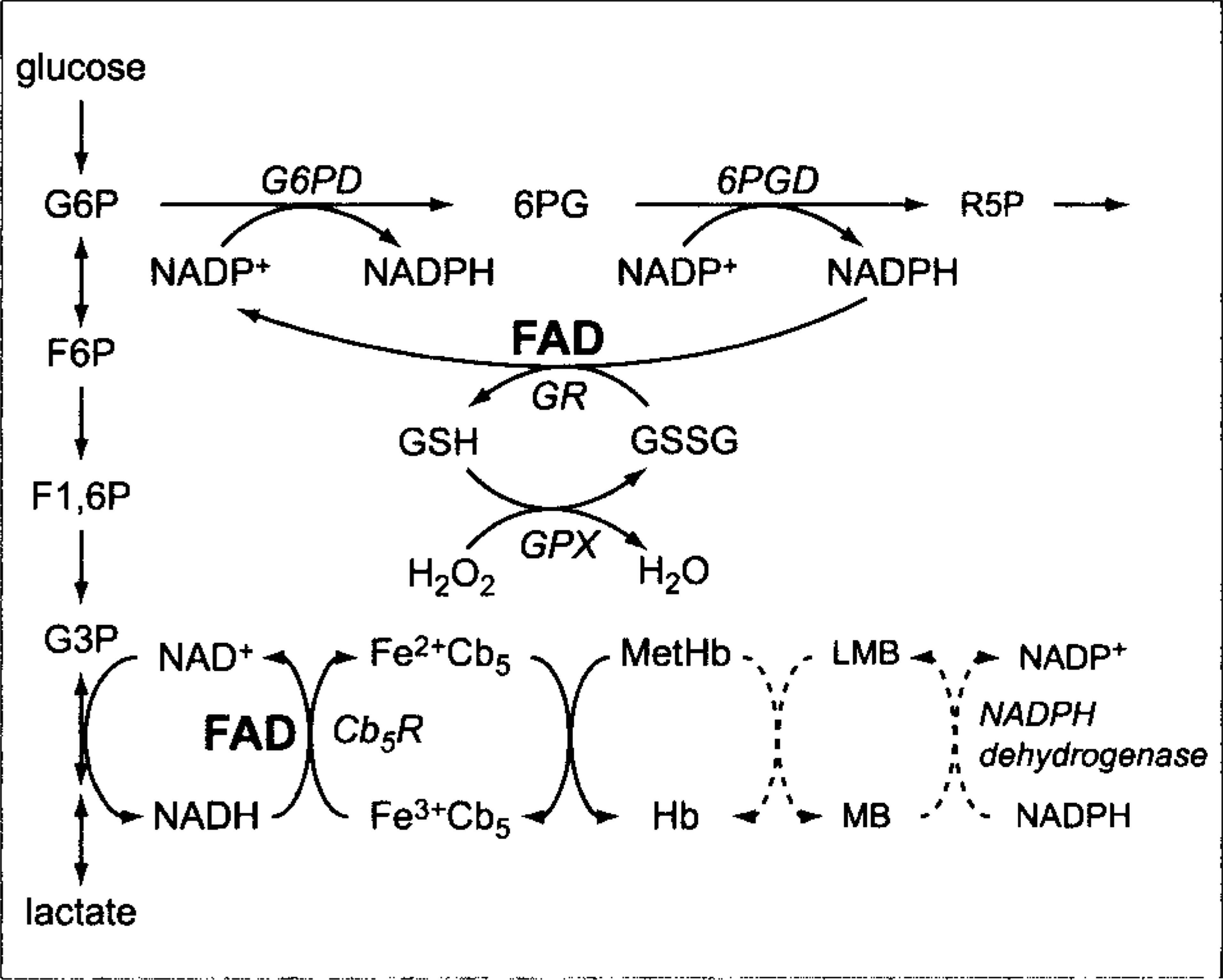
Flavin adenine dinucleotide roles in reductive pathways of erythrocytes. Glycolytic and pentose phosphate pathways are abbreviated. 6PGD, 6-phosphogluconate dehydrogenase; Cb5R, cytochrome-b5 reductase; F1,6P, fructose 1,6-diphosphate; F6P, fructose 6-phosphate; Fe2+Cb5, ferrocytochrome-b5; Fe3+Cb5, ferricytochrome-b5; FAD, flavin adenine dinucleotide; G3P, glyceraldehyde 3-phosphate; G6P, glucose 6-phosphate; G6PD, glucose 6-phosphate dehydrogenase; GPX, glutathione peroxidase; GR, glutathione reductase; GSH, reduced glutathione; GSSG, oxidized glutathione; H2O2, hydrogen peroxide; Hb, hemoglobin; MB, methylene blue; MetHb, methemoglobin; NAD+, nicotinamide adenine dinucleotide; NADH, reduced nicotinamide adenine dinucleotide; NADP+, nicotinamide adenine dinucleotide phosphate; NADPH, reduced nicotinamide adenine dinucleotide phosphate; and R5P, ribose 5-phosphate.
Sulfhydryl groups in proteins and unsaturated lipids in membranes are especially susceptible to oxidation. Oxidative denaturation and the precipitation of the globin portion of hemoglobin into large aggregates result in the formation of Heinz bodies that can bind to and alter membranes. Membrane structure also is altered by the oxidation of sulfhydryl groups and by lipid peroxidation. 18 Oxidative injury to erythrocyte membranes may result in the formation of eccentrocytes and pyknocytes. 14,15
Reduced glutathione (GSH) is a tripeptide of glutamic acid, cysteine, and glycine that is synthesized de novo from constituent amino acids through two enzymatic reactions requiring adenosine triphosphate (ATP). GSH has a highly reactive (easily oxidizable) sulfhydryl group that protects against oxidant injury by nonenzymatic and enzymatic mechanisms. GSH is constantly being oxidized to a disulfide (oxidized glutathione [GSSG]) in response to oxidative reactions in the body. The GSSG formed is reduced to GSH by reduced nicotinamide adenine dinucleotide phosphate (NADPH) and the FAD-dependent glutathione reductase (GR) reaction (Fig. 1). NADPH is produced by the initial enzyme reactions of the pentose phosphate pathway, and erythrocytes increase pentose phosphate–pathway metabolism in response to oxidants to provide the NADPH necessary for the regeneration of GSH. In healthy humans and animals, most glutathione in erythrocytes is maintained as GSH, with low concentrations of GSSG being present. 18
Erythrocytes from horses are slower than erythrocytes from other species studied in their ability to regenerate GSH after it has been oxidized in vitro. 1,31 Horse erythrocytes also appear less able to protect themselves against oxidative injury induced by incubation with high levels of ascorbate, which stimulates the GR reaction by the oxidation of GSH. 22 These reduced abilities may be related to the fact that horse erythrocytes have lower GR activities than erythrocytes from humans and most domestic animal species, and the Michaelis-Menton constant (Km) of GSSG for GR is higher in horses than in three other species measured. 21
This report describes erythrocyte biochemical findings in an adult horse that exhibited persistent methemoglobinemia, eccentrocytosis, and pyknocytosis that was not related to the consumption or administration of an exogenous oxidant. Eccentrocytosis and pyknocytosis have previously been recognized in a horse with erythrocyte glucose-6-phosphate dehydrogenase (G6PD) deficiency, but methemoglobinemia was not present in that animal. 36,43 The methemoglobinemia in the present horse was attributed to a deficiency in Cb5R activity, and the eccentrocytes and pyknocytes were attributed to a marked deficiency in GR activity. Both enzymes use FAD as a cofactor (Fig. 1), and erythrocytes from the affected horse were deficient in FAD. This deficiency in FAD results from a defect in erythrocyte riboflavin metabolism, which has not been documented previously in animals.
A Spanish mustang mare of about 20 years of age was presented to the University of Missouri–Columbia, College of Veterinary Medicine, with a persistent mucopurulent nasal discharge and an inflamed epiglottis associated with empyema in the left guttural pouch. Abnormal findings in a routine complete blood count (CBC) included mild neutropenia without left shift and poikilocytosis, consisting of moderate eccentrocytosis, moderate pyknocytosis, and low numbers of erythrocytes containing hemoglobin crystals that were often eccentrically located in the cell (Fig. 2A–C). The hematocrit was 33% (reference interval, 32–53%), and erythrocyte indices were within reference intervals. Neither Heinz bodies nor reticulocytes were seen by using a new methylene blue (NMB) reticulocyte stain. An absence of blood reticulocytes is expected in horses, whether or not anemia is present. Hemoglobin crystals did not stain blue with NMB, but a thin rim of intense blue staining was often apparent on the periphery (Fig. 2D). Pyknocytes and eccentrocytes were obvious, staining darker than other erythrocytes with NMB (Fig. 2E). Erythrocytes from a horse with suspected spherocytic hemolytic anemia were assessed the same day and did not stain dark like the mustang's eccentrocytes and pyknocytes (Fig. 2F). The chondroids were not surgically removed because of the unexplained hemic abnormalities and the client's reluctance to agree to surgery.
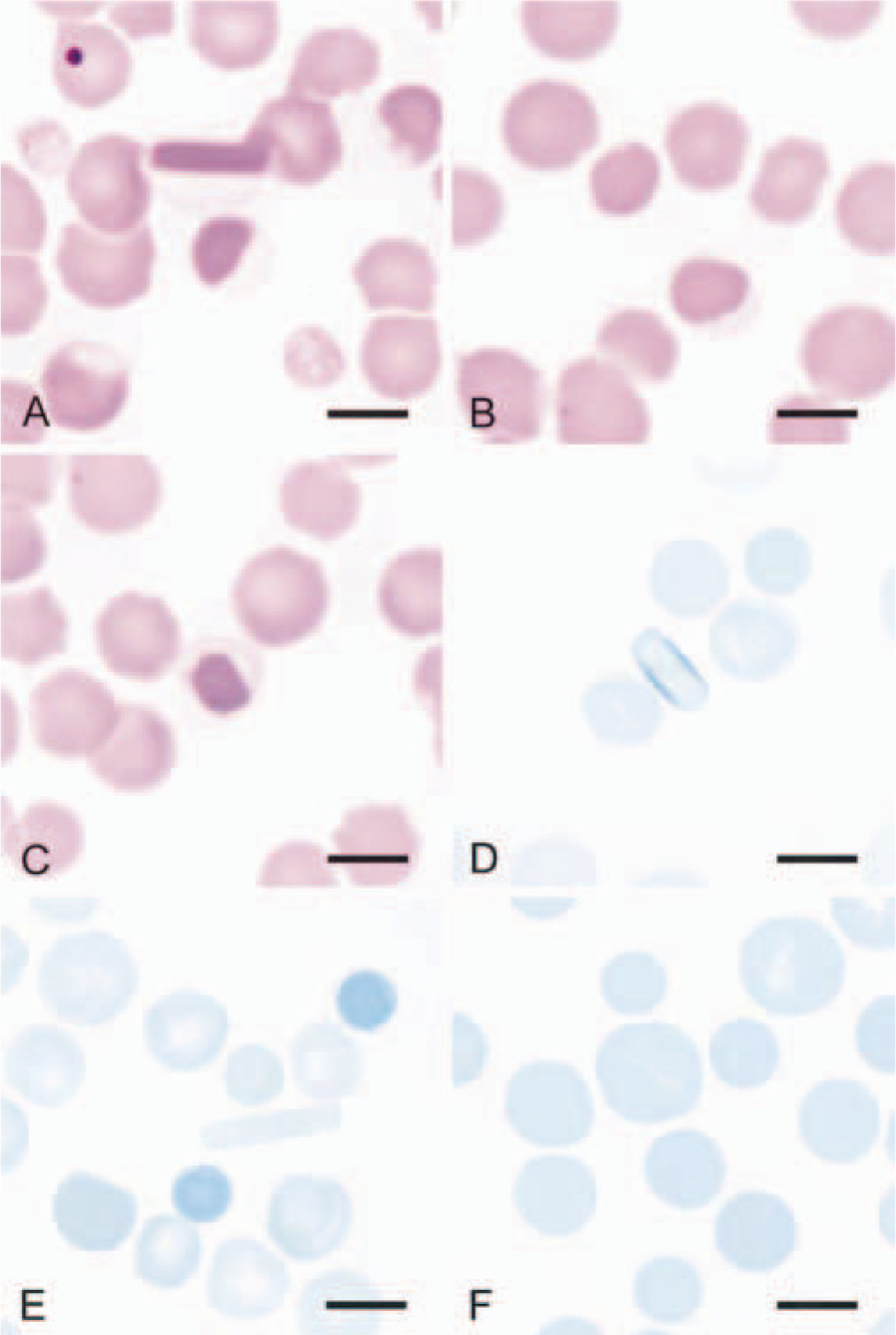
Erythrocytes from blood films of the affected mustang and (
An intermittent mucopurulent nasal discharge persisted, and the horse was evaluated 8 months after initial presentation. Guttural pouch cultures were positive for Streptococcus equi. Surgical drainage was recommended, but the client elected to try an indwelling catheter for irrigation of the guttural pouch with sterile saline. Abnormal findings in the CBC again included mild neutropenia and similar poikilocytosis. A bone marrow aspirate smear contained hypercellular particles with a myeloid : erythroid ratio of 0.3 (reference interval, 0.5–1.5), indicating that erythroid hyperplasia was present.
Because of the persistence of poikilocytes and the lack of an identifiable source of oxidants in the diet, a defect in erythrocyte metabolism protecting against endogenous oxidants was considered. Consequently, blood was drawn to measure GSH and enzymes involved in the protection of erythrocytes against oxidants. Venous blood appeared brownish, suggesting that methemoglobinemia was also present. A spot test confirmed the presence of methemoglobinemia, 19 prompting the addition of methemoglobin and Cb5R to the assays being planned. Findings from the erythrocyte assays are presented in the Results section.
The horse was examined again 10 months after initial presentation. Clinical findings were similar, and the poikilocytosis persisted, but the horse was no longer neutropenic, and the hematocrit was 27%. The horse was donated to the University of Missouri 24 months after initial examination so that additional research studies could be performed in accordance with university animal use guidelines. At the time of donation, the clinical findings and poikilocytosis persisted, and the absolute neutrophil count remained within the reference interval, but the hematocrit (35%) was higher and within the reference interval. Clinical chemistry profiles from the initial examination, 10 months later, and 24 months later were within reference intervals, except for isolated mild hyperglycemia and hyperchloremia and persistent moderate hyperproteinemia and hyperglobulinemia, characterized as polyclonal gammopathy by serum protein electrophoresis. The dysproteinemia was attributed to persistent guttural pouch infection.
Materials and Methods
Blood sample collection and handling
Blood samples for quantitative erythrocyte biochemical assays were collected in potassium ethylenediaminetetraacetic acid (K3EDTA)–containing tubes, kept at around 4 C, and shipped the same day by overnight carrier to the University of Florida for analysis. Blood extracts for total nicotinamide adenine dinucleotide phosphate (NADP) (NADP+ + NADPH) and NADPH assays 50 were prepared rapidly after collection, immediately frozen (−70 C), and shipped with dry ice by overnight carrier to the University of Florida, where they were kept frozen at −70 C until assayed. Blood samples from healthy horses were collected, shipped, and analyzed exactly as the mustang's samples were each time that samples from the mustang were analyzed. These samples served as control samples for processing and handling. Control horses used were adult male and female hot-blooded horses of various breeds. The number of control horses used varied according to the assay being performed; consequently, numbers are given in appropriate places in the Results section and in the tables.
Electron microscopy
Erythrocytes in K3EDTA-anticoagulated blood were fixed with 2% paraformaldehyde-2% glutaraldehyde in 0.1 M cacodylate buffer, pH 7.35, for 2 hours at 4 C. After primary fixation, specimens were washed thoroughly with buffer and postfixed with aqueous 1% osmium tetroxide and 1% sucrose for 1 hour at 4 C. For scanning electron microscopy samples, erythrocytes were dehydrated in a graded ethanol series and critical point–dried. For transmission electron microscopy, samples were en bloc stained with aqueous 1% uranyl acetate for 1 hour, dehydrated in a graded ethanol series, and embedded in resin.
Erythrocyte biochemical assays
Erythrocyte G6PD, 6-phosphogluconate dehydrogenase (6PGD), GR with and without 1 µM FAD added, glutathione peroxidase (GPX), catalase, NADPH dehydrogenase, hexokinase (HK), and pyruvate kinase (PK) activities were measured spectrophotometrically (DU-64 Spectrophotometer, Beckman Instruments, Inc., Fullerton, CA) at 37 C in reaction mixtures containing hemolysates prepared after the removal of leukocytes by filtration through microcrystalline cellulose and alpha-cellulose columns. 5 Cb5R was measured as NADH-ferricyanide reductase at 30 C. 5,6 Methemoglobin percentage 24 and GSH, 39 total NADP, and NADPH 44 concentrations were measured spectrophotometrically using established methods.
Methylene blue injections
To determine the presence or absence of an abnormal hemoglobin (hemoglobin M) that might be spontaneously oxidized to methemoglobin and to confirm the presence of a functional NADPH dehydrogenase, the mustang was injected intravenously with methylene blue (8.8 mg/kg body weight) as a 3% (wt/vol) solution. Methemoglobin percentage was determined before injection of methylene blue and 0.5 and 2 hours after injection.
Dietary riboflavin studies
Dietary riboflavin deficiencies in humans and animals result in low GR activity secondary to low FAD concentrations. 12,29 Despite the lack of clinical signs typical of systemic riboflavin deficiency, the horse was treated with riboflavin (100 mg/day) through daily 20 ml intramuscular injections of a B-complex vitamin supplement (Super-B Complex, Vedco, St. Joseph, MO) for 24 days. Microscopic appearance of erythrocytes, erythrocyte GR (with and without FAD added) and Cb5R activities, methemoglobin percentage, and erythrocyte GSH concentration were evaluated immediately before and after the treatment period.
Neutrophil isolation and enzyme assays
Blood for neutrophil isolation was drawn into sterile containers with acid-citrate-dextrose. The whole blood was allowed to stand undisturbed for 30 minutes on the bench top. The plasma was harvested and mixed with an equal volume of a solution containing 6% dextran (Sigma T500) and placed on the bench top and allowed to stand for an additional 45 minutes at room temperature. The supernatant was collected and centrifuged at 250 × g for 10 minutes at 4 C. The cell pellet was resuspended in 2 ml of a lysis solution consisting of 0.15 M NH4Cl, 10 mM NaHCO3, and 0.1 M EDTA for 10 minutes on ice. At the end of the lysis procedure, the lysis buffer-cell mixture was diluted with an equal volume of phosphate-buffered saline (PBS) to which 2% glucose was added. The cells were pelleted at 400 × g for 10 minutes, resuspended in PBS–2% glucose, and the wash cycle repeated a second time. Neutrophil purity was verified by examination of Wright-stained cytospin preparations. Neutrophil suspensions were kept at around 4 C and shipped the same day by overnight carrier to the University of Florida for enzyme analysis. Neutrophils in suspension were counted, and samples were sonicated, frozen in dry ice and acetone, and thawed to room temperature in a water bath. Enzyme assay conditions for GR (with and without FAD addition) and Cb5R were the same as for erythrocyte assays.
Flavin assays in erythrocytes and plasma
Blood for flavin assays was collected in K3EDTA-containing tubes, protected from light during transit to the laboratory, and immediately processed in dim light. Blood was centrifuged to separate cells from plasma. Plasma was removed from the tube, placed in polypropylene tubes, and frozen (−70 C). The remaining plasma, buffy coat, and a thin layer of erythrocytes were removed and discarded. Remaining erythrocytes were washed three times in chilled isotonic saline. After the third wash, packed erythrocytes were placed in polypropylene tubes and frozen (−70 C). Frozen aliquots were shipped on dry ice to the NIH Clinical Nutrition Research Unit, University of California, Davis, California.
Flavin (riboflavin, flavin mononucleotide [FMN], and FAD) concentrations were measured with a Hewlett Packard series 1100 high-pressure liquid chromatograph (HPLC) and a 1046A fluorescence detector. 16 Samples were prepared using a modified form of the method of Zempleni et al. 49
Results
Eccentrocytosis, pyknocytosis, and hemoglobin crystals persisted in stained blood over the 32 months that the mustang was studied. Scanning and transmission electron microscopy verified the presence of eccentrocytes, pyknocytes, and hemoglobin crystals (Fig. 3A–F). The eccentric placement of hemoglobin crystals noted by light microscopy was confirmed by scanning electron microscopy (Fig. 3E). The hematocrit ranged from 27% to 38% (mean 32%), with 3 of the 10 values below the reference interval of 32–53%. The mean cell volume and mean cell hemoglobin concentration values were consistently within reference intervals over the time period.
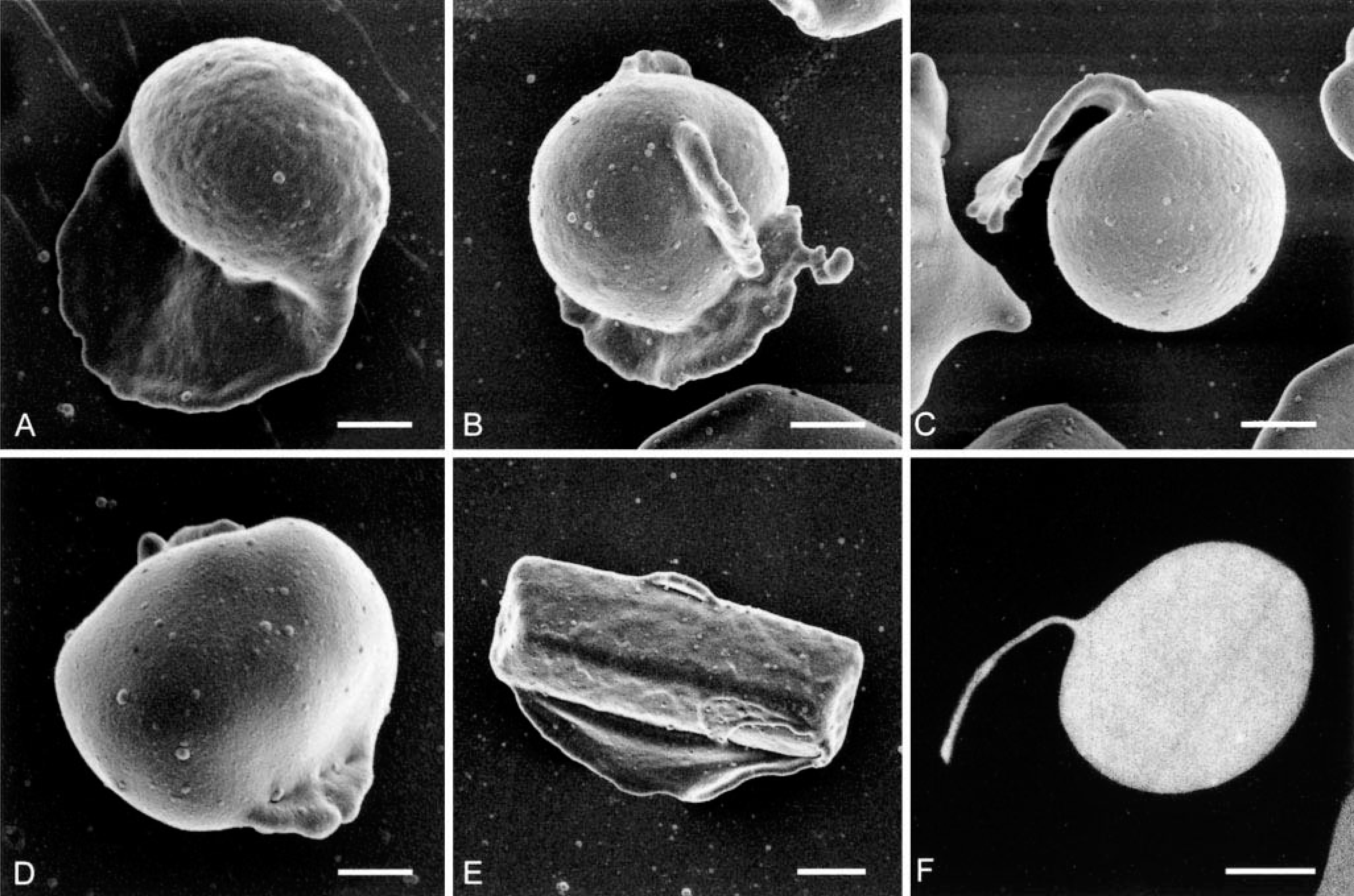
Scanning electron micrographs and transmission electron micrograph
Erythrocyte G6PD activity was about 70% higher than that measured in two healthy control horses and above the reference interval established previously at the University of Florida (Table 1). The mustang's PK activity was also above the reference interval, and HK activity was at the upper limit of the reference interval. Results of erythrocyte 6PGD and catalase assays were within the respective reference intervals (Table 1). GPX activity was slightly below the reference interval, but so was the value from one of the two healthy control horses. GSH concentration was low. Erythrocyte GR activity was not detectable in the mustang unless the cofactor FAD was added to the assay, indicating that a severe erythrocyte FAD deficiency was present. The GR activity increased markedly after 1 µM FAD addition to the assay from the mustang but remained below the reference interval. The GR activity increased by about 70% after 1 µM FAD addition to assays from healthy horses. The addition of 1, 5, and 10 µM FMN to assays from healthy horses had no effect on GR activity (data not shown).
Results of erythrocyte biochemical assays from a mustang horse with erythrocyte FAD deficiency and from clinically healthy horses of various breeds. ∗

∗ Reported values are medians followed by minimum and maximum values in parentheses. Superscript numbers represent the number of times the mustang's samples were assayed (first column of data) or the number of samples assayed from other individuals (last two columns of data). Individual values are given if the number of samples was less than three. For the first five analytes, samples from the mustang and accompanying healthy control horses were assayed five times over 17 months. Blood samples from healthy control horses were collected and shipped by overnight express along with the mustang's blood samples. Blood for the reference intervals was collected from University of Florida horses and assayed on the day of collection. Reference intervals represent the lowest and highest values measured.
† FAD = flavin adenine dinucleotide; GR = glutathione reductase; Hb = hemoglobin; ND = not detectable; GSH = reduced glutathione; Cb5R = cytochrome-b5 reductase; HK = hexokinase; PK = pyruvate kinase; G6PD = glucose-6-phosphate dehydrogenase; 6PGD = 6-phosphogluconate dehydrogenase; GPX = glutathione peroxidase.
Methemoglobin percentage was markedly increased in the mustang, and Cb5R activity was below that of the controls and the reference interval (Table 1). Cb5R activity was not altered by the addition of 1 µM FAD to assays from the mustang and two healthy horses (data not shown). Abnormal findings in erythrocyte GSH concentration, methemoglobin percentage, GR activity, and Cb5R activity were consistently present when measured on five different occasions over a period of 17 months. A reference interval was not established for erythrocyte NADPH dehydrogenase activity in horses, but the median activity (0.5 U/g Hb) of three assays from the mustang was similar to the median assay (0.4 U/g Hb) measured in erythrocytes from three healthy control horses. The total erythrocyte NADP concentration was 64 µmol/liter in the mustang, similar to 67 µmol/liter and 51 µmol/liter in two healthy control horses. In separate assays, erythrocyte NADPH concentration values for the mustang and healthy control horses were somewhat higher than the respective total erythrocyte NADP concentration values, indicating that essentially all the NADP was in the form of NADPH.
Methemoglobin percentage in the mustang dropped from 26% of total hemoglobin concentration before intravenous methylene blue injection to 3% and 2% of total hemoglobin concentration at 0.5 and 2 hours after injection, respectively.
Riboflavin was administered intramuscularly on a daily basis to the mustang, in the event that the horse had a defect in intestinal riboflavin absorption. The 24-day treatment with riboflavin (100 mg/day) had no apparent effect on erythrocyte GR activity (with or without FAD added), Cb5R activity, GSH concentration, or blood methemoglobin percentage (data not shown).
In contrast to results found in erythrocytes, the Cb5R activity in mustang neutrophils was higher than the activity measured in neutrophils from a healthy horse, but without an established reference interval, it is not possible to determine whether the mustang's neutrophil Cb5R activity was increased above normal. The neutrophil GR activity of the mustang was essentially the same as that found in a healthy control horse (Table 2).
Glutathione reductase (GR) and cytochrome-b5 reductase (Cb5R) activities in neutrophils from a mustang horse with erythrocyte flavin adenine dinucleotide (FAD) deficiency and from a clinically healthy control horse. ∗

∗ Reported values represent the results of duplicate assays done on consecutive days, beginning each day with the centrifugation of samples to concentrate intact cells before sonication and freezing.
Results of the flavin assays determined by HPLC are given in Table 3. FMN and FAD concentrations were too low to be accurately quantified individually, necessitating the measurement of their combined concentrations after heating to convert FAD to FMN. There were no apparent differences between plasma concentrations of riboflavin or FAD + FMN of the mustang and healthy horses. Likewise, there was no apparent difference in erythrocyte concentrations of riboflavin, but the combined concentration of erythrocyte FAD + FMN was lower in the mustang than in 20 healthy horses. Although individual FAD and FMN concentrations could not be quantified, both FAD and FMN peaks appeared to be lower in the mustang than in healthy horses when HPLC scans were visually inspected.
Concentrations of flavin compounds in erythrocytes and plasma from a mustang horse with FAD-deficient erythrocytes and from clinically healthy horses of various breeds. ∗
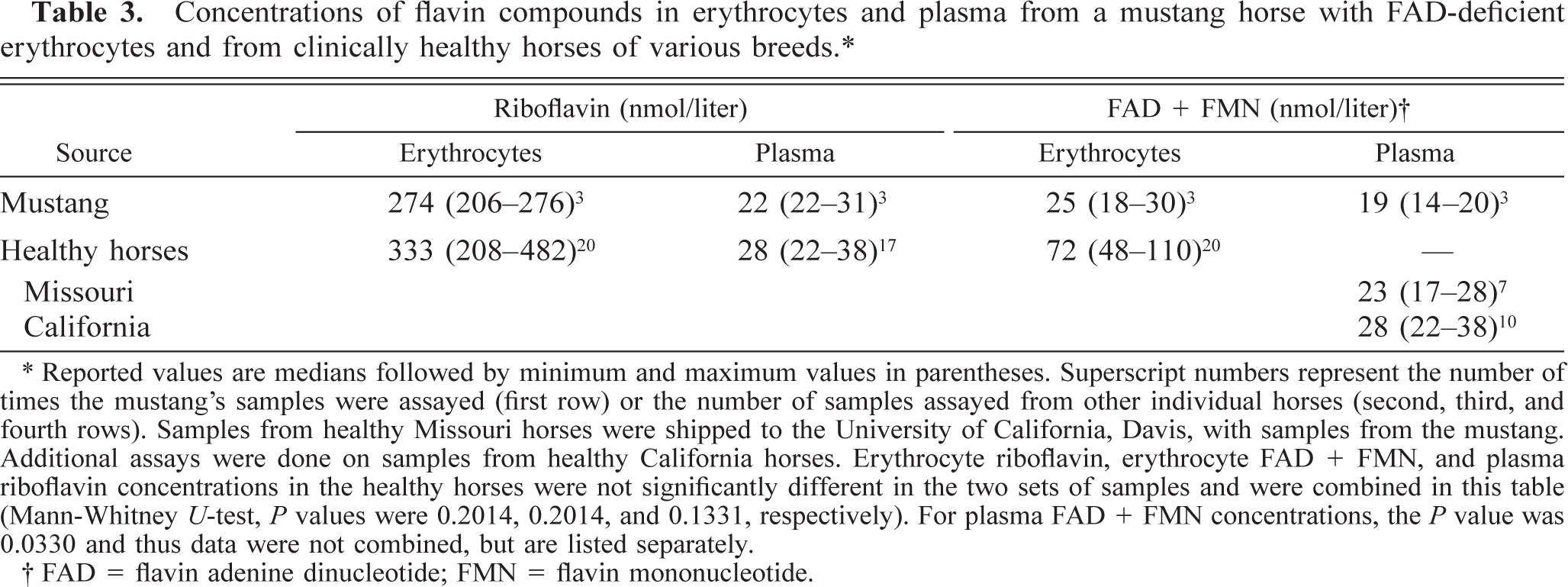
∗ Reported values are medians followed by minimum and maximum values in parentheses. Superscript numbers represent the number of times the mustang's samples were assayed (first row) or the number of samples assayed from other individual horses (second, third, and fourth rows). Samples from healthy Missouri horses were shipped to the University of California, Davis, with samples from the mustang. Additional assays were done on samples from healthy California horses. Erythrocyte riboflavin, erythrocyte FAD + FMN, and plasma riboflavin concentrations in the healthy horses were not significantly different in the two sets of samples and were combined in this table (Mann-Whitney U-test, P values were 0.2014, 0.2014, and 0.1331, respectively). For plasma FAD + FMN concentrations, the P value was 0.0330 and thus data were not combined, but are listed separately.
† FAD = flavin adenine dinucleotide; FMN = flavin mononucleotide.
Discussion
An eccentrocyte is an erythrocyte in which the hemoglobin is located eccentrically, leaving a hemoglobin-poor area in the remaining part of the cell. 20 Eccentrocytes develop secondary to oxidant damage to erythrocyte membranes. An eccentrocyte apparently forms when opposing areas of the cytoplasmic face of the erythrocyte membrane adhere together, concentrating the hemoglobin in the remaining volume of the erythrocyte. The cross-bonding of erythrocyte membranes requires both an alteration in membrane skeletal proteins to make them adhesive and a force that will bring opposing sides of the membrane together. 14,15 It is suggested that membranes can be brought together in vivo by osmotic shrinkage in the kidney and/or when squeezing through the microcirculation. 15 Pyknocytes appear to develop from eccentrocytes after the loss of much of the fused membrane. They are irregularly spheroid, with only a tag of fused membrane remaining. 20,43 It is not known why the eccentrocytes and pyknocytes stained more intensely with NMB, but similar staining of eccentrocytes and pyknocytes has been seen in other patients, including the horse with G6PD deficiency. 43
The significance of the hemoglobin crystals is not known. The presence of large hemoglobin crystals within erythrocytes is commonly recognized in blood films from cats and llamas and rarely recognized in blood films from dogs. 20 No hemoglobin abnormalities have been recognized by hemoglobin electrophoresis, and no pathologic significance has been established for hemoglobin crystals in blood films from these domestic animals. Hemoglobin crystals have not been documented as forming in association with oxidant injury; however, rare hemoglobin crystals also were recognized in blood films from the G6PD-deficient horse. 43 The presence of crystals in an eccentric location and in association with collapsed or fused membranes similar to those of eccentrocytes suggests that at least some of the crystals in the mustang were formed in eccentrocytes.
Eccentrocytes have been seen in animals ingesting or receiving oxidants, including onions, acetaminophen, and vitamin K in dogs; intravenous hydrogen peroxide as a “home remedy” in a cow; and red maple leaves in horses. 20 Eccentrocytes and pyknocytes also have been seen in an American Saddlebred Horse with G6PD deficiency in which no exogenous oxidants were identified. 43 G6PD is the rate-limiting enzyme in the pentose phosphate pathway, thus the G6PD-deficient horse was unable to regenerate the NADPH needed in the protection against endogenous oxidants. The presence of eccentrocytes in the mustang in the absence of administered or consumed oxidants indicates the likelihood of inadequate metabolic protection against endogenously generated oxidants in this horse as well. However, erythrocyte G6PD deficiency was not a factor because G6PD activity was about 70% higher than that measured in two healthy control horses and above the reference interval established previously for horses.
Eccentrocytes and pyknocytes are presumed to have shortened life spans in the circulation. The mild anemia that was sometimes detected in the erythrocyte FAD–deficient mustang was likely the result of this more rapid removal of damaged cells. The increased G6PD and PK activities in the mustang erythrocytes were attributed to a younger average age of the circulating erythrocytes. 27,40,43 The finding of erythroid hyperplasia in the bone marrow was also consistent with a compensated hemolytic anemia.
GR exists in two forms, an active form bound to its cofactor FAD and an inactive form not bound to FAD. 47 The active enzyme activity is measured when GR activity is measured without the addition of FAD to the assay. Total GR activity is measured after the addition of excess FAD to the assay, and the percentage of active enzyme is determined by dividing the GR activity without added FAD by the GR activity with added FAD. On average, about 60% of total GR in healthy horses had bound FAD. The reciprocal, obtained by dividing GR activity with FAD by GR activity without FAD, has been called the activation coefficient. The activation coefficient has been used for many years as a sensitive indicator of riboflavin status in people with high values, indicating dietary riboflavin deficiency. 12 An activation coefficient could not be calculated for the mustang because GR activity was not detectable in assays without FAD addition; however, the substantial increase in GR activity in assays from the mustang after FAD addition indicates that the GR deficiency primarily resulted from a deficiency in erythrocyte FAD. The somewhat lower total GR activity measured in the mustang after FAD addition, as compared with healthy horses, can be explained if GR without bound FAD is less stable than GR with bound FAD. A similar phenomenon has been reported in rats with dietary riboflavin deficiency and resultant GR deficiency. 37
The administration of oral riboflavin supplements to healthy humans not previously receiving dietary riboflavin supplementation caused an increase in erythrocyte GR activity within 1 or 2 days that was associated with an increase in erythrocyte FAD concentration. 4 Treatment of the mustang with injectable riboflavin did not result in detectable erythrocyte GR activity in this animal, but healthy horses were not studied to determine whether this was a unique finding in the mustang. However, these results excluded a defect in riboflavin absorption from the diet as the cause of the erythrocyte FAD deficiency.
A virtual absence of GR activity has been reported in three human siblings, and the deficient activity was not corrected by addition of FAD to assays before GR determination. 30 Erythrocytes from these patients were more susceptible to oxidant stress in vivo and in vitro, but resting GSH concentrations were within reference intervals in the absence of exogenous oxidants. The authors speculated that either extremely low resting activity of GR is sufficient for maintenance of GSH concentrations under physiologic conditions or that the synthesis of GSH from constituent amino acids can keep up with the oxidation of GSH in vivo. Excess GSSG that cannot be reduced to GSH is transported from erythrocytes by an active process. 42
Approximately 3% of blood hemoglobin is oxidized to methemoglobin each day in dogs and humans, 23 but the methemoglobin percentage is usually less than 1% of the total hemoglobin concentration because the methemoglobin that forms is continuously reduced back to hemoglobin, primarily by the flavoprotein-containing Cb5R. 18 In this reaction, electrons are transferred from NADH to ferricytochrome-b5 using FAD as the enzyme-bound prosthetic group, and then the resulting ferrocytochrome-b5 reduces methemoglobin nonenzymatically to hemoglobin (Fig. 1). 25
Methemoglobinemia usually results from either increased production of methemoglobin by oxidants or decreased reduction of methemoglobin because of a deficiency in erythrocyte Cb5R. 19 Methemoglobinemia also has been reported in people with various hemoglobinopathies (referred to as hemoglobin M) that result in methemoglobin formation that is not amenable to reduction. 34 No source of exogenous oxidants was found in the mustang, and the rapid reduction of the methemoglobinemia with intravenous methylene blue excluded a similar hemoglobinopathy. The methemoglobinemia in this mustang was attributed to Cb5R deficiency caused by an erythrocyte FAD deficiency.
Oral therapy with pharmacologic doses of riboflavin resulted in the reduction of blood methemoglobin percentage in some human patients with Cb5R deficiency. 2,26 This methemoglobin reduction has been attributed to NADPH dehydrogenase activity. NADPH dehydrogenase activity is not considered to be important in the reduction of methemoglobin under physiologic conditions because the Km of this enzyme for flavins is many times higher than the concentration of free flavins in erythrocytes. However, NADPH dehydrogenase can reduce methemoglobin in the presence of nonphysiologic concentrations of flavins, which appear to act as substrates rather than as bound cofactors. 48 Although the NADPH dehydrogenase activity in the mustang's erythrocytes was similar to those found in healthy horses, treatment with injectable riboflavin had no detectable effect on blood methemoglobin percentage. A lack of response to riboflavin therapy also was reported in dogs with Cb5R deficiency. 23
In contrast to the stimulation that FAD addition produced in erythrocyte GR assays, the addition of FAD to erythrocyte Cb5R assays from the mustang and healthy horses did not stimulate Cb5R activity. Similarly, insignificant changes in Cb5R activity occurred after the addition of FAD to erythrocyte assays from rats, and short-term riboflavin deficiency in rats did not result in a decrease in erythrocyte Cb5R activity. 35 Low GR activity (without FAD addition) has been reported in some humans with inherited Cb5R deficiency. 2 Oral riboflavin supplementation in one study of Cb5R-deficient people resulted in a rapid increase in erythrocyte GR activity (without FAD addition) but no change in Cb5R activity. 2 This group of investigators speculated that a relative accumulation of NADH, associated with inherited Cb5R deficiency, might interfere with FAD binding to erythrocyte GR in vivo. 17
On the basis of GR studies, it is clear that erythrocytes from the mustang are deficient in FAD. A lack of systemic signs attributable to a generalized defect in riboflavin metabolism and apparently normal activities of GR and Cb5R in neutrophils from the mustang suggested that the defect in the mustang might be limited to erythrocytes. After transport into human erythrocytes, riboflavin is first converted to FMN by riboflavin kinase and then to FAD by FMN adenylyltransferase (Fig. 4). Both enzymes require ATP. 32 A deficiency of either enzyme could result in FAD-deficient erythrocytes. Although unlikely, an upregulation of either FAD pyrophosphatase, allowing for the increased conversion of FAD back to FMN, or acid phosphatase, allowing dephosphorylation of FMN back to riboflavin, 33 also could explain a decrease in FAD concentration (Fig. 4). Total concentrations of FAD + FMN were low in the mustang's erythrocytes when measured by HPLC, and visual inspection of HPLC scans indicated a reduction in concentration of both analytes, compared with erythrocytes from healthy control horses. This finding is consistent with a defect in the conversion of riboflavin to FMN, as has been reported in humans. 3 The more severe deficiency of GR activity versus Cb5R activity in the mustang might be explained if equine erythrocyte Cb5R has a higher affinity for FAD than does GR, as has been proposed in humans. 38
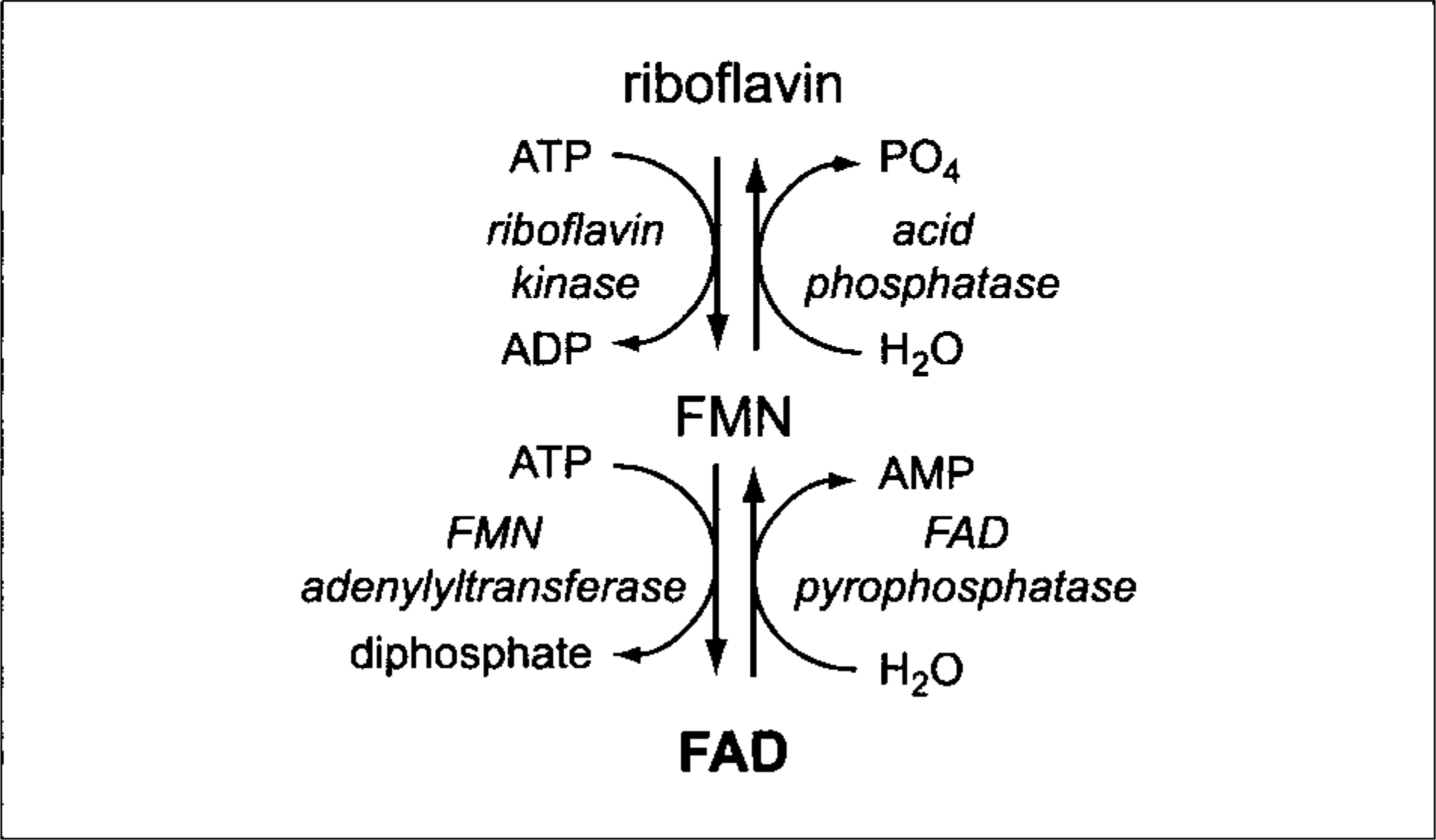
Flavin biochemical pathways in erythrocytes. ADP, adenosine diphosphate; AMP, adenosine monophosphate; ATP, adenosine triphosphate; FAD, flavin adenine dinucleotide; FMN, flavin mononucleotide; and PO4, inorganic phosphate.
Laboratory findings for this mustang are similar to findings for two related trotter mares that were reported to have hemolytic anemia, methemoglobinemia, GR deficiency, and decreased GSH concentrations within erythrocytes. 11 Hematocrit values tended to be lower, and the GR deficient-states were milder in the trotters. Basophilic stippling was identified in erythrocytes from one trotter, but poikilocytosis was not reported. In contrast to the mustang, Cb5R activity was reportedly within reference intervals in the trotters. The investigators measured Cb5R using horse methemoglobin-ferrocyanide complex as a substrate. Hemoglobin for this substrate was presumably prepared after removing Cb5R activity from a healthy horse hemolysate. Failure to adequately remove the Cb5R activity before performing the assay might mask Cb5R deficiency.
In conclusion, the methemoglobinemia in the mustang was attributed to a deficiency in Cb5R activity, and the eccentrocytes and pyknocytes were attributed to a marked deficiency in erythrocyte GR activity. It is not clear whether the low GSH concentration alone explains the increased susceptibility to endogenous oxidants in this animal or whether there might be another effect of GR deficiency that contributes to this defect, such as an impaired ability to reduce lipoic acid to the potent membrane-free radical scavenger dihydrolipoic acid. 9 The decreased Cb5R and GR erythrocyte enzyme activities resulted from an erythrocyte deficiency of their cofactor FAD. The erythrocyte FAD deficiency results from a defect in riboflavin metabolism inside the erythrocytes and does not appear to affect other cells. This syndrome of methemoglobinemia and erythrocyte membrane injury secondary to erythrocyte FAD deficiency has not been reported previously in animals or humans.
Footnotes
Acknowledgements
We thank Ms. Melanie Pate, Dr. Jeff Lakritz, and Dr. Mary Christopher for technical assistance. This work was supported by the Nutritional Assessment Core of the University of California–Davis Clinical Nutrition Research Unit (NIH #DDK35747).