
Abstract
Animal models have played a critical role in elucidating the complex pathogenesis of traumatic brain injury, the major cause of death and disability in young adults in Western countries. This review discusses how different types of animal models are useful for the study of neuropathologic processes in traumatic, blunt, nonmissile head injury.
There are two main reasons for conducting research on traumatic, blunt, nonmissile brain injury. First, traumatic brain injury (TBI) is the leading cause of death and severe disability in people under 45 years of age in Western industrialized countries, affecting the young and adults in the most productive years of their lives and predominantly caused by motor vehicle accidents. For every fatality, there are many survivors with severe brain damage and many more with moderate or mild injury.30,51,63 Although TBI is a problem of major medical and socioeconomic significance, its pathogenesis is incompletely understood, and it is often difficult to reconstruct the events leading to the primary and secondary lesions of varying severity and regional distribution that constitute TBI.11,50 In contrast, the mechanical input in animal models is quantifiable and subject to manipulation, and head injuries are produced under controlled experimental conditions. Second, although a host of potential neuroprotective agents have been studied, the few that have shown promise under experimental conditions have failed to provide consistent and significant improvement in human clinical trials.14,16,29,108 Testing the efficacy of new drugs in animal models will therefore continue to be an essential precursor to their use in humans.
TBI is also frequently encountered in veterinary practice as a result of automobile accidents, falls, assaults, bites, and crushing injuries.19,110 But head injury in animals has generally received scant attention in the veterinary literature, and most of our knowledge is derived from their use as experimental models of human TBI.34 This review highlights the contribution of different types of animal models to our understanding of the neuropathology of head injury.
Types of Animal Models
TBI may be produced by the head impacting or coming into contact with an object (contact phenomena) or acceleration/deceleration forces producing vigorous movement of the brain (acceleration/deceleration or inertial phenomena), or varying combinations of these mechanical forces.11,34,50,51,91
In most models, the mechanical input is controlled and results in injury that is reproducible, quantifiable, and clinically relevant. No single animal model can reliably replicate the full spectrum of human TBI.40,42,67,68,87,88,90
Animal models of TBI can be broadly classified as
impact acceleration models inertial (nonimpact) acceleration models direct brain deformation models.
Impact acceleration models involve direct head impacts using a piston, humane stunner or captive bolt pistol, calibrated pendulum, or weight drop onto the skull. This focal mechanical loading causes deformation of the brain which, being almost incompressible, is particularly vulnerable to strain injury. These models resemble closed head injury in motor vehicle accidents or falls where there is rapid acceleration/deceleration of the head after impact to an intact skull, but they sometimes fail to produce a highly repeatable injury. Impact acceleration models have been used in primates,54,62,81,83,84,101 sheep,32,33,35,36,64 cats,113 rats,8,71,102 and rabbits.55
Head impact models in primates were pioneered by Denny-Brown and Russell who showed that brain damage was more likely to be produced in a freely mobile head than in one that is constrained.20,21 Head impact models have reproduced several important features of TBI in primates, cats, and sheep, including contusions, subarachnoid hemorrhage, and widespread axonal injury (AI) and, in contrast to inertial acceleration models, skull fractures are commonly found. Diffuse brain injury was also produced in an impact-acceleration rat model,37,71 whereas previous impact models in rats had been largely unsuccessful because the high acceleration levels required to produce brain damage resulted in a slender margin between injury and death.40
Inertial acceleration models involve acceleration of the head without impact, unlike most human motor vehicle accident situations.78 Inertial acceleration devices generate a repeatable pathologic response, especially when the direction and distance of head movement is constrained. Angular acceleration, especially in the coronal plane, has been shown to be particularly injurious in nonhuman primates,43,44,47,81,82,83,85,86 pigs,98 and cats.80 In these models, the acceleration required to produce injury experimentally is found only in human TBI when head impact occurs.67
Inertial injury models were first developed in primates by Ommaya and Gennarelli, and subsequent modifications by Gennarelli and colleagues succeeded in reproducing many features of human TBI.38,41,43,44,46,47,83,86 By sudden deceleration of a moving frame to which the primate body was firmly attached, coma was produced by the whiplash motion of a freely mobile head. In these studies, coma was more readily produced by angular acceleration in the coronal plane and was associated with diffuse axonal injury (DAI) in the subcortical white matter.3,4,41,44,59 Angular or translational acceleration could be generated in these primates and, by controlling head motion, acute subdural hematoma, brief unconsciousness or prolonged coma, and DAI were induced.
Direct brain deformation models include both fluid percussion and rigid indentation types and use either a fluid pulse or mechanically driven piston, respectively, to rapidly compress the exposed dura or cortex through a craniotomy site.58,70,100 These models produce well-controlled levels of localized injury rather than diffuse damage. Brain deformation models have been used primarily in rats18,23–25,48,66,76,77,111 and also in cats15,56,57,89 and ferrets.65
In fluid percussion models, contusions of varying size can be produced; these being smaller and less frequent with a central (midline) impact compared with a lateral impact. In centrally directed models, scattered AI is principally found in the brainstem with modest involvement of cortical and subcortical areas.23,89,92,106,112 A lateral approach tends to produce hippocampal injury with less brainstem damage, and unilateral AI occurs in the central white matter. Contusions are also produced with central and lateral rigid indentation models, but AI is more localized than diffuse.77,99
An optic nerve stretch model has also been developed in guinea pigs, enabling AI to be studied under very controlled conditions.39,72,74
Thus, although no single animal model of nonmissile head injury produced by mechanical energy can express the great diversity of neural damage constituting human TBI, multiple animal models are nevertheless capable of replicating specific features of TBI that can be analyzed.
Types of TBI
Primary traumatic brain damage
This type of TBI is the result of mechanical forces producing tissue deformation at the moment of injury. These deformations may directly damage the blood vessels, axons, neurons, and glia in a focal, multifocal, or diffuse pattern of involvement and initiate dynamic and evolving processes that differ for each component part and result in complex cellular, inflammatory, neurochemical, and metabolic alterations.11,51 Primary TBI is summarized in Table 1.
Classification of traumatic brain injury.

Secondary traumatic brain damage
This type of TBI occurs as a complication of the different types of primary brain damage and includes ischemic and hypoxic damage and cerebral swelling, the consequences of raised intracranial pressure, hydrocephalus, and infection.11,51 These events are summarized in Table 1.
Experimental models of TBI have been devised using a variety of techniques and species, with the aim of producing repeatable lesions resembling those found in head-injured man. Although primary damage produced by mechanical forces operating at the moment of impact is largely refractory to treatment and relies on preventive measures, most brain injury evolves as a progressive cascade of events that is potentially reversible with adequate treatment. The recognition of the therapeutic window created by these delayed, secondary complications of the initial injury has led to attempts to pharmacologically manipulate some of the putative factors involved.52
Traumatic AI
Complete transection or avulsion of neural tissue produces instantaneous complete axotomy of all the nerve fibers (primary axotomy), as well as disruption of blood vessels and glial cells. The AI in less severe mechanical insults is termed secondary axotomy, and in most cases the process takes many hours or even days to be completed,73 creating a potential window for therapeutic intervention.51,92,94
The detection of AI in human TBI patients and animal models has been greatly enhanced by the introduction of immunocytochemical techniques using antibodies to axonally transported proteins, such as amyloid precursor protein (APP), neurofilament protein, and synaptophysin.51 APP immunostaining (Figs. 1–3) is the most sensitive marker of AI,105 identifying axonal swellings within 30 minutes postimpact and with minimal background interference because uninjured axons do not stain with this technique.33,36,64,114,115 But APP immunoreactive axons can still be identified many months after head injury.17
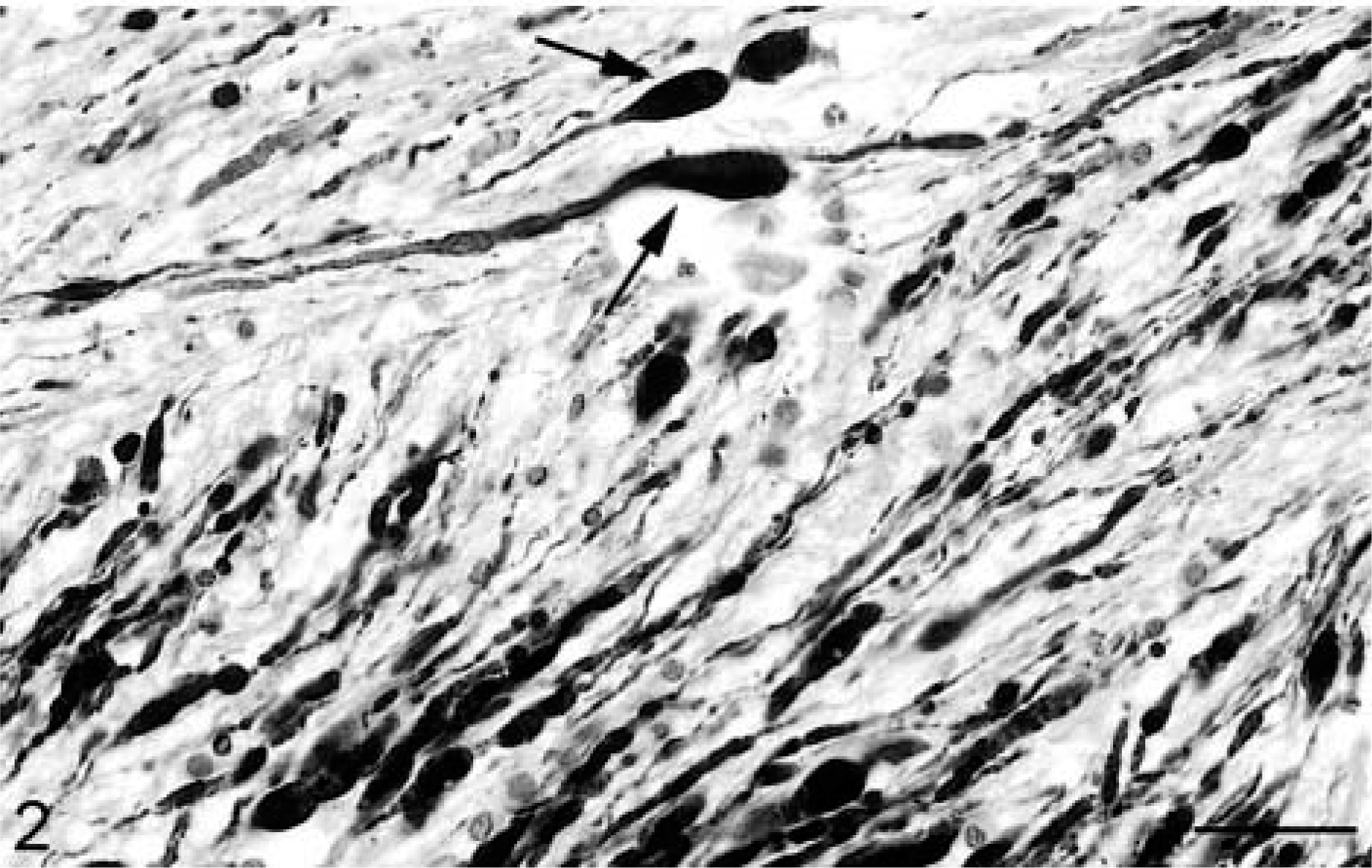
Sheep. APP-positive axonal swellings, with most axons running longitudinally in the same direction. Some axons are irregularly beaded (arrows). Avidin–biotin–peroxidase method, hematoxylin counterstain. Bar = 40, 80 μm.
Sheep. APP-positive axonal swellings, with most axons running longitudinally in the same direction. Some axons are irregularly beaded (arrows). Avidin–biotin–peroxidase method, hematoxylin counterstain. Bar = 40, 80 μm.
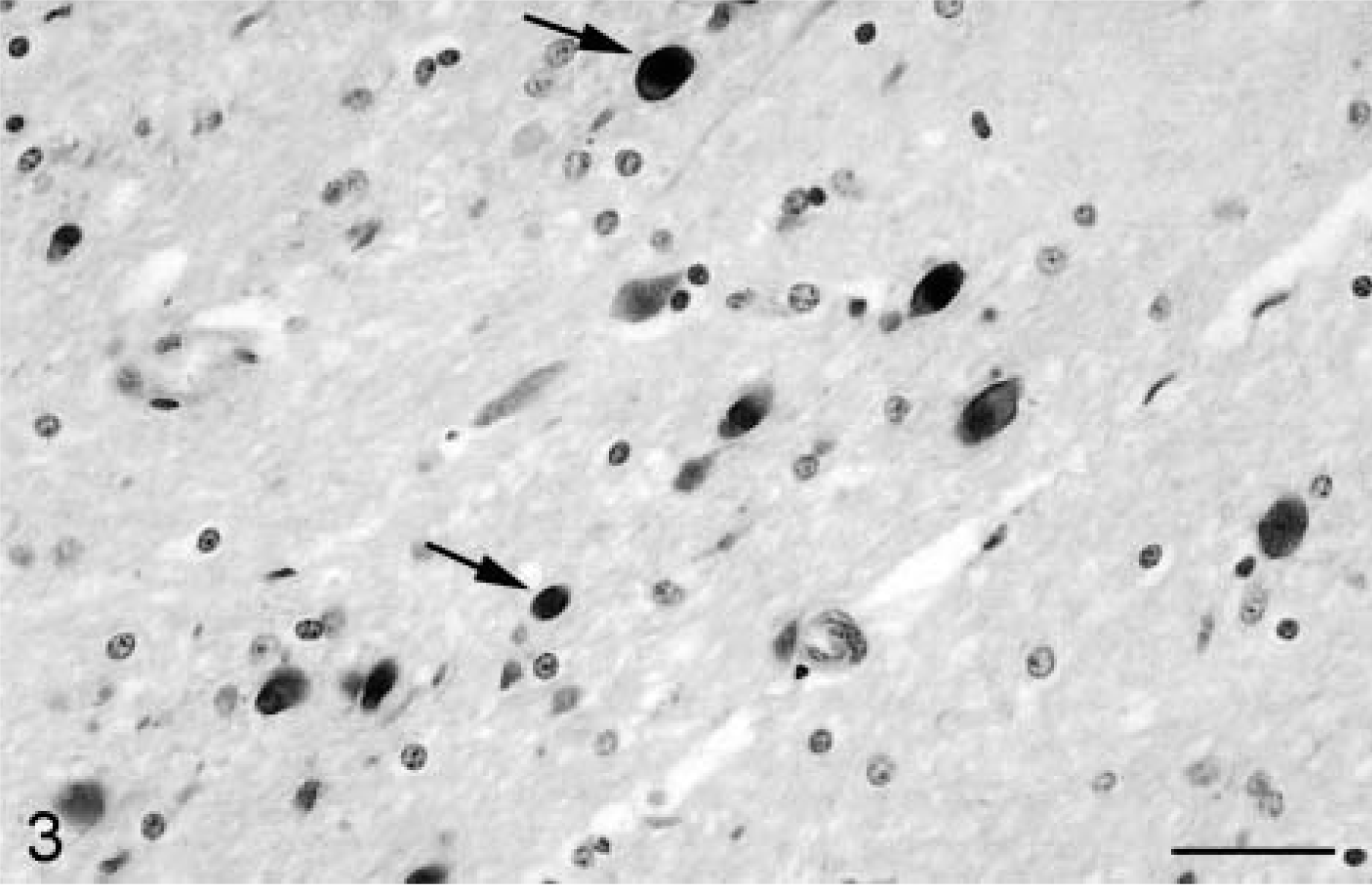
Sheep. APP-positive axonal swellings cut in transverse section (arrows); uninjured axons are not stained with this immunohistochemical technique. Bar = 72 μm.
The total burden of AI in a given brain has been underestimated using traditional, less sensitive staining methods such as hematoxylin and eosin and silver impregnation.105 But focal nontraumatic AI around hematomas, infarcts, and abscesses cannot be distinguished from AI due to mechanical deformation of the brain, and thus APP accumulation in axons is not confined to head-injured patients and is not a specific marker for head injury.11,12,51,105
Animal models have defined the early changes after traumatic AI. Mechanical deformation and cell membrane depolarization by shearing forces lead to a marked calcium influx into cells, which may be receptor-mediated, voltage-dependent, or via transient defects in the plasmalemma (mechanoporation). As a result of this traumatic calcium overload, enzyme and gene activation occur. Proteases damage the cytoskeletal architecture leading to interruption of axonal transport and accumulation of cytoskeletal components and membranous organelles over 3–6 hours, which is manifest as axonal swellings. Axotomy becomes apparent 6–12 hours after injury, and the distal segment undergoes Wallerian degeneration.94,95 This temporal progression of axonal reaction is also variable and species-dependent, occurring more rapidly in rodents than higher order animals.88 Axons that change direction during their course appear to be more vulnerable to injury, and it is not uncommon to find APP-positive axons in one tract and few or none in an adjacent tract with a different orientation. AI is followed by widespread and intense microglial reaction.
DAI is defined as widely distributed AI in the cerebral hemispheres, corpus callosum, brainstem, and cerebellum and is an important determinant of clinical outcome after human TBI.2 It spans a clinical spectrum from concussion to severe disability, permanent unconsciousness, or the vegetative state. In severe human DAI, there may also be hemorrhage associated with tears in the corpus callosum and dorsolateral quadrants of the rostral brainstem, which serve as useful markers for the postmortem and radiologic diagnosis of DAI; no macroscopic abnormalities may be found in milder forms.9–12,51 These focal hemorrhages are rarely reported in animal models, but brainstem and cerebellar hemorrhage observed in an ovine head impact model may have been due, in part at least, to impact against the well-developed tentorium cerebelli,33,35,36,64 which closely embraces the brainstem in animals.60
The total amount of AI in a given brain may be a combination of mechanical deformation and ischemic injury, especially because ischemia-hypoxia due to failure of cerebral perfusion is one of the major secondary insults after head injury. But the current histologic methods for detecting AI are unable to distinguish between these types of AI.12 Continuous physiologic monitoring is thus mandatory when modeling AI to ensure that no complicating hypoxic episodes supervene. Injured axons also seem to be much more vulnerable to ischemia than normal axons.31
In general, widely distributed AI has been produced with some fidelity by inertial and impact acceleration models, whereas AI after direct brain deformation tends to be unilateral and localized to the impact site and subjacent white matter. Modification of the lateral rigid percussion model by opening the contralateral cranium sometimes extends AI across the midline in subcortical areas.45
Brain acceleration of the type that produces human DAI seems to be largely confined to animal models that have a relatively large, gyrencephalic brain such as nonhuman primates (whose ratio of brain mass to head mass is similar to man), sheep, and pigs, in part because shearing forces and inertial loading are related to brain mass.34 DAI is rarely produced by inertial loading in small animals, such as cats, ferrets, and rats.42 Gyrencephalic brains have a complex, species-specific pattern of surface convolutions resembling man,13 in contrast to the almost lissencephalic rodent brains that can tolerate much greater acceleration/deceleration forces than nonhuman primates and man. In quadrupeds, the long axes of the brain and spinal cord are parallel, whereas in man and nonhuman primates they are almost at right angles. This almost linear neuraxis in lower species may impede rotational shearing after head injury and render them less vulnerable to traumatic injury.40,90,88
Traumatic loss of consciousness
This was reliably reproduced in acceleration and percussion concussion primate models by Denny-Brown and Russell at the beginning of the modern era of neurotrauma research at Oxford in the early 1940s.20,21 These studies also demonstrated the importance of head acceleration in TBI, for impact to a freely mobile head was much more likely to produce loss of consciousness than to one constrained. Head acceleration may be either translational (linear) when the impact force passes through the center of gravity of the head or angular (rotational) when it does not. Ommaya, Gennarelli, and colleagues in Philadelphia found that angular (rotational) acceleration of the head of primates, especially in the coronal plane, produced coma more readily.1,43,44,81,85
The Philadelphia primate studies highlighted the critical importance of acceleration/deceleration in TBI, for the head does not need to strike or be struck by an object to produce widespread AI. In these studies, the greater the number of reactive axons, the more persistent the neurologic abnormalities and the duration of unconsciousness after head impact correlated with the degree of DAI. Loss of consciousness persists for seconds to minutes with mild DAI but days to weeks with more severe DAI. Ommaya and Gennarelli found that acceleration in the primate model over longer time periods (20–25 ms) as may occur in motor vehicle accidents produced prolonged traumatic coma and DAI, whereas a short acceleration time (5–10 ms) resembling that in falls caused subdural hematoma.51
Contusions
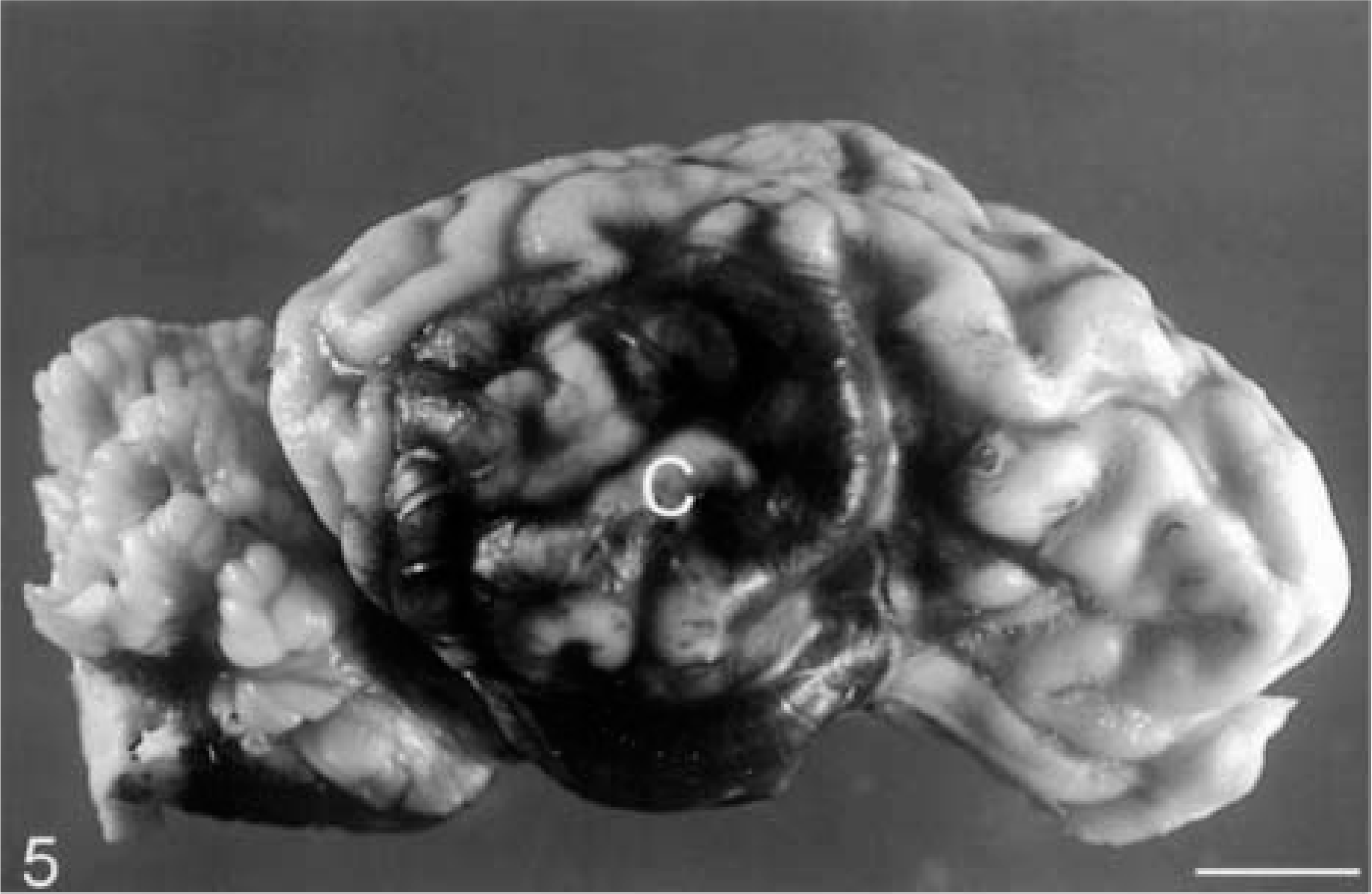
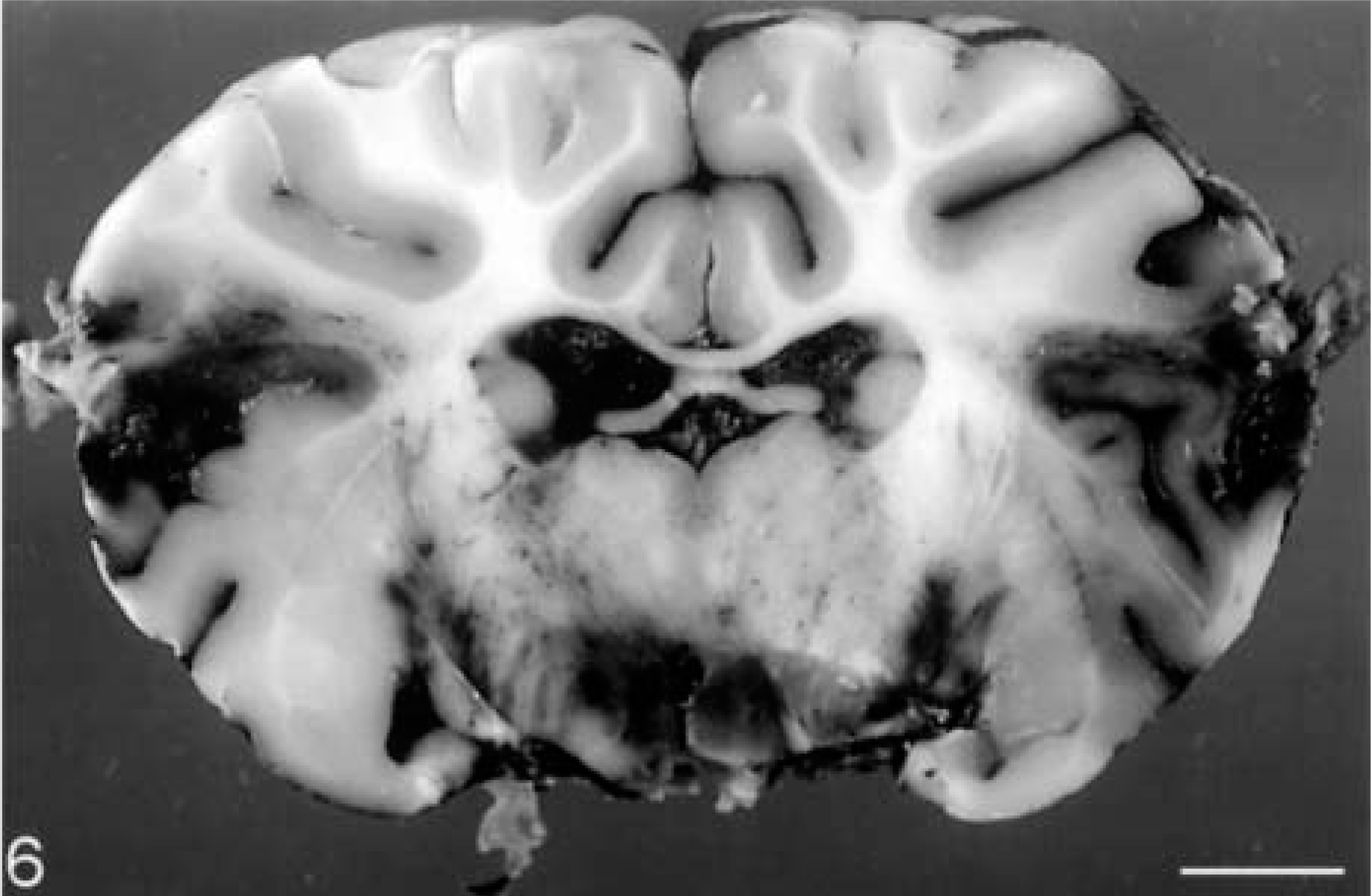
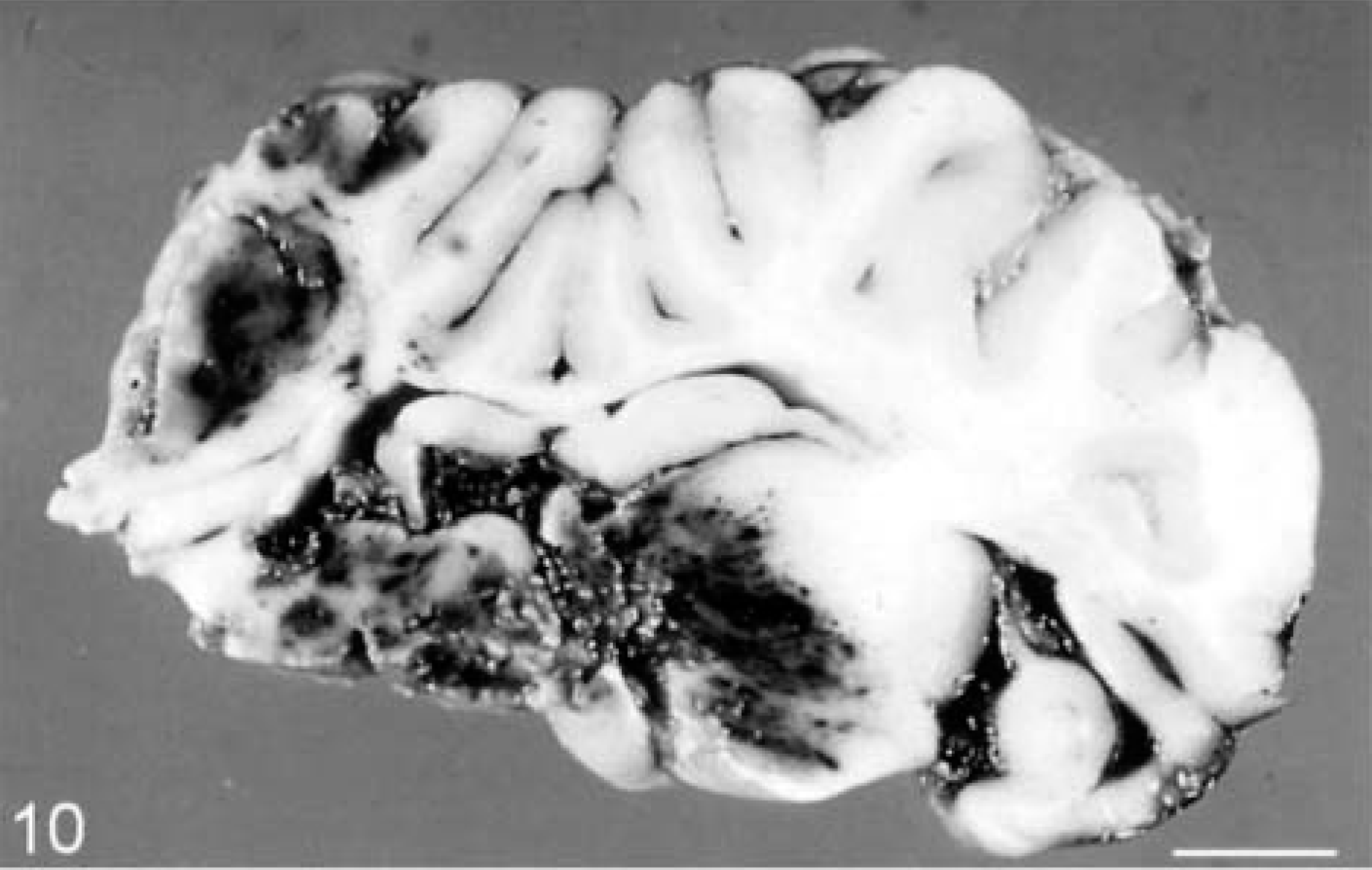
Contusions (Figs. 4–6) are regular features of TBI, and their presence confirms that a head injury has occurred. They have been observed to varying degrees in most animal models of TBI, although some brain deformation models produce hemorrhage at the gray-white matter interface rather than in the cortical mantle.18,23,77 Contusions are focal surface injuries resulting from damage to small blood vessels (Figs. 7, 8), with hemorrhages often disposed at right angles to the cortical surface. Contusions typically affect gyral crests and frequently progress to a wedge-shaped necrotic area involving the subjacent white matter, the initial focal lesion expanding due to continuing hemorrhage, ischemic necrosis, and vasogenic edema. They often occur beneath the impact site (coup contusions) and because contusions are mainly impact-related phenomena, they are often associated with skull fractures (fracture contusions) in impact acceleration models of closed head injury. Contrecoup contusions may also be found more or less opposite the impact site (Fig. 6), whereas herniation contusions occur in the margins of brain hernias. But contusions may be minimal or absent with fatal diffuse brain injuries. Laceration results when there is physical disruption of the neural parenchyma, and laceration-contusion are part of a continuous spectrum of damage at the surface of the brain.11,51
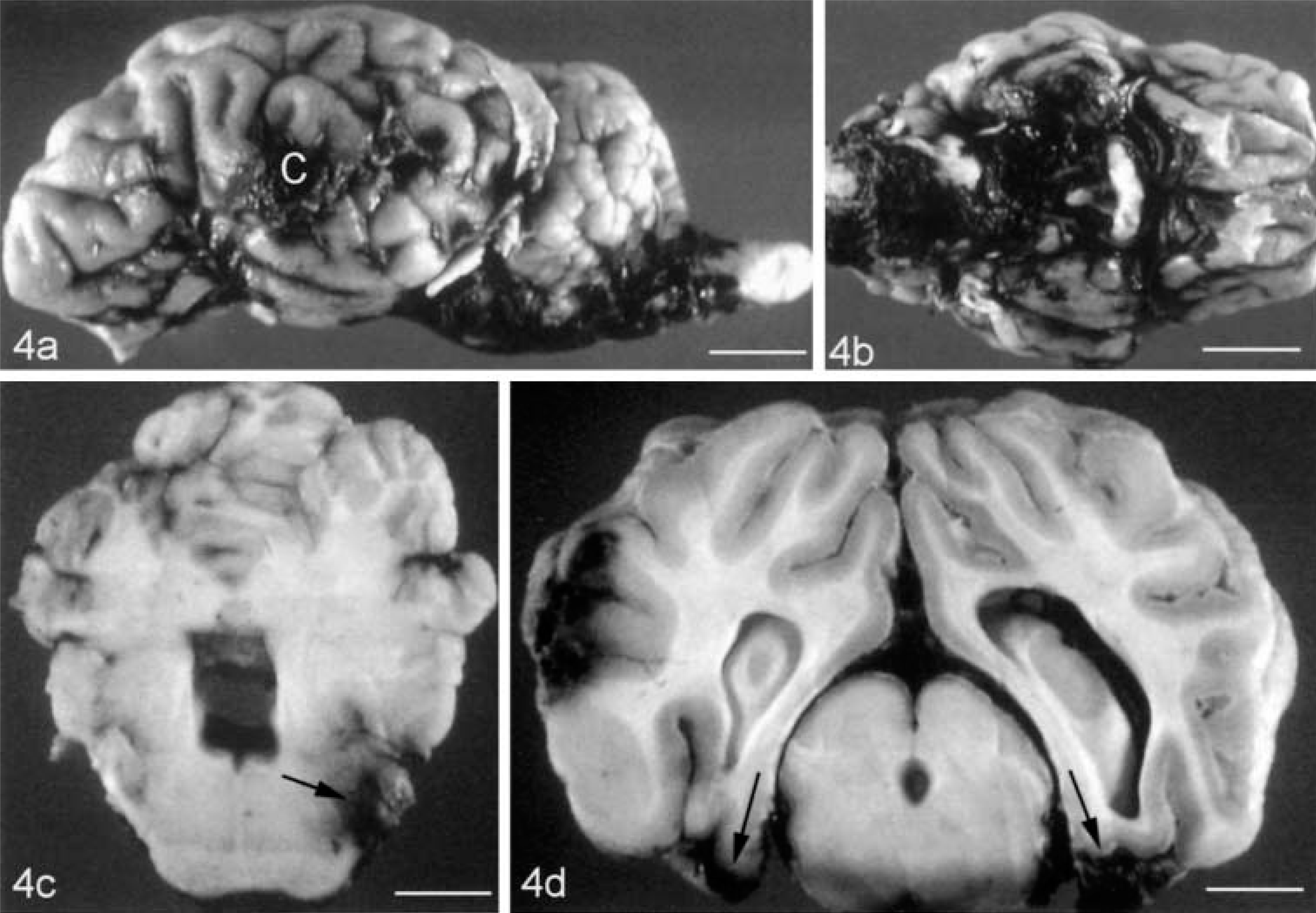
Sheep (clockwise from top left); lateral cerebral hemisphere impact contusion (C), with subarachnoid hemorrhage around the brainstem (bar = 11 cm); ventral aspect of the brain showing abundant subarachnoid hemorrhage (bar = 12.5 cm); focal hemorrhage in the lateral pons (arrow) (bar = 7.5 cm); left lateral hemisphere impact contusion, with contusions on the inferior aspect of the pyriform lobes (arrows) (bar = 4 cm).
Sheep. Large lateral hemisphere impact contusion (C). Bar = 4 cm.
Sheep. Left lateral hemisphere impact contusion, with contrecoup contusion directly opposite. Bar = 7 cm.
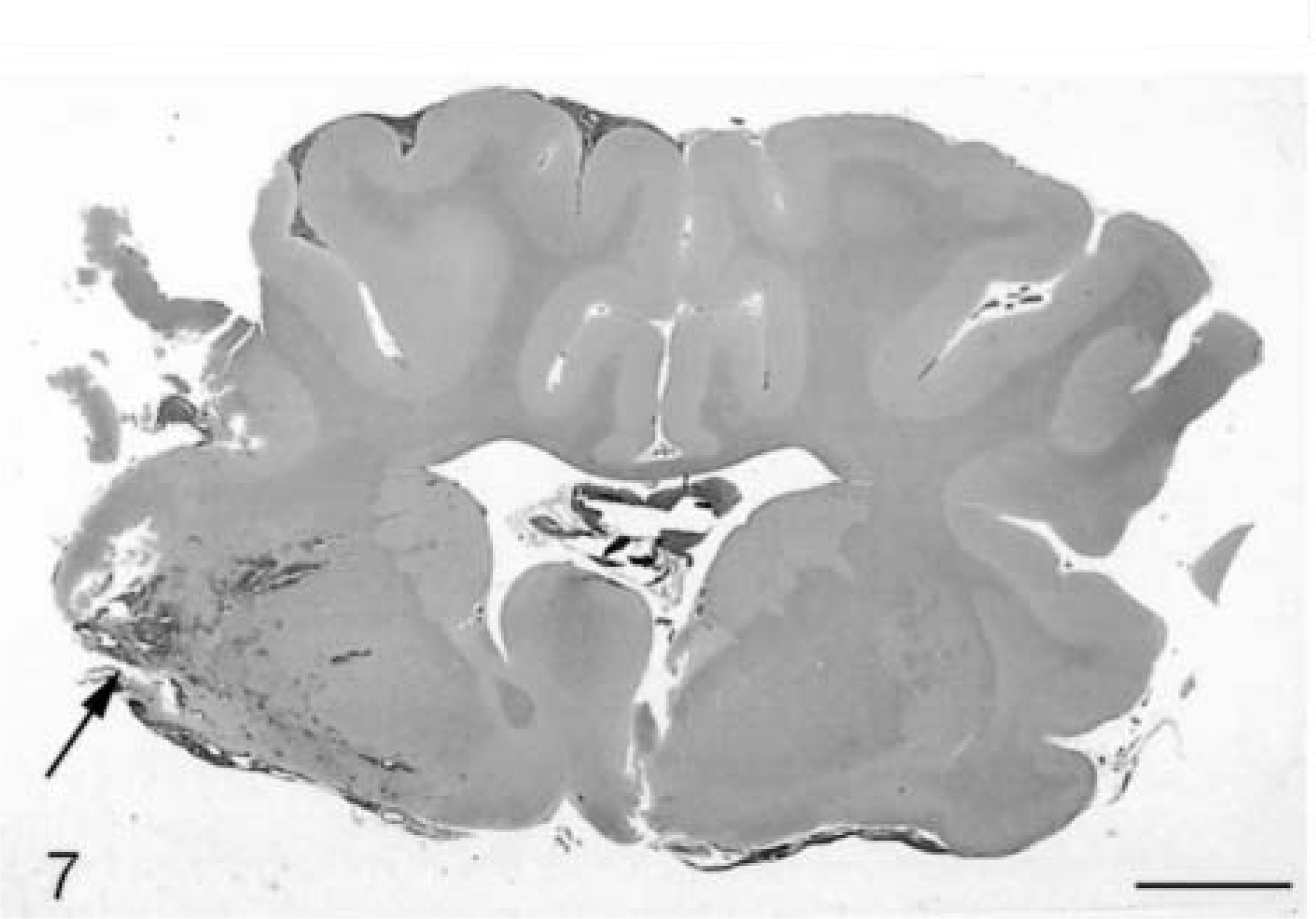
Sheep. Coronal section of brain showing left lateral hemisphere impact contusion and laceration (arrow), extending into the basal ganglia. HE. Bar = 5 cm.

Sheep. Higher power view of the contusion in Fig. 4, showing numerous perivascular hemorrhages. HE. Bar = 2 cm.
Traumatic brain hemorrhage
Traumatic brain hemorrhage (Figs. 9–11) generally results from tearing of blood vessels at the moment of head impact. But gradually expanding, delayed posttraumatic hematomas may not be manifest clinically until hours or days after the initial injury when they cause elevated intracranial pressure and herniation. Bleeding into the subarachnoid space (subarachnoid hemorrhage) is the most common form of vascular injury (Fig. 4) after head trauma and is found in many types of animal models. It is usually minor but may evolve into a significant space-occupying lesion. Subdural hematoma is produced in inertial acceleration models,44 where bridging veins are ruptured by rapid angular acceleration forces and the hemorrhage may extend over an entire hemisphere. Subdural hematoma may also form adjacent to contusions in, for example, a rigid indentation model.65 Because the skull is not fractured in most animal models, epidural hematomas with progressive separation of the dura from the skull after tearing of meningeal blood vessels are not observed. But skull fractures do occur with many impact acceleration models, unless the skull is protected by a metal plate or molded cap. Intraventricular hemorrhage is a frequent complication after human and animal head impact, and hematomas may also develop in the brain substance (intracerebral hematomas). Intraparenchymal hemorrhage is often distributed throughout the central white matter and basal ganglia in inertial models and principally in the brainstem in percussion models. In human patients who die within minutes after head impact, there may be numerous petechial hemorrhages scattered throughout the brain (diffuse vascular injury), but there appears to be no animal model counterpart.11,51
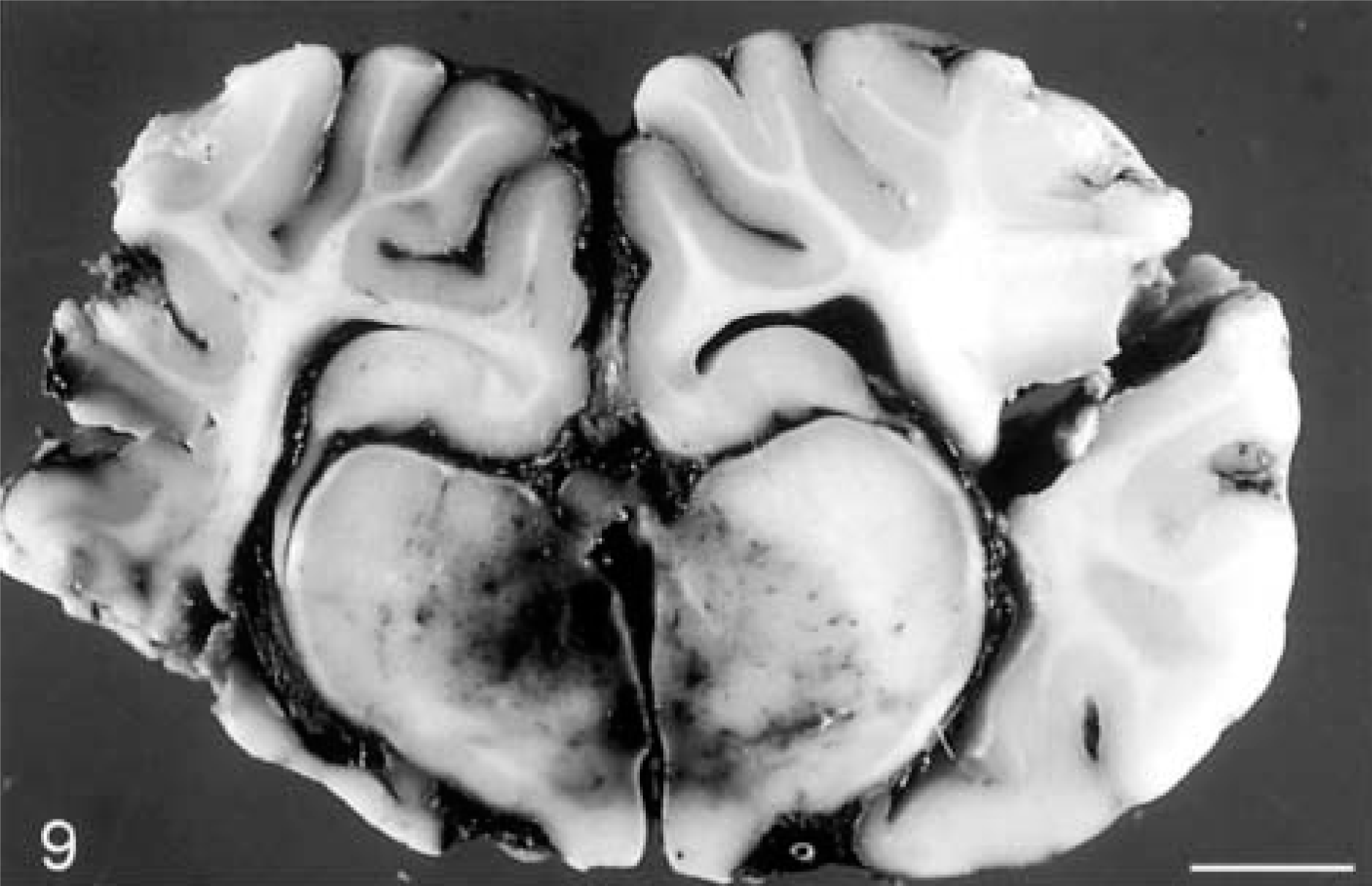
Sheep. Extensive traumatic midbrain hemorrhage. Bar = 7 cm.
Sheep. Severe traumatic thalamic hemorrhage, with midline shift and distortion of the brain. Bar = 8.5 cm.
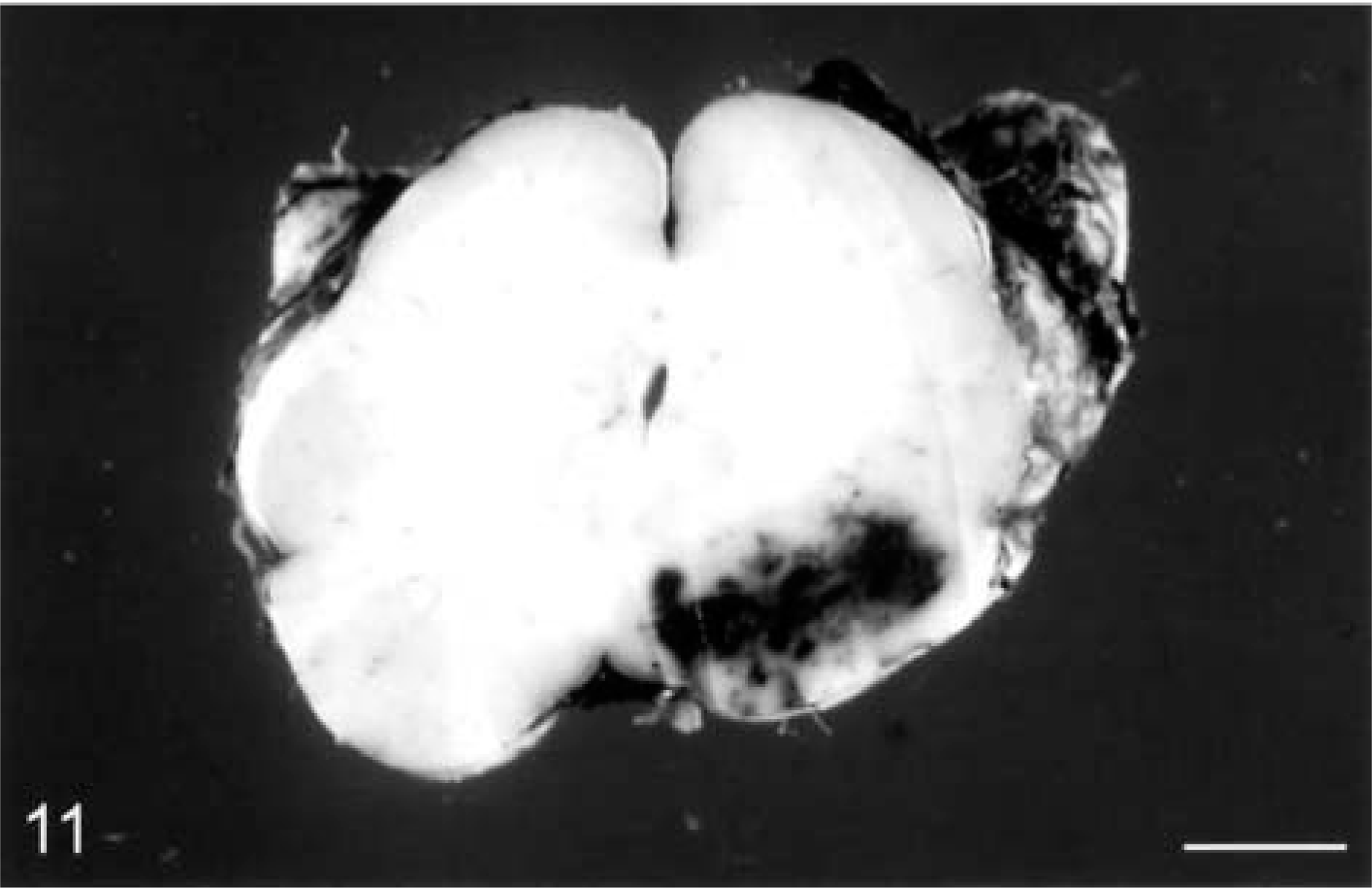
Sheep. Focal traumatic hemorrhage in the lateral pons. Bar = 4 cm.
There is widespread breakdown of the blood-brain barrier to circulating proteins very early after TBI in many animal models,18,76,93,100,109 often attended by brain swelling. Brain swelling encompasses both edema and congestion and, together with hematomas, is the major contributor to increased intracranial pressure. It may eventually lead to distortion, shift, and herniation of the brain. Edematous swelling often occurs around contusions and intracerebral hemorrhages in humans and animal models, whereas swelling of one hemisphere in man may result from a combination of congestion and edema. Diffuse swelling of the entire brain due to hyperemia occurs in young children, even after an apparently trivial injury, but is uncommon in adults. Generalized brain swelling produces gyral flattening, sulcal narrowing, and ventricular collapse.11,51
Reactive microglia also influence clinical outcome after TBI because they are the principal immune effector elements of the brain, are the major source of central nervous system–derived proinflammatory cytokines, assist wound healing in neural tissues, and produce many neurotoxic agents.53 Excitotoxicity may be a further important mediator of TBI. Traumatic brain deformation in animal models results in massive impact depolarization and excessive release of excitatory amino acid neurotransmitters, especially glutamate, into the extracellular fluid.28,61 Although the storage, transmission, and uptake of glutamate is normally finely regulated, a marked rise in extracellular glutamate after TBI is highly toxic to neurons.96
Chronic brain injury
The long-term effects of TBI depend on a combination of the different types of primary and secondary damage sustained and their relative severity, but modeling of chronic brain injury has rarely been attempted.40,88 But in a fluid percussion model eliciting AI, reorganization of the axonal cytoskeleton occurred over a 28-day period with a spontaneous regenerative attempt.15
TBI is a major risk factor for Alzheimer's disease (AD),75 and diffuse β-amyloid deposits have been reported to occur between 4 hours and 2.5 years in human TBI.97 Transgenic mice engineered to overexpress human APP twofold (APP-YAC mice) failed to produce amyloid plaques or cognitive change after injury,79 whereas transgenic mice overexpressing human APP 10-fold (PD-APP mice) showed hippocampal neuronal loss and decreased cognitive function.107 The use of genetically engineered mice to study the link between TBI and neurodegenerative diseases, and the pathogenesis of TBI more generally, has recently been reviewed.69
Pediatric TBI modeling
Although there are important age-related differences in the response of the brain to TBI, there have been few animal models using an immature brain, and differences in the stage of maturity of the brain at birth between species are important.7 Species can be categorized as prenatal (sheep and guinea pigs) or postnatal (rat, rabbit, pig, and man) brain developers in relation to the growth spurt, the period when brain growth is most rapid.26 The pig may be suitable as a model of human pediatric TBI because its brain development closely parallels that of humans, and both species have a peak in brain growth at the time of birth.22 In man, the long postnatal growth period of the brain renders it particularly vulnerable to traumatic injury and recovery is more complicated because it is superimposed on continuing developmental events. Moreover, although the sequence of brain development is well characterized, the effect of trauma on these immensely intricate developmental processes and the resumption of the orderly progression after injury is unknown.49,103,116
Models of pediatric TBI have produced contusional injury in the juvenile rat and pig by controlled cortical impact,6,27 and a model of subdural hematoma has also been reported in piglets.104 Impact-acceleration brain damage has been produced in young rats and lambs.5,33,36 In the rat model, AI was largely confined to the brainstem,5 whereas in the lamb model there was early and widespread AI,36 the severity of which was substantially influenced by the region of the head impacted.35
Conclusion
The complexity and diversity of TBI pathology will ensure a continuing role for animal models to define more accurately the cascade of morphologic and biochemical events occurring after a traumatic insult. One of the most important insights into the pathogenesis of TBI provided by animal models has been the realization that many structural changes, particularly AI, are not immediate and irreversible but often time-dependent evolving processes that may be amenable to therapeutic intervention. Animal models will be essential to evaluate potential neuroprotective agents.