
Abstract
Polyarteritis nodosa (PAN) is an idiopathic necrotizing vasculitis affecting small- to medium-sized arteries. The disease is well recognized in humans, and PAN-like syndromes have been described in a number of other species. This report describes a case of PAN in a 6-year-old male cynomolgus macaque. The animal had necrotizing arteritis affecting vessels in the kidney, small intestine, colon, heart, spleen, mesentery, urinary bladder, and pancreas. The lesions were segmental in distribution and of varying severity and stage of development. A transmural mixed inflammatory cell infiltrate was present, often accompanied by fibrinoid necrosis of the tunica media and loss of the internal elastic lamina. Immunohistochemical staining showed that many of the infiltrating cells were T lymphocytes and histiocytes, suggesting a cell-mediated component to the pathogenesis.
Polyarteritis nodosa (PAN) is a systemic necrotizing vasculitis of humans affecting small- and medium-sized arteries. 6,8,12 Renal arteries are most commonly affected, with involvement of other visceral arteries and relative sparing of the lungs. Areas of arterial branching are predilection sites, and the inflammation often results in grossly visible nodular thickening of affected arteries. The lesions are often segmental within an artery and polyphasic, with acute and chronic lesions present within the same tissue section. PAN is most common in adults, and the pathogenesis is uncertain. In animals, reports of PAN-like diseases have included rats, 4,16 mice, 11,14,15 dogs, 13 cats, 2 and pigs. 9 Among nonhuman primates, the disease has been described only in the cynomolgus macaque (Macaca fascicularis). 1 Here, we present the second report of PAN in a cynomolgus macaque.
A 6-year-old male cynomolgus macaque housed at the Southwest Foundation for Biomedical Research was presented with an acute onset of lethargy, dehydration, and fever. The animal was purchased from a commercial vendor 15 months before onset of illness. It was a member of a breeding colony and had no known history of experimental treatment. There were no significant prior health problems, and tests for cercopithicine herpesvirus 1 (herpes B virus); simian immunodeficiency virus; simian T-cell leukemia virus; and simian retroviruses 1, 2, and 5 performed 14 months before the onset of illness were negative. Blood work at presentation revealed elevated blood urea nitrogen and creatinine and decreased serum protein, red blood cell count, hemoglobin, and hematocrit (Table 1). Renal failure was diagnosed, and intravenous fluids and a blood transfusion were administered. On day 23, edema of the face, scrotum, and subcutis developed. The blood work was repeated on days 25 and 33 and revealed continuing renal failure, hypoproteinemia, regenerative anemia, and thrombocytopenia (Table 1). The animal did not respond to treatment and was euthanatized on day 34.
Selected blood and serum chemistry values. ∗
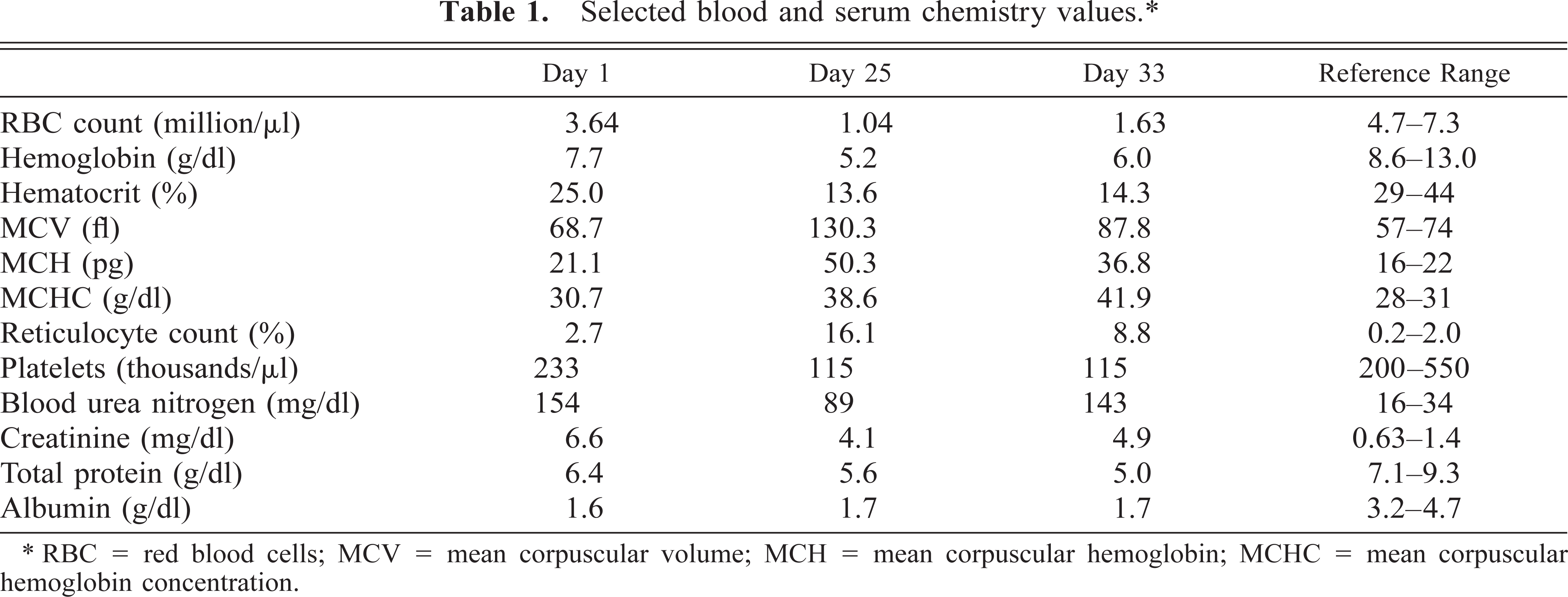
∗ RBC = red blood cells; MCV = mean corpuscular volume; MCH = mean corpuscular hemoglobin; MCHC = mean corpuscular hemoglobin concentration.
On necropsy, the most significant findings were severe subcutaneous edema and excessive clear fluid in all body cavities. Representative tissue sections were fixed in formalin, processed routinely, sectioned at 5 µm, and stained with hematoxylin and eosin (HE). Selected sections were stained with Verhoeff–van Gieson and Masson trichrome stains. Sections of kidney were sectioned at 2 µm and stained with HE, Jones methenamine silver, and Masson trichrome. Small intestinal samples were stained immunohistochemically using a streptavidin peroxidase method according to the manufacturer's instructions (Histostain SP Kit; Zymed Laboratories, San Francisco, CA). The primary antibodies were against CD3 (rabbit anti-human polyclonal at 1 : 100; DAKO, Carpinteria, CA), CD79α (mouse anti-human monoclonal at 1 : 25; DAKO), and MAC 387 (mouse anti-human monoclonal at 1 : 300; DAKO). CD3 and CD79α stain T and B lymphocytes, respectively, whereas MAC 387 has specificity for macrophages and neutrophils. The antigen-retrieval method was pepsin treatment for CD3, pronase treatment for MAC 387, and microwave treatment for CD79α. The chromagen was 3,3′-diaminobenzidine, and the slides were lightly counterstained with Gill's hematoxylin. Human tonsil was used as a positive control. For negative controls, the primary antibody was substituted with normal rabbit serum or normal mouse immunoglobulin G (Santa Cruz Biotechnology, Inc., Santa Cruz, CA).
Histologically, small- to medium-sized arteries within the kidney, small intestine, colon, heart, spleen, mesentery, urinary bladder, and pancreas exhibited inflammatory changes of varying severity and stage of development. Arteries within the lung, liver, adrenal gland, brain, skeletal muscle, salivary glands, and lymph nodes were normal. The mildest change noted was infiltration of the tunica adventitia and tunica media with small to moderate numbers of lymphocytes, histiocytes, and fewer neutrophils, eosinophils, and plasma cells. This change was often segmental (Figs. 1, 2). In some affected vessels, inflammatory cell infiltrates were accompanied by intimal proliferation, which was characterized by increased numbers of fusiform cells within a small amount of mucinous stroma (Fig. 2). Occasional vessels exhibited marked intimal proliferation, with minimal inflammatory cell infiltration of the vessel wall. In more severe lesions, marked fibrinoid necrosis was present in the tunica media, with loss of the internal elastic lamina (Fig. 3). Necrotic vessels had edema and numerous inflammatory cells in the tunica adventitia and surrounding tissues. In severely affected vessels, the lumen was often occluded by intimal proliferation, inflammatory cells, necrotic debris, and thrombus formation. Rare arteries exhibited aneurysmal dilation. In the kidney, many arteries exhibited chronic changes, with replacement of the vessel wall by a thick layer of mature fibrous connective tissue (Fig. 4). The distribution of arterial lesions appeared random and segmental, with normal arterial profiles sometimes present adjacent to severely affected ones. Veins, venules, arterioles, and capillaries were not affected. Immunostaining showed moderate to large numbers of CD3-positive cells (T lymphocytes), primarily within the tunica adventitia of affected arteries. Numerous adventitial MAC 387–positive cells were also present, which were interpreted to be macrophages and neutrophils. A few CD3- and MAC 387–positive cells were visible in the tunica media and tunica adventitia. CD3- and MAC 387–positive cells were present in both mild and more severe lesions. Only occasional CD79α-positive cells (B lymphocytes) were evident, primarily within the tunica adventitia. Other changes in the kidney included marked interstitial fibrosis, mild mesangial thickening, mild nonsuppurative interstitial nephritis, and tubular proteinosis.
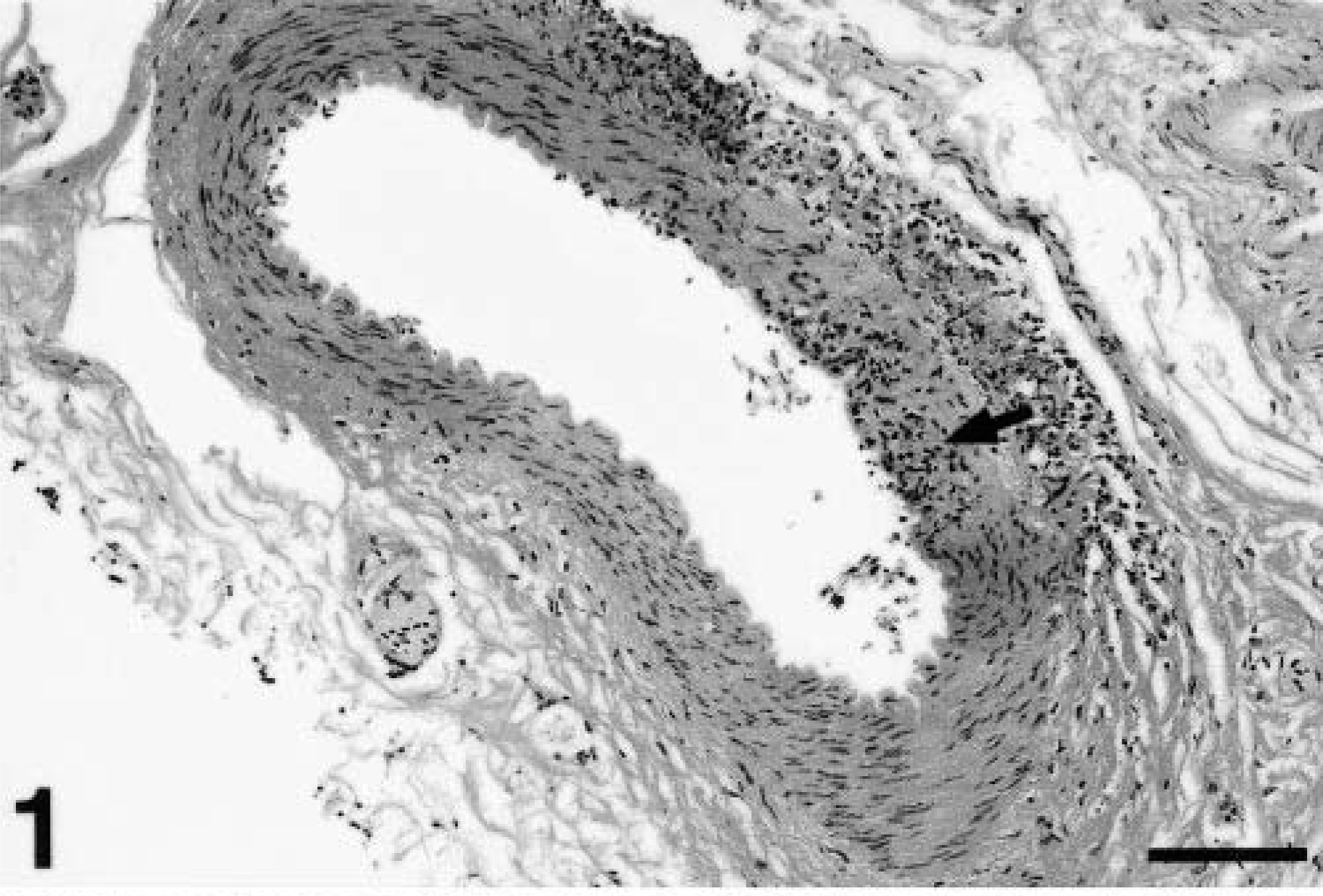
Mesenteric artery; cynomolgus macaque. Segmental inflammatory cell infiltrates are present within the tunica adventitia and tunica media (arrow). HE. Bar = 128 µm.
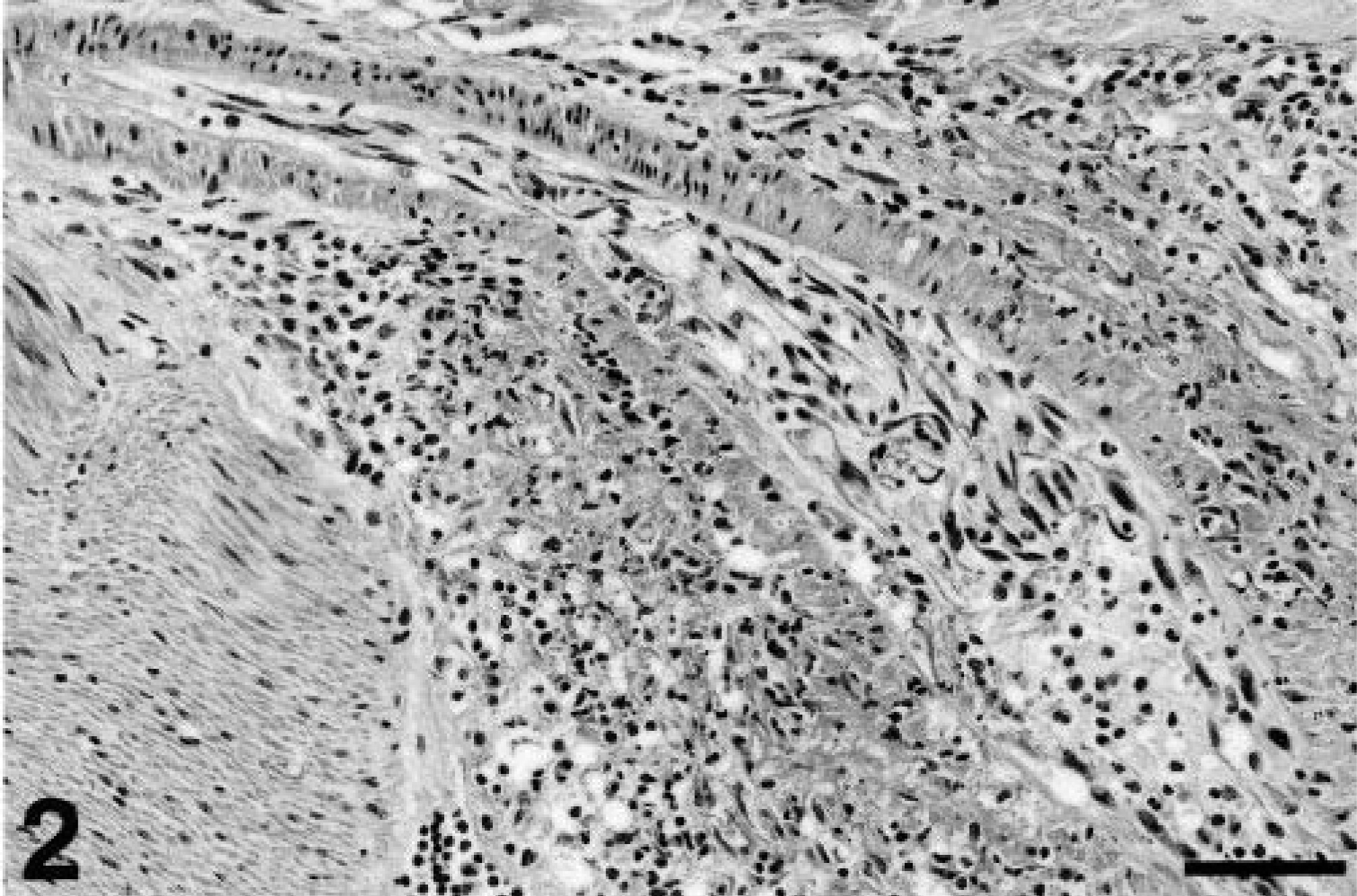
Small intestine; cynomolgus macaque. Inflammatory cell infiltrates are present within the tunica adventitia and tunica media, accompanied by mild fibrinoid necrosis and intimal proliferation. Note the normal vessel wall at the upper left. HE. Bar = 64 µm.
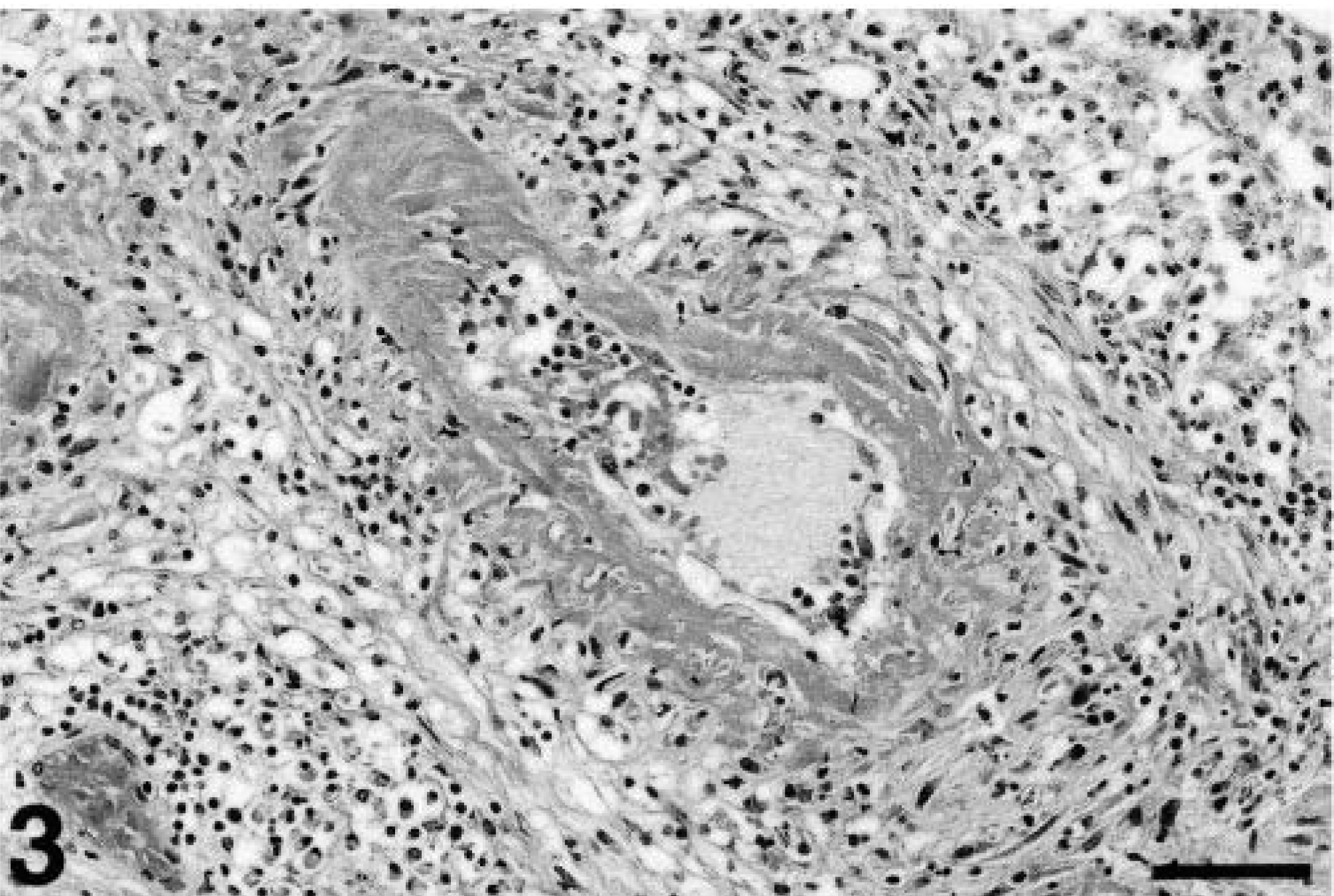
Small intestine; cynomolgus macaque. Marked fibrinoid necrosis is evident, obliterating the vessel wall and narrowing the lumen. Numerous lymphocytes, histiocytes, and fewer neutrophils, eosinophils, and plasma cells are present in the tunica adventitia and surrounding tissue. HE. Bar = 64 µm.
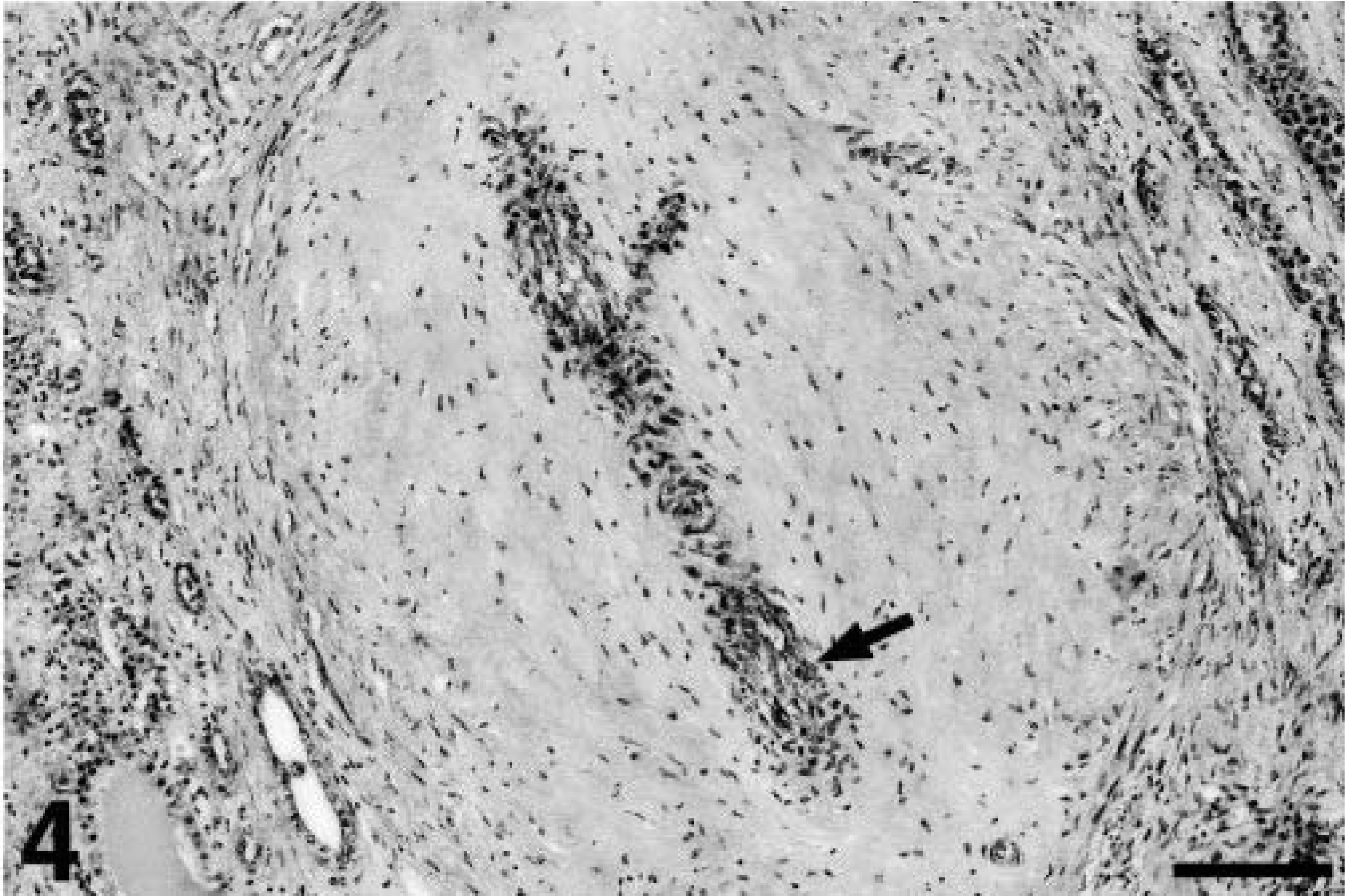
Kidney; cynomolgus macaque. The arterial wall is markedly thickened and sclerotic, with a very narrow residual lumen (arrow). Mild intimal proliferation is present. Marked fibrosis is evident in the adjacent cortical interstitium. Masson trichrome. Bar = 128 µm.
The pattern and character of the lesions in this macaque were very similar to those described in human cases of PAN. 6,8,12 Key features of PAN in humans are involvement of small- to medium-sized arteries, segmental distribution within affected arteries, a predilection for arterial branching points, and the coexistence of acute and chronic lesions. Lesions are most severe in the kidney, gastrointestinal tract, and heart, with sparing of the lung vasculature. In most cases, fibrinoid necrosis of the tunica media can be demonstrated, accompanied by a transmural mixed inflammatory infiltrate. Intimal proliferation, thrombosis, and aneurysmal dilation are often present. Possible manifestations of PAN in humans that were not observed in this case include peripheral neuropathy, seizures, distal gangrene of the extremities, and infarctions in multiple organs. 8 Renal involvement is a major cause of death in human PAN patients, 12 with renal failure occurring as a consequence of chronic renal ischemia. 8 Likewise, the animal in this case had clinical evidence of renal failure. Although urinalysis was not performed, urine protein loss was the presumptive cause of the hypoproteinemia and edema observed. In a human case of PAN, the inflammatory infiltrate included large numbers of T lymphocytes and macrophages. 5 Similarly, the inflammatory infiltrate in this case included many T lymphocytes and macrophages, which was confirmed with immunohistochemistry.
The findings described in the previous report of PAN in a cynomolgus macaque 1 are similar to those described here. The animal in the previous report had segmental arteritis of small- to medium-sized arteries in the kidney, heart, gall bladder, stomach, pancreas, large intestine, and uterus, with sparing of the pulmonary vessels. A significant difference between that case and the present one was the lack of fibrinoid necrosis in affected vessels. Glomeruli were hypercellular and contained increased matrix. Renal arterial sclerosis, interstitial fibrosis, and functional renal deficits were not evident. Cases of PAN-like disease in other species have been reported, and the term has occasionally been applied to conditions that differ significantly from the human disease. In rats, lesions occur most commonly as incidental findings in aged animals, with a predilection for mesenteric arteries. 4,16 The disease in mice has been reported in several different strains and has often included involvement of the aorta. 11,14,15 A polyarteritis syndrome has been recognized in juvenile Beagles. 13 This syndrome is characterized by fever and consistent involvement of cardiac, cranial mediastinal, and cervical meningeal vessels. Affected dogs often exhibit neck pain, causing the disease to be referred to as Beagle pain syndrome. Single case reports of PAN-like disease in a cat 2 and a pig 9 included pulmonary arterial involvement.
Inflammatory diseases affecting medium-sized arteries in humans include PAN, allergic granulomatosis and angiitis (Churg-Strauss syndrome), Kawasaki disease, and giant-cell arteritis. 6 Microscopic polyangiitis, a disease closely resembling PAN, is restricted to small vessels and includes glomerular and pulmonary involvement. 8 As with most forms of vasculitis, the pathogenesis of PAN is poorly understood and likely involves heterogeneous immunopathogenic mechanisms. 3,6,8,10 Most speculation centers on immune complex deposition, with subsequent activation of the complement cascade, neutrophil and monocyte chemotaxis, and the release of lysosomal enzymes, oxygen-free radicals, and proinflammatory mediators. The release of vasoactive amines from platelets likely increases vascular permeability, allowing intramural deposition of immune complexes. The inciting antigen in most cases of PAN is unknown, but 30% of human cases occur in patients with hepatitis B antigen in their serum. 12 The tendency for PAN lesions to occur at arterial branching sites may be the result of hydrostatic forces 3 or may relate to the increased expression of adhesion molecules and increased numbers of intimal macrophages at these sites. 10 Antineutrophil cytoplasmic antibodies have been identified in patients with some forms of vasculitis but are rare in PAN. 8,12 Cell-mediated mechanisms likely contribute to the pathogenesis. Patients with PAN have been shown to have marked elevations in serum levels of interferon-γ and interleukin-2, 7 and an immunohistochemical study showed that the inflammatory infiltrates in PAN lesions were composed primarily of macrophages and T cells, mostly CD4+ cells. 5
The cause for marked regenerative anemia in this case was not identified. Renal failure likely contributed, but the severity of the anemia suggests that other factors may have played a role. Vasculitis-induced hemorrhage is a consideration, but significant hemorrhages were not evident at necropsy. A nosebleed was observed on day 8, and thrombocytopenia was present on days 25 and 33. Hemostasis tests, examination of bone marrow, and testing for simian parvovirus, a recognized cause of anemia in cynomolgus macaques, were not performed. Blood smears revealed marked anisocytosis of erythrocytes and no evidence of blood parasites. Reticulocytosis was the presumptive cause of the elevated mean corpuscular volume. In human PAN patients, anemia is a common but poorly characterized finding. 8 Gastrointestinal hemorrhage is evident in some cases of human PAN 8 but was not observed in this case. Anemia was not noted in the previous case of PAN in a cynomolgus macaque. 1
The pathogenesis of PAN remains enigmatic, and a reliable animal model of the disease has not been identified. Of the reported cases of PAN-like syndromes in animals, the disease in cynomolgus macaques appears to most closely resemble the human disease. However, the sporadic nature of the disease in this species prevents the development of a useful animal model.
Footnotes
Acknowledgements
We appreciate the help of Dr. Sean McDonough and Dr. Clive Huxtable in interpretation of the immunohistochemical stains and renal changes, respectively. We also thank Pat Fisher for her expertise with immunohistochemistry and Alexis Wenski-Roberts of the Cornell University Image Lab for assistance with photography.