
Abstract
The effects of immunosuppression were examined in 1.5-month-old calves that were given dexamethasone (DM) before endobronchial inoculation with bovine adenovirus type 3 (BAV-3). Immunohistopathologically, severe necrotizing bronchiolitis with eosinophilic and basophilic intranuclear inclusion bodies was observed both in DM-treated 1.5-month-old infected calves and in non-DM-treated 7-day-old infected calves. These inclusion bodies were correlated with the detection of BAV-3 antigen and viral particles. The presence of inclusion bodies in the desquamated epithelial cells or of BAV-3 antigen, or both, correlated well with the isolated level of BAV-3 in bronchoalveolar lavage (BAL) fluid. Few immunoglobulin (IgG, IgM, and IgA)-containing B lymphocytes or CD8+ T lymphocytes infiltrated the pneumonic lesion in both the 7-day-old and the DM-treated 1.5-month-old infected calves. Thus, depletion of CD8+ T lymphocytes in calves might influence the clearance of BAV-3 from respiratory tissues.
Keywords
Introduction
Disseminated adenoviral infection is frequently as sociated with fatal interstitial pneumonia in transplant recipients, children, and young animals. 2–4 , 8 , 10 , 13 , 21 , 25 In calves, two types (types 3 and 5) of adenovirus appear to be more pathogenic than others, producing disease concentrated in the respiratory and gastrointestinal tracts of these animals. 5–7 , 9 , 11 The occurrence of respiratory adenoviral infection has been reported in colostrum-deprived calves experimentally infected with bovine adenovirus type 3 (BAV-3). 5–7 However, these calves failed to produce signs of disease when they were at 1 week of age or older, except those that had undergone endobronchial inoculation with BAV-3. 19 In considering the pathogenesis of BAV-3 in calves, it is interesting to investigate what factors in addition to age correlate with susceptibility.
The purpose of this study was to examine respiratory tissues from 7-day-old, dexamethasone (DM)-treated 1.5-month-old, and non–DM-treated 1.5-month-old calves inoculated endobronchially (EB) with BAV-3. Pathologic changes and the distribution of BAV-3 antigen, IgG-, IgM-, and IgA-containing B cells, and CD4+ and CD8+ T lymphocytes in the pneumonic lesions were described using an immunoperoxidase technique. We also examined bronchoalveolar lavage (BAL) fluid for virus isolation, detection of viral inclusion bodies, and BAV-3 antigen in the desquamated epithelial cells.
Materials and Methods
Virus
BAV-3 was isolated from a calf in Japan that exhibited signs of pneumonia, coughing, and serious rhinitis. After plaque purification, the virus was identified as BAV by biophysical characterization and more specifically as BAV-3 by a cross-neutralization test. After the virus was propagated in bottle cultures of Madin-Darby bovine kidney (MDBK) cells, aliquots of the virus were kept frozen (at −80 C) until they were used as inoculum. Bovine viral diarrhea virus-free was confirmed by the methods described previously. 19 , 24
Animals
Twelve male Holstein calves were used in this experiment and were divided into four groups (Tables 1 and 2). Group 1 consisted of three 7-day-old infected calves (calf Nos. 1–3); groups 2 (calf Nos. 4–6) and 3 (calf Nos. 7–9) consisted of six 1.5-month-old infected calves; and group 4 consisted of three uninfected calves used as controls—one 7-day-old (calf No. 10) and two 1.5-month-old (calf Nos. 11 and 12). All 12 calves had received colostrum within a few hours of birth and had neutralizing antibody against BAV-3 at levels ranging from 1:4 to 1:64. The nine experimental calves were kept in separate isolation pens throughout the experiment, and the three control calves were likewise kept in similar pens to prevent cross-infection.
Cytologic and virologic analyses of BAL fluid cells.
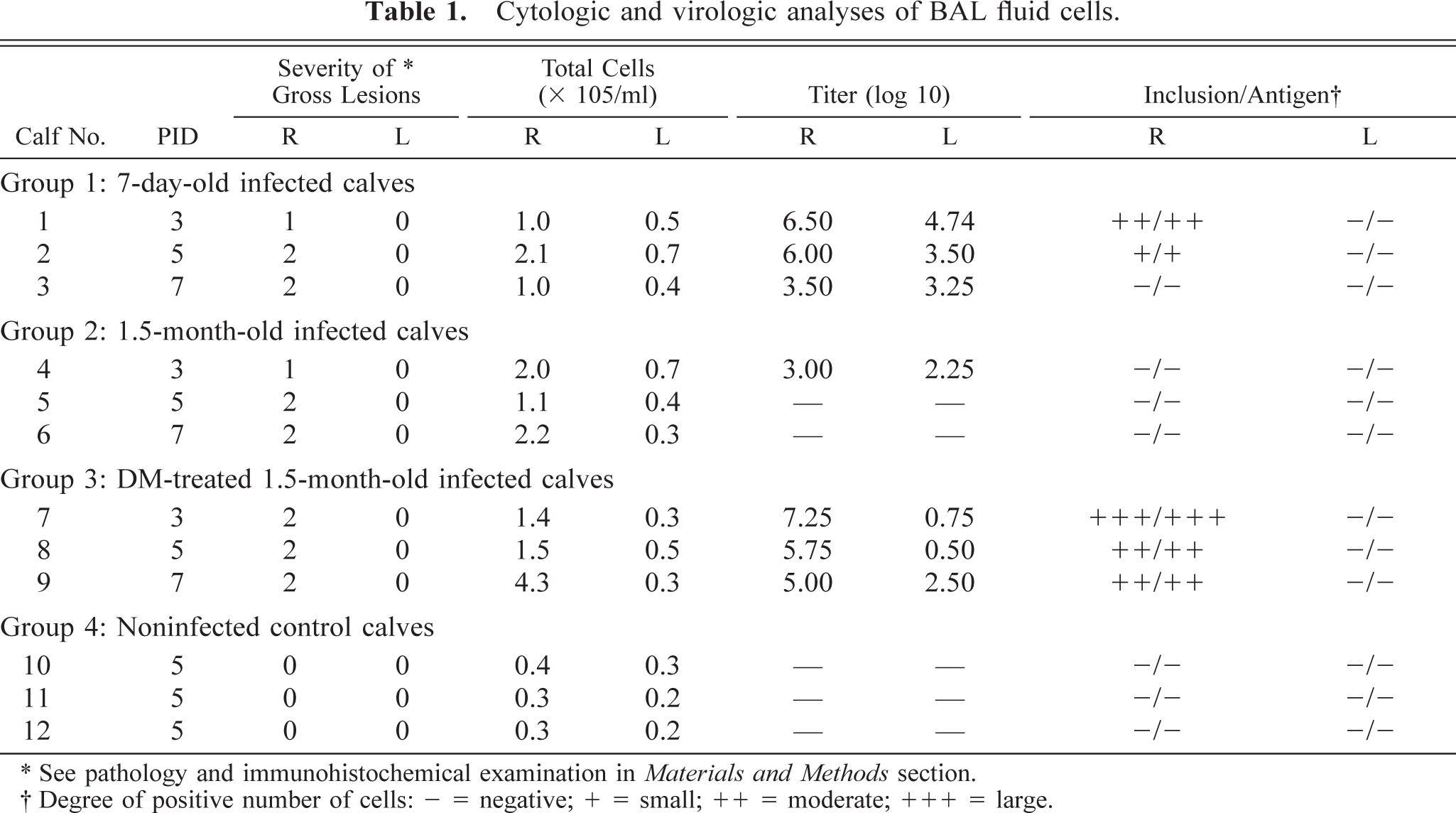
∗ See pathology and immunohistochemical examination in Materials and Methods section.
† Degree of positive number of cells: − = negative; + = small; ++ = moderate; +++ = large.
Distribution of histologic lesions, BAV-3 antigen, and T-lymphocytes in the lungs.
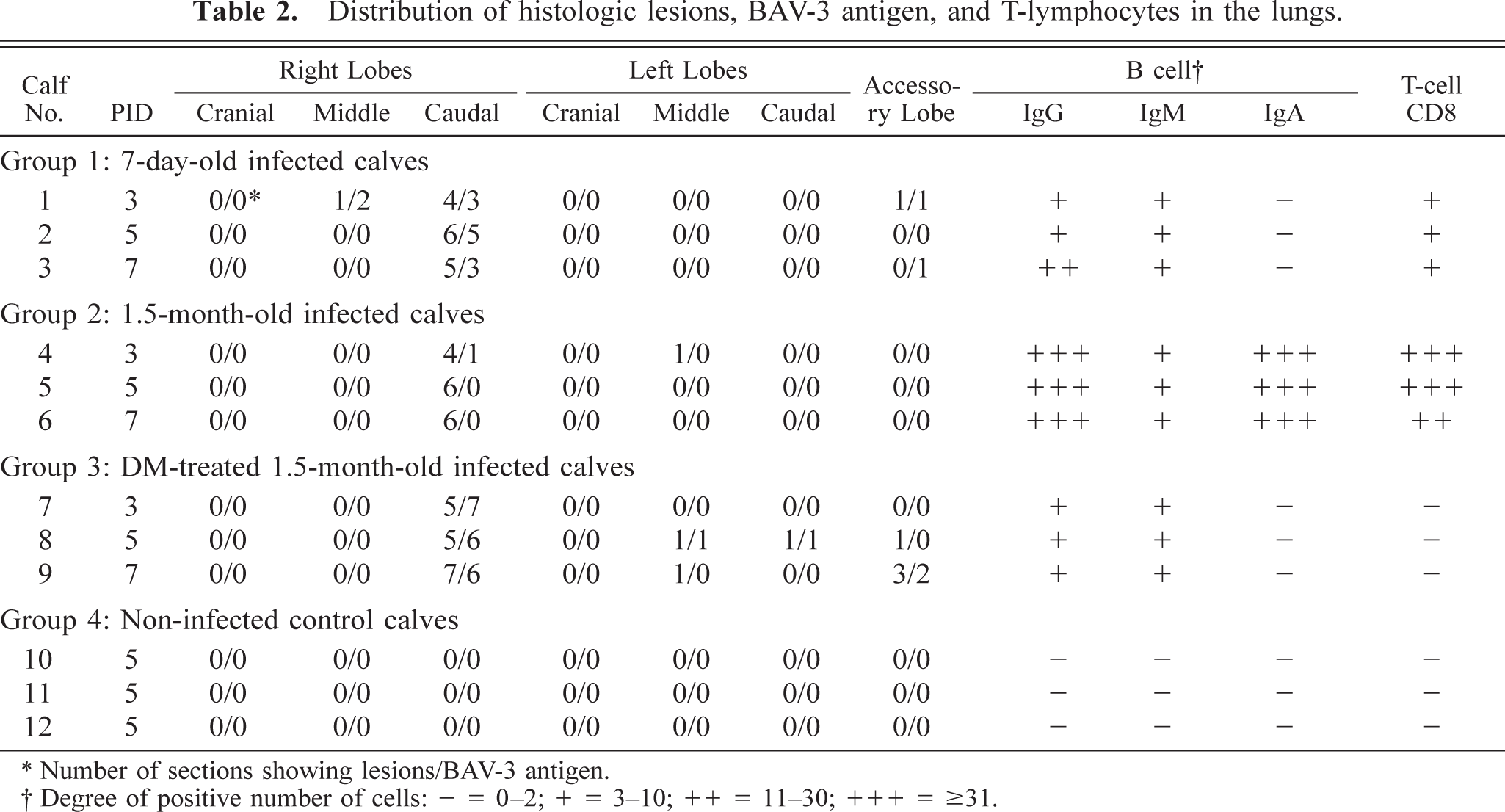
∗ Number of sections showing lesions/BAV-3 antigen.
† Degree of positive number of cells: − = 0–2; + = 3–10; ++ = 11–30; +++ = ≥31.
Experimental procedure
Four 1.5-month-old calves (calf Nos. 7–9 and 12) were injected intravenously with seven consecutive daily doses of 0.1 mg of DM/kg of body weight. On day 5 of DM treatment, three of these calves (calf Nos. 7–9) were inoculated EB with BAV-3. Before inoculation with BAV-3, all 12 calves were given atropine hydrochloride (2 mg in a 4-ml dose volume, intramuscularly) and were anaesthetized with xylazin (2 ml/head, intravenously). The six calves in groups 1 and 2 (calf Nos. 1–6) and the three 5th DM-treated calves in group 3 (calf Nos. 7–9) were inoculated EB with 10 ml (105.0 TCID50/0.1 ml) of viral suspension by means of a bronchofibroscope (Olympus type 10; Olympus, Tokyo, Japan). This instrument was inserted into the right main bronchus of the caudal lobe. The procedure was completed within 10 minutes for each calf. One calf from each group was killed by an intravenous overdose of barbiturates on each of the three postinoculation days (PIDs), days 3, 5, and 7. The three control calves in group 4 (calf Nos. 10–12), including 5th DM-treated calf (calf No. 12), also were inoculated EB with 10 ml of noninfected maintenance medium and killed on PID 5. The clinical signs and body temperatures of all calves were observed at least twice daily during the experiment.
Cytologic evaluation of BAL fluid
After removal of the lungs at necropsy, a cannula (4 mm in diameter) was inserted into each of the right and left bronchi on the caudal lobes. Thirty milliliters of sterile phosphate-buffered saline was then introduced and immediately withdrawn. The BAL fluid was recovered, 20 ml was placed in a sterile bottle, and the number of nucleated cells per microliter of BAL was counted. One slide prepared from the centrifuged cells was stained with a Diff-Quik kit (International Reagent Corporation, Kobe, Japan) and another was fixed with cold acetone for 5 minutes and stored at 20 C for subsequent immunohistochemical examination. Evaluation included a 200-cell differential count and a morphologic description of the cells.
Viral isolation
BAL fluid samples (1 ml each) collected separately from the right and left caudal lobes were centrifuged at 3,000 rpm, and 1 ml of maintenance medium without bovine serum was added to each cell pellet. The cells were then stored at −80 C until use. The samples were subjected to ultrasonic disruption for 20 seconds at 4 C, and 0.1 ml decimal dilution of each sample was inoculated onto confluent cultures of MDBK cells in a 96-well microtitre plate. The cells were examined daily for 1 week for cytopathogenic effects to determine the presence and titer (TCID50) of BAV-3.
Histopathology and immunohistochemistry
All calves was subjected to necropsy, and the extent of pneumonia in the caudal lobe was recorded as follows: 0, nil; 1, 1–25%; 2, 26–50%; 3, 51–75%; and 4, 76–100%.
Specimens from each calf were collected and fixed in buffered 10% formalin. Three tissue samples of the right and left cranial lobes were collected from uniform sites from each calf, in addition to three from each of the right and left middle lobes, nine from each of the right and left caudal lobes, and three from the accessory lobe. All specimens were embedded in paraffin wax, and sections were cut and stained with hematoxylin and eosin (HE staining).
The presence of BAV-3 antigens in the formalin-fixed, paraffin wax–embedded tissues and cold acetone-fixed BAL fluid cells was demonstrated by the strept–avidin–biotin complex immunoperoxidase (SAB-IP) method, using an SAB kit (Nichirei Corporation, Tokyo, Japan). Anti–BAV-3 rabbit serum was used as the primary antibody at a dilution of 1:8,000. 19 The presence of antibovine IgG, IgM, and IgA in the pneumonic lesions in paraffin sections was demonstrated using a simple stain MAXPO kit (Nichirei Corp.). Rabbit antibovine IgG-Fab fragment (Bethyl Laboratories Inc., Montgomery, TX), rabbit anti-IgM mu chain specific (Bethyl Laboratories Inc.), and rabbit antibovine IgA-alpha chain specific (Bethyl Laboratories Inc.) were used as the primary antibodies at a dilution of 1:2,000 each. The presence of CD4+ and CD8+ T lymphocytes in three parts of the pneumonic lesions from right caudal lobe of the cryostat sections was demonstrated by using SAB kit (Nichirei Corporation). Antibovine CD4 (VMRD Inc., Pullman, WA) and anti-CD8 (VMRD Inc.) monoclonal antibodies were used at dilution of 1:200 and 1:1,600, respectively. The samples were counterstained with methyl green. Tissue sections from the uninfected control calves and sera from nonimmunized rabbits and mice were used as controls. The mean positive number of B and T lymphocytes was counted from five fields of 200×.
Small portions of the pneumonic lesions from the right caudal lobes were prepared for electron microscopy by post-fixing them in 1% osmium tetroxide, then dehydrating and embedding them in Epon mixture. Ultrathin sections were stained with uranyl acetate and lead citrate and examined with a TEM-100 CX electron microscope.
Results
Clinical observations
None of the calves in any group showed clinical evidence for respiratory disease or diarrhea at any time during the course of the experiments. Several infected calves in each group exhibited a slight rise in rectal temperature (more than 39.5 C) that lasted for 1–2 days. The control calves remained normal.
Analysis of BAL cells
Table 1 shows the results of the analysis of BAL cells. The total number of cells in the BAL fluid from the right and left caudal lobes ranged from 0.2 × 105.0/ml to 0.4 × 105.0/ml in the noninfected control calves (group 4, calf Nos. 10–12). In group 4, the percentage of macrophage in the right and left lobes was 90–93% and that of neutrophil was 2–3%. The total cell number from the right caudal lobe had increased 2–5 times (in comparison with control levels) in group 1 and group 2 infected calves (calf Nos. 1–6) and 3–9 times in group 3 infected calves (calf Nos. 7–9). The ratio of neutrophils from the right caudal lobe had increased greatly in groups 1–3, giving levels of 11–28%, 10–30%, and 15–32%, respectively. The ratio of desquamated cells from the right caudal lobe was 13–16% in group 1, 10–12% in group 2, and 11–36% in group 3. The total number of cells from the left caudal lobe in groups 1–3 ranged from 0.3 × 105.0 to 0.7 × 105.0 and did not increase the number of neutrophils.
Cytology
Eosinophilic and basophilic intranuclear inclusion bodies from the right caudal lobe were found in the desquamated epithelial cells of two calves from group 1 (calf Nos. 1 and 2) and of all three calves from group 3 (calf Nos. 7–9) but were not found in cells of any calf from group 2 or from the three noninfected controls (Fig. 1). No inclusion bodies were observed in the left caudal lobe of any of the infected calves. Immunohistochemical examination revealed a high number of BAV-3 antigens in the nuclei and cytoplasm of the desquamated epithelial cells, a finding that corresponds well to the detection of intranuclear inclusion bodies. No such inclusion bodies were detected in the left caudal lobes of any of the infected or control calves.
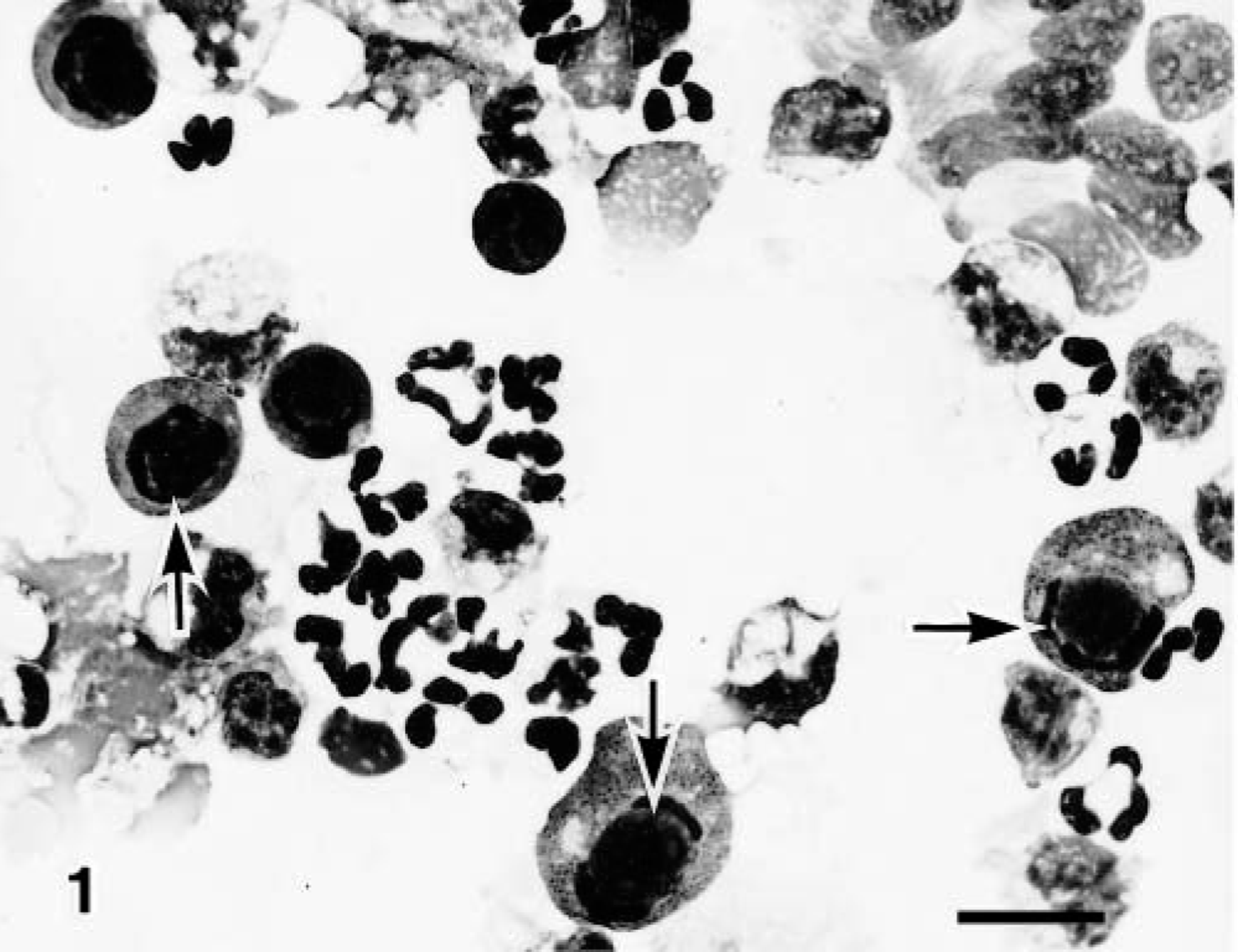
Bronchoalveolar fluid cells of the right caudal lobe; calf No. 7. Eosinophilic intranuclear inclusion bodies (arrows) in the desquamated epithelial cells. Diff-Quik. Bar = 15 μm.
BAV-3 was isolated from BAL fluid cells of the right and left caudal lobes of all three group 1 calves (calf Nos. 1–3), one group 2 calf (calf No. 4), and all three group 3 calves (calf Nos. 7–9) but was not detected in two of the infected group 2 calves (calf Nos. 5 and 6) or in the three noninfected controls (group 4, calf Nos. 10–12). The highest titers were 106.5 TCID50/ml in group 1, 103.0 TCID50/ml in group 2, and 107.25 TCID50/ml in group 3 (Table 1).
Pathology
Gross lesions were confined to the lungs of the virus-inoculated side (Table 1). Dark red, depressed, irregular areas of consolidation, primarily located in the caudal area, were found in the right caudal lobe in all infected calves but not in the left caudal lobe (Fig. 2). By PID 7, the lesions in calf No. 6 were found to be smaller than those in other infected calves. The lungs of the noninfected calves were normal in appearance. The bronchiolar and mediastinal lymph nodes were slightly swollen and congested in all infected calves. Moreover, the three DM-treated infected calves (calf Nos. 7–9) and the single DM-treated noninfected control calf (calf No. 12) showed severe atrophy of the thymus.
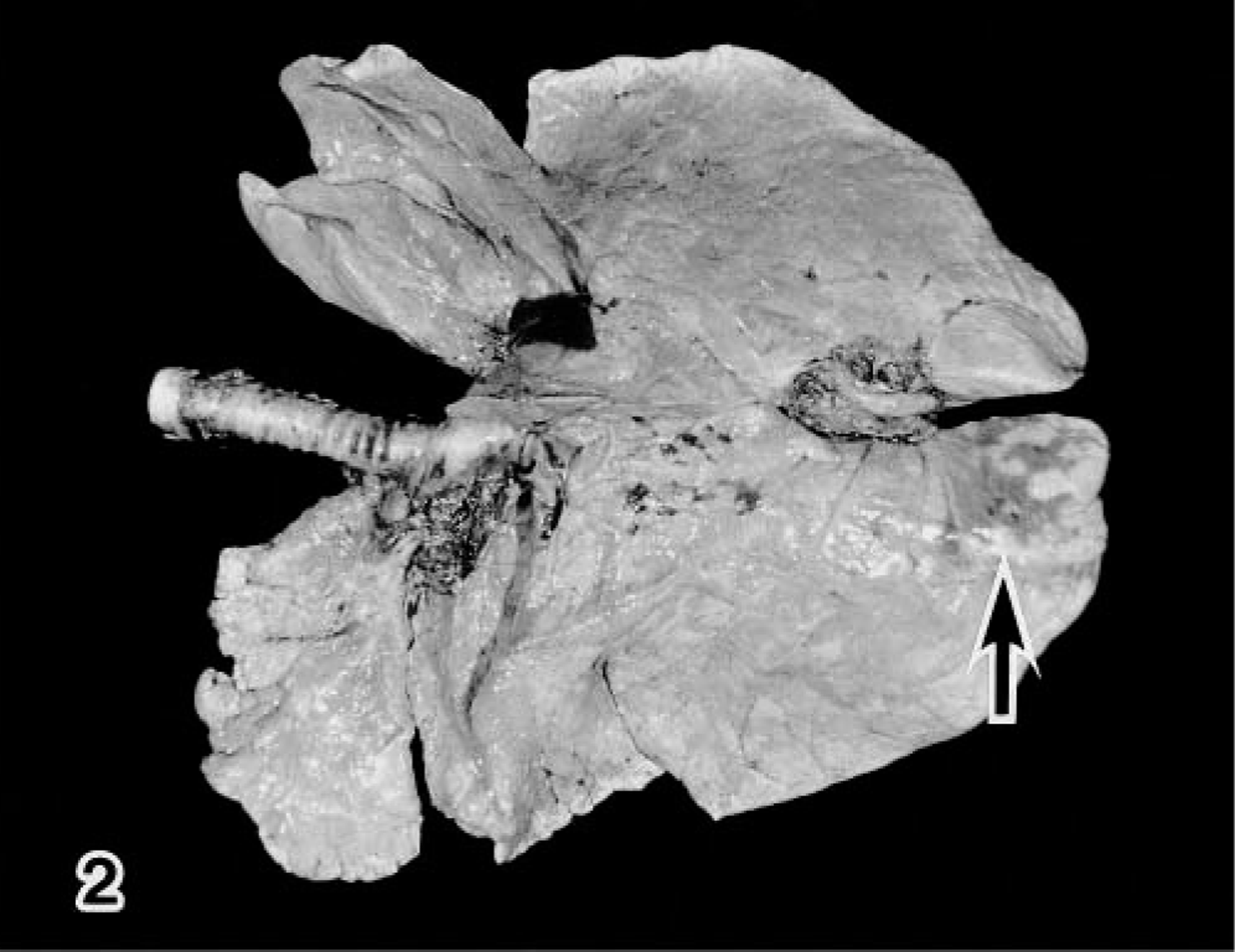
Lung; calf No. 7. Pneumonic lesion (arrow) is located in the caudal part of the right caudal lobe.
In groups 1 and 3, the characteristic pneumonic lesions in experimentally infected calves appeared as an acute, necrotizing, exfoliative bronchiolitis and bronchitis with intranuclear inclusion bodies (see Table 2, Fig. 3). There was extensive necrosis and desquamation of the respiratory epithelium in affected airways, with scant lining epithelial cells remaining intact and viable. The lumina were frequently occluded by cellular and streaming nuclear debris, admixed with degenerative leukocytes and fibrin. The recognizable, sloughed epithelium consisted of deeply eosinophilic, basophilic, round to ovoid, intranuclear inclusion bodies with margination of nuclear chromatin. The surrounding parenchymal cells were markedly congested with a light, diffuse infiltration of alveolar interstitium by inflammatory cells, primarily lymphocytes with plasma cells, macrophages, and neutrophils. By PIDs 5 and 7, many intranuclear inclusion bodies were found in the alveolar pneumocytes in the parenchymal pneumonic lesions in DM-treated infected calves (calf Nos. 8 and 9).
Lung; calf No. 7. Degenerated bronchilar epithelial cells (arrows) contained eosinophilic intranuclear inclusion bodies. HE. Bar = 15 μm.
In group 2, moderate proliferative bronchitis and bronchiolitis were found, accompanied by epithelial hyperplastic lesions and severe infiltration of leukocytes and lymphocytes with thickening of the alveolar wall. By PIDs 5 and 7, calf Nos. 5 and 6 (calf No. 4 had been killed on PID 3) exhibited varying degrees of peribronchial and perivascular cuffings (Fig. 4). However, no inclusion bodies were found in any of the infected group 2 calves (calf Nos. 4–6).
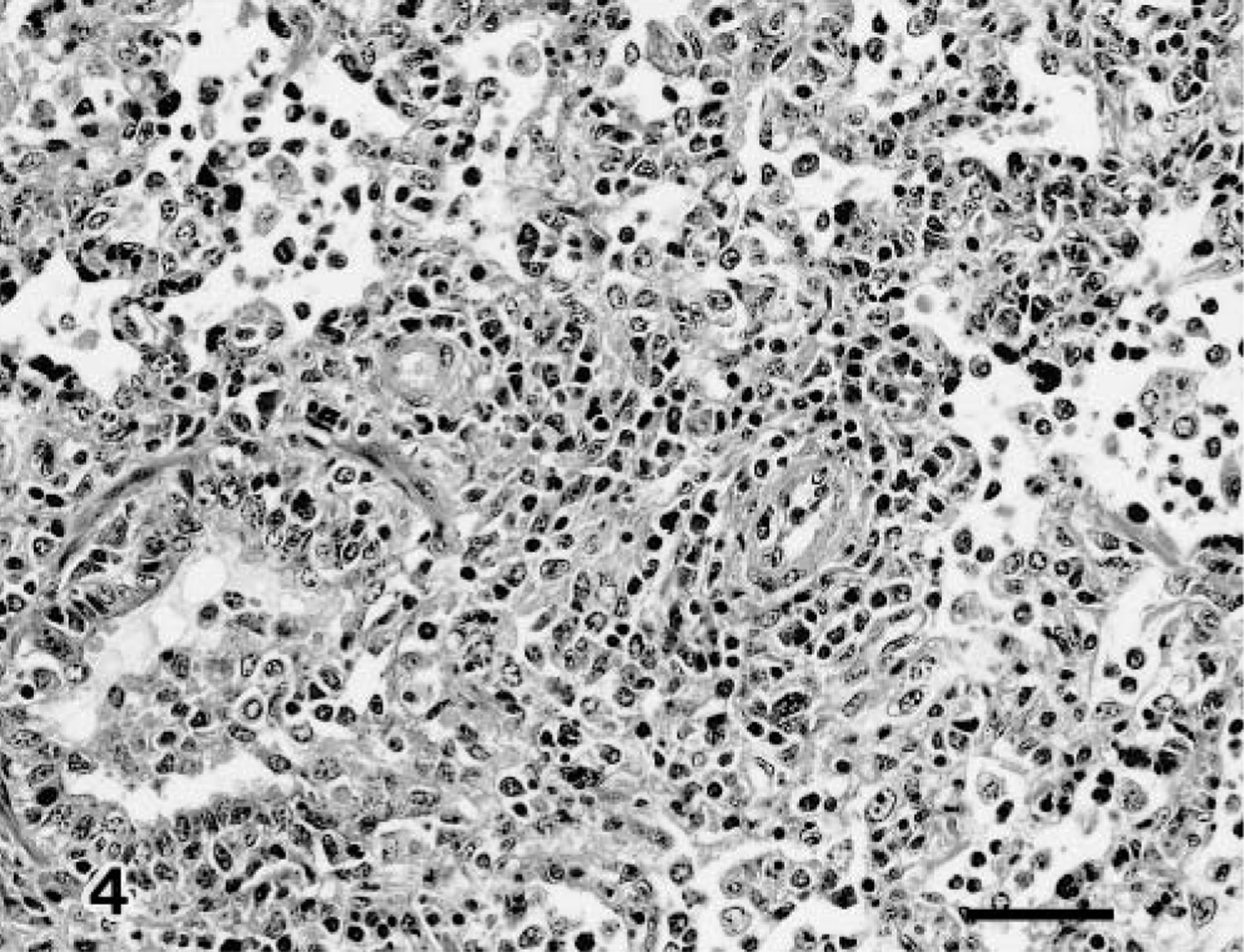
Lung; calf No. 6. Interstitial pneumonia with peribronchial and perivascular cuffing. HE. Bar = 50 μm.
In addition to pneumonia, a slight suppurative lymphadenitis was observed in the bronchial and mediastinal lymph nodes of all infected calves. Severe atrophy of the thymus was observed in the four DM-treated calves (calf Nos. 7–9 and 12). Neither gross nor histopathologic changes was found in the intestinal mucous membrane or brain of any of the infected animals (calf Nos. 1–9). No significant pneumonic lesions were found in the three control calves (calf Nos. 10–12).
Electron microscopic examination
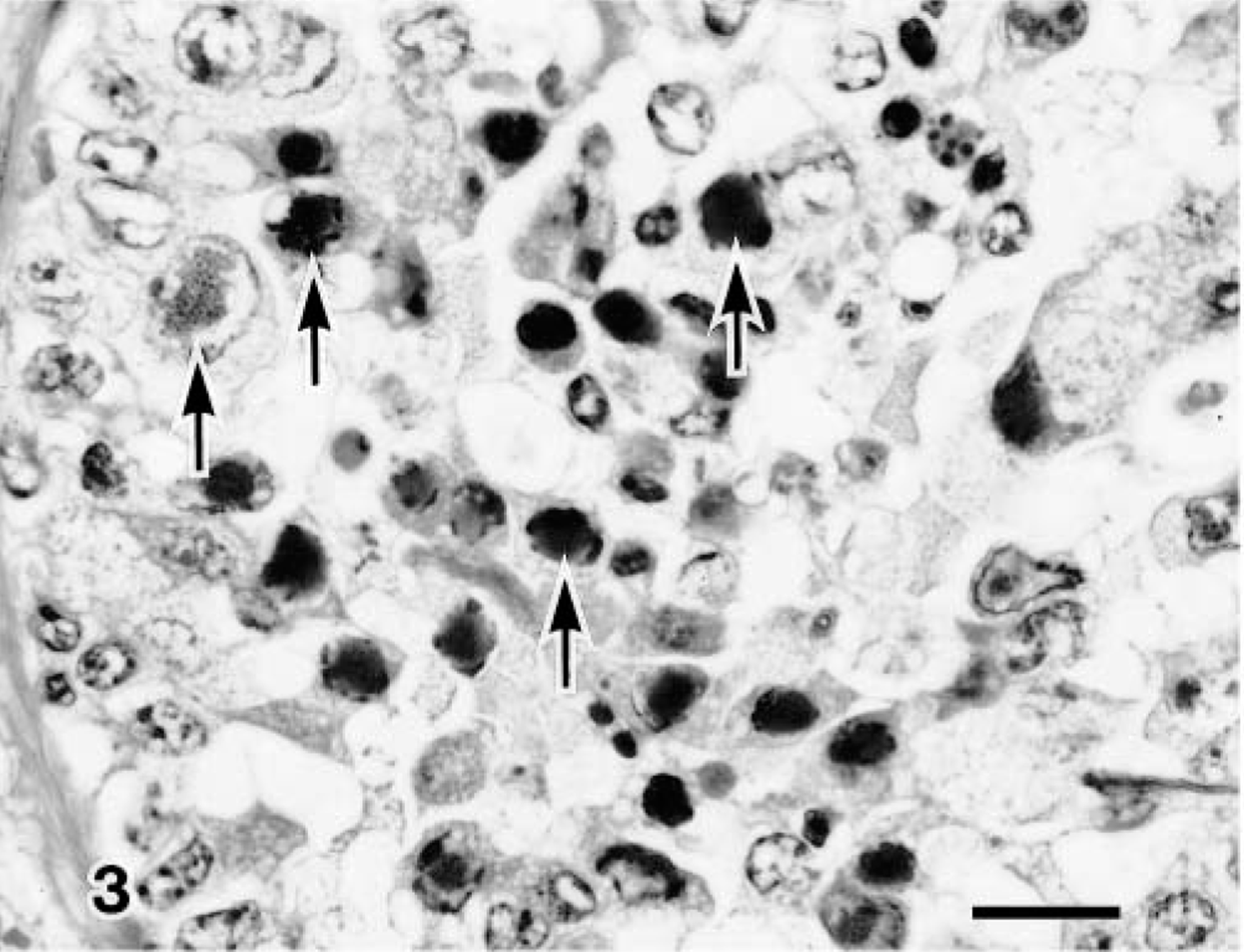
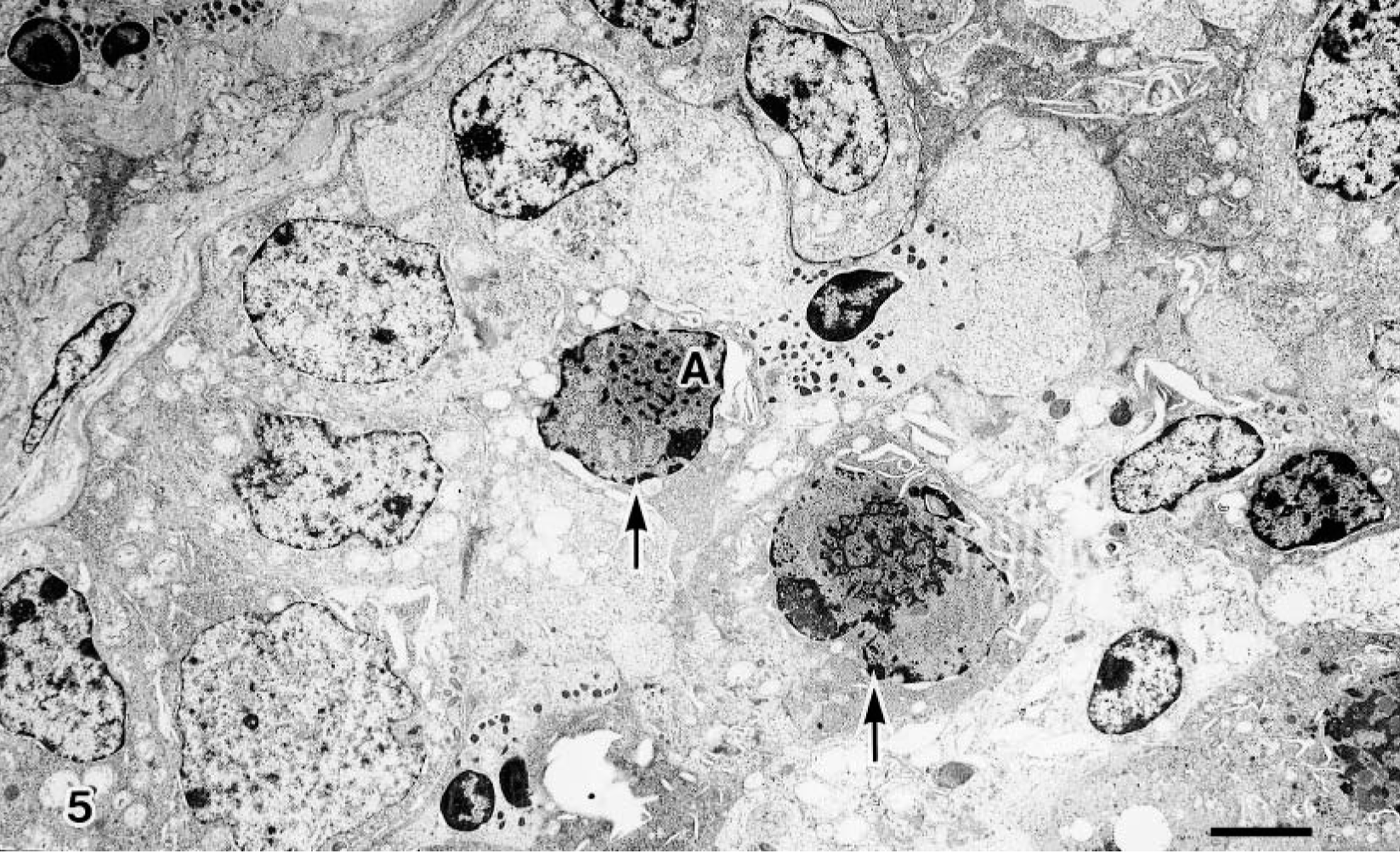
BAV-3–infected terminal bronchiolar epithelial cells showed many intranuclear inclusion bodies, revealing cellular degeneration, manifested by undulation of the nuclear membrane and margination of nuclear chromatin (Fig. 5). Neutrophils and macrophages were infiltrated in areas adjacent to cell degeneration. Virus particles, approximately 70 nm in diameter, were found in the surface and desquamated areas of bronchiolar epithelial cells and occasionally in membrane-bound vacuoles in the cytoplasm (Fig. 6).
Transmission electron micrograph. Lung; calf No. 7. Viral inclusion bodies (arrows) are found in the desquamated bronchiolar epithelial cells. Bar = 3 μm.
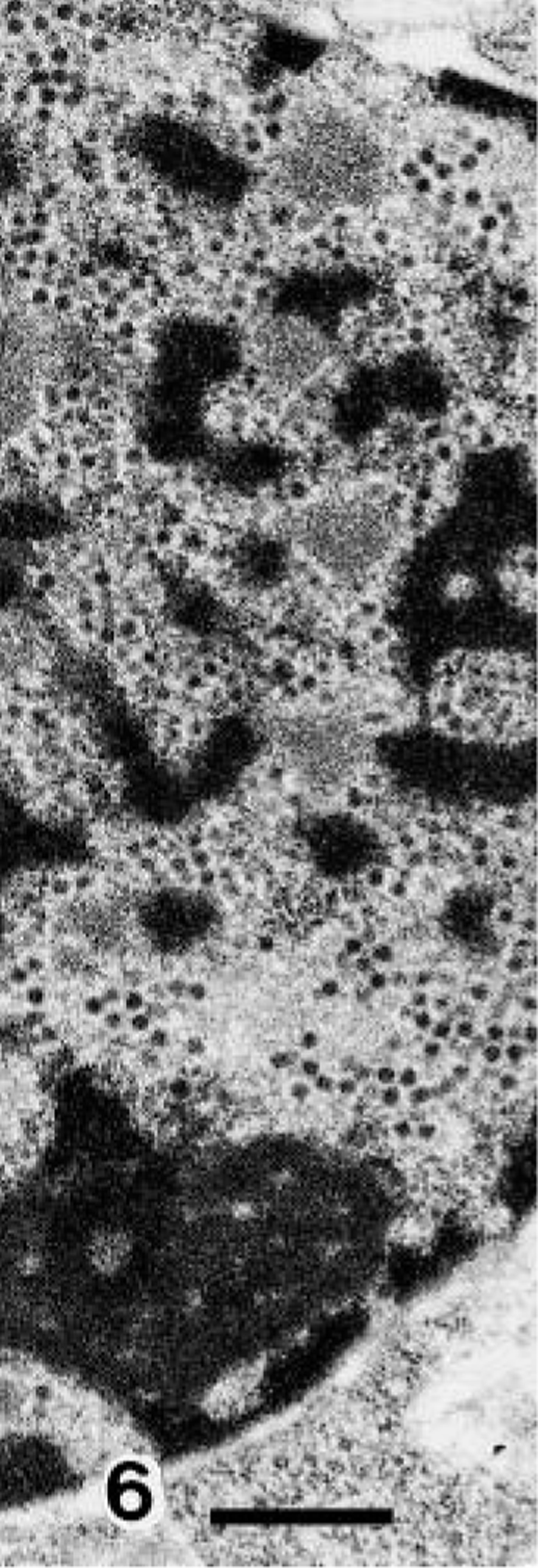
Transmission electron micrograph. Lung; calf No. 7. High magnification of cell A in Fig. 6. Viral particles are loosely arranged. Bar = 500 μm.
Immunohistochemistry
The BAV-3 antigen in the infected calves of groups 1 and 3 (calf Nos. 1–3 and 7–9, respectively) was first detected in the epithelial cells in the bronchioles (see Table 2, Fig. 7). BAV-3 antigen was observed in calves killed after PID 5 in neighboring alveolar macrophages and alveoli. The positive number of BAV-3 antigens was decreased in calves examined at PID 7. No BAV-3 antigen was found in group 2 infected calves (calf Nos. 4–6) or in group 4 noninfected calves (calf Nos. 10–12).
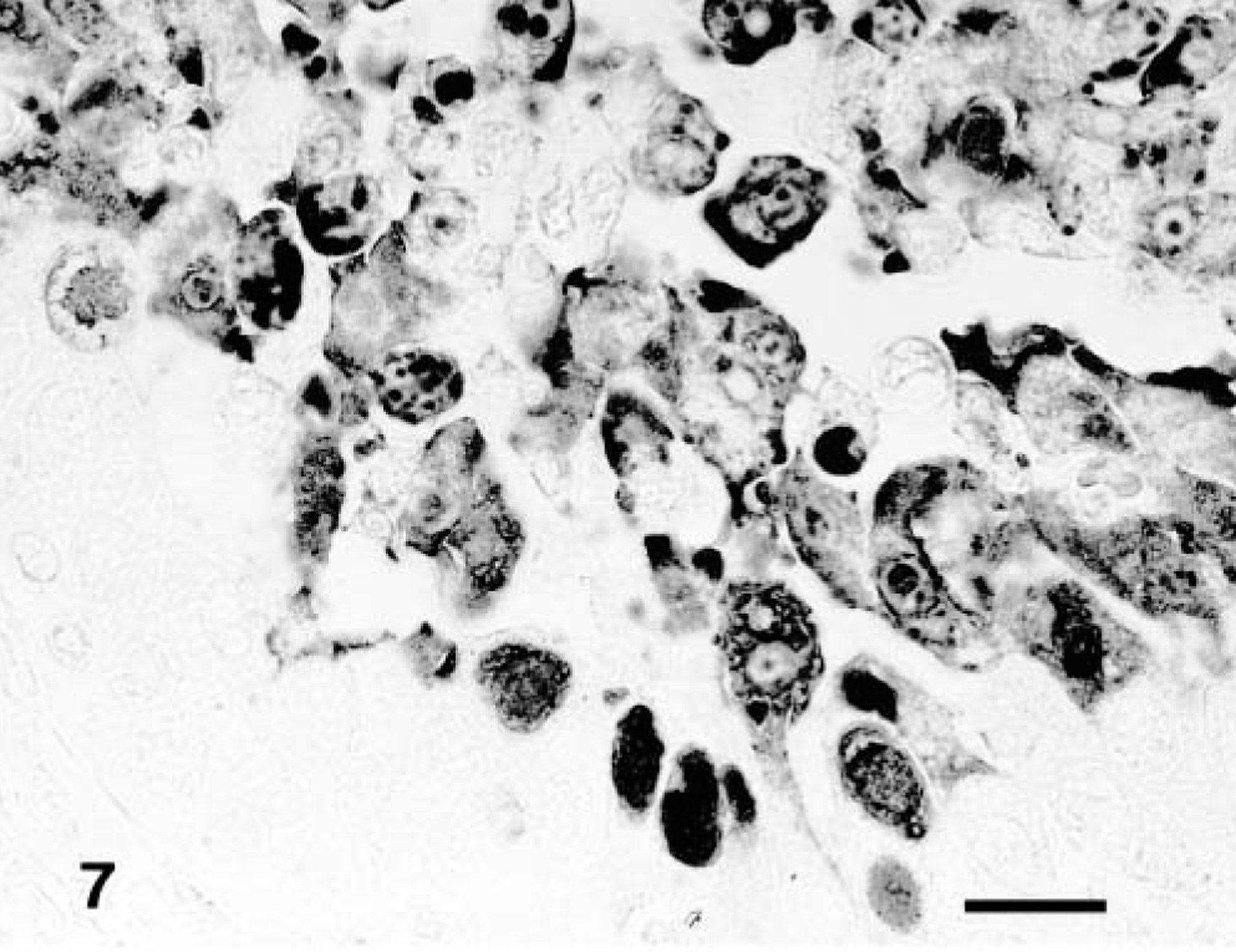
Lung; calf No. 7. BAV-3 antigen is detected in the bronchiolar epithelial cells. SAB-IP. Bar = 15 μm.
The IgG- and IgM-containing cells were first detected in the pneumonic lesions in all infected claves but not IgA-containing cells in groups 1 and 3. Immunoglobulin-containing cells were scattered and other cells were aggregated, coincident with the inflammatory response. Moreover, the positive number of immunoglobulin-containing cells in group 2 had increased significantly more than those of groups 1 and 3 (Table 2) (Figs. 8, 9). The number of CD8+ T lymphocytes in the pneumonic lesions were higher in calves of group 2 than in those of groups 1 or 3 (Fig. 10). A low number of CD4+ T lymphocytes was detected in the pneumonic lesions in all infected calves, but this number did not differ substantially among the groups of infected calves. Neither CD8+ nor CD4+ T lymphocytes was present in the three noninfected calves (Table 2).
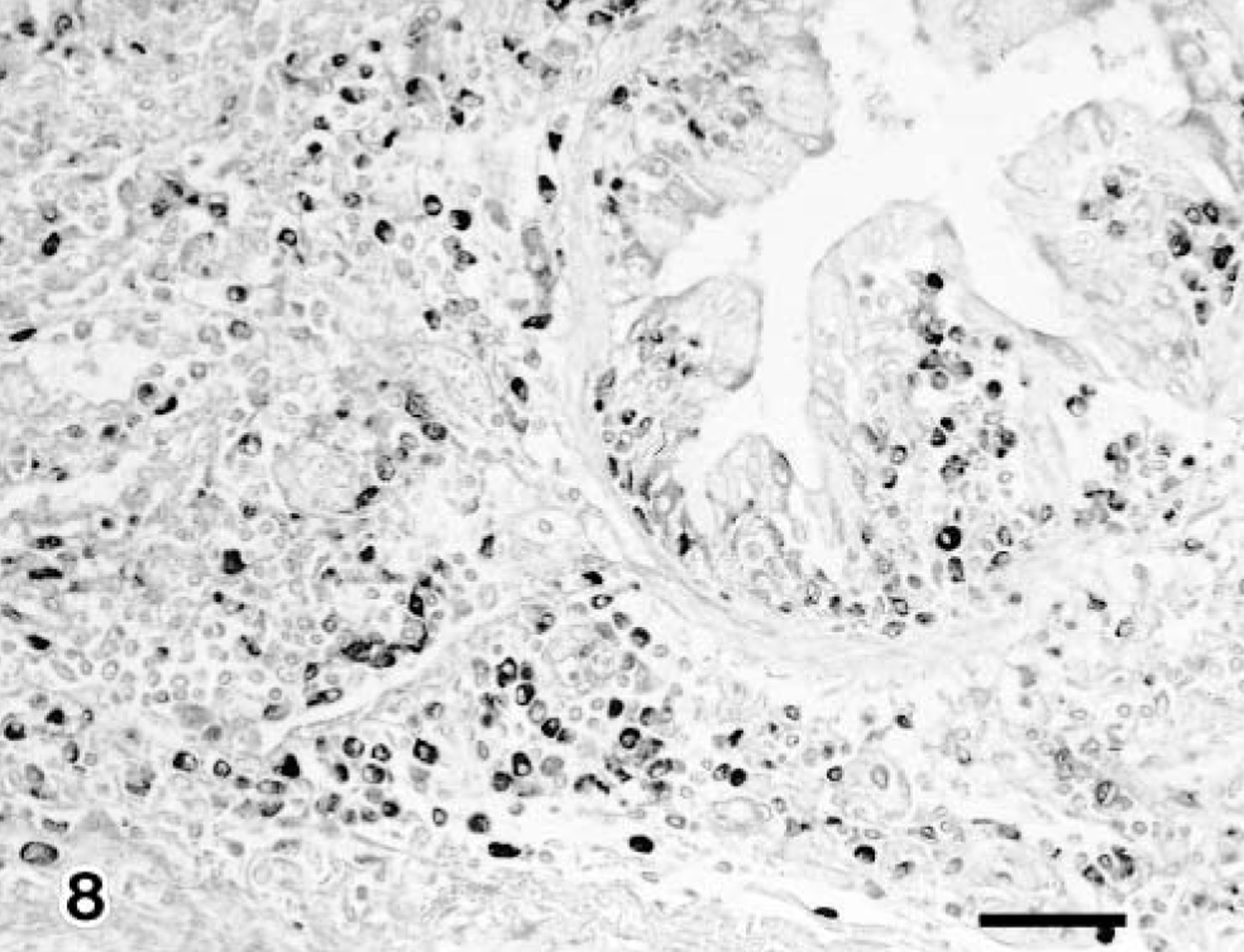
Lung; calf No. 5. Many IgG-containing cells are detected in the peribronchiolar area. SAB-IP. Bar = 50 μm.
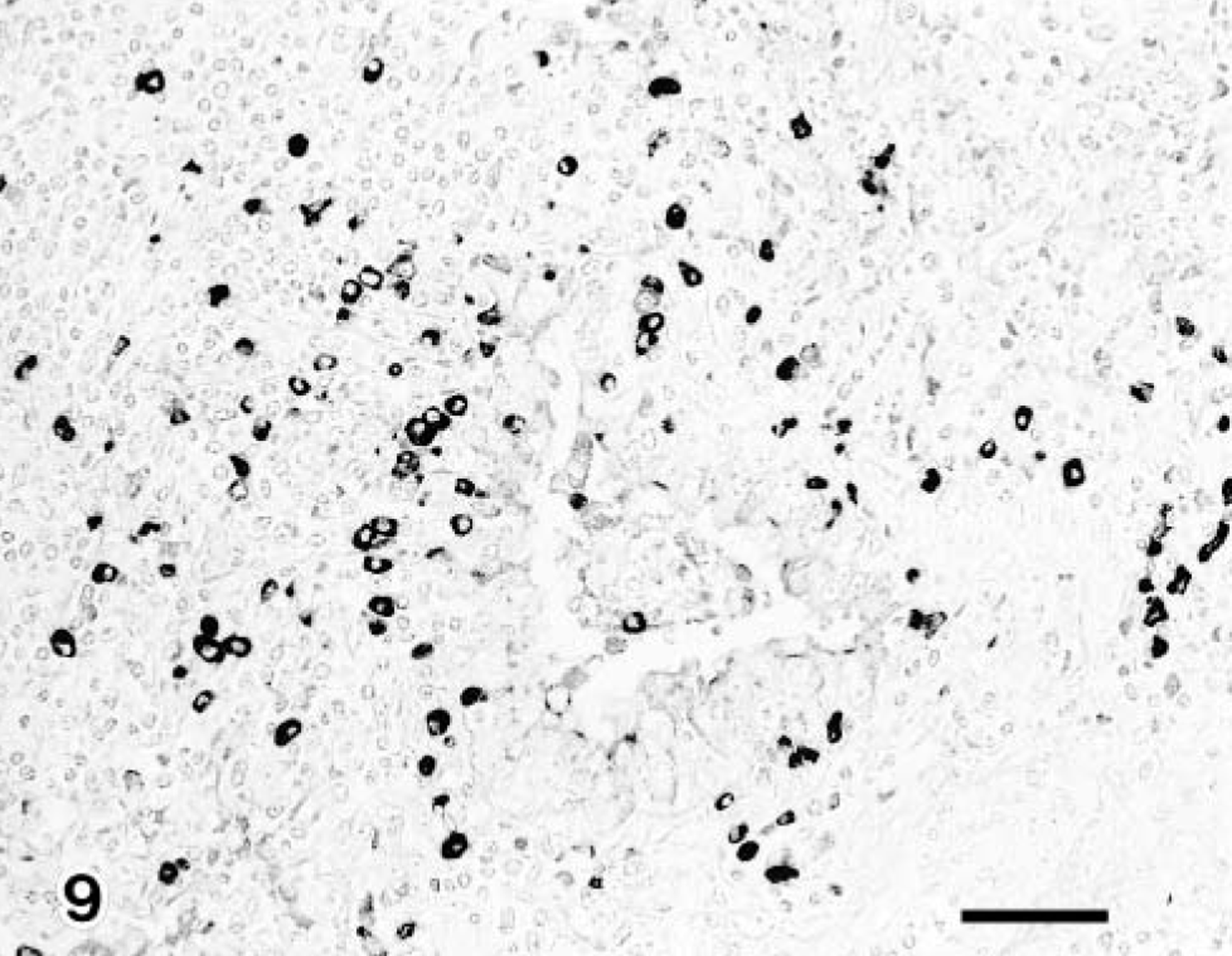
Lung; calf No. 6. Many IgA-containing cells are detected in the bronchus associated pneumonic lesion. SAB-IP. Bar = 50 μm.
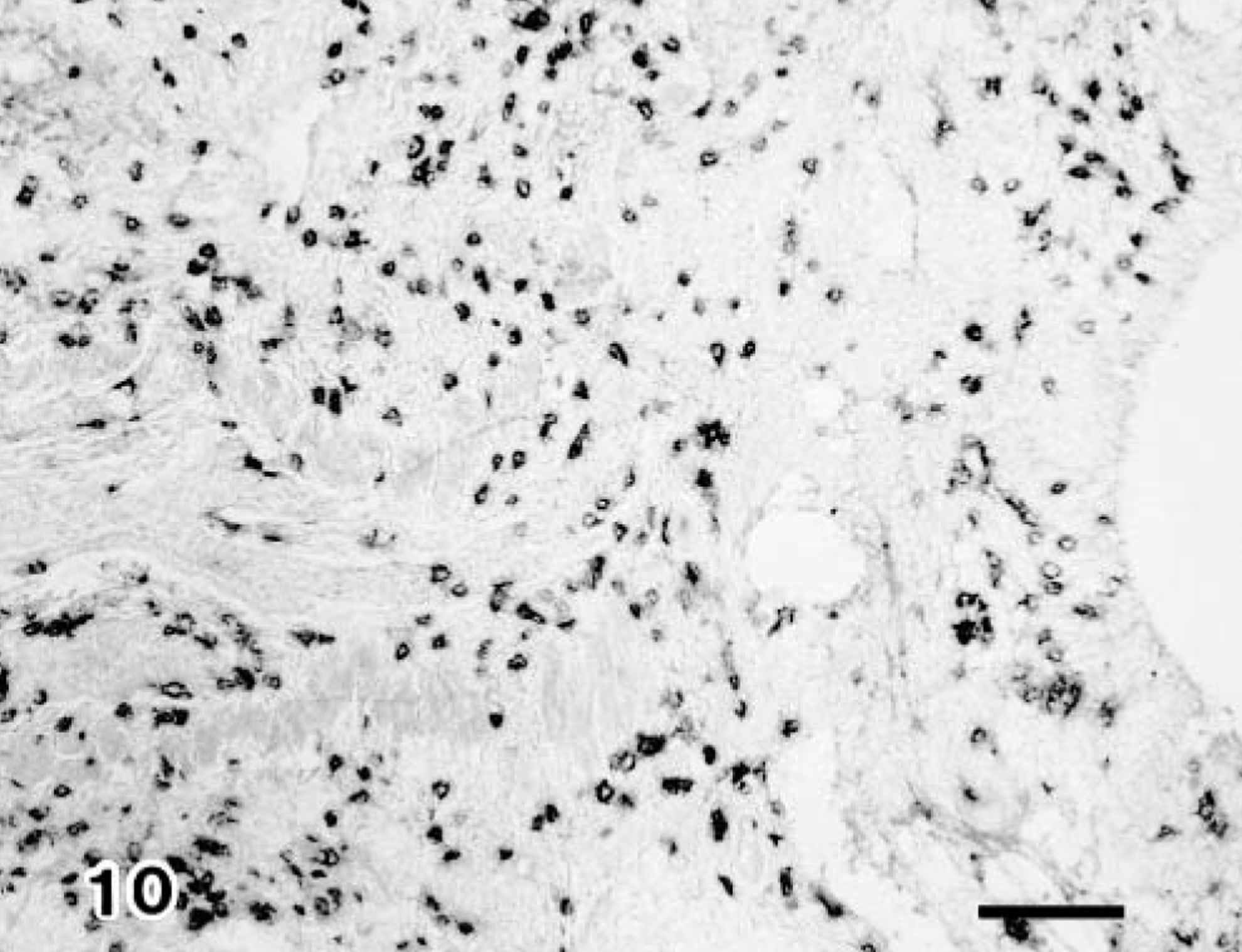
Lung; calf No. 6. Many CD8+ T lymphocytes are detected in the pneumonic lesion. SAB-IP. Bar = 50 μm.
Discussion
Adenovirus is known to be an important pathogen in immunocompromised hosts, including newborn infants as well as recipients of bone marrow, kidney, and liver transplants, infecting not only the transplanted organs but also other organs. 2 , 13 , 21 The effects of immunosuppression in domestic animals also have been demonstrated under experimental treatment with corticosteroids. 14 , 16 , 20 In the present study, no prominent clinical respiratory signs were observed in any BAV-3 infected calves. A high level of BAV-3 was isolated from BAL fluid cells of DM-treated 1.5-month-old infected calves as well as from those of 7-day-old infected calves but no BAV-3 was isolated from 1.5-month-old infected calves. These results correspond well to the detection of intranuclear inclusion bodies in the BAL fluid cells. Thus, it is clearly demonstrated that treatment with DM enhances the replication of BAV-3 in respiratory tissues.
Respiratory lesions induced by adenovirus infection in calves, lambs, dogs, and other small animals are characterized by focal necrosis of the epithelium in the lower respiratory tract associated with the presence of intranuclear inclusion bodies. 3–7 , 9 , 19 , 25 In the present study, pneumonic lesions were divided into two types. The first type was characteristic of severe necrotizing bronchiolitis with numerous intranuclear inclusion bodies and was found in non–DM-treated 7-day-old infected calves and in DM-treated 1.5-month-old infected calves. The presence of inclusion bodies correlated with that of BAV-3 antigen and viral particles. These results are similar to those found in the case of colostrum-deprived calves infected on the day of birth with BAV-3 and in EB-infected 7-day-old calves. 5–7 , 19 The second type of pneumonic lesions were those associated with intrastitial pneumonia with severe cellular infiltration and were similar to those found in 3-month-old calves EB infected with BAV-3 or colostrum-deprived calves infected with trypsin-treated BAV-3, or both. 5 , 9 , 11 , 19
In recent studies, examination of BAL fluid cells has been used as a technique for understanding the pathophysiology of respiratory diseases. 17–19 In the present study, the total cell count of BAL fluid cells from the right caudal lobe increased after BAV-3 infection, and the cells consisted of neutrophilic and desquamated epithelial cells containing intranuclear inclusion bodies. Detection of BAV-3 antigen was closely associated with the presence of intranuclear inclusion bodies in the degenerated epithelial cells. Moreover, the presence of a positive number of inclusion bodies or BAV-3 antigen, or both, in the desquamated epithelial cells correlated well with the isolated level of BAV-3 in BAL fluid cells. Therefore, we conclude that examination of BAL fluid may be a valuable method of diagnosis for respiratory viral diseases. 17–19
It is well known that immunosuppression with corticosteroids influences the immune system and enhances susceptibility to viral infection. 14 , 20 Distribution of the cells containing heavy and light polypeptide chains of immunoglobulin also has been described in lung tissues from domestic animals. 1 , 15 CD8+ T lymphocytes have been shown to be important for adenovirus type 7 clearance from respiratory tissues in children and BAV-3 in calves. 12 , 19 , 22 In the present study, the 1.5-month-old calves infected with BAV-3 had the highest number of immunoglobulin (IgG, IgM, and IgA)-containing cells and CD8+ T lymphocytes in their pneumonic lesions, but their lungs neither contained detectable BAV-3 antigen nor any intranuclear inclusion bodies. A low number of immunoglobulin-containing B lymphocytes and CD8+ T lymphocytes was found in the non–DM-treated 7-day-old infected calves, in the DM-treated 1.5-month-old infected calves, nor in either of the noninfected controls. These results are similar to those found in 3-month-old calves infected with BAV-3 and in cyclophosphamide-treated chickens infected with the avian nephritis virus. 16 , 19 These results strongly suggest that DM treatment might influence immunologic function in 1.5-month-old calves with BAV-3 infection. The reduction in lymphocyte numbers after infection may be due to decreased infiltration of lymphocytes in pneumonic lesion. Thus, depletion of CD8+ T lymphocytes in calves might influence the clearance of BAV-3 infection from respiratory tissues, as well as bovine respiratory syncytial virus. 23
Footnotes
Acknowledgements
We thank Mr. M. Kobayashi and Miss M. Shimada for preparing the pathologic sections and Dr. Y. Ando and Mr. T. Fujisawa for preparing the photographs.