
Abstract
The leaves of Acer rubrum (red maple), especially when wilted in the fall, cause severe oxidative damage to equine erythrocytes, leading to potentially fatal methemoglobinemia and hemolytic anemia. Gallic acid and tannins from A. rubrum leaves have been implicated as the toxic compounds responsible for red maple toxicosis, but the mechanism of action and toxic principle(s) have not been elucidated to date. In order to investigate further how red maple toxicosis occurs, aqueous solutions of gallic acid, tannic acid, and ground dried A. rubrum leaves were incubated with contents of equine ileum, jejunum, cecum, colon, and liver, and then analyzed for the metabolite pyrogallol, as pyrogallol is a more potent oxidizing agent. Gallic acid was observed to be metabolized to pyrogallol maximally in equine ileum contents in the first 24 hr. Incubation of tannic acid and A. rubrum leaves, individually with ileum contents, produced gallic acid and, subsequently, pyrogallol. Ileum suspensions, when passed through a filter to exclude microbes but not enzymes, formed no pyrogallol, suggesting a microbial basis to the pathway. Bacteria isolated from ileum capable of pyrogallol formation were identified as Klebsiella pneumoniae and Enterobacter cloacae. Therefore, gallotannins and free gallic acid are present in A. rubrum leaves and can be metabolized by K. pneumoniae and E. cloacae found in the equine ileum to form pyrogallol either directly or through a gallic acid intermediate (gallotannins). Identification of these compounds and their physiological effects is necessary for the development of effective treatments for red maple toxicosis in equines.
Red maple (Acer rubrum), a deciduous tree native to eastern North America, is often found near pastures where it can cause poisoning if its leaves are ingested by horses.1,14 Affected horses present with methemoglobinemia and Heinz body hemolysis, typical of oxidant-induced damage to erythrocytes.1,14 Previous studies suggest that 1,500 mg of A. rubrum leaves per kg of body weight can induce clinical signs of toxicosis in ponies, especially after mid-September, when harvested leaves were demonstrated to cause death within 18 hr of ingestion.1,14
Tannic acid and gallic acid have been implicated as the primary oxidants in A. rubrum leaves, although metabolites of these compounds, gallic acid and pyrogallol, respectively, were considered more likely to be the ultimate toxins. 7 Ex vivo decomposition of gallotannins, such as tannic acid, and gallic acid appears improbable under normal environmental conditions,16,17 but in vivo metabolism of both compounds via microbial pathways has been well documented in the literature.3,4,11 The presence of gallotannins in general and free gallic acid has been well established in A. rubrum leaves, but it is unknown if the leaves contain tannic acid. 2
Tannins are defined as naturally occurring water-soluble polyphenolic compounds, which typically have molecular weights between 300 and 3,000 Da. 19 Structurally, tannins are divided into 2 classes—hydrolysable and condensed. Hydrolysable tannins contain a sugar core (usually glucose) to which either gallic acid as in tannic acid (Fig. 1A) or ellagic acid as in 1,3-di-O-galloyl-4,6-(S)-hexahydroxydiphenoyl-β-D-glucopyranose (Fig. 1B) moieties are attached through ester linkages (gallotannins and ellagitannins, respectively). Acidic or enzymatic hydrolysis of tannins usually forms monomeric products through a deglycosylation reaction, such as in the formation of gallic acid (Fig. 1C) from the deglycosylation of tannic acid. Further metabolism of these products is possible, such as in the formation of pyrogallol (Fig. 1D) from the decarboxylation of gallic acid. 4 Based on the hypothesis that gallotannins and free gallic acid in A. rubrum leaves are metabolized in vivo to more toxic compounds after ingestion by a horse, the objectives of the current study were to identify the parent compound and toxic metabolite, determine the pathway and location of metabolism, and determine if metabolism occurs due to enzymatic or microbial activity.
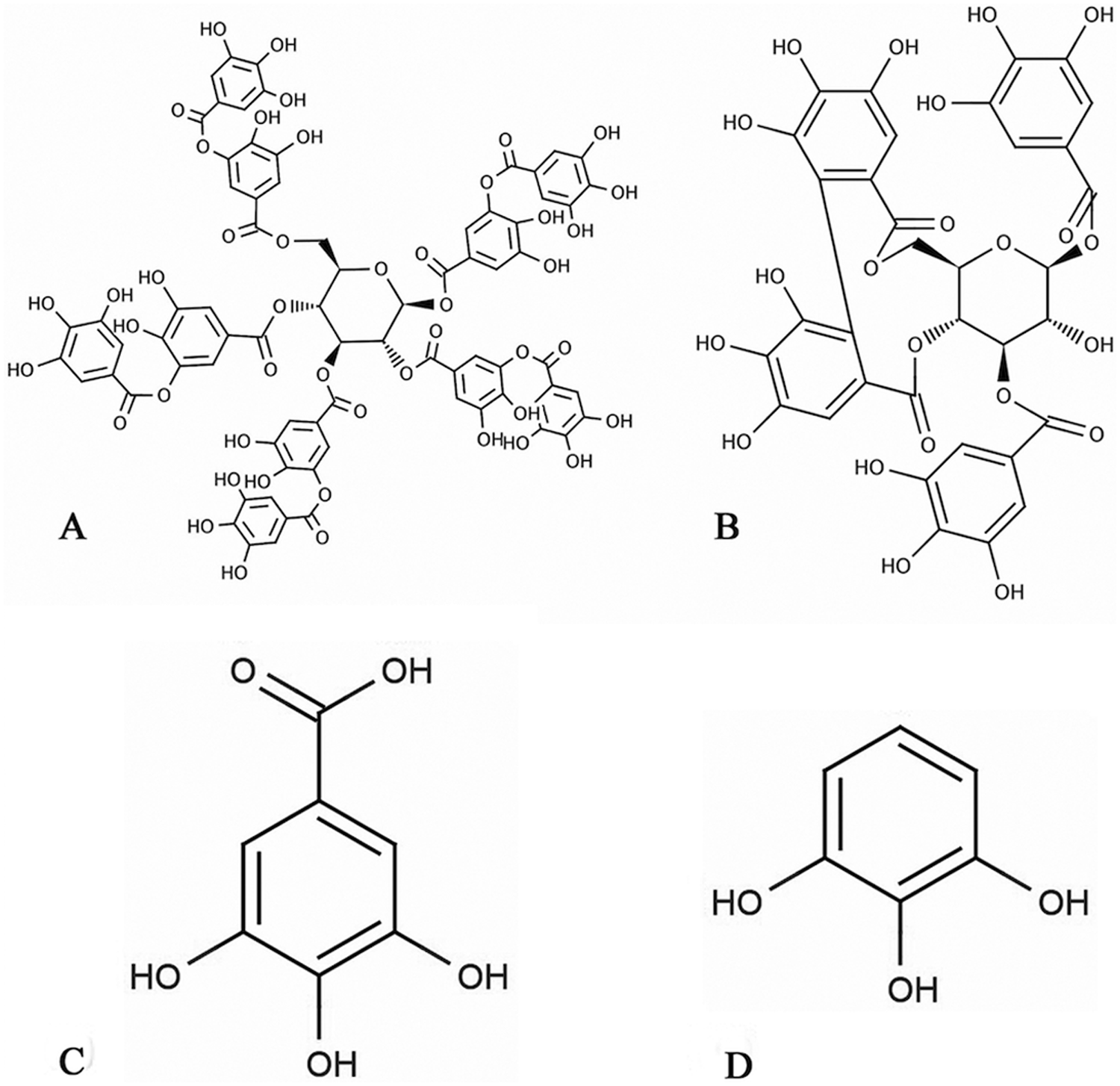
Examples of the 2 major groups of hydrolysable tannins and the metabolites of gallotannins:
Necropsy samples of equine gastrointestinal contents and liver were obtained from 3 horses submitted to the Anatomic Pathology Service of the College of Veterinary Medicine at Cornell University (CU; Ithaca, New York) or the Veterinary Diagnostic Laboratory at the University of Kentucky (Lexington, Kentucky). Samples were taken within 24 hr of death, and selected horses did not undergo recent antibiotic treatment or have evidence of metabolic disease. Samples were stored at 4°C within 1 hr of collection and analyzed within 24 hr of collection. Equine blood samples were submitted to the CU Animal Health Diagnostic Center for routine clinical pathology analysis or drug screening. Leaves from A. rubrum were collected from Cornell Plantations (CU). Within 24 hr of collection, leaves were dried at 60°C in a forced air drying oven a for 18 hr, ground in a Wiley mill b fitted with a 3-mm screen, and stored at −20°C.
The following standards were used throughout the current study: gallic acid, c tannic acid, d pyrogallol, c catechol, c ferulic acid, e p-coumaric acid, e and trans-cinnamic acid. c Each standard was prepared in water f at a final concentration of 1 mg/ml and stored at 4°C.
Analysis was performed using capillary gas chromatography–mass spectrometry (GC-MS). g A 1-µl aliquot of sample was injected via an autosampler using a splitless injection technique into a (5%-phenyl)-methylpolysiloxane 30 m × 0.25 mm × 0.25-µm capillary column. g The carrier gas was helium at a flow rate of 1 ml/min. The injector port was set at 250°C. The initial oven temperature was set at 80°C, held for 2 min, ramped to 290°C at 15°C/min and held for 10 min. A solvent delay of 3.5 min was used. Temperature of the MS source and MS Quad was 230°C and 150°C, respectively. The MS was operated in the full scan mode using electron impact ionization. Compounds were identified as their trimethylsilyl derivatives on the basis of their retention time and electron impact spectra by comparison with authentic standards and our online reference spectra contained in the Wiley Registry of Mass Spectral Data. h Quantification of compounds were done using a standard curve.
Data was analyzed using two-tailed paired Student t-tests performed using a commercial spreadsheet application. i Mean concentrations of gallic acid and pyrogallol were compared between time points for each experiment. Mean concentrations of gallic acid and pyrogallol were compared between individual gastrointestinal tract content and liver samples at each time point.
To identify compounds in A. rubrum capable of inducing methemoglobinemia, 4 sets of equine blood samples obtained from different horses were combined to form a composite equine blood sample. The composite sample was suspended in an equal amount of 0.9% NaCl solution to form a 1:1 suspension. Eleven 1-ml aliquots of this suspension were placed in the wells of a 24-well plate and spiked with one of the above standards as shown in Figure 2 in triplicate. Incubation of the plate at 37°C in a convection oven j for 3 hr followed by qualitative assessment based on color change (from red to brown-black) indicated methemoglobin formation in the tannic acid, gallic acid, pyrogallol, and catechol spikes. Based on the amount of standard required to induce color change and the intensity of the change, pyrogallol was the most potent oxidant and therefore a candidate as one of the ultimate toxicants. Formation and persistence of pyrogallol was therefore considered to be the endpoint for the metabolic pathway of toxicant synthesis and was monitored throughout the study.

Results of the incubation of simple phenolics with equine blood samples at 37°C for 3 hr. Dark color is indicative of methemoglobin formation. A: tannic acid (200 µg/ml); B: gallic acid (200 µg/ml); C: pyrogallol (25 µg/ml); D: pyrogallol (50 µg/ml); E: pyrogallol (100 µg/ml); F: pyrogallol (200 µg/ml); G: catechol (200 µg/ml); H: ferulic acid (200 µg/ml); I: p-coumaric acid (200 µg/ml); J: trans-cinnamic acid (200 µg/ml); K: control blood sample.
To identify free pyrogallol in A. rubrum leaves, 500 mg of dried and ground leaves collected in the summer and fall were extracted in 10 ml of methanol j and water f (70:30) solution by agitation and sonication. k One-milliliter aliquots of each extract were dried under nitrogen, reconstituted with 50 µl of N,O-Bis(trimethylsilyl)trifluoroacetamide + 1% trimethylchlorosilane l and 50 µl of ethyl acetate, j and refluxed for 1 hr at 80°C to form a trimethylsilyl derivative. Gas chromatography–mass spectrometry analysis indicated that both seasonal samples lacked free pyrogallol but had nearly identical chemical compositions. Therefore, pyrogallol is not present in red maple leaves, nor is it synthesized ex vivo in A. rubrum during the wilt stage. If pyrogallol is the causative agent of red maple toxicosis, it is synthesized by in vivo metabolism.
The gastrointestinal tract contents and liver were examined for their ability to metabolize gallic acid to pyrogallol. Two grams of content samples from the ileum, jejunum, cecum, and colon were individually suspended in 40 ml of water f and agitated to form a homogenous suspension, and 2 g of liver were homogenized in 40 ml of water. f Aliquots of 10 ml of these mixtures were spiked with 500 µl of gallic acid standard to achieve a final concentration of 1 µg of gallic acid per mg of homogenate. Samples were incubated in standing culture at 38 ± 2°C in a heating/stirring module m for 120 hr. Aliquots of 1 ml were drawn every 24 hr, acidified with 0.6 N HCl to pH 2.5, and extracted with 5 ml ethyl acetate. j These extracts were evaporated to dryness under nitrogen, reconstituted, derivatized, and analyzed by GC-MS.
Contents from each section of the gastrointestinal tract and liver metabolized the gallic acid to pyrogallol at different rates (Fig. 3), satisfying the pathway proposed in a previous study. 7 The liver formed the maximum pyrogallol in 24 hr (588 ± 685 µg/ml), but the ileum formed the next highest concentration of pyrogallol with less variation between samples (160 ± 112 µg/ml), although the difference was not statistically significant (p > 0.05). The initial 24-hr period is important in red maple toxicosis: death can occur within 18 hr of ingestion of leaves. 14 There was marked variation in the amount of pyrogallol formed in the first 24 hr in the liver homogenate, which can be partly explained by genetic and environmental factors affecting hepatic enzyme production. Because of this variation, and the limited supply of equine tissues available, the liver was not further investigated in the present study. The ileum was chosen for further study because it had consistently high pyrogallol production, making it a more suitable choice for further experimentation. Additionally, gastrointestinal metabolism and absorption of gallotannins and pyrogallol has been demonstrated as a pathway for tannin-induced toxicosis in monogastric animals,5,12,24 which supports the decision to choose the ileum for further study rather than the liver.
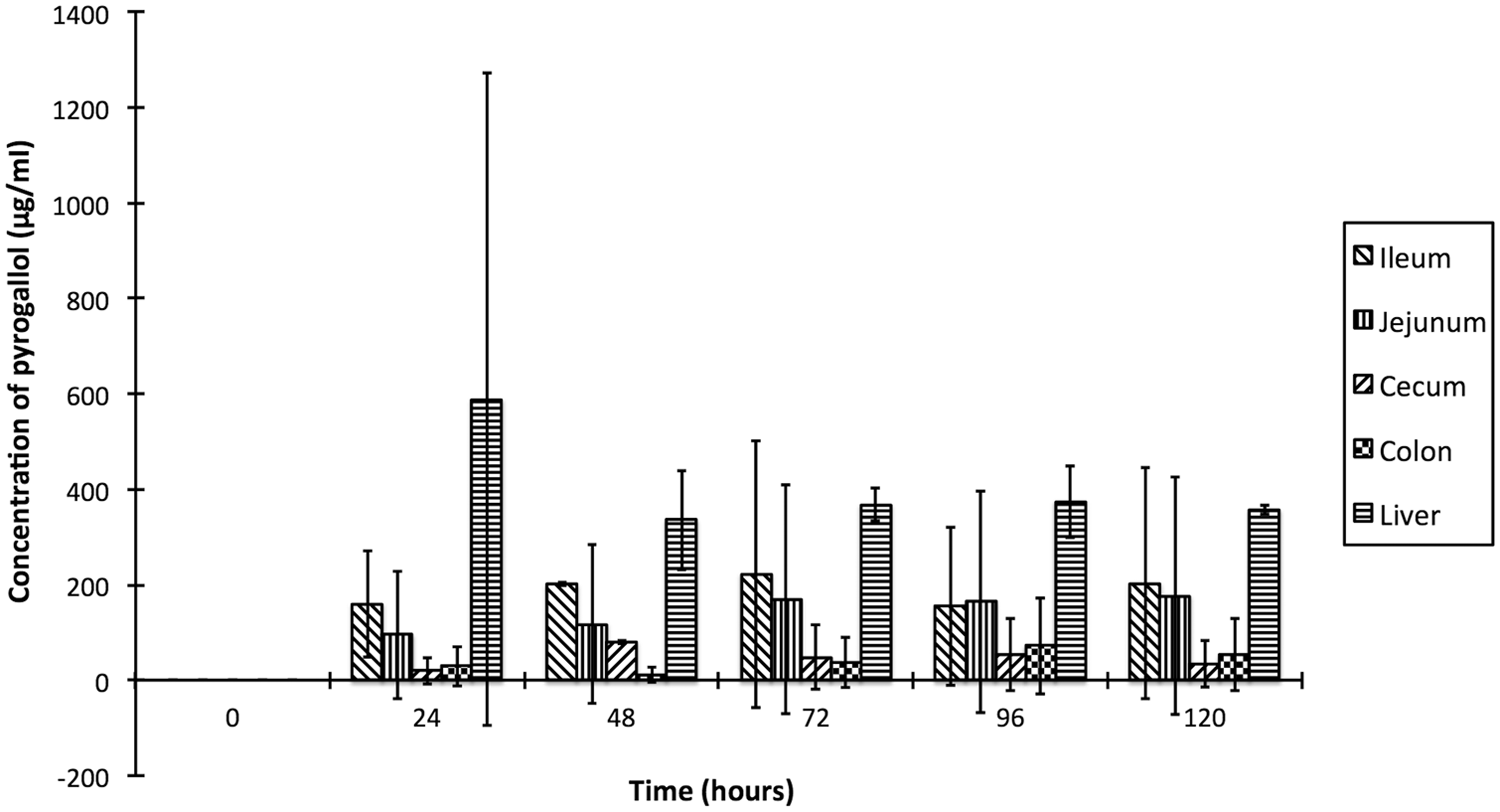
Formation of pyrogallol from the metabolism of gallic acid by equine ileum, jejunum, cecum and colon contents, and liver homogenate.
Metabolism of tannic acid to gallic acid has been proposed as an alternate pathway for red maple toxicosis. 7 Repeating the previous experiment with 500 µl of the tannic acid standard in place of the gallic acid standard using ileum content homogenate, the gallic acid concentration increased from 0 hr (77 ± 16 µg/ml) to 24 hr (185 ± 128 µg/ml), after which the gallic acid concentration decreased until the end of the incubation period at 120 hr (6 ± 2 µg/ml). The pyrogallol concentration increased between 0 hr (0 ± 0 µg/ml) and 48 hr (34 ± 39 µg/ml), then decreased slightly by 72 hr (5 ± 8 µg/ml), and increased again at 120 hr (48 ± 68 µg/ml), suggesting that this pathway occurs in the ileum, although none of the differences were statistically significant (p > 0.05).
Interestingly, metabolism of tannic acid by ileum content produced gallic acid and pyrogallol, suggesting that the previously proposed pathways are complimentary rather than mutually exclusive, with gallic acid as the intermediate. The gallic acid observed at 0 hr when tannic acid was incubated with ileum contents was probably attributable to impurities in the tannic acid sample, which was 99% pure. Gallotannins are too large and polar to be analyzed by the GC-MS used in the current study, which has an m/z range of 1.6–800 Da (https://www.chem.agilent.com/Library/specifications/Public/5989_0082EN_low.PDF), thus the presence of gallotannins was indicated by formation of gallic acid metabolites.
To confirm that these metabolic pathways were complimentary, 100 mg of ground dried A. rubrum leaves collected in the fall (estimated to contain approximately 530 µg of gallic acid based on a previous study 7 ) were incubated in standing culture at 38 ± 2°C with ileum contents. Pyrogallol formation was measured over time. Gallic acid increased from 0 hr (382 ± 281 µg/ml) to 48 hr (1067 ± 877 µg/ml), then decreased until the end of the incubation period at 120 hr (132 ± 111 µg/ml; Fig. 4), but only the decrease between 48 and 72 hr was statistically significant (p < 0.05). Pyrogallol increased from 0 hr (0 ± 0 µg/ml) to 48 hr (2562 ± 253 µg/ml), although only the increase between 24 and 48 hr was significant (p < 0.05), and reached a plateau at 72 hr (2286 ± 603 µg/ml). The increase in gallic acid over the first 48 hr is attributable to the deglycosylation of gallotannins, and the simultaneous increase in pyrogallol is attributed to the decarboxylation of free gallic acid from the leaves. Based on these results, gallotannins and free gallic acid in A. rubrum leaves are metabolized in the equine ileum to pyrogallol either directly (from free gallic acid) or through a gallic acid intermediate (from gallotannins).
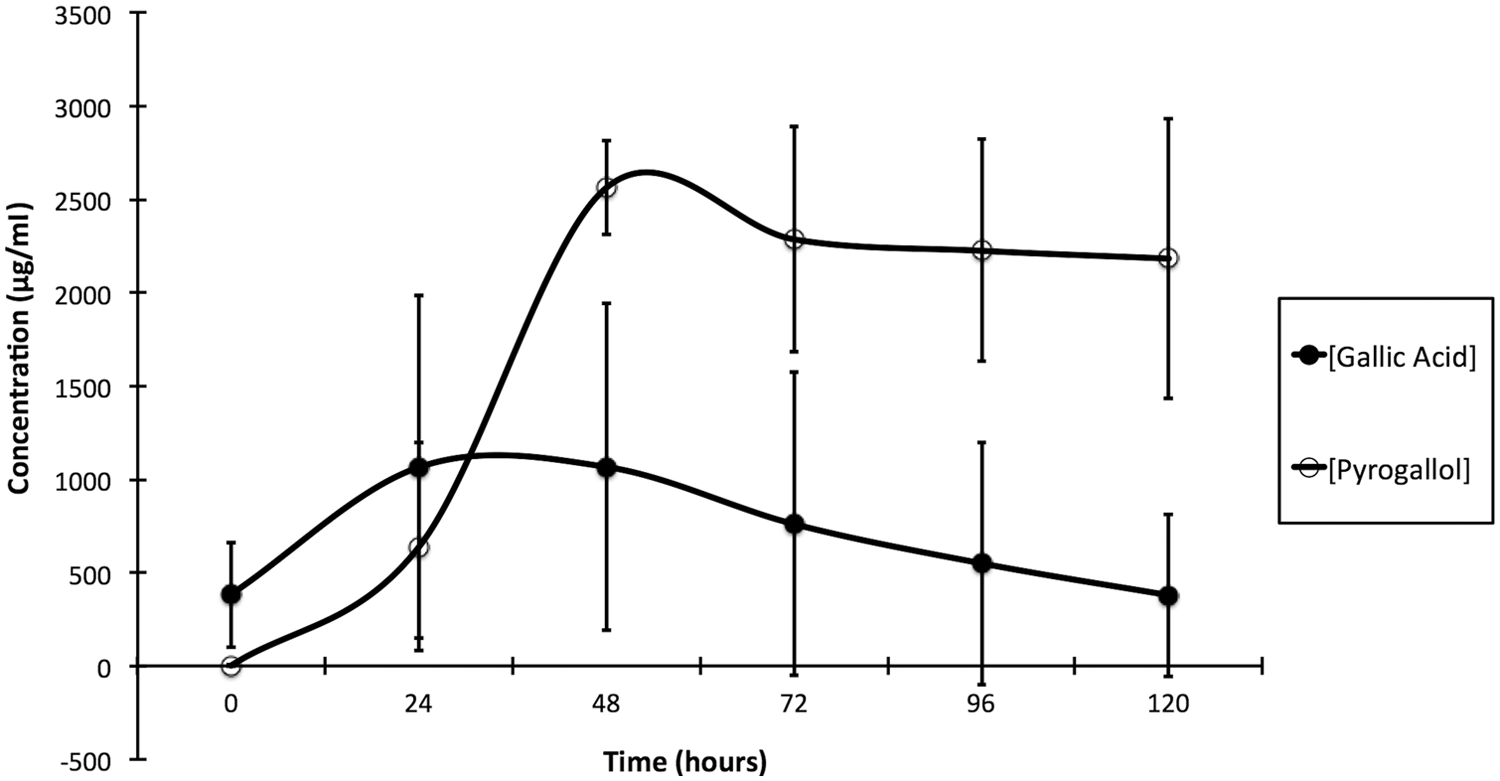
Metabolism of 100 mg of Acer rubrum leaves by the equine ileum contents.
Metabolic processes occur either due to either inherent enzymatic activity of the animal or microbial metabolism. A 40-ml aliquot of ileum suspension was centrifuged n (3,000 × g), the supernatant filtered through a 0.45-µm cellulose filter, o and the filtrate passed through a 0.22-µm filterf,p to remove bacteria (approximately 1 µm in length). Matrix-soluble enzymes pass through the filter. Five hundred ml of gallic acid standard and 100 mg of dried ground fall A. rubrum leaves were each added to 10-ml aliquots of the filtrate. Both aliquots were incubated and analyzed via GC-MS as previously described, and results were compared to results from the previous experiments with unfiltered ileum contents.
The incubation of the gallic acid standard with filtered and unfiltered suspensions of ileum contents resulted in more rapid metabolism of gallic acid in the unfiltered suspension, though changes in the gallic acid concentrations were not significantly different between filtered and unfiltered suspensions (p > 0.05). There was a significant decrease in gallic acid in the unfiltered suspension between 48 and 72 hr (p < 0.01). Pyrogallol was only formed in the unfiltered suspension, suggesting that microbes were necessary for metabolism of gallotannins to pyrogallol (Fig. 5). It is unknown why no pyrogallol was formed in the filtered suspension despite metabolism of gallic acid, but it is possible that enzymes present in the suspension may have metabolized the gallic acid to a different endpoint that was not considered in this assay.
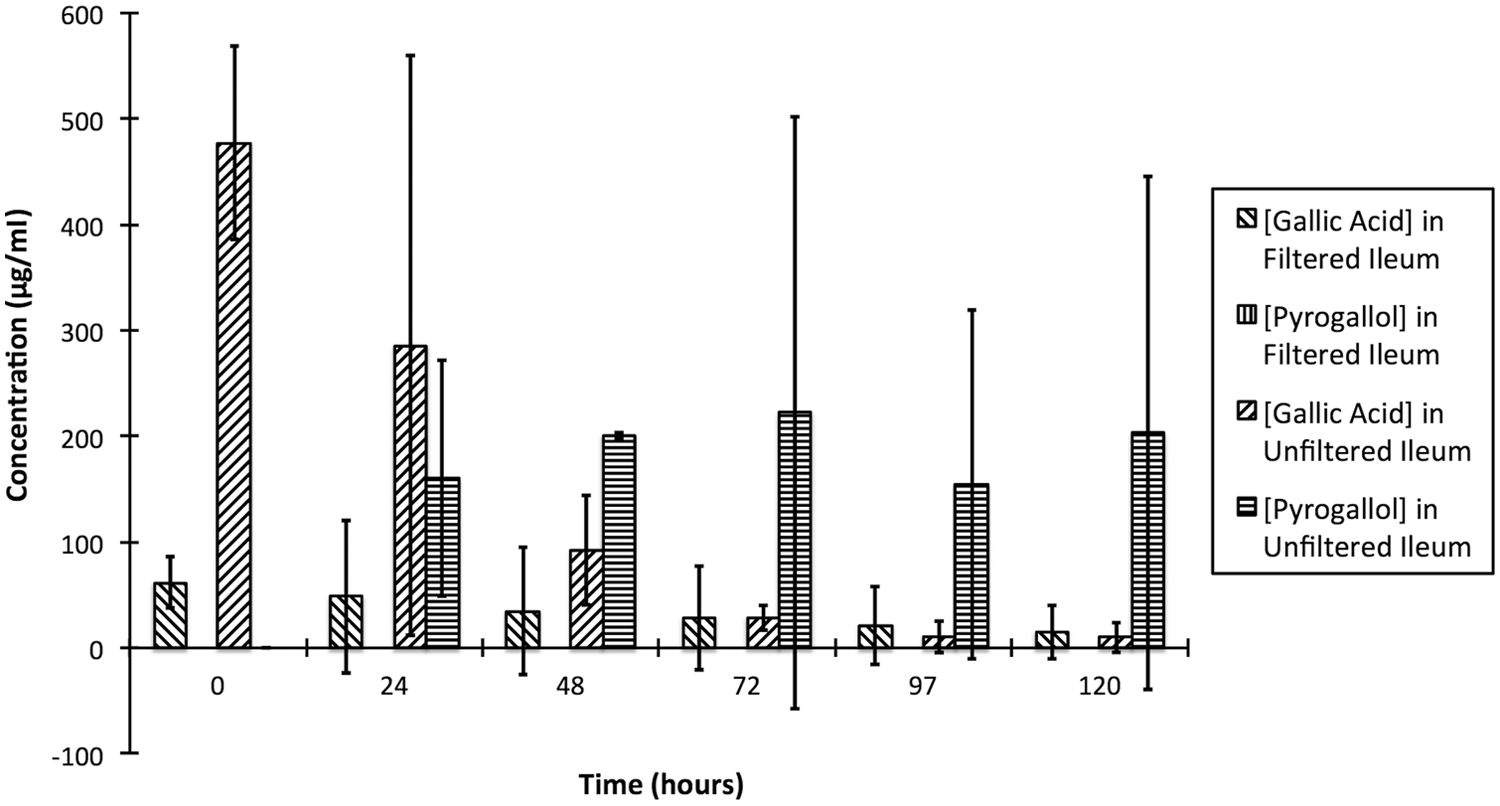
Comparison of the enzymatic and microbial metabolism of gallic acid in the equine ileum contents.
Incubation of A. rubrum leaves with the filtered ileum content suspension resulted in inconsistent changes in gallic acid and pyrogallol concentrations, compared to the unfiltered sample. The results of the unfiltered ileum suspension assay are as shown in Figure 4. The concentration of gallic acid in the filtered ileum suspension initially increased from 365 ± 292 µg/ml at 0 hr to 1,772 ± 57 µg/ml at 24 hr, then decreased to 1,158 ± 845 µg/ml at 48 hr and again increased to 2,097 ± 737 µg/ml at 72 hr before decreasing to 727 ± 769 µg/ml at 96 hr. Simultaneously, the concentration of pyrogallol increased slightly from 0 ± 0 µg/ml at 0 hr to 24 ± 34 µg/ml at 24 hr, then increased sharply to 1,312 ± 1,441 µg/ml at 48 hr before decreasing slightly to 1,209 ± 877 µg/ml at 72 hr and then increasing to 2,400 ± 455 µg/ml at 96 hr. These changes in concentration are similar to the system approaching a state of equilibrium between gallic acid and pyrogallol; a mechanism for the carboxylation of pyrogallol to gallic acid in a closed system has been demonstrated previously. 25 Statistically significant changes in gallic acid concentrations were observed between 0 and 24 hr and again between 72 and 96 hr (p < 0.05) for the filtered sample.
Pyrogallol production in the filtered ileum content suspension incubated with leaves was unexpected, but K. pneumoniae, which has been demonstrated to metabolize gallic acid and gallotannins to pyrogallol, 11 was cultured and isolated from the incubation tube. The presence of K. pneumoniae could be due to contamination or improper filtration, but K. pneumoniae was not isolated from the incubation tube containing the filtered suspension and gallic acid standard, thus the A. rubrum leaves were the likely source of the bacteria. When A. rubrum leaves from the same stock were cultured, however, E. cloacae was isolated, but not K. pneumoniae, suggesting that different bacteria colonized different leaves within the stock. Both are enteric organisms in horses and other animals and are capable of gallotannin metabolism and pyrogallol production.3,11
It was beyond the scope of the current study to determine the source of microbial contamination of the maple leaves. Although sterile technique was not used to collect the leaves, and leaves were stored frozen for up to 9 months before analysis, other investigators have reported deposition of aerosolized bacteria (including normal flora of the gastrointestinal tracts of human beings and other animals) on plants, especially near water treatment facilities.6,9,21 Both the CU Animal Hospital (and associated outdoor paddocks) and the Cornell Water Filtration Plant are within 1.25 km of Cornell Plantations where the A. rubrum leaves were collected, and there are several other wastewater treatment and filtration plants within an 8 km radius of Cornell Plantations, and these are likely sources of enteric bacteria (including K. pneumoniae and E. cloacae) deposited on the leaves. Additionally, an exploratory study incubated A. rubrum leaves in distilled, filtered water f at 38 ± 2°C and indicated an increased gallic acid concentration and pyrogallol formation over a 48-hr period, followed by a decrease in the concentration of both compounds at 72 hr. These results strongly suggest the presence of microbes on A. rubrum leaves capable of metabolizing hydrolysable tannins and gallic acid to pyrogallol, though it is possible that the leaves were contaminated due to lack of sterile collection techniques and long-term freezer storage before analysis.
To identify specific bacteria in the equine ileum capable of catalyzing metabolism of gallotannins and gallic acid to pyrogallol, samples of an ileum suspension capable of metabolizing gallic acid to pyrogallol were inoculated onto trypticase soy agar II with 5% sheep blood q and modified eosin methylene blue agar. q Plates were incubated at 35°C with 5% CO2 for 48 hr. Bacterial colonies of differing morphologies were purified onto trypticase soy agar II with 5% sheep blood. q Isolates were tested for their ability to metabolize gallic acid, tannic acid and other gallotannins, and gallic acid in ground dried A. rubrum leaves to pyrogallol as described below. Bacterial isolates capable of pyrogallol formation were identified using a commercial identification system r as K. pneumoniae with 96.00% certainty for 2 of the equine ileum content samples and as E. cloacae with 100.00% certainty for 1 of the equine ileum content samples.
To confirm the ability of K. pneumoniae and E. cloacae to metabolize gallotannins and gallic acid to pyrogallol, nine 10-ml aliquots of N-minimal medium, prepared using a previously described method, 20 were mixed with 20 µl of tryptic soy broth q and each was individually inoculated with approximately 108 cultured K. pneumoniae or E. cloacae. Aliquots were individually spiked with 500 µl of gallic acid standard, 500 µl of tannic acid standard, or 10 mg of fall A. rubrum leaves (estimated to contain approximately 53 µg of gallic acid based on a previous study 7 ) and allowed to incubate for 120 hr at 38 ± 2°C. Sampling, extraction, and analysis were conducted as described above.
Isolates of K. pneumoniae metabolized gallic acid and tannic acid standards, and gallotannins and free gallic acid from leaves. Gallic acid metabolism was rapid: gallic acid decreased from 121 ± 17 µg/ml at 0 hr to 0 ± 0 µg/ml at 24 hr. Tannic acid metabolism to gallic acid was slower: tannic acid decreased from 99 ± 3 µg/ml at 0 hr to 10 ± 1 µg/ml at 24 hr and 3 ± 4 µg/ml at 96 hr. No pyrogallol formation was observed in either sample. Incubation of K. pneumoniae with A. rubrum leaves resulted in slow metabolism of gallic acid and tannic acid, but pyrogallol was produced and persisted for 120 hr despite the trace concentration of gallic acid (near 0 µg/ml).
Isolates of E. cloacae metabolized gallic acid and tannic acid standards, and gallotannins and free gallic acid from leaves. Gallic acid metabolism was slow: gallic acid decreased from 63 µg/ml at 0 hr to 48 µg/ml at 48 hr to 26 µg/ml at 120 hr. Tannic acid metabolism to gallic acid was more rapid: gallic acid increased from 61 µg/ml at 0 hr to 154 µg/ml at 48 hr to 159 µg/ml at 120 hr. No pyrogallol formation was detected in either sample. The incubation of E. cloacae with leaves produced an initial increase in gallic acid followed by a sharp decrease, while pyrogallol increased steadily until the 96 hr, then decreased slightly.
Statistical analysis was inappropriate due to the small number of samples (n = 2 for K. pneumoniae, n = 1 for E. cloacae). However, both K. pneumoniae and E. cloacae were capable of metabolizing the gallotannins and gallic acid from A. rubrum leaves.
Klebsiella pneumoniae and E. cloacae metabolize gallotannins and gallic acid to pyrogallol and other compounds,3,8 but observed results differ from those in the literature because no pyrogallol was detected when either organism was incubated with pure standards, although pyrogallol was produced when either organism was incubated with A. rubrum leaves. It is not specifically known why pyrogallol could not be detected in the assays utilizing gallic or tannic acid standards. Contrasting with the complex matrix of the leaves and microflora of the ileum, the rapid metabolism of a limited quantity of pure gallic or tannic acid and their metabolites by pure cultures of E. cloacae or K. pneumoniae may have precluded the detection of pyrogallol. It is possible that the pyrogallol pathway was bypassed in favor of a pathway involving the formation of 4-O-methyl gallic acid as described in a previous study. 5 Alternately, pyrogallol could have been metabolized to other products by K. pneumoniae or E. cloacae very rapidly after formation, meaning the presence of pyrogallol was too transient for detection at the selected time points, as has been demonstrated in other studies.8,15 Further research will be necessary to determine the true pathways for gallic acid and tannic acid metabolism by these organisms.
The results of the present study, with rare exception, did not demonstrate any statistical significance due to the small sample size (n = 3). Tissue samples came from horses from different locations in New York and Kentucky with different management situations, which was expected to affect their hepatic enzymes and intestinal microflora.10,18 However, general trends support the proposed metabolic pathway summarized in Figure 6. Pyrogallol, which persisted in ileum content in vitro for ≥120 hr, is likely absorbed into the blood where it forms free radicals in the presence of Fe, leading to oxidation of Fe2+ in hemoglobin to Fe3+ to form methemoglobin. 13
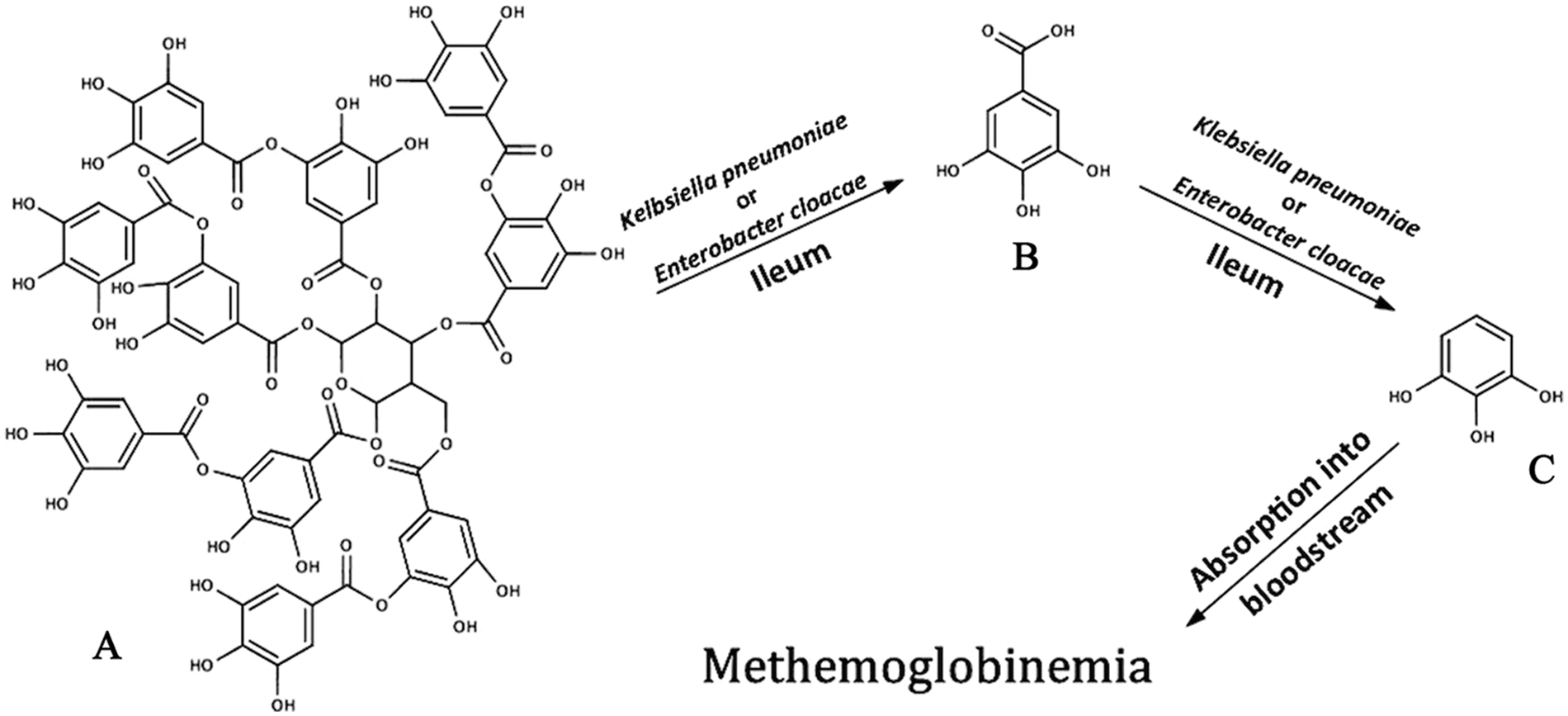
Proposed pathway for red maple toxicosis in equines. Tannic acid (
The current study covers only a limited aspect of red maple toxicosis: the metabolism of gallotannins and free gallic acid to pyrogallol by intestinal organisms such as K. pneumoniae and E. cloacae. Given the myriad microbes in the ileum, dependent upon factors such as diet and habitat,10,18 other microorganisms likely contribute to pyrogallol production. Further research is needed to determine how well pyrogallol is absorbed in the equine intestine. There was evidence of hepatic involvement in gallotannin metabolism as well, which is another area for future research. Additionally, other compounds in A. rubrum leaves could be involved in toxicosis. 7 More work is needed to determine the complete pathophysiology, absorption, and sites of metabolism of gallotannins in monogastric organisms.5,12,24 The recent isolation of the tannase enzyme (the enzyme catalyzing the first step in the proposed pathway) from E. cloacae and K. pneumoniae22,23 could be important in the development of effective treatment options for red maple toxicosis, possibly including antimicrobials and drugs that inhibit intestinal absorption of pyrogallol.
Footnotes
Acknowledgements
The authors thank Drs. Motoko Mukai and Dan Brown for their help and advice on study design and interpretation of results; Joseph Hillebrandt, Kerry Manzell, Melissa Aprea, Lorrie Miller, Dr. Cynthia Gaskill, and Patrick Burke for technical assistance; and Dr. HG Kang for his support.
a.
The Grieve Corporation, Round Lake, IL.
b.
Arthur H. Thomas Co., Philadelphia, PA.
c.
Acros Organics, Fair Lawn, NJ.
d.
Mallinckrodt, St. Louis, MO.
e.
MP Biomedicals LLC, Solon, OH.
f.
Millipore Corp., Billerica, MA.
g.
HP-5ms, Agilent Technologies Inc., Santa Clara, CA.
h.
Copyright © 1999-2012 by John Wiley and Sons Inc.; ed. McLafferty FW; online ISBN: 9780470175217.
i.
Microsoft Excel, Microsoft Corp., Redmond, WA.
j.
Fisher Scientific, Pittsburgh, PA.
k.
Branson Cleaning Equipment Co., Shelton, CT.
l.
Thermo Scientific, Bellefonte, PA.
m.
Reacti-Therm, Pierce Chemical Co., Rockford, IL.
n.
International Equipment Co., Needham Heights, MA.
o.
Grace Davison Discovery Science, Deerfield, IL.
p.
VWR, Radnor, PA.
q.
BD, Franklin Lakes, NJ.
r.
Sensititre, TREK Diagnostics Inc., Cleveland, OH.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by a memorandum of understanding with the National Veterinary Research and Quarantine Service of the Republic of Korea.